Characterization, Expression Pattern and Antiviral Activities of Mx Gene in Chinese Giant Salamander, Andrias davidianus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
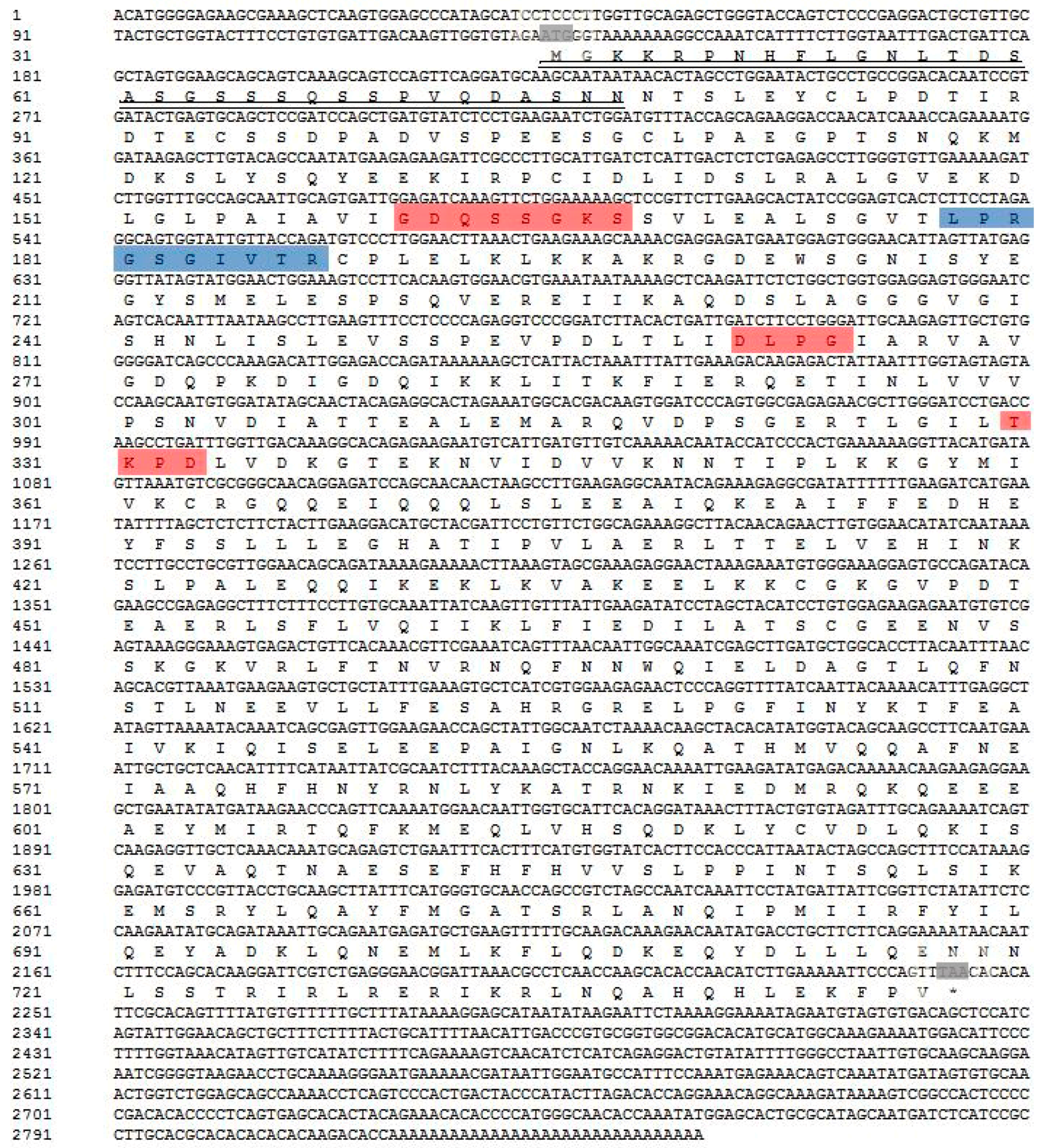
