Identification and Characterization of NPR1 and PR1 Homologs in Cymbidium orchids in Response to Multiple Hormones, Salinity and Viral Stresses

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
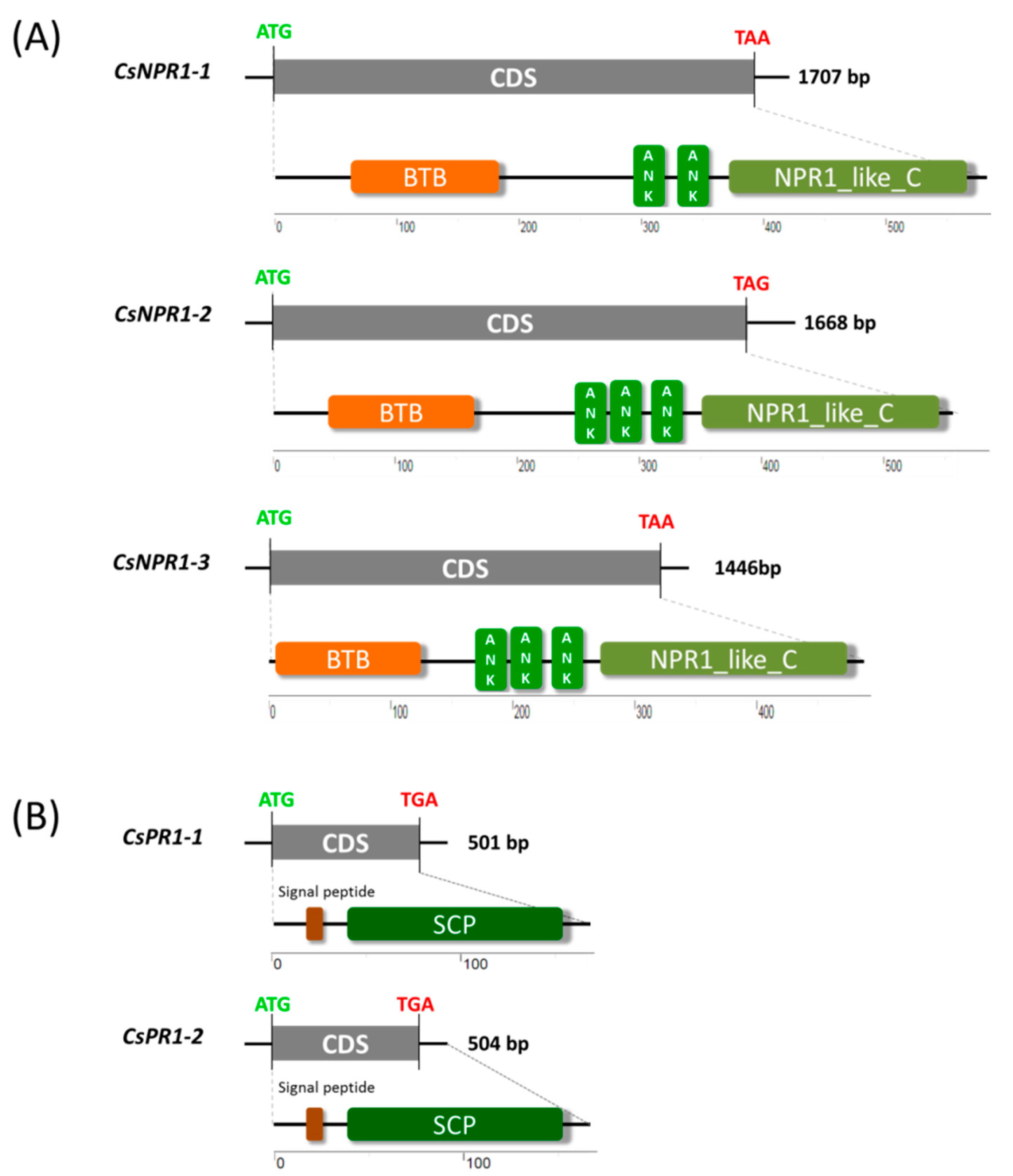
2.1. Genome-Wide Identification of NPR1 and PR1 Homologs in Cymbidium Orchids
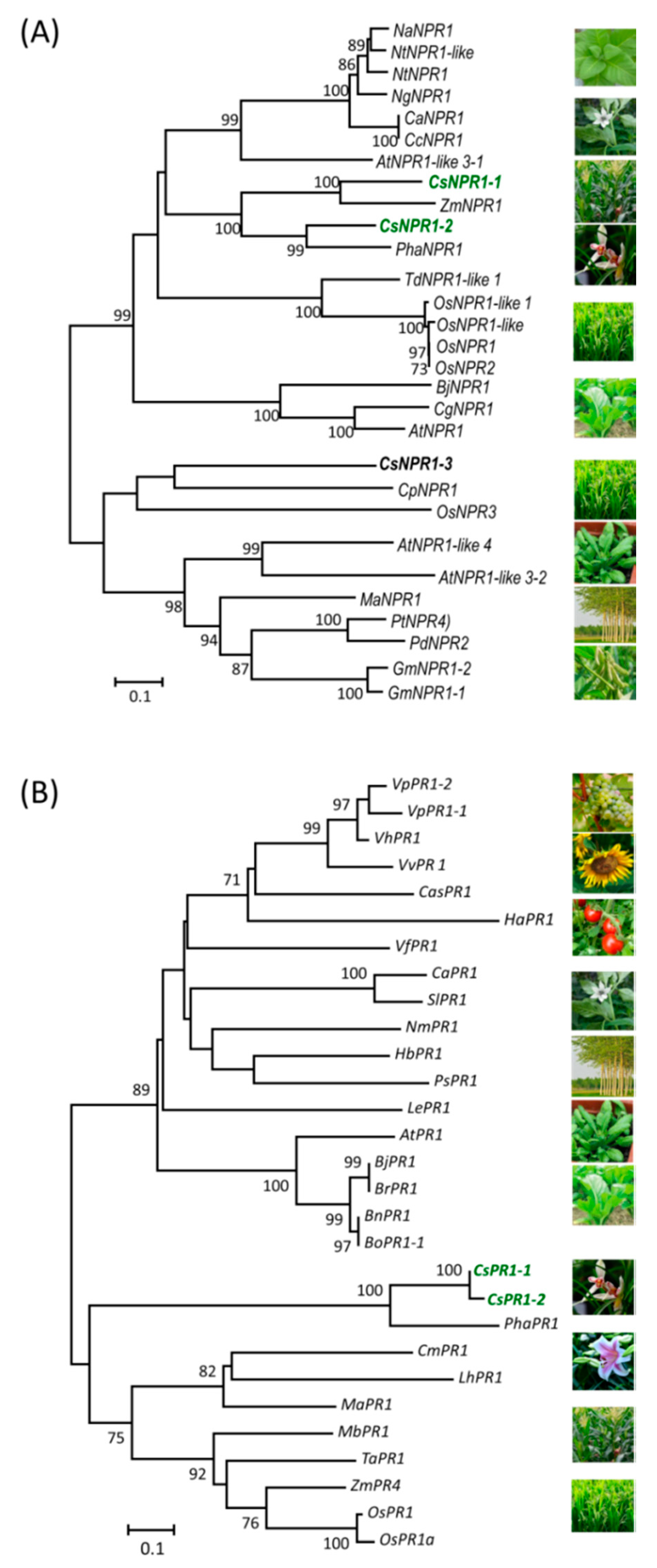
2.2. Phylogenetic Analysis and Sequence Alignment of NPR1 and PR1 Homologs
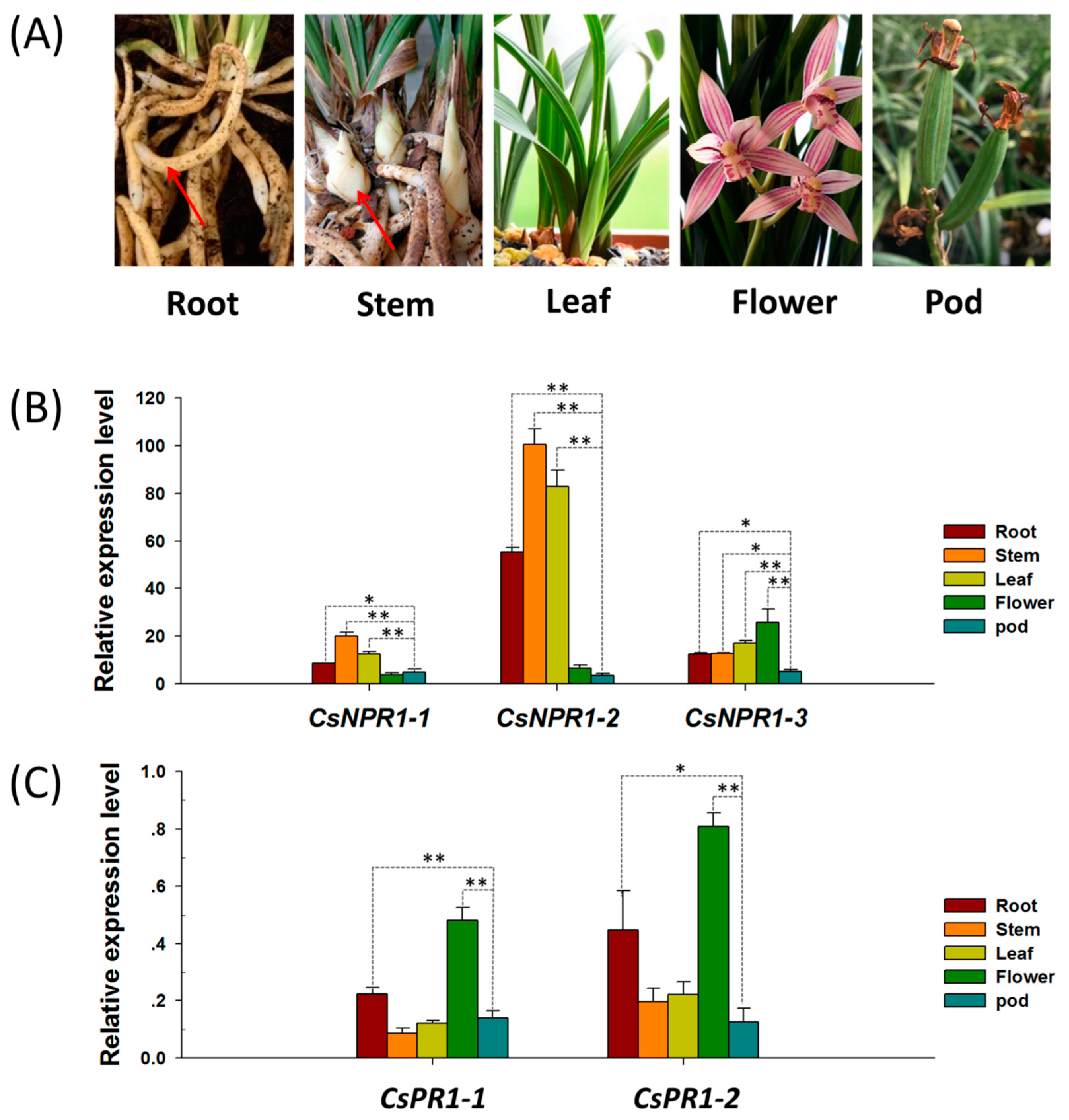
2.3. Tissues/Organs-Specific Expression Analysis of CsNPR1 and CsPR1 Genes
2.4. SA and ROS Significantly Upregulates Expression of CsNPR1 and CsPR1 Genes
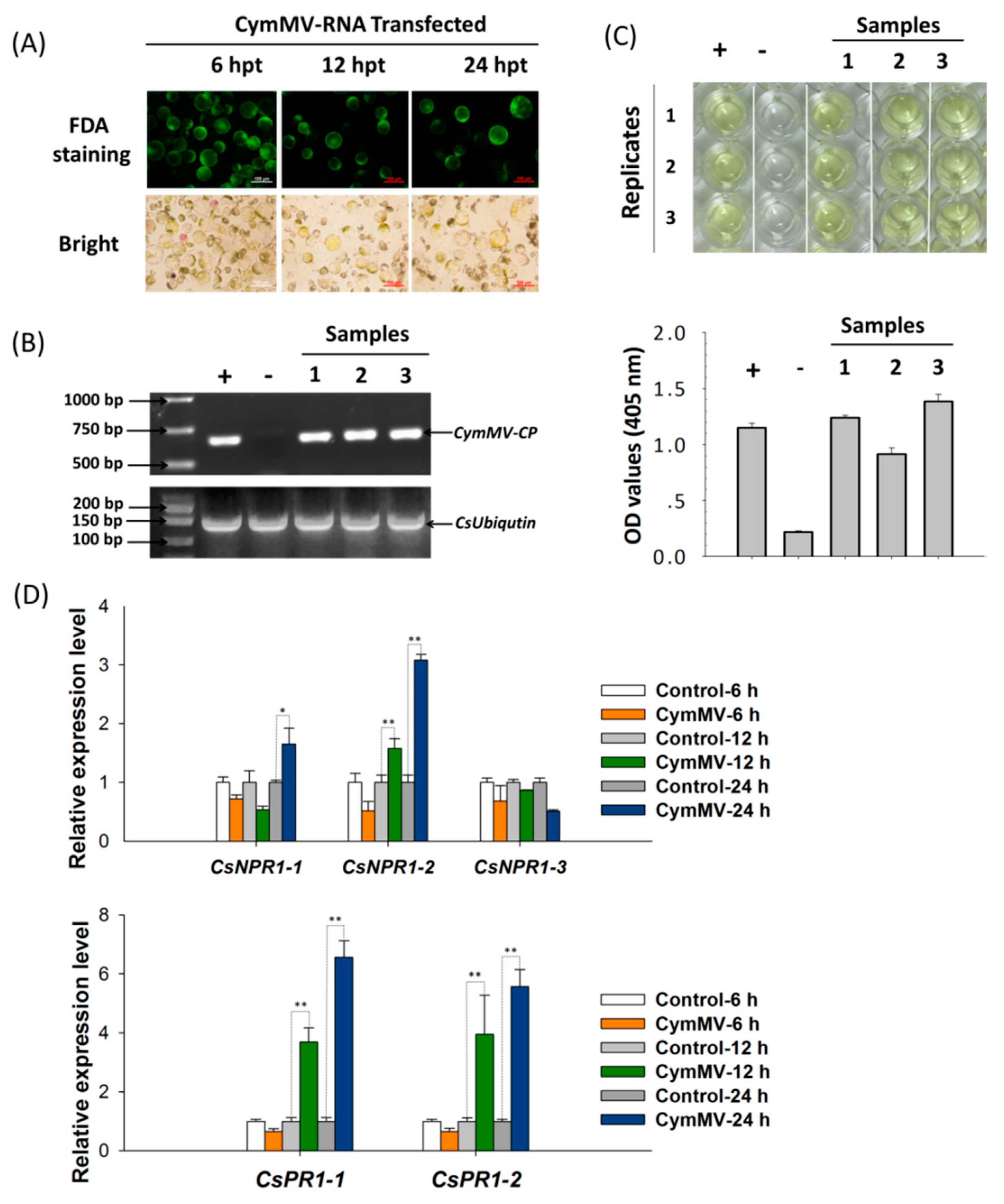
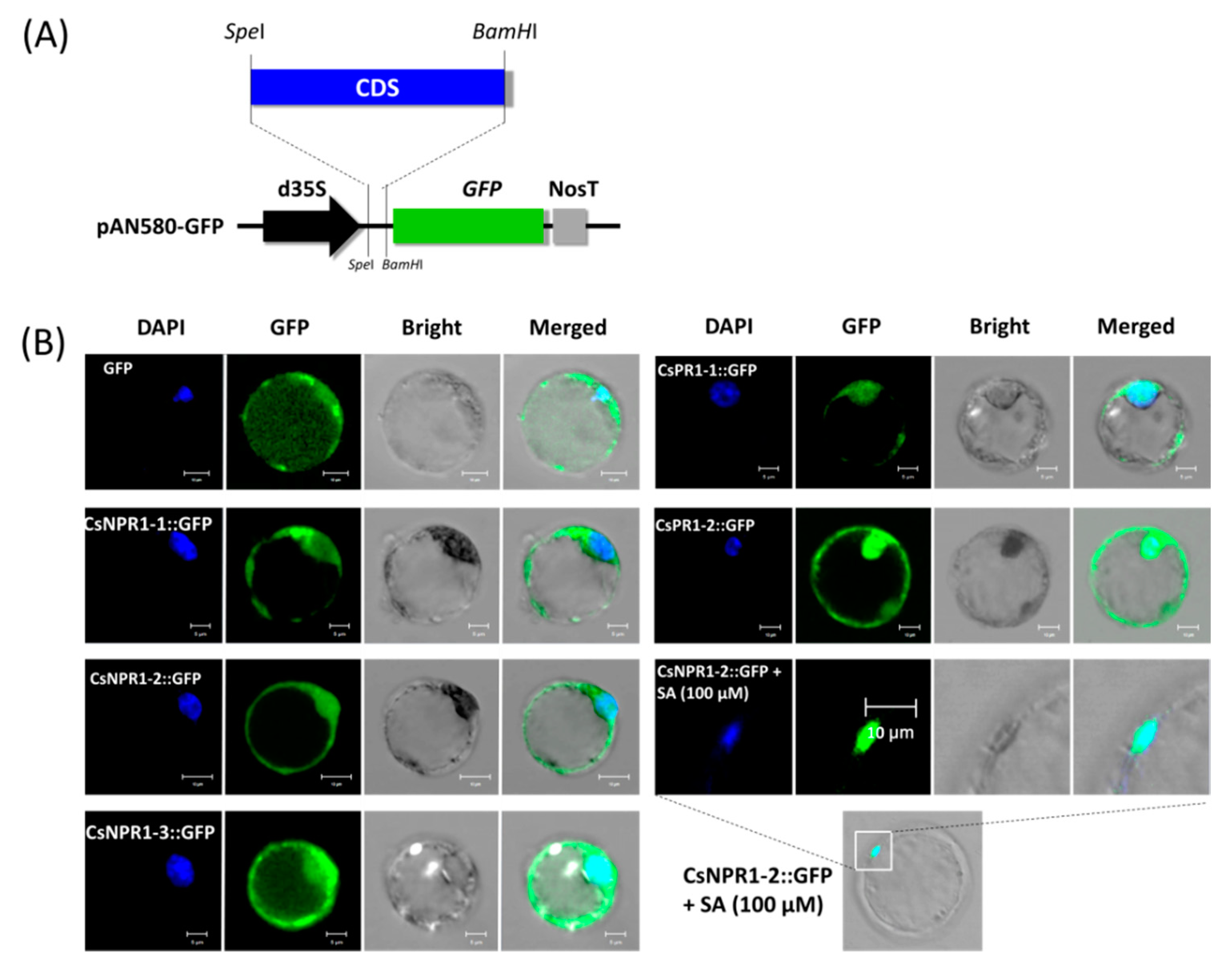
2.5. CsNPR1-2-Mediated Singling Pathway Confers the Response of Cymbidium orchids Against CymMV Infection
2.6. Nuclear Translocation of CsNPR1-2 in the Presence of SA
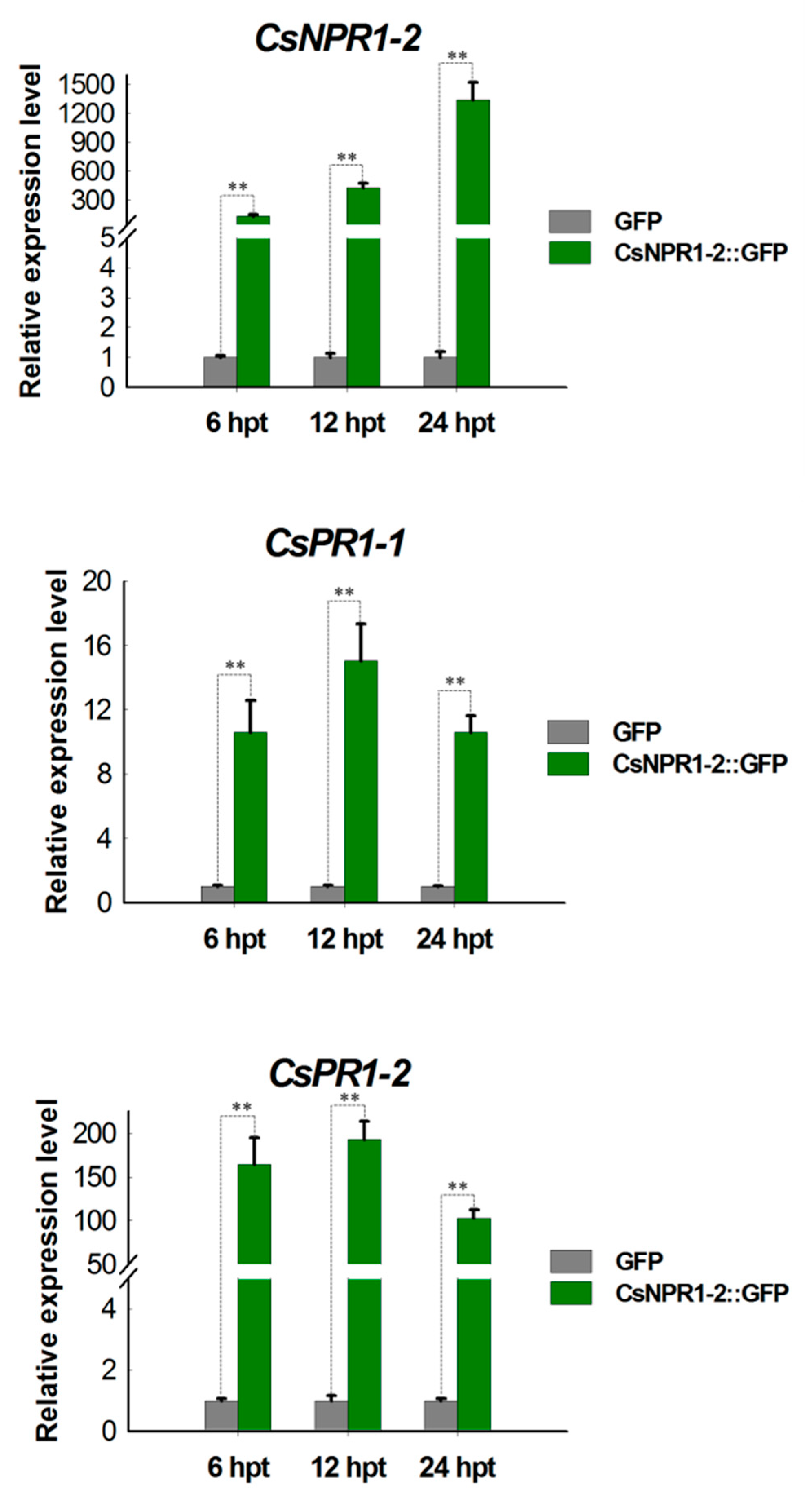
2.7. CsNPR1-2 Positively Regulate the Expression of CsPR1 Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Genome-Wide Identification of NPR1-like and PR1-like Genes in Cymbidium Sinense
4.3. Sequence and Phylogenetic Analysis
4.4. Tissues Collection and Protoplast Isolation
4.5. Protoplast Measurements and Treatments
4.6. In Vitro Transcription of CymMV-RNA and Vector Preparation
4.7. PEG-Mediated Protoplast Transfection
4.8. RT-PCR and qRT-PCR
4.9. DAS-ELISA
4.10. Statistical analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| ANK | Ankyrin repeat |
| AsA | Ascorbic acid |
| BTB/POZ | Bric-a-brac/poxvirus and zinc finger |
| CDS | Coding sequence |
| CymMV | Cymbidium mosaic virus |
| DAS-ELISA | Double antibody sandwich-enzyme linked immunosorbent assay |
| FDA | Fluorescein diacetate |
| GFP | Green fluorescent protein |
| PR1 | Pathogenesis-associated 1 |
| qRT-PCR | Quantitative reverse transcription -polymerase chain reaction |
| ROS | Reactive oxygen species |
| SA | Salicylic acid |
| SAR | Systemic acquired resistance |
| SCP | Sterol carrier protein |
| SD | Standard deviation |
| SP | Signal peptide |
References
- Roberts, D.L.; Dixon, K.W. Orchids. Curr. Biol. 2008, 18, 325–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.M. Orchid viruses—A compendium. In Orchid Biology VIII: Reviews and Perspectives; Kull, T., Arditti, J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 505–546. [Google Scholar]
