Structures of Human Transglutaminase 2: Finding Clues for Interference in Cross-linking Mediated Activity



College of Pharmacy, Chung-Ang University, Seoul 06974, Korea
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(6), 2225; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062225
Submission received: 17 February 2020
/
Revised: 19 March 2020
/
Accepted: 20 March 2020
/
Published: 23 March 2020
(This article belongs to the Special Issue The World of Transglutaminases: From Basic Biological and Medical Research to Applied Sciences)
Abstract
:Human transglutaminase 2 (TGase2) has various functions, including roles in various cellular processes such as apoptosis, development, differentiation, wound healing, and angiogenesis, and is linked to many diseases such as cancer. Although TGase2 has been considered an optimized drug target for the treatment of cancer, fibrosis, and neurodegenerative disorders, it has been difficult to generate TGase2-targeted drugs for clinical use because of the relatively flat and broad active site on TGase2. To design more specific and powerful inhibitors, detailed structural information about TGase2 complexed with various effector and inhibitor molecules is required. In this review, we summarized the current structural studies on TGase2, which will aid in designing drugs that can overcome the aforementioned limitations.
1. Introduction
Transglutaminases (TGases) are enzymes that catalyze the cross-linking reaction between the γ-carboxamide of a glutamine residue and the ε-amino group of a lysine residue; they also catalyze the deamidation reaction of glutamine residues to generate glutamic acid in the presence of water [1,2]. These enzymes exhibit cross-linking activity, which helps create stable, barrier-like structures such as blood-clots, skin, and hair [1,3,4]. In humans, eight isoforms of TGases, TGase1~7 and Factor XIII, have been identified and characterized [5].
Human TGase2 (also known as tissue TGase) is a multi-functional, ubiquitous enzyme that is important for various cellular processes, including apoptosis [6,7,8], development [9,10], differentiation [11,12], wound healing [13,14], and angiogenesis [14,15]. Human TGase2 comprises 687 amino acids (~70 kDa) and has a four-domain organization. It contains the N-terminal β-sandwich domain, the catalytic core domain containing the catalytic triad, and two β-barrel domains (β-barrel1 and β-barrel2) [16]. Using this domain organization, TGase2 performs multiple functions; it acts as a GTPase [17,18,19], kinase [20,21], protein disulfide isomerase [22], and scaffolding factor [23], and performs the traditional activity of cross-linking by transamidation. The multiple functions of TGase2 are controlled in various ways in the cell, such as by cellular cofactors [24,25], spatial localization of TGase2 [13,26], endogenous amine compounds and proteins [27,28,29,30,31], and post-translational modifications [32]. Non-functional and dysregulated TGase2 is related to various human diseases, such as celiac disease [33], inflammatory disease [34], many types of cancers [3,35,36,37,38], tissue fibrosis [39], diabetes [40], pulmonary hypertension by fibrogenic remodeling [41], and neurodegenerative diseases [42,43,44]. It has been reported that overexpression or over-activity due to mutations in TGase2 leads to celiac disease [45], inflammatory disease [34], and tissue fibrosis [39]. Both downregulated and upregulated TGase2 are known to cause many types of cancer [8,46]. TGase2 has been found to be downregulated in sarcoma [35,47], whereas its overactivation has been observed in pancreatic and breast cancer [46,48]. Hyperactivity of TGase2 in the pathogenic stages of neurodegenerative diseases, including Alzheimer’s disease (AD), has also been reported [49,50,51,52]. These studies show that hyperactivated TGase2 cross-links several AD-related proteins, including tau, Aβ, and α-synuclein, causing the accumulation of these proteins in patients with AD [50,51,52]. Misregulated TGase2 activity due to mutations has been found in the early stages of type 2 diabetes, indicating that its activity is also linked to metabolic diseases [40].
Owing to its biologically and pathogenically important functions, TGase2 has been considered a major therapeutic target for the treatment of many human diseases caused by dysregulated TGase2 [29,53,54,55,56]. The aim of this review was to summarize the recent progress in the structural studies of TGase2, which can aid in the development of drugs targeting this enzyme. We will discuss its structural complexity and several TGase2 structures complexed with effector molecules.
2. Multi-functional TGase2
TGase2 is a multi-functional protein. Cross-linking, deamination, and amine incorporation activities are the main functions of TGase2, GTPase, disulfide isomerase, and kinase. Scaffolding activities are also reported as minor functions of TGase2 [16,57]. The principal transamidation activity is performed by the catalytic triad, composed of C277, H335, and D358, on the active site of TGase2. The thiol functional group of C277 on TGase2 works as a nucleophile and attacks the carboxamide of a glutamine residue, forming a thioester intermediate during the transamidation reaction. Then, this unstable intermediate is broken down by the ε-amino group of the lysine residue, producing a stable isopeptide bond that cross-links the two substrates (Figure 1A).
The protein disulfide isomerase (PDI) activity of TGase2, which was initially reported in studies of RNase A [22] and adenine nucleotide translocator 1 (ANT1) [58], is another important function of TGase2. An initial PDI activity study showed that reduced, denatured, inactive RNase A was converted into the renatured, active form by the PDI activity of TGase2 [58]. Neither Ca2+ (activator of transamidase activity) nor guanosine triphosphate (GTP, inhibitor of transamidase activity) affect the PDI activity of TGase2 [58]. A mutagenesis study showed that the cysteine residue in the active site is not critical for the PDI activity of TGase2, indicating that the two reactions, transamidation and PDI, are independent. Another TGase2 substrate for PDI activity is the ANT1 protein, which is one of the components of the mitochondrial membrane permeability transition pore (PTP) [59,60]. During apoptosis, ANT1 is oligomerized and forms a pore on the mitochondrial membrane owing to the cross-linking and PDI activities of TGase2 [58]. This causes the cell to release various pro-apoptotic molecules, leading to apoptosis. This indicates that the PDI activity of TGase2 is involved in mitochondria-dependent apoptosis.
When TGase2 was first characterized, it was assumed that the nucleotide-binding pocket on TGase2 strongly indicated that it might have GTPase and kinase activities. Since then, GTPase activity during signal transduction has been reported [17]. In this study, TGase2 mediated between the α1-adrenergic receptor and phospholipase Cδ in the liver during proliferation signaling of hepatocytes, which is critical for the GTPase activity of TGase2 [17]. A study on the kinase activity of TGase2 on insulin-like growth factor-binding protein-3 (IGFBP-3) was the first to report the kinase activity of TGase2 [61]. The study indicated that TGase2 phosphorylated both IGFBP-3 and IGFBP-5 at multiple serine residues. The kinase activity of TGase2 was inhibited by Ca2+, which is known to be an activator of the transamidation activity of TGase2 [61]. Many additional substrates, including P53 [21], histones [20], retinoblastoma protein (Rb) [62], the transmembrane protein CDH1 [63], and the extracellular protein MMP-3 [63], have since been reported. TGase2 can form a complex with fibronectin and collagen in the extracellular matrix (ECM) and mediate the interaction between fibronectin and integrins during the cell adhesion process. Its scaffolding activity, which is not related to the cross-linking activity for ECM turnover, has also been highlighted [23,64].