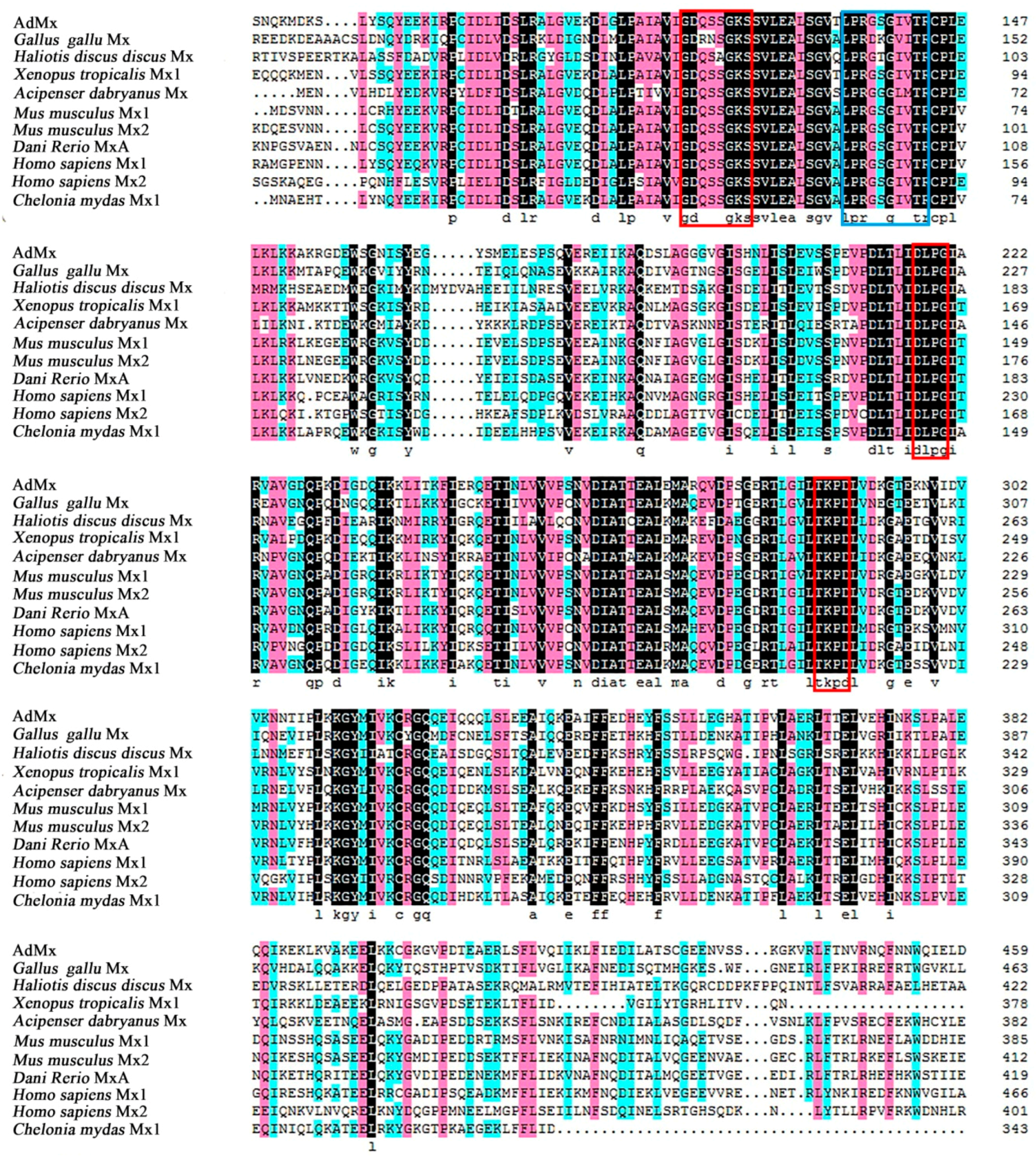
2.1. Identification and Molecular Characterization of AdMx