- Chen, S.; Liu, Z.; Zhu, G.; Lang, K.; Ji, Z.; Luo, Y.; Jin, X.; Cribb, P.J.; Wood, J.J.; Gale, S.W.; et al. Orchidaceae. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2009; pp. 211–235. [Google Scholar]
- Cribb, P.J. Subtribe Polystachyinae, Taxonomic notes. In Genera Orchidacearum, Volume 6: Epidendroideae (Part 3); Pridgeon, A.M., Cribb, P.J., Chase, M.W., Rasmussen, F.N., Eds.; Oxford University Press: Oxford, UK, 2014; p. 544. [Google Scholar]
- Tuhid, N.H.; Abdullah, N.E.; Khairi, N.M.; Saaid, M.F.; Shahrizam, M.S.B.; Hashim, H. A statistical approach for orchid disease identification using RGB color. In Proceedings of the 2012 IEEE Control and System Graduate Research Colloquium, Shah Alam, Selangor, Malaysia, 16–17 July 2012; pp. 382–385. [Google Scholar]
- Koh, K.W.; Lu, H.C.; Chan, M.T. Virus resistance in orchids. Plant Sci. 2014, 228, 26–38. [Google Scholar] [CrossRef]
- Lim, S.T.; Wong, S.M.; Goh, C.J. Elimination of Cymbidium mosaic virus and Odontoglossum ringspot virus from orchids by meristem culture and thin section culture with chemotherapy. Ann. Appl. Biol. 1993, 122, 289–297. [Google Scholar] [CrossRef]
- Panattoni, A.; Luvisi, A.; Triolo, E. Elimination of viruses in plants: Twenty years of progress. Span. J. Agri. Res. 2013, 1, 173–188. [Google Scholar]
- Gurr, S.J.; Rushton, P.J. Engineering plants with increased disease resistance: What are we going to express? Trends Biotechnol. 2005, 23, 275–282. [Google Scholar] [CrossRef]
- Pavan, S.; Jacobsen, E.; Visser, R.G.; Bai, Y. Loss of susceptibility as a novel breeding strategy for durable and broad-spectrum resistance. Mol. Breed. 2010, 25, 1. [Google Scholar] [CrossRef] [Green Version]
- Collinge, D.B.; Jørgensen, H.J.; Lund, O.S.; Lyngkjær, M.F. Engineering pathogen resistance in crop plants: Current trends and future prospects. Ann. Rev. Phytopathol. 2010, 48, 269–291. [Google Scholar] [CrossRef] [Green Version]
- Dong, X. NPR1, all things considered. Curr. Opin. Plant Boil. 2004, 7, 547–552. [Google Scholar] [CrossRef]
- Hao, L.; Zhao, Y.; Jin, D.; Zhang, L.; Bi, X.; Chen, H.; Xu, Q.; Ma, C.; Li, G. Salicylic acid-altering Arabidopsis mutants response to salt stress. Plant Soil 2012, 354, 81–95. [Google Scholar] [CrossRef]