As the functions of TGase2 are critical for the survival of the organism owing to its involvement in various cellular processes, TGase2 activities have to be finely controlled. It has been reported that the activity of TGase2 is controlled by cellular cofactors, spatial localization, endogenous protein regulators, and post-translational modifications [13,24,25,26,27,28,29,30,31,32]. Various cellular ions and nucleotides are the main regulators of the multi-functional activities of TGase2. Although the activation mechanism is still unclear, the most well-known activator of the transamidation activity of TGase2 is the Ca2+ ion. Ca2+ was initially assumed to promote the structural transition of TGase2 to its active form. A recent study, however, showed that Ca2+ binding to TGase2 did not lead to a dramatic structural transition [65]. In contrast, nucleotides such as GTP, GDP (guanosine diphosphate), and ATP (adenosine triphosphate) are well-known inhibitors of the transamidation activity of TGase2 [66]. Nucleotide binding to the relatively well-defined nucleotide-binding site on TGase2, which is located between the catalytic core domain and the first β-barrel, maintains the compact, inactive form of TGase2 via the formation of hydrogen bonds between a cysteine in the active site and a tyrosine residue nearby; this blocks the structural transition to the active form [67,68,69].
Endogenous amine compounds, such as cystamine, cysteamine, spermidine, and histamine, are another class of TGase2 inhibitors [53,70]. As these amine compounds can work as amine donors that can cross-link with glutamine in the substrate, they can inhibit the transamidase activity of TGase2 by competing with natural substrates [28]. Another known inhibition mechanism used by the amine compounds is the direct modification of the active site cysteine (C277) and other surface-exposed cysteines on TGase2, which are crucial for TGase2 activity, by the formation of a disulfide bond [71,72]. It has been reported that several disulfide bonds on TGase2, which are formed or unformed based on the oxidation/reduction states, were critical for the regulation of allosteric activity. This indicates that the amine compound-mediated cysteine modification and the formation of disulfide bonds are important processes for TGase2 activity control [31,73]. Endogenous proteins can also control TGase2 activity. Ribosomal proteins RPL7a and RPL13, which are the most well-known endogenous protein inhibitors of TGase2, inhibit transamidase activity by binding to the β-barrel2 domain of TGase2 [29]. Besides the direct activity control by endogenous small molecules, macromolecules, and ions, cytokine- and reactive oxygen species (ROS)-mediated indirect mechanisms of TGase2 activity control have also been suggested [30,74,75].
3. Structures of TGase2 Complexed with Various Effectors
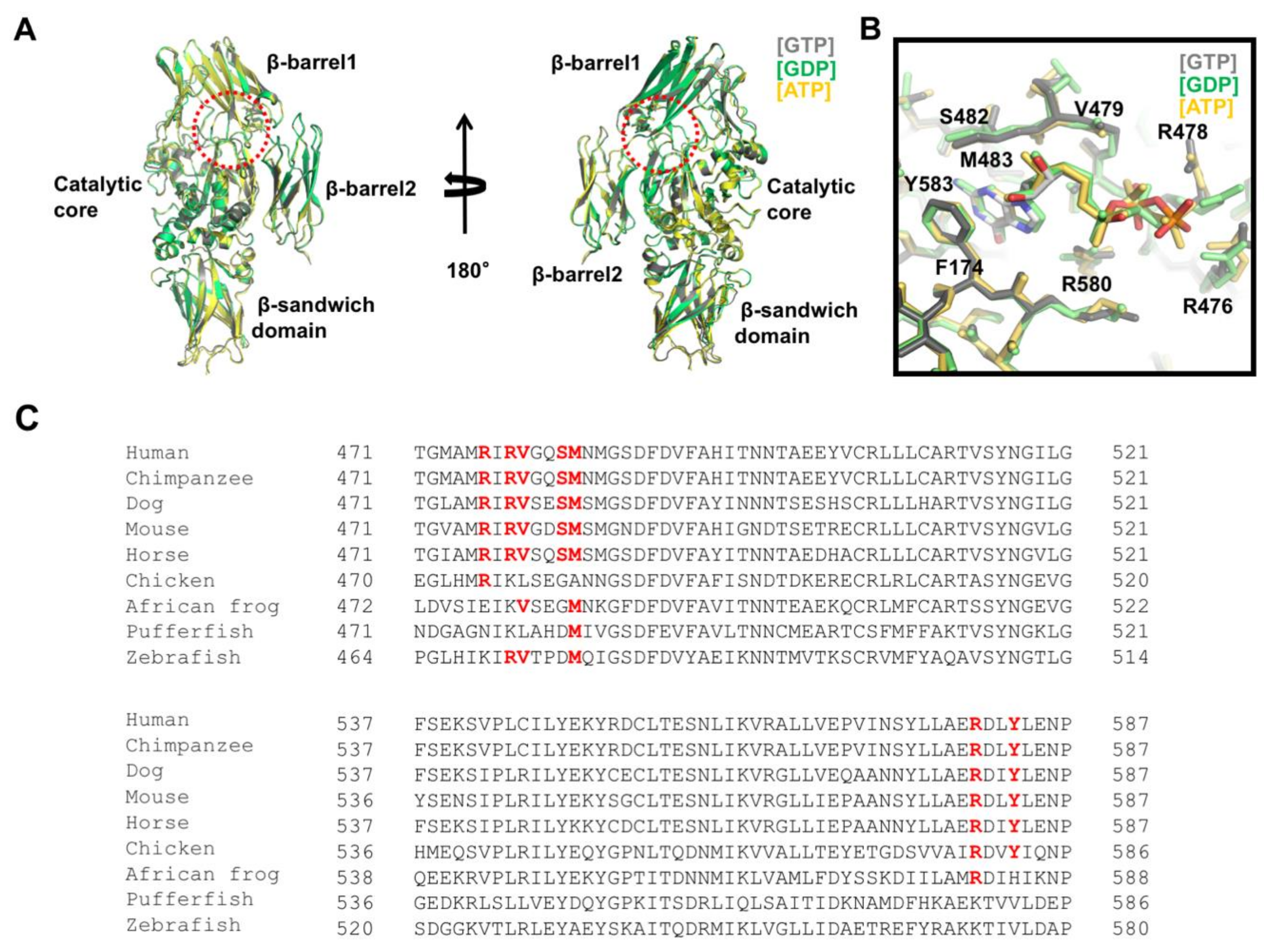
The multi-functional activity of TGase2 is finely controlled by various molecules in the cell. To understand the molecular mechanism of the activity control of TGase2 by various effector molecules, structural studies of TGase2 complexed with effector molecules have been performed. The structures of three nucleotides, GTP, GDP, and ATP-binding TGase2 have been reported [67,69,76]. These structures showed that a unique nucleotide-binding site was present in between the catalytic core and the first β–barrel (Figure 2A). The three structures were nearly identical, having a root mean square deviation of ~0.5 Å with each other, indicating that different nucleotides did not affect the overall structure of TGase2 (Figure 2A).
Although TGase2 possesses GTPase and ATPase activities, the major role of the nucleotides is to inactivate its activity by blocking structural transition, which is important for retaining the activity of TGase2. Structural analysis of the GTP-binding site of TGase2 indicated that three positively charged arginines, R476, R478, and R580, surround the negatively charged phosphate moieties of the nucleotide by forming hydrogen bonds, whereas hydrophobic residues from TGase2, including F174, V479, and M483, stabilize the hydrophobic base moiety on the nucleotides via hydrophobic interactions (Figure 2B) [67]. Among the residues accommodating the nucleotides, R580 was the most critical residue for the nucleotide interaction. Mutations in R580, which formed two hydrogen bonds with the phosphate of the nucleotides, abolished the nucleotide-binding capacity and eliminated its sensitivity to GTP [77,78]. R478 is involved in nucleotide binding via the formation of hydrogen bonds with the γ-phosphate of GTP. The structure of GDP-bound TGase2 showed that the side chain of R478 moved toward the γ-phosphate position, which is the most critical structural difference between GTP-bound and GDP-bound TGase2 (Figure 2B) [69]. Binding preference between GTP and ATP on TGase2 was identified by a structural study [67]. S482 and Y583 residues are involved in the interaction with only GTP, not ATP, indicating that GTP is the preferable nucleotide owing to a tighter interaction. Indeed, TGase2 activity is more strongly inhibited by GTP than ATP [25]. For nucleotide binding, Mg2+ is not necessary, unlike other GTP-binding proteins, including various small G proteins [67].