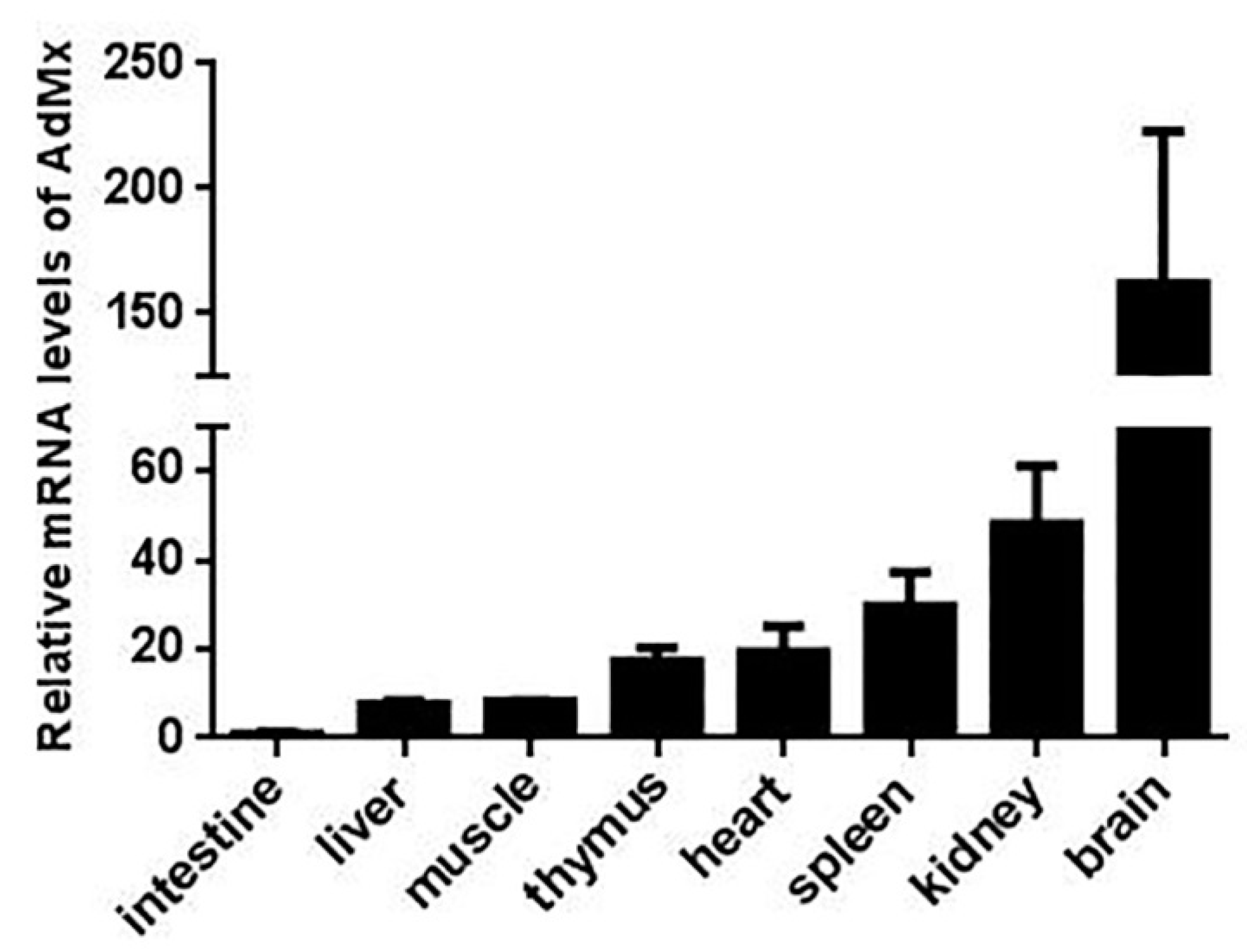
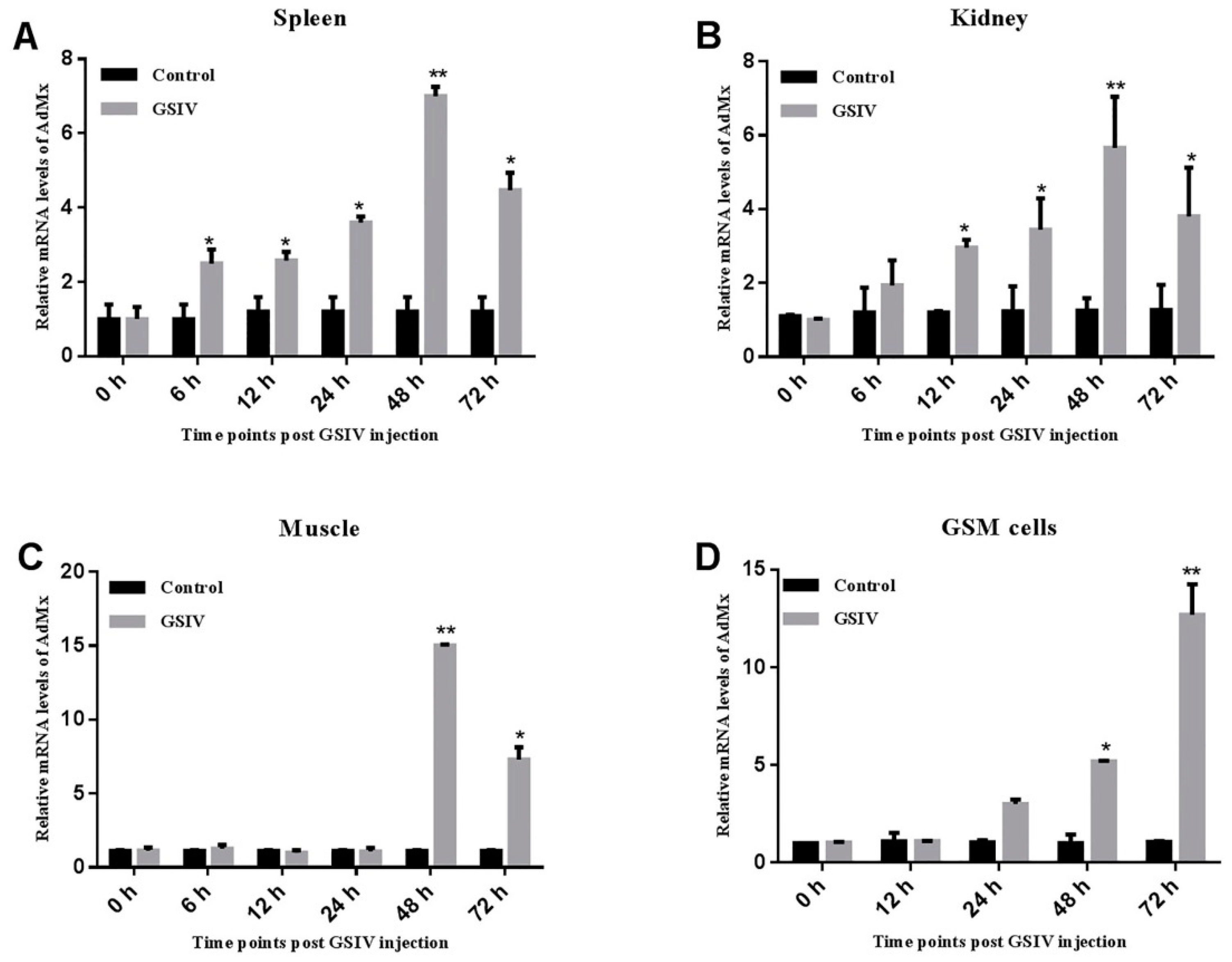