- Jayakannan, M.; Bose, J.; Babourina, O.; Shabala, S.; Massart, A.; Poschenrieder, C.; Rengel, Z. The NPR1-dependent salicylic acid signaling pathway is pivotal for enhanced salt and oxidative stress tolerance in Arabidopsis. J. Exp. Bot. 2015, 66, 1865–1875. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Holub, E.B.; Alonso, J.M.; Ecker, J.R.; Fobert, P.R. An Arabidopsis NPR1-like gene, NPR4, is required for disease resistance. Plant J. 2005, 41, 304–318. [Google Scholar] [CrossRef] [PubMed]
- Mou, Z.; Fan, W.; Dong, X. Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell 2003, 113, 935–944. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, D.; Tasma, I.M.; Frasch, R.; Bhattacharyya, M.K. Systemic acquired resistance in soybean is regulated by two proteins, orthologous to Arabidopsis NPR1. BMC Plant Boil. 2009, 9, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishina, T.E.; Zeier, J. Pathogen-associated molecular pattern recognition rather than development of tissue necrosis contributes to bacterial induction of systemic acquired resistance in Arabidopsis. Plant J. 2007, 50, 500–513. [Google Scholar] [CrossRef] [PubMed]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Altenbach, D.; Robatzek, S. Pattern recognition receptors: From the cell surface to intracellular dynamics. Mol. Plant Microbe In. 2007, 20, 1031–1039. [Google Scholar] [CrossRef] [Green Version]
- Bigeard, J.; Colcombet, J.; Hirt, H. Signaling mechanisms in pattern-triggered immunity (PTI). Mol. Plant 2015, 8, 521–539. [Google Scholar] [CrossRef] [Green Version]
- Sticher, L.; Mauch-Mani, B.; Métraux, A.J. Systemic acquired resistance. Annu. Rev. Phytopathol. 1997, 35, 235–270. [Google Scholar] [CrossRef]
- Loake, G.; Grant, M. Salicylic acid in plant defence—The players and protagonists. Curr. Opin. Plant Biol. 2007, 10, 466–472. [Google Scholar] [CrossRef]
- Cao, H.; Bowling, S.A.; Gordon, A.S.; Dong, X. Characterization of an Arabidopsis mutant that is nonresponsive to inducers of systemic acquired resistance. Plant Cell 1994, 6, 1583–1592. [Google Scholar] [CrossRef]