Amino acid sequence analysis of TGase2 indicates that the nucleotide-binding pocket generated by the residues R476, R478, V479, S482, M483, R580, and Y583 is only conserved in mammals (Figure 2C). This site is not formed in birds, frogs, or fish, indicating that the use of nucleotides in TGase2 has evolved over a long period of time.
4. Structures of TGase2 Complexed with Inhibitors Derived from Peptide Mimetics
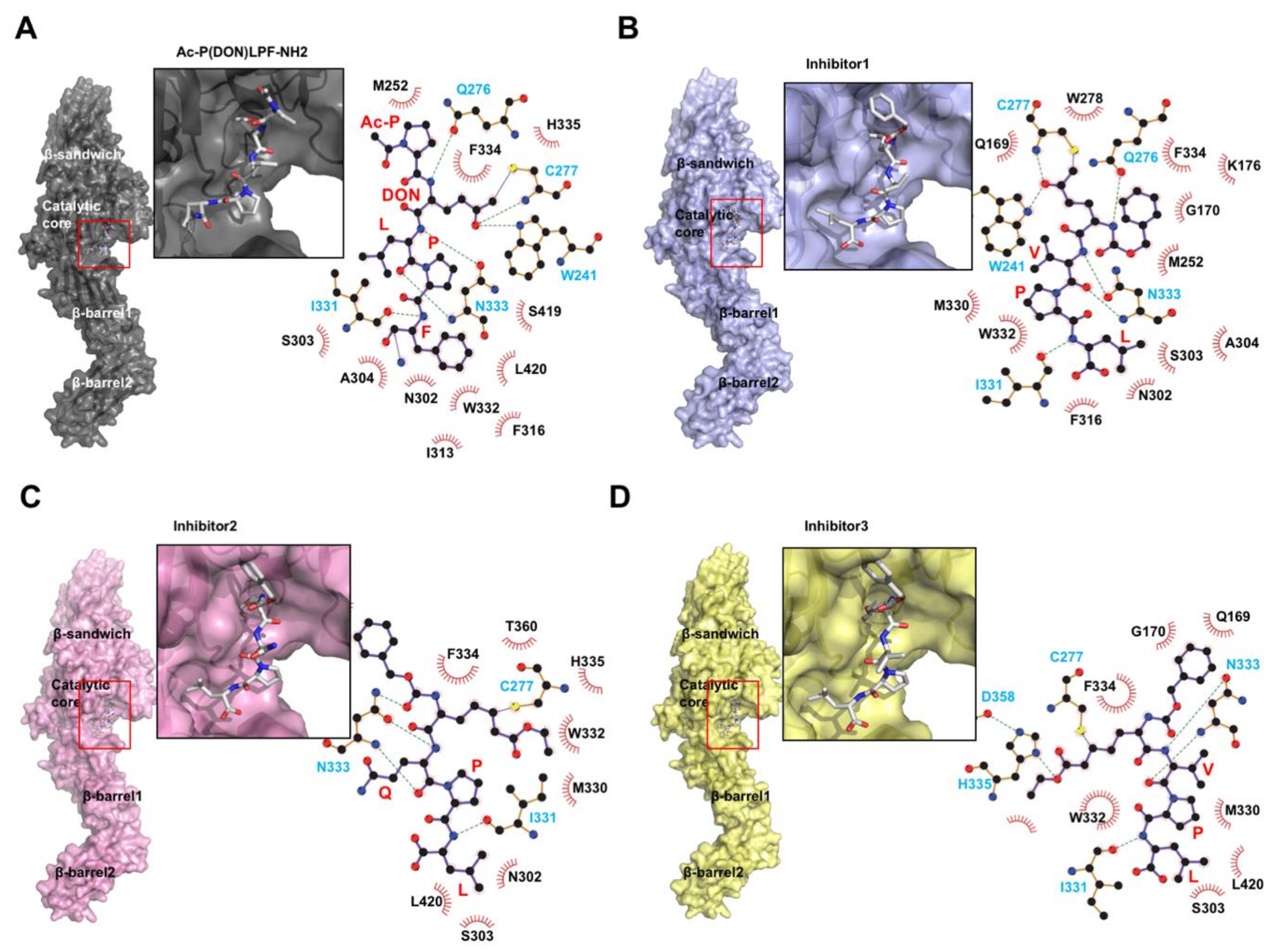
While nucleotides maintain the closed form and inhibit TGase2 activities, substrate derived-peptide mimetics, which work as irreversible inhibitors, promote the open, active conformation of TGase2 (Figure 1B and Figure 3A) [79]. The first introduced inhibitor peptide, known as Ac-P(DON)LPF-NH2, where DON is the electrophilic amino acid 6-diazo-5-oxo-L-norleucine, derived from the penta-peptide PQLPY from gluten, is one of the substrates of TGase2. The TGase2/Ac-P(DON)LPF-NH2 complex structure shows that the peptide mimetic attaches to the active site C277 via a thioether covalent linkage (Figure 3A) [79]. The LPF peptide region of the inhibitor is well-positioned toward the hydrophobic pocket formed by S303, A304, I313, F316, I331, and L420 on TGase2. W241, Q276, C277, and N333 form hydrogen bonds with the inhibitor. Since this inhibitor-bound, open form structure was introduced, three more TGase2 structures complexed with peptide mimetic inhibitors have been deposited in The Protein Data Bank without publication. All three peptide mimetic inhibitors are modified versions of Ac-P(DON)LPF-NH2. The first inhibitor is N-[(benzyloxy)carbonyl]-6-diazonio-5-oxo-L-norleucyl-L-valyl-L-prolyl-L-leucine. This inhibitor is also a penta-peptide mimetic containing a VPL sequence instead of an LPF sequence. Similar to Ac-P(DON)LPF-NH2 binding, this inhibitor also irreversibly binds to C277 on TGase2 via a thioether linkage (Figure 3B). Hydrogen bonds are formed between the inhibitor ketone and the indole of W241, and the backbone amide of C277. The peptide-binding pocket of the other three inhibitors, formed by S303, A304, I313, F316, I331, and W332, is also similar to the Ac-P(DON)LPF-NH2 binding pocket. The second peptide mimetic inhibitor is N~2~-[(2S)-2-{[(benzyloxy)carbonyl]amino}-7-ethoxy-7-oxoheptanoyl]-L-glutaminyl-L-prolyl-L-leucine. This inhibitor contains a QPL sequence. QPL binds to the conserved substrate- or peptide mimetic inhibitor-binding pocket. Although this peptide mimetic also covalently binds to C277 on TGase2, which is similar to that observed in Ac-P(DON)LPF-NH2 binding, because of the absence of indole, the peptide inhibitor cannot form hydrogen bonds with C277 and the neighboring W241 (Figure 3C). Instead, N333 is more tightly involved in the peptide binding, forming three hydrogen bonds (Figure 3C). The last peptide mimetic inhibitor is N-[(2R)-2-{[(benzyloxy)carbonyl]amino}-7-ethoxy-7-oxoheptanoyl]-L-valyl-L-prolyl-L-leucine. This peptide contains a VPL sequence, and its interaction is similar to that of the second inhibitor. The modified, long side chain at the second position of this penta-peptide inhibitor interacts with C277 via a thioether linkage, and with H335 and D358 via hydrogen bonds in the deep active site (Figure 3D). This is the only peptide inhibitor that can interact with all three residues from the catalytic triad in the active site of TGase2.