2.2. Expression of AdMx in Chinese Giant Salamander Tissues and GSM Cells
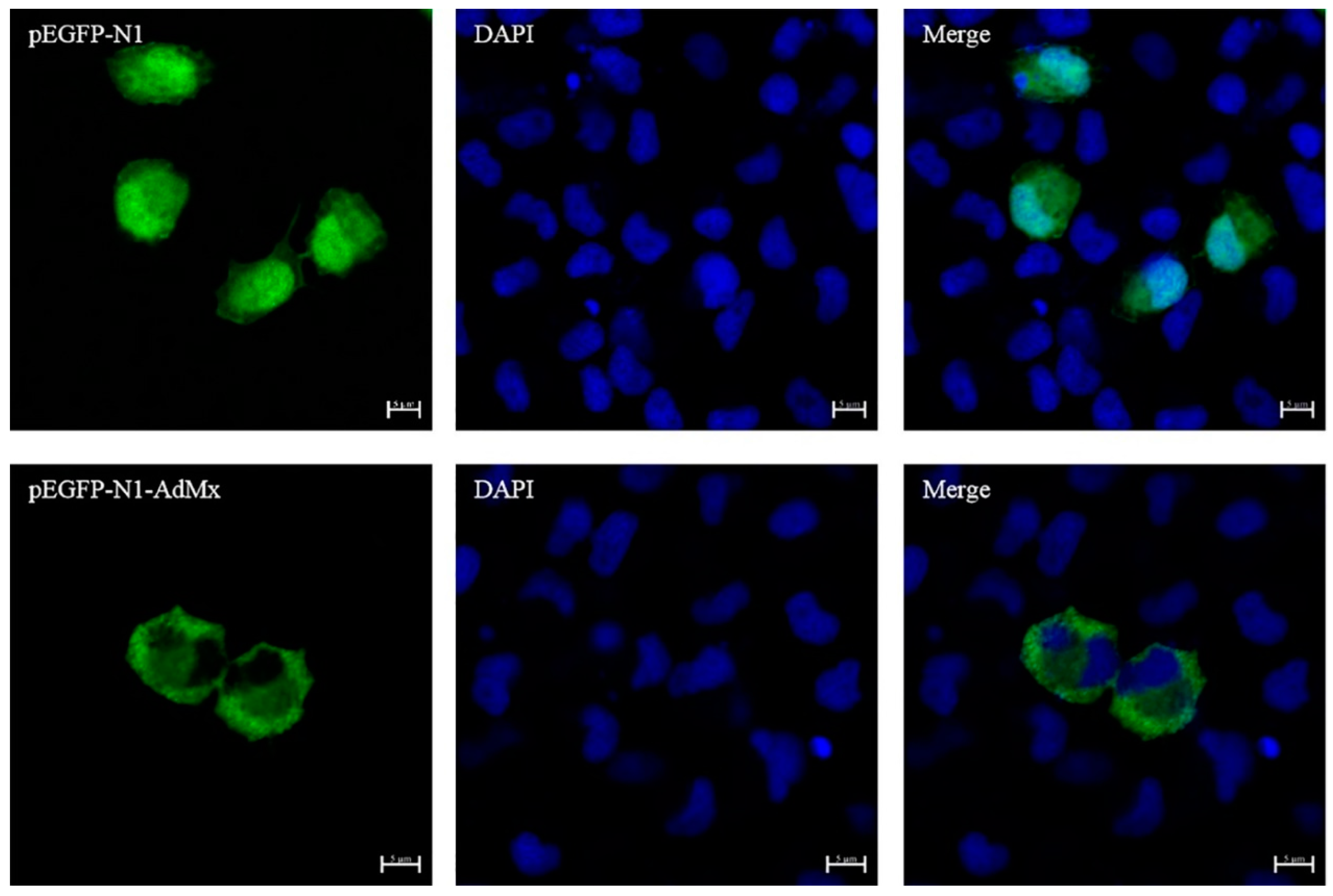