- Chen, J.; Lu, H.; Chen, C.; Hsu, H.F.; Chen, H.H.; Yeh, H.H. The NPR1 ortholog PhaNPR1 is required for the induction of PhaPR1 in Phalaenopsis aphrodite. Bot. Stud. 2013, 54, 31. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Hsieh, M.H.; Chen, C.E.; Chen, H.H.; Wang, H.I.; Yeh, H.H. A high-throughput virus-induced gene-silencing vector for screening transcription factors in virus-induced plant defense response in orchid. Mol. Plant Microbe In. 2012, 25, 738–746. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Li, X.; Dong, X. Generation of broad-spectrum disease resistance by overexpression of an essential regulatory gene in systemic acquired resistance. Proc Natl. Acad. Sci. USA 1998, 95, 6531–6536. [Google Scholar] [CrossRef] [Green Version]
- Grant, M.; Lamb, C. Systemic immunity. Curr. Opin. Plant Biol. 2006, 9, 414–420. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, D.; Chu, J.Y.; Boyle, P.; Wang, Y.; Brindle, I.D.; Luca, V.D.; Després, C. The Arabidopsis NPR1 protein is a receptor for the plant defense hormone salicylic acid. Cell Rep. 2012, 1, 639–647. [Google Scholar] [CrossRef] [Green Version]
- Kinkema, M.; Fan, W.; Dong, X. Nuclear localization of NPR1 is required for activation of PR gene expression. Plant Cell 2000, 12, 2339–2350. [Google Scholar] [CrossRef] [Green Version]
- Beltagi, M.S. Exogenous ascorbic acid (vitamin C) induced anabolic changes for salt tolerance in chick pea (Cicer arietinum L.) plants. Afr. J. Plant Sci. 2008, 2, 118–123. [Google Scholar]
- Barbetti, M.J.; Banga, S.S.; Salisbury, P.A. Challenges for crop production and management from pathogen biodiversity and diseases under current and future climate scenarios–case study with oilseed Brassicas. Field Crop. Res. 2012, 127, 225–240. [Google Scholar] [CrossRef]
- Silva, K.J.P.; Mahna, N.; Mou, Z.; Folta, K.M. NPR1 as a transgenic crop protection strategy in horticultural species. Horti. Res. 2018, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Chern, M.S.; Fitzgerald, H.A.; Yadav, R.C.; Canlas, P.E.; Dong, X.; Ronald, P.C. Evidence for a disease-resistance pathway in rice similar to the NPR1-mediated signaling pathway in Arabidopsis. Plant J. 2001, 27, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Lu, C.; Wu, J.; Cheng, M.; Lin, Y.; Yang, N.; Black, L.; Green, S.K.; Wang, J.F.; Cheng, C.P. Transgenic tomato plants expressing the Arabidopsis NPR1 gene display enhanced resistance to a spectrum of fungal and bacterial diseases. Transgenic Res. 2004, 13, 567–581. [Google Scholar] [CrossRef]