5. Future Perspectives
With important roles in many pathological conditions and various activities, TGase2 has been considered an optimized target for therapeutic intervention [53,54,81]. In this effort, TGase2-targeting drugs have developed. Although several small molecules and peptide mimetic inhibitors have shown inhibiting capacity in animal models and clinical trials, no drugs are currently in use in clinical settings.
The screening and discovery of TGase2-targeting drugs by examining small molecules was not initially successful. Owing to its relatively flat and broad active site, finding small molecules that target the active site of TGase2 has always been challenging. To overcome this issue, many researchers have used peptide mimetics to cover the broad active site. Because of this effort, several potential peptide mimetic inhibitors are currently in clinical trials [82]. The nucleotide-binding site on TGase2 has also been targeted by non-hydrolyzable GTP analogs, which can bind to and maintain the inactive (closed) form of TGase2. However, this strategy was not successful owing to specificity [83]. To design high affinity optimized inhibitors more efficiently, structural information is critical and has been employed (Table 1).
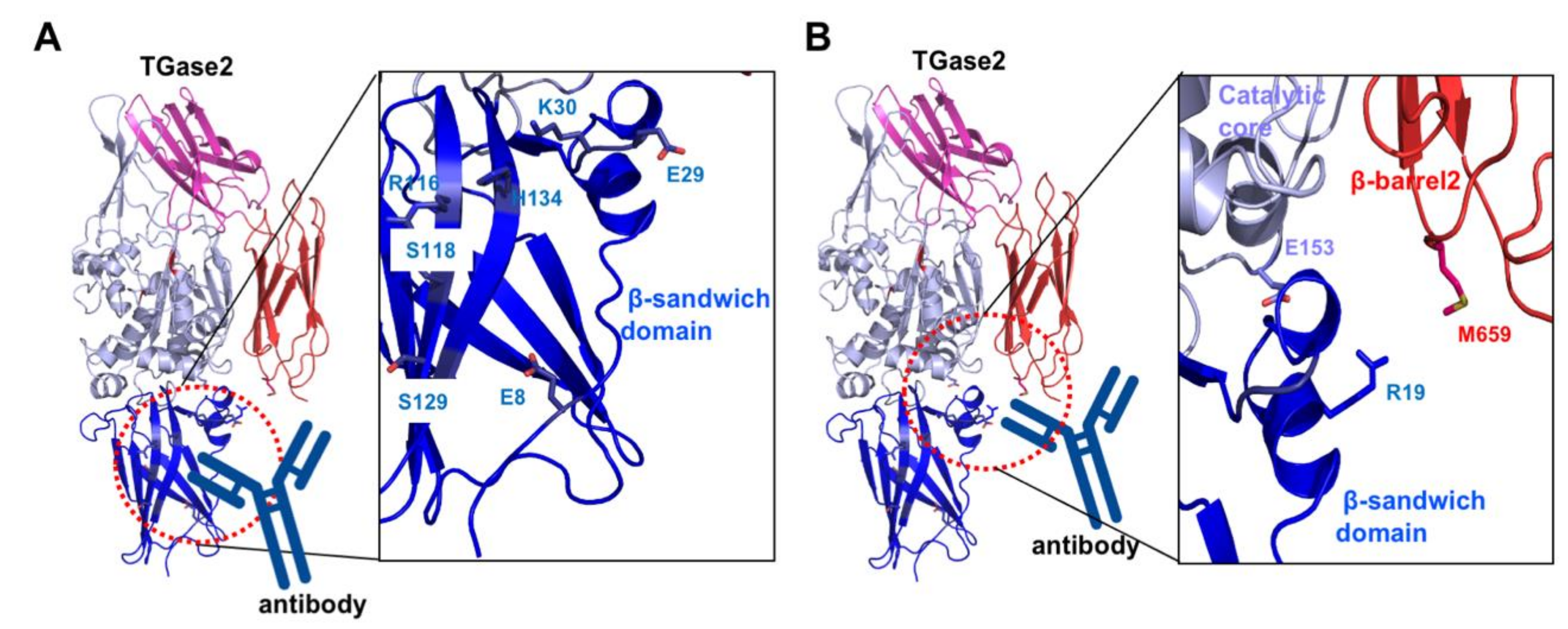
Besides finding small molecules and peptide mimetics that target the active site or the nucleotide-binding site of TGase2 for pharmaceutical purposes, various approaches to discover drugs for TGase2 inhibition are undergoing. One of the representative cases is the search for an antibody against TGase2. Celiac disease is caused by an autoimmune response to dietary gluten antigens. This antigen produces autoantibodies against TGase2 [84]. The effect of autoantibody in the fibroblasts from celiac disease patient [85], and B cell tolerance and autoantibody production in mouse models have been studied recently [86]. Recent structural and biophysical studies mapped the autoantibody-binding sites to TGase2 [87]. The first site is located in the N-terminal β-sandwich domain. Residues E8, E29, K30, R116, S118, S129, and H134 are involved in the antibody binding (Figure 4A) [87]. The second autoantibody-binding site is the cleft region formed by the β-sandwich, catalytic core, and β-barrel2 domains (Figure 4B) [88]. R19, E153, and M659 are the residues responsible for binding to the autoantibodies [88]. The inhibition of autoantibody interactions with TGase2 might be a good strategy for the treatment of celiac disease. The investigation of small interfering RNA (siRNA) targeting TGase2 expression has also been attempted for the development of drugs for the treatment of several types of cancers related to the aberrant expression of TGase2 [89]. Endogenous TGase2 inhibitors, such as the ribosomal proteins RPL7a and RPL13 might be important sources for TGase2 targeting [29]. Structural studies of TGase2 with various endogenous inhibitors need to be performed to aid drug design.
Author Contributions
Conceptualization, H.H.P.; Writing—Original Draft Preparation, G.E.K. and H.H.P. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Basic Science Research Program of the National Research Foundation of Korea (NRF), funded by the Korea government (MSIT) (NRF-2017M3A9D8062960 and NRF-2018R1A2B2003635), and a grant from the Korea Healthcare Technology R&D Project, Ministry of Health & Welfare, Republic of Korea (HI17C0155).