2.3. Evaluation of AdMx Subcellular Localization
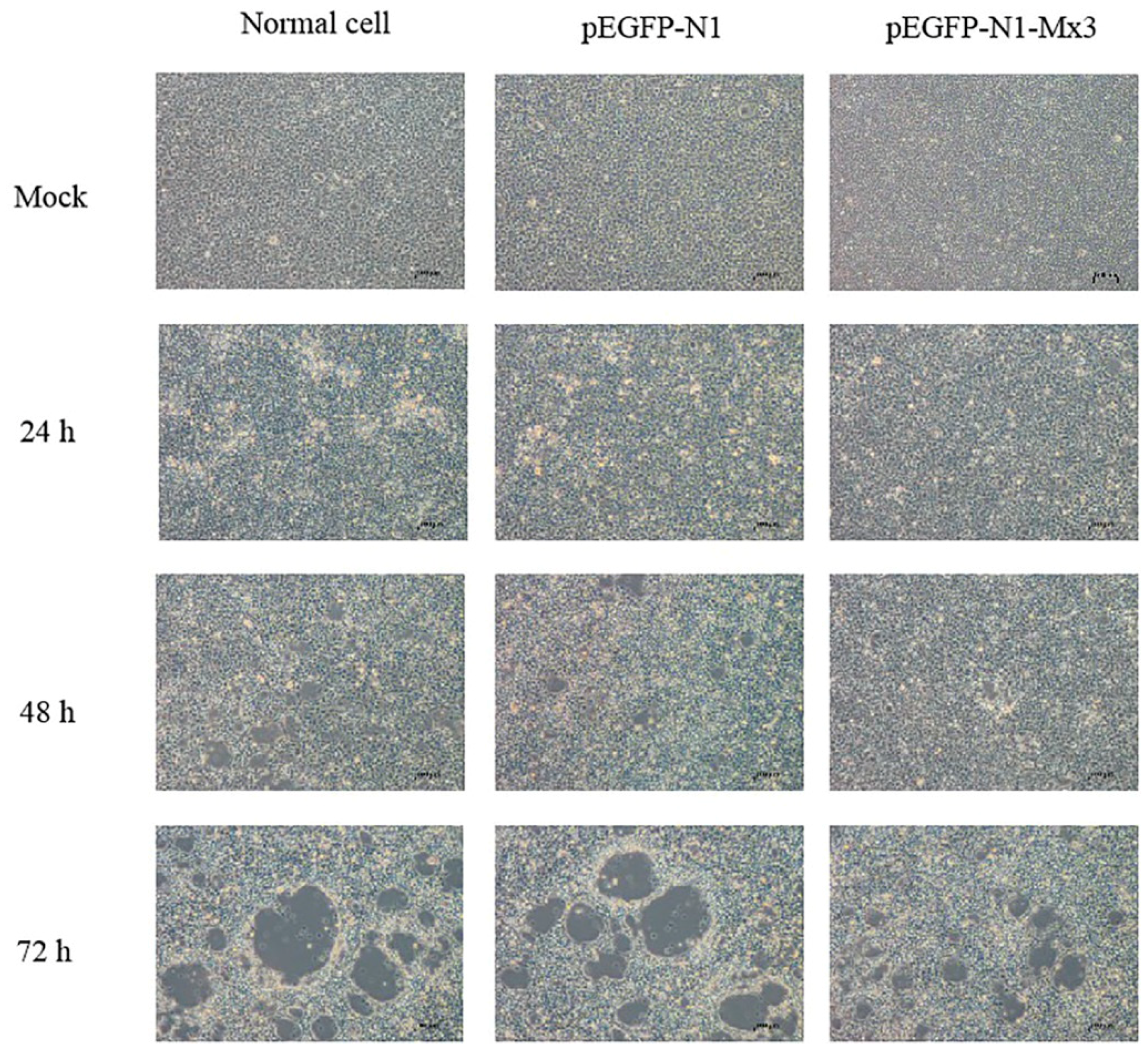