- Liu, Q.; Xue, Q. Computational identification of novel PR-1-type genes in Oryza sativa. J. Genetics 2006, 85, 193. [Google Scholar] [CrossRef]
- Zhang, X.; Francis, M.I.; Dawson, W.O.; Graham, J.H.; Orbović, V.; Triplett, E.W.; Mou, Z. Over-expression of the Arabidopsis NPR1 gene in citrus increases resistance to citrus canker. Eur. J. Plant Pathol. 2010, 128, 91–100. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhang, H.; Yao, J.; Wang, X.; Xu, J.; Han, Q.; Wei, G.; Huang, L.; Kang, Z. Characterization of non-host resistance in broad bean to the wheat stripe rust pathogen. BMC Plant Biol. 2012, 12, 96. [Google Scholar] [CrossRef] [Green Version]
- Quilis, J.; Peñas, G.; Messeguer, J.; Brugidou, C.; San Segundo, B. The Arabidopsis AtNPR1 inversely modulates defense responses against fungal, bacterial, or viral pathogens while conferring hypersensitivity to abiotic stresses in transgenic rice. Mol. Plant Microbe In. 2008, 21, 1215–1231. [Google Scholar] [CrossRef] [Green Version]
- Wally, O.; Jayaraj, J.; Punja, Z.K. Broad-spectrum disease resistance to necrotrophic and biotrophic pathogens in transgenic carrots (Daucus carota L.) expressing an Arabidopsis NPR1 gene. Planta 2009, 231, 131–141. [Google Scholar] [CrossRef]
- Backer, R.; Naidoo, S.; van den Berg, N. The nonexpressor of pathogenesis-related genes 1 (NPR1) and related family: Mechanistic insights in plant disease resistance. Front. Plant Sci. 2019, 10, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.Q.; Wang, H.; Qin, R.L.; Fang, L.J.; Liu, Z.; Yuan, S.S.; Ji, X.L. Characterization of NPR1 and NPR4 genes from mulberry (Morus multicaulis) and their roles in development and stress resistance. Physiol. Plant. 2019, 167, 302–316. [Google Scholar] [CrossRef]
- Ding, Y.; Sun, T.; Ao, K.; Peng, Y.; Zhang, Y.; Li, X.; Zhang, Y. Opposite roles of salicylic acid receptors NPR1 and NPR3/NPR4 in transcriptional regulation of plant immunity. Cell 2018, 173, 1454–1467. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Rejeb, I.; Pastor, V.; Mauch-Mani, B. Plant responses to simultaneous biotic and abiotic stress: Molecular mechanisms. Plants 2014, 3, 458–475. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Kollist, H.; Zandalinas, S.I.; Sengupta, S.; Nuhkat, M.; Kangasjärvi, J.; Mittler, R. Rapid responses to abiotic stress: Priming the landscape for the signal transduction network. Trends Plant Sci. 2019, 24, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Fan, W.; Kinkema, M.; Li, X.; Dong, X. Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene. Proc. Natl. Aca. Sci. USA 1999, 96, 6523–6528. [Google Scholar] [CrossRef] [Green Version]