Acknowledgments
The authors would like to thank the members of Park’s lab for helping with reference collection and their helpful discussions.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Griffin, M.; Casadio, R.; Bergamini, C.M. Transglutaminases: Nature’S biological glues. Biochem. J. 2002, 368, 377–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorand, L.; Graham, R.M. Transglutaminases: Crosslinking enzymes with pleiotropic functions. Nat. Rev. Mol. Cell Biol. 2003, 4, 140–156. [Google Scholar] [CrossRef] [PubMed]
- Birckbichler, P.J.; Orr, G.R.; Conway, E.; Patterson, M.K., Jr. Transglutaminase activity in normal and transformed cells. Cancer Res. 1977, 37, 1340–1344. [Google Scholar]
- Sicker, T.; Hilgenfeld, R. Blood coagulation factor XIII: Activation, substrates and structure of a transglutaminase. Hamostaseologie 2002, 22, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Beninati, S.; Piacentini, M. The transglutaminase family: An overview: Minireview article. Amino Acids 2004, 26, 367–372. [Google Scholar] [CrossRef]
- Oliverio, S.; Amendola, A.; Di Sano, F.; Farrace, M.G.; Fesus, L.; Nemes, Z.; Piredda, L.; Spinedi, A.; Piacentini, M. Tissue transglutaminase-dependent posttranslational modification of the retinoblastoma gene product in promonocytic cells undergoing apoptosis. Mol. Cell. Biol. 1997, 17, 6040–6048. [Google Scholar] [CrossRef] [Green Version]
- Nemes, Z., Jr.; Adany, R.; Balazs, M.; Boross, P.; Fesus, L. Identification of cytoplasmic actin as an abundant glutaminyl substrate for tissue transglutaminase in HL-60 and U937 cells undergoing apoptosis. J. Biol. Chem. 1997, 272, 20577–20583. [Google Scholar] [CrossRef] [Green Version]
- Piacentini, M.; Fesus, L.; Farrace, M.G.; Ghibelli, L.; Piredda, L.; Melino, G. The expression of “tissue” transglutaminase in two human cancer cell lines is related with the programmed cell death (apoptosis). Eur. J. Cell Biol. 1991, 54, 246–254. [Google Scholar]
- Kaartinen, M.T.; El-Maadawy, S.; Rasanen, N.H.; McKee, M.D. Tissue transglutaminase and its substrates in bone. J. Bone Miner. Res. 2002, 17, 2161–2173. [Google Scholar] [CrossRef]
- Aeschlimann, D.; Mosher, D.; Paulsson, M. Tissue transglutaminase and factor XIII in cartilage and bone remodeling. Semin. Thromb. Hemost. 1996, 22, 437–443. [Google Scholar] [CrossRef]
- Matic, I.; Sacchi, A.; Rinaldi, A.; Melino, G.; Khosla, C.; Falasca, L.; Piacentini, M. Characterization of transglutaminase type II role in dendritic cell differentiation and function. J. Leukoc. Biol. 2010, 88, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tee, A.E.; Marshall, G.M.; Liu, P.Y.; Xu, N.; Haber, M.; Norris, M.D.; Iismaa, S.E.; Liu, T. Opposing effects of two tissue transglutaminase protein isoforms in neuroblastoma cell differentiation. J. Biol. Chem. 2010, 285, 3561–3567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upchurch, H.F.; Conway, E.; Patterson, M.K., Jr.; Maxwell, M.D. Localization of cellular transglutaminase on the extracellular matrix after wounding: Characteristics of the matrix bound enzyme. J. Cell. Physiol. 1991, 149, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Haroon, Z.A.; Hettasch, J.M.; Lai, T.S.; Dewhirst, M.W.; Greenberg, C.S. Tissue transglutaminase is expressed, active, and directly involved in rat dermal wound healing and angiogenesis. FASEB J. 1999, 13, 1787–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.A.; Kotsakis, P.; Johnson, T.S.; Chau, D.Y.; Ali, S.; Melino, G.; Griffin, M. Matrix changes induced by transglutaminase 2 lead to inhibition of angiogenesis and tumor growth. Cell Death Differ. 2006, 13, 1442–1453. [Google Scholar] [CrossRef]
- Lee, C.S.; Park, H.H. Structural aspects of transglutaminase 2: Functional, structural, and regulatory diversity. Apoptos. Int. J. Program. Cell Death 2017, 22, 1057–1068. [Google Scholar] [CrossRef]
- Nakaoka, H.; Perez, D.M.; Baek, K.J.; Das, T.; Husain, A.; Misono, K.; Im, M.J.; Graham, R.M. Gh: A GTP-binding protein with transglutaminase activity and receptor signaling function. Science 1994, 264, 1593–1596. [Google Scholar] [CrossRef]
- Mian, S.; el Alaoui, S.; Lawry, J.; Gentile, V.; Davies, P.J.; Griffin, M. The importance of the GTP-binding protein tissue transglutaminase in the regulation of cell cycle progression. FEBS Lett. 1995, 370, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Hitomi, K.; Ikura, K.; Maki, M. GTP, an inhibitor of transglutaminases, is hydrolyzed by tissue-type transglutaminase (TGase 2) but not by epidermal-type transglutaminase (TGase 3). Biosci. Biotechnol. Biochem. 2000, 64, 657–659. [Google Scholar] [CrossRef]
- Mishra, S.; Saleh, A.; Espino, P.S.; Davie, J.R.; Murphy, L.J. Phosphorylation of histones by tissue transglutaminase. J. Biol. Chem. 2006, 281, 5532–5538. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Murphy, L.J. The p53 oncoprotein is a substrate for tissue transglutaminase kinase activity. Biochem. Biophys. Res. Commun. 2006, 339, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, G.; Suwa, M.; Ichikawa, Y.; Ohtsuka, T.; Kumagai, S.; Kikuchi, M.; Sato, Y.; Saito, Y. A novel function of tissue-type transglutaminase: Protein disulphide isomerase. Biochem. J. 2003, 373, 793–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akimov, S.S.; Krylov, D.; Fleischman, L.F.; Belkin, A.M. Tissue transglutaminase is an integrin-binding adhesion coreceptor for fibronectin. J. Cell Biol. 2000, 148, 825–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiraly, R.; Csosz, E.; Kurtan, T.; Antus, S.; Szigeti, K.; Simon-Vecsei, Z.; Korponay-Szabo, I.R.; Keresztessy, Z.; Fesus, L. Functional significance of five noncanonical Ca2+-binding sites of human transglutaminase 2 characterized by site-directed mutagenesis. FEBS J. 2009, 276, 7083–7096. [Google Scholar] [CrossRef] [Green Version]
- Begg, G.E.; Carrington, L.; Stokes, P.H.; Matthews, J.M.; Wouters, M.A.; Husain, A.; Lorand, L.; Iismaa, S.E.; Graham, R.M. Mechanism of allosteric regulation of transglutaminase 2 by GTP. Proc. Natl. Acad. Sci. USA 2006, 103, 19683–19688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesort, M.; Attanavanich, K.; Zhang, J.; Johnson, G.V. Distinct nuclear localization and activity of tissue transglutaminase. J. Biol. Chem. 1998, 273, 11991–11994. [Google Scholar] [CrossRef] [Green Version]
- Lentini, A.; Abbruzzese, A.; Caraglia, M.; Marra, M.; Beninati, S. Protein-polyamine conjugation by transglutaminase in cancer cell differentiation: Review article. Amino Acids 2004, 26, 331–337. [Google Scholar] [CrossRef]
- Jeitner, T.M.; Delikatny, E.J.; Ahlqvist, J.; Capper, H.; Cooper, A.J. Mechanism for the inhibition of transglutaminase 2 by cystamine. Biochem. Pharmacol. 2005, 69, 961–970. [Google Scholar] [CrossRef]
- Hitomi, K.; Kojima, S.; Fesus, L. Transglutaminase, Multiple Functional Modifiers and Targets for New Drug Discovery, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Diraimondo, T.R.; Klock, C.; Khosla, C. Interferon-gamma activates transglutaminase 2 via a phosphatidylinositol-3-kinase-dependent pathway: Implications for celiac sprue therapy. J. Pharmacol. Exp. Ther. 2012, 341, 104–114. [Google Scholar] [CrossRef] [Green Version]