- Després, C.; Chubak, C.; Rochon, A.; Clark, R.; Bethune, T.; Desveaux, D.; Fobert, P.R. The Arabidopsis NPR1 disease resistance protein is a novel cofactor that confers redox regulation of DNA binding activity to the basic domain/leucine zipper transcription factor TGA1. Plant Cell 2003, 15, 2181–2191. [Google Scholar] [CrossRef] [Green Version]
- Fan, W.; Dong, X. In vivo interaction between NPR1 and transcription factor TGA2 leads to salicylic acid–mediated gene activation in Arabidopsis. Plant Cell 2002, 14, 1377–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; Huh, S.U.; Kojima, M.; Sakakibara, H.; Paek, K.H.; Hwang, I. The cytokinin-activated transcription factor ARR2 promotes plant immunity via TGA3/NPR1-dependent salicylic acid signaling in Arabidopsis. Dev. Cell 2010, 19, 284–295. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, I.; Durner, J.; Lindermayr, C. Crosstalk between nitric oxide and glutathione is required for nonexpressor of pathogenesis-related genes 1 (NPR 1)-dependent defense signaling in Arabidopsis thaliana. New Phytol. 2015, 208, 860–872. [Google Scholar] [CrossRef] [PubMed]
- Mehdy, M.C. Active oxygen species in plant defense against pathogens. Plant Physiol. 1994, 105, 467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, J.A.; Rubio, M.; Olmos, E.; Ros-Barceló, A.; Martínez-Gómez, P. Oxidative stress induced by long-term plum pox virus infection in peach (Prunus persica). Physiol. Plant. 2004, 122, 486–495. [Google Scholar] [CrossRef] [Green Version]
- Ren, R.; Yin, J.; Zheng, H.; Wang, T.; Liu, S.; Karthikeyan, A.; Yang, Q.; Gao, L.; Zhi, H.; Li, K. Characterization of broad-spectrum resistance to Soybean mosaic virus in soybean [Glycine max (L.) Merr.] cultivar ‘RN-9′. Plant Breed. 2018, 137, 605–613. [Google Scholar] [CrossRef]
- Hu, W.W.; Wong, S.M.; Loh, C.S.; Goh, C.J. Synergism in replication of cymbidium mosaic potexvirus (CymMV) and odontoglossum ringspot tobamovirus (ORSV) RNA in orchid protoplasts. Arch. Virol. 1998, 143, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.C.; Koh, A.; Loh, C.S.; Wong, S.M. Cucurbit protoplast isolation for the study of plant virus replication. J. Virology. Methods 2001, 91, 21–27. [Google Scholar] [CrossRef]
- Liang, X.; Ding, S.; Wong, S. Development of a kenaf (Hibiscuscannabinus L.) protoplast system for a replication study of Hibiscus chlorotic ringspot virus. Plant Cell Rep. 2002, 20, 982–986. [Google Scholar] [CrossRef]
- Chen, S.; Tao, L.; Zeng, L.; Vega-Sanchez, M.E.; Umemura, K.; Wang, G. A highly efficient transient protoplast system for analyzing defence gene expression and protein–protein interactions in rice. Mol. Plant Pathol. 2006, 7, 417–427. [Google Scholar] [CrossRef]
- Raghupathy, M.B.; Griffiths, J.S.; Stobbs, L.W.; Brown, D.C.; Brandle, J.E.; Wang, A. Transfection of Arabidopsis protoplasts with a Plum pox virus (PPV) infectious clone for studying early molecular events associated with PPV infection. J. Virol. Methods 2006, 136, 147–153. [Google Scholar] [CrossRef]
- Babu, M.; Griffiths, J.S.; Huang, T.S.; Wang, A. Altered gene expression changes in Arabidopsis leaf tissues and protoplasts in response to Plum pox virus infection. BMC Genom. 2008, 9, 325. [Google Scholar] [CrossRef] [Green Version]
- Iskra-Caruana, M.L.; Baurens, F.C.; Gayral, P.; Chabannes, M. A four-partner plant–virus interaction: Enemies can also come from within. Mol. Plant Microbe In. 2010, 23, 1394–1402. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Zhu, G. Digital gene expression analysis based on de novo transcriptome assembly reveals new genes associated with floral organ differentiation of the orchid plant Cymbidium ensifolium. PLoS ONE 2015, 10, e0142434. [Google Scholar] [CrossRef]
- Yang, F.; Zhu, G.; Wang, Z.; Liu, H.; Xu, Q.; Zhao, C. Integrated mRNA and microRNA transcriptome variations in the multi-tepal mutant provide insights into the floral patterning of the orchid Cymbidium goeringii. BMC Genom. 2017, 18, 367. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nature Prot. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.Y.; Chen, J.C.; Fang, S.C. A protoplast transient expression system to enable molecular, cellular, and functional studies in Phalaenopsis orchids. Front. Plant Sci. 2018, 9, 843. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.H.; Senft, J.A. An improved method to determine cell viability by simultaneous staining with fluorescein diacetate-propidium iodide. J. Histochem. Cytochem. 1985, 33, 77–79. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.H.; Hwang, D.J. Isolation and characterization of the rice NPR1 promoter. Plant Biotechnol. Rep. 2010, 4, 29–35. [Google Scholar] [CrossRef]
- Rui, R.; Liu, S.; Karthikeyan, A.; Wang, T.; Niu, H.; Yin, J.; Yang, Y.; Wang, L.; Yang, Q.; Zhi, H. Fine-mapping and identification of a novel locus Rsc15 underlying soybean resistance to Soybean mosaic virus. Theor. Appl. Genet. 2017, 130, 2395–2410. [Google Scholar] [CrossRef]
- Yu, H.H.; Wong, S.M. A DNA clone encoding the full-length infectious genome of Odontoglossum ringspot tobamovirus and mutagenesis of its coat protein gene. Arch. Virol. 1998, 143, 163–171. [Google Scholar] [CrossRef]
- Otto, F. DAPI staining of fixed cells for high-resolution flow cytometry of nuclear DNA. Methods Cell Biol. 1990, 33, 105–110. [Google Scholar]
- Sherpa, A.R.; Hallan, V.; Pathak, P.; Zaidi, A.A. Characterization of the coat protein gene of Cymbidium mosaic virus isolates from India. J. Phytopathol. 2006, 154, 275–280. [Google Scholar] [CrossRef]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, R.; Wei, Y.; Ahmad, S.; Jin, J.; Gao, J.; Lu, C.; Zhu, G.; Yang, F. Identification and Characterization of NPR1 and PR1 Homologs in Cymbidium orchids in Response to Multiple Hormones, Salinity and Viral Stresses. Int. J. Mol. Sci. 2020, 21, 1977. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21061977
Ren R, Wei Y, Ahmad S, Jin J, Gao J, Lu C, Zhu G, Yang F. Identification and Characterization of NPR1 and PR1 Homologs in Cymbidium orchids in Response to Multiple Hormones, Salinity and Viral Stresses. International Journal of Molecular Sciences. 2020; 21(6):1977. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21061977
Chicago/Turabian StyleRen, Rui, Yonglu Wei, Sagheer Ahmad, Jianpeng Jin, Jie Gao, Chuqiao Lu, Genfa Zhu, and Fengxi Yang. 2020. "Identification and Characterization of NPR1 and PR1 Homologs in Cymbidium orchids in Response to Multiple Hormones, Salinity and Viral Stresses" International Journal of Molecular Sciences 21, no. 6: 1977. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21061977