- Plugis, N.M.; Palanski, B.A.; Weng, C.H.; Albertelli, M.; Khosla, C. Thioredoxin-1 selectively activates transglutaminase 2 in the extracellular matrix of the small intestine: Implications for celiac disease. J. Biol. Chem. 2017, 292, 2000–2008. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.C.; Chiang, B.Y.; Chou, C.C.; Chen, T.C.; Chen, Y.J.; Chen, Y.J.; Lin, C.H. Glutathionylspermidine in the modification of protein SH groups: The enzymology and its application to study protein glutathionylation. Molecules 2015, 20, 1452–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieterich, W.; Ehnis, T.; Bauer, M.; Donner, P.; Volta, U.; Riecken, E.O.; Schuppan, D. Identification of tissue transglutaminase as the autoantigen of celiac disease. Nat. Med. 1997, 3, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y. Transglutaminase 2 in inflammation. Front. Biosci. 2006, 11, 3026–3035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, R.N.; Bungay, P.J.; Elliott, B.M.; Walton, P.L.; Griffin, M. Alterations in the distribution and activity of transglutaminase during tumour growth and metastasis. Carcinogenesis 1985, 6, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Mangala, L.S.; Mehta, K. Tissue transglutaminase (TG2) in cancer biology. Prog. Exp. Tumor Res. 2005, 38, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Delaine-Smith, R.; Wright, N.; Hanley, C.; Hanwell, R.; Bhome, R.; Bullock, M.; Drifka, C.; Eliceiri, K.; Thomas, G.; Knight, M.; et al. Transglutaminase-2 mediates the biomechanical properties of the colorectal cancer tissue microenvironment that contribute to disease progression. Cancers 2019, 11, 701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, S.; Moon, Y.; Choi, J.; Yoon, S.; Jung, K.H.; Cheon, J.; Kim, W.; Kim, D.; Lee, C.H.; Kim, S.W.; et al. The GTP binding activity of transglutaminase 2 promotes bone metastasis of breast cancer cells by downregulating microRNA-205. Am. J. Cancer Res. 2019, 9, 597–607. [Google Scholar]
- Griffin, M.; Smith, L.L.; Wynne, J. Changes in transglutaminase activity in an experimental model of pulmonary fibrosis induced by paraquat. Br. J. Exp. Pathol. 1979, 60, 653–661. [Google Scholar]
- Porzio, O.; Massa, O.; Cunsolo, V.; Colombo, C.; Malaponti, M.; Bertuzzi, F.; Hansen, T.; Johansen, A.; Pedersen, O.; Meschi, F.; et al. Missense mutations in the TGM2 gene encoding transglutaminase 2 are found in patients with early-onset type 2 diabetes. Mutation in brief no. 982. Online. Hum. Mutat. 2007, 28, 1150. [Google Scholar] [CrossRef]
- Bhedi, C.D.; Nasirova, S.; Toksoz, D.; Warburton, R.R.; Morine, K.J.; Kapur, N.K.; Galper, J.B.; Preston, I.R.; Hill, N.S.; Fanburg, B.L.; et al. Glycolysis regulated transglutaminase 2 activation in cardiopulmonary fibrogenic remodeling. FASEB J. 2020, 34, 930–944. [Google Scholar] [CrossRef] [Green Version]
- Lesort, M.; Tucholski, J.; Miller, M.L.; Johnson, G.V. Tissue transglutaminase: A possible role in neurodegenerative diseases. Prog. Neurobiol. 2000, 61, 439–463. [Google Scholar] [CrossRef]
- Hoffner, G.; Djian, P. Transglutaminase and diseases of the central nervous system. Front. Biosci. 2005, 10, 3078–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basso, M.; Milelli, A. Transglutaminases, neuronal cell death and neural repair: Implications for traumatic brain injury and therapeutics. Curr. Opin. Neurol. 2019, 32, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Reif, S.; Lerner, A. Tissue transglutaminase--the key player in celiac disease: A review. Autoimmun. Rev. 2004, 3, 40–45. [Google Scholar] [CrossRef]
- .Mehta, K.; Kumar, A.; Kim, H.I. Transglutaminase 2: A multi-tasking protein in the complex circuitry of inflammation and cancer. Biochem. Pharmacol. 2010, 80, 1921–1929. [Google Scholar] [CrossRef]
- Birckbichler, P.J.; Bonner, R.B.; Hurst, R.E.; Bane, B.L.; Pitha, J.V.; Hemstreet, G.P., III. Loss of tissue transglutaminase as a biomarker for prostate adenocarcinoma. Cancer 2000, 89, 412–423. [Google Scholar] [CrossRef]
- Mangala, L.S.; Fok, J.Y.; Zorrilla-Calancha, I.R.; Verma, A.; Mehta, K. Tissue transglutaminase expression promotes cell attachment, invasion and survival in breast cancer cells. Oncogene 2007, 26, 2459–2470. [Google Scholar] [CrossRef] [Green Version]
- Ishizawa, T.; Mattila, P.; Davies, P.; Wang, D.; Dickson, D.W. Colocalization of tau and alpha-synuclein epitopes in Lewy bodies. J. Neuropathol. Exp. Neurol. 2003, 62, 389–397. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.S.; Dickson, D.W.; Malter, J.S. Tissue transglutaminase, protein cross-linking and Alzheimer’s disease: Review and views. Int. J. Clin. Exp. Pathol. 2008, 1, 5–18. [Google Scholar]
- Dudek, S.M.; Johnson, G.V. Transglutaminase facilitates the formation of polymers of the beta-amyloid peptide. Brain Res. 1994, 651, 129–133. [Google Scholar] [CrossRef]
- Norlund, M.A.; Lee, J.M.; Zainelli, G.M.; Muma, N.A. Elevated transglutaminase-induced bonds in PHF tau in Alzheimer’s disease. Brain Res. 1999, 851, 154–163. [Google Scholar] [CrossRef]
- Siegel, M.; Khosla, C. Transglutaminase 2 inhibitors and their therapeutic role in disease states. Pharmacol. Ther. 2007, 115, 232–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Hwang, H.; Im, C.Y.; Kim, S.Y. Recent progress in the development of transglutaminase 2 (TGase2) inhibitors. J. Med. Chem. 2017, 60, 554–567. [Google Scholar] [CrossRef] [PubMed]
- Katt, W.P.; Antonyak, M.A.; Cerione, R.A. The diamond anniversary of tissue transglutaminase: A protein of many talents. Drug Discov. Today 2018, 23, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Palucci, I.; Maulucci, G.; De Maio, F.; Sali, M.; Romagnoli, A.; Petrone, L.; Fimia, G.M.; Sanguinetti, M.; Goletti, D.; De Spirito, M.; et al. Inhibition of Transglutaminase 2 as a Potential Host-Directed Therapy Against Mycobacterium tuberculosis. Front. Immunol. 2020, 10. [Google Scholar] [CrossRef] [Green Version]
- Tatsukawa, H.; Furutani, Y.; Hitomi, K.; Kojima, S. Transglutaminase 2 has opposing roles in the regulation of cellular functions as well as cell growth and death. Cell Death Dis. 2016, 7, e2244. [Google Scholar] [CrossRef] [Green Version]
- Malorni, W.; Farrace, M.G.; Matarrese, P.; Tinari, A.; Ciarlo, L.; Mousavi-Shafaei, P.; D’Eletto, M.; Di Giacomo, G.; Melino, G.; Palmieri, L.; et al. The adenine nucleotide translocator 1 acts as a type 2 transglutaminase substrate: Implications for mitochondrial-dependent apoptosis. Cell Death Differ. 2009, 16, 1480–1492. [Google Scholar] [CrossRef] [Green Version]
- Dorner, A.; Schultheiss, H.P. Adenine nucleotide translocase in the focus of cardiovascular diseases. Trends Cardiovasc. Med. 2007, 17, 284–290. [Google Scholar] [CrossRef]
- Tsujimoto, Y.; Shimizu, S. Role of the mitochondrial membrane permeability transition in cell death. Apoptos. Int. J. Program. Cell Death 2007, 12, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Murphy, L.J. Tissue transglutaminase has intrinsic kinase activity: Identification of transglutaminase 2 as an insulin-like growth factor-binding protein-3 kinase. J. Biol. Chem. 2004, 279, 23863–23868. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Melino, G.; Murphy, L.J. Transglutaminase 2 kinase activity facilitates protein kinase A-induced phosphorylation of retinoblastoma protein. J. Biol. Chem. 2007, 282, 18108–18115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, S.H.; Ji, S.H.; Han, E.T.; Park, W.S.; Hong, S.H.; Kim, Y.M.; Ha, K.S. Real-time monitoring of glucose-6-phosphate dehydrogenase activity using liquid droplet arrays and its application to human plasma samples. Biosens. Bioelectron. 2016, 79, 930–937. [Google Scholar] [CrossRef]
- Wang, Z.; Collighan, R.J.; Gross, S.R.; Danen, E.H.; Orend, G.; Telci, D.; Griffin, M. RGD-independent cell adhesion via a tissue transglutaminase-fibronectin matrix promotes fibronectin fibril deposition and requires syndecan-4/2 alpha5beta1 integrin co-signaling. J. Biol. Chem. 2010, 285, 40212–40229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, E.M.; Lee, K.B.; Kim, G.E.; Kim, C.M.; Lee, J.H.; Kim, H.J.; Shin, J.W.; Kwon, M.A.; Park, H.H.; Kim, I.G. Competitive binding of magnesium to calcium binding sites reciprocally regulates transamidase and GTP hydrolysis activity of transglutaminase 2. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [Green Version]
- Achyuthan, K.E.; Greenberg, C.S. Identification of a guanosine triphosphate-binding site on guinea pig liver transglutaminase. Role of GTP and calcium ions in modulating activity. J. Biol. Chem. 1987, 262, 1901–1906. [Google Scholar] [PubMed]
- Jang, T.H.; Lee, D.S.; Choi, K.; Jeong, E.M.; Kim, I.G.; Kim, Y.W.; Chun, J.N.; Jeon, J.H.; Park, H.H. Crystal structure of transglutaminase 2 with GTP complex and amino acid sequence evidence of evolution of GTP binding site. PLoS ONE 2014, 9, e107005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Venere, A.; Rossi, A.; De Matteis, F.; Rosato, N.; Agro, A.F.; Mei, G. Opposite effects of Ca(2+) and GTP binding on tissue transglutaminase tertiary structure. J. Biol. Chem. 2000, 275, 3915–3921. [Google Scholar] [CrossRef] [Green Version]
- Han, B.G.; Cho, J.W.; Cho, Y.D.; Jeong, K.C.; Kim, S.Y.; Lee, B.I. Crystal structure of human transglutaminase 2 in complex with adenosine triphosphate. Int. J. Biol. Macromol. 2010, 47, 190–195. [Google Scholar] [CrossRef]
- Karpuj, M.V.; Becher, M.W.; Springer, J.E.; Chabas, D.; Youssef, S.; Pedotti, R.; Mitchell, D.; Steinman, L. Prolonged survival and decreased abnormal movements in transgenic model of Huntington disease, with administration of the transglutaminase inhibitor cystamine. Nat. Med. 2002, 8, 143–149. [Google Scholar] [CrossRef]
- Lorand, L. DRPLA aggregation and transglutaminase, revisited. Nat. Genet. 1998, 20, 231. [Google Scholar] [CrossRef]
- Lorand, L.; Conrad, S.M. Transglutaminases. Mol. Cell. Biochem. 1984, 58, 9–35. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Stamnaes, J.; Klock, C.; DiRaimondo, T.R.; Sollid, L.M.; Khosla, C. Activation of extracellular transglutaminase 2 by thioredoxin. J. Biol. Chem. 2011, 286, 37866–37873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, E.M.; Kim, C.W.; Cho, S.Y.; Jang, G.Y.; Shin, D.M.; Jeon, J.H.; Kim, I.G. Degradation of transglutaminase 2 by calcium-mediated ubiquitination responding to high oxidative stress. FEBS Lett. 2009, 583, 648–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, D.M.; Jeon, J.H.; Kim, C.W.; Cho, S.Y.; Lee, H.J.; Jang, G.Y.; Jeong, E.M.; Lee, D.S.; Kang, J.H.; Melino, G.; et al. TGFbeta mediates activation of transglutaminase 2 in response to oxidative stress that leads to protein aggregation. FASEB J. 2008, 22, 2498–2507. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Cerione, R.A.; Clardy, J. Structural basis for the guanine nucleotide-binding activity of tissue transglutaminase and its regulation of transamidation activity. Proc. Natl. Acad. Sci. USA 2002, 99, 2743–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begg, G.E.; Holman, S.R.; Stokes, P.H.; Matthews, J.M.; Graham, R.M.; Iismaa, S.E. Mutation of a critical arginine in the GTP-binding site of transglutaminase 2 disinhibits intracellular cross-linking activity. J. Biol. Chem. 2006, 281, 12603–12609. [Google Scholar] [CrossRef] [Green Version]
- Ruan, Q.; Tucholski, J.; Gundemir, S.; Johnson Voll, G.V. The differential effects of R580A mutation on transamidation and GTP binding activity of rat and human type 2 transglutaminase. Int. J. Clin. Exp. Med. 2008, 1, 248–259. [Google Scholar]
- Pinkas, D.M.; Strop, P.; Brunger, A.T.; Khosla, C. Transglutaminase 2 undergoes a large conformational change upon activation. PLoS Biol. 2007, 5, e327. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Zhuang, R.; Khosla, C. Substrates, inhibitors, and probes of mammalian transglutaminase 2. Anal. Biochem. 2020, 591, 113560. [Google Scholar] [CrossRef]
- Ventura, M.A.E.; Sajko, K.; Hils, M.; Pasternack, R.; Greinwald, R.; Tewes, B.; Schuppan, D. The oral transglutaminase 2 (Tg2) inhibitor Zed1227 blocks Tg2 activity in a mouse model of intestinal inflammation. Gastroenterology 2018, 154, S490. [Google Scholar] [CrossRef]
- Duval, E.; Case, A.; Stein, R.L.; Cuny, G.D. Structure-activity relationship study of novel tissue transglutaminase inhibitors. Bioorg. Med. Chem. Lett. 2005, 15, 1885–1889. [Google Scholar] [CrossRef] [PubMed]
- Martucciello, S.; Paolella, G.; Esposito, C.; Lepretti, M.; Caputo, I. Anti-type 2 transglutaminase antibodies as modulators of type 2 transglutaminase functions: A possible pathological role in celiac disease. Cell. Mol. Life Sci. 2018, 75, 4107–4124. [Google Scholar] [CrossRef] [PubMed]
- Paolella, G.; Lepretti, M.; Barone, M.V.; Nanayakkara, M.; Di Zenzo, M.; Sblattero, D.; Auricchio, S.; Esposito, C.; Caputo, I. Celiac anti-type 2 transglutaminase antibodies induce differential effects in fibroblasts from celiac disease patients and from healthy subjects. Amino Acids 2017, 49, 541–550. [Google Scholar] [CrossRef] [PubMed]
- du Pre, M.F.; Blazevski, J.; Dewan, A.E.; Stamnaes, J.; Kanduri, C.; Sandve, G.K.; Johannesen, M.K.; Lindstad, C.B.; Hnida, K.; Fugger, L.; et al. B cell tolerance and antibody production to the celiac disease autoantigen transglutaminase 2. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Hnida, K.; Graewert, M.A.; Andersen, J.T.; Iversen, R.; Tuukkanen, A.; Svergun, D.; Sollid, L.M. Structural basis for antigen recognition by transglutaminase 2-specific autoantibodies in celiac disease. J. Biol. Chem. 2015, 290, 21365–21375. [Google Scholar] [CrossRef] [Green Version]
- Simon-Vecsei, Z.; Kiraly, R.; Bagossi, P.; Toth, B.; Dahlbom, I.; Caja, S.; Csosz, E.; Lindfors, K.; Sblattero, D.; Nemes, E.; et al. A single conformational transglutaminase 2 epitope contributed by three domains is critical for celiac antibody binding and effects. Proc. Natl. Acad. Sci. USA 2012, 109, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.; Guha, S.; Diagaradjane, P.; Kunnumakkara, A.B.; Sanguino, A.M.; Lopez-Berestein, G.; Sood, A.K.; Aggarwal, B.B.; Krishnan, S.; Gelovani, J.G.; et al. Therapeutic significance of elevated tissue transglutaminase expression in pancreatic cancer. Clin. Cancer Res. 2008, 14, 2476–2483. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Reaction mechanism of transglutaminase 2 (TGase2). (A) Transamidation reaction mechanism of TGase2. Residues in green are from TGase2. Two consecutive nucleophilic attacks by the thiol group from the active site on TGase2 and the amine donor are shown. (B) Figures of the closed inactive form and the open active form of TGase2 showing the catalytic triad composed of C277, H335, and D358. The four distinct domains, β-sandwich domain, catalytic core domain, β-barrel1, and β-barrel2, are visually distinguished by the different colors.
Figure 1.
Reaction mechanism of transglutaminase 2 (TGase2). (A) Transamidation reaction mechanism of TGase2. Residues in green are from TGase2. Two consecutive nucleophilic attacks by the thiol group from the active site on TGase2 and the amine donor are shown. (B) Figures of the closed inactive form and the open active form of TGase2 showing the catalytic triad composed of C277, H335, and D358. The four distinct domains, β-sandwich domain, catalytic core domain, β-barrel1, and β-barrel2, are visually distinguished by the different colors.

Figure 2.
Structural analysis of the nucleotide-binding site on TGase2. (A) Structural comparison of TGase2 complexed with different nucleotides. Three different structures were superimposed and compared. The dotted red circle indicates the nucleotide-binding site located in between the catalytic core domain and the β-barrel1 domain. (B) Close-up view of the nucleotide-binding site on TGase2. Residues involved in the interaction with the nucleotides are shown. (C) Sequence comparison of TGase2 in various species. Residues important for binding with nucleotides are shown in red bold font.
Figure 2.
Structural analysis of the nucleotide-binding site on TGase2. (A) Structural comparison of TGase2 complexed with different nucleotides. Three different structures were superimposed and compared. The dotted red circle indicates the nucleotide-binding site located in between the catalytic core domain and the β-barrel1 domain. (B) Close-up view of the nucleotide-binding site on TGase2. Residues involved in the interaction with the nucleotides are shown. (C) Sequence comparison of TGase2 in various species. Residues important for binding with nucleotides are shown in red bold font.

Figure 3.
Structural analysis of TGase2 complexed with peptide mimetic inhibitors. Structure of TGase2 complexed with peptide mimetic, Ac-P(DON)LPF-NH2 (A), with peptide inhibitor1, N-[(benzyloxy)carbonyl]-6-diazonio-5-oxo-L-norleucyl-L-valyl-L-prolyl-L-leucine (B), with peptide inhibitor2, N~2~-[(2S)-2-{[(benzyloxy)carbonyl]amino}-7-ethoxy-7-oxoheptanoyl]-L-glutaminyl-L-prolyl-L-leucine (C), and with peptide inhibitor3, N-[(2R)-2-{[(benzyloxy)carbonyl]amino}-7-ethoxy-7-oxoheptanoyl]-L-valyl-L-prolyl-L-leucine (D). The red box indicates the inhibitor-binding site. The magnified figure of the inhibitor in the inhibitor-binding site of TGase2 is provided. The binding details, generated by LIGPLOT+ [80], are shown at the right side of each panel. The black and cyan residues indicate the hydrophobic interaction and hydrogen bond, respectively.
Figure 3.
Structural analysis of TGase2 complexed with peptide mimetic inhibitors. Structure of TGase2 complexed with peptide mimetic, Ac-P(DON)LPF-NH2 (A), with peptide inhibitor1, N-[(benzyloxy)carbonyl]-6-diazonio-5-oxo-L-norleucyl-L-valyl-L-prolyl-L-leucine (B), with peptide inhibitor2, N~2~-[(2S)-2-{[(benzyloxy)carbonyl]amino}-7-ethoxy-7-oxoheptanoyl]-L-glutaminyl-L-prolyl-L-leucine (C), and with peptide inhibitor3, N-[(2R)-2-{[(benzyloxy)carbonyl]amino}-7-ethoxy-7-oxoheptanoyl]-L-valyl-L-prolyl-L-leucine (D). The red box indicates the inhibitor-binding site. The magnified figure of the inhibitor in the inhibitor-binding site of TGase2 is provided. The binding details, generated by LIGPLOT+ [80], are shown at the right side of each panel. The black and cyan residues indicate the hydrophobic interaction and hydrogen bond, respectively.

Figure 4.
Structural analysis of celiac autoantibody-binding sites, site 1 (A) and site 2 (B). Close-up view of the autoantibody-binding sites (right panel). Residues that are involved in the interaction with the autoantibody are shown.
Figure 4.
Structural analysis of celiac autoantibody-binding sites, site 1 (A) and site 2 (B). Close-up view of the autoantibody-binding sites (right panel). Residues that are involved in the interaction with the autoantibody are shown.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Structures of TGase2 complexed with effectors and inhibitors.
| Effectors/Inhibitors | Closed/Open | PDB | References |
|---|---|---|---|
| GDP | Closed | 1KV3 | [76] |
| GTP | Closed | 4PYG | [67] |
| ATP | Closed | 3LY6 | [69] |
| Ac-P(DON)LPF-NH2 | Open | 2Q3Z | [79] |
| Inhibitor1 | Open | 3S3J | N.A. |
| Inhibitor2 | Open | 3S3P | N.A. |
| Inhibitor3 | Open | 3S3S | N.A. |
PDB, The Protein Data Bank; GDP, guanosine diphosphate; GTP, guanosine triphosphate; N.A., Non-Available.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, G.E.; Park, H.H. Structures of Human Transglutaminase 2: Finding Clues for Interference in Cross-linking Mediated Activity. Int. J. Mol. Sci. 2020, 21, 2225. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062225
AMA Style
Kim GE, Park HH. Structures of Human Transglutaminase 2: Finding Clues for Interference in Cross-linking Mediated Activity. International Journal of Molecular Sciences. 2020; 21(6):2225. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062225
Chicago/Turabian StyleKim, Gi Eob, and Hyun Ho Park. 2020. "Structures of Human Transglutaminase 2: Finding Clues for Interference in Cross-linking Mediated Activity" International Journal of Molecular Sciences 21, no. 6: 2225. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062225
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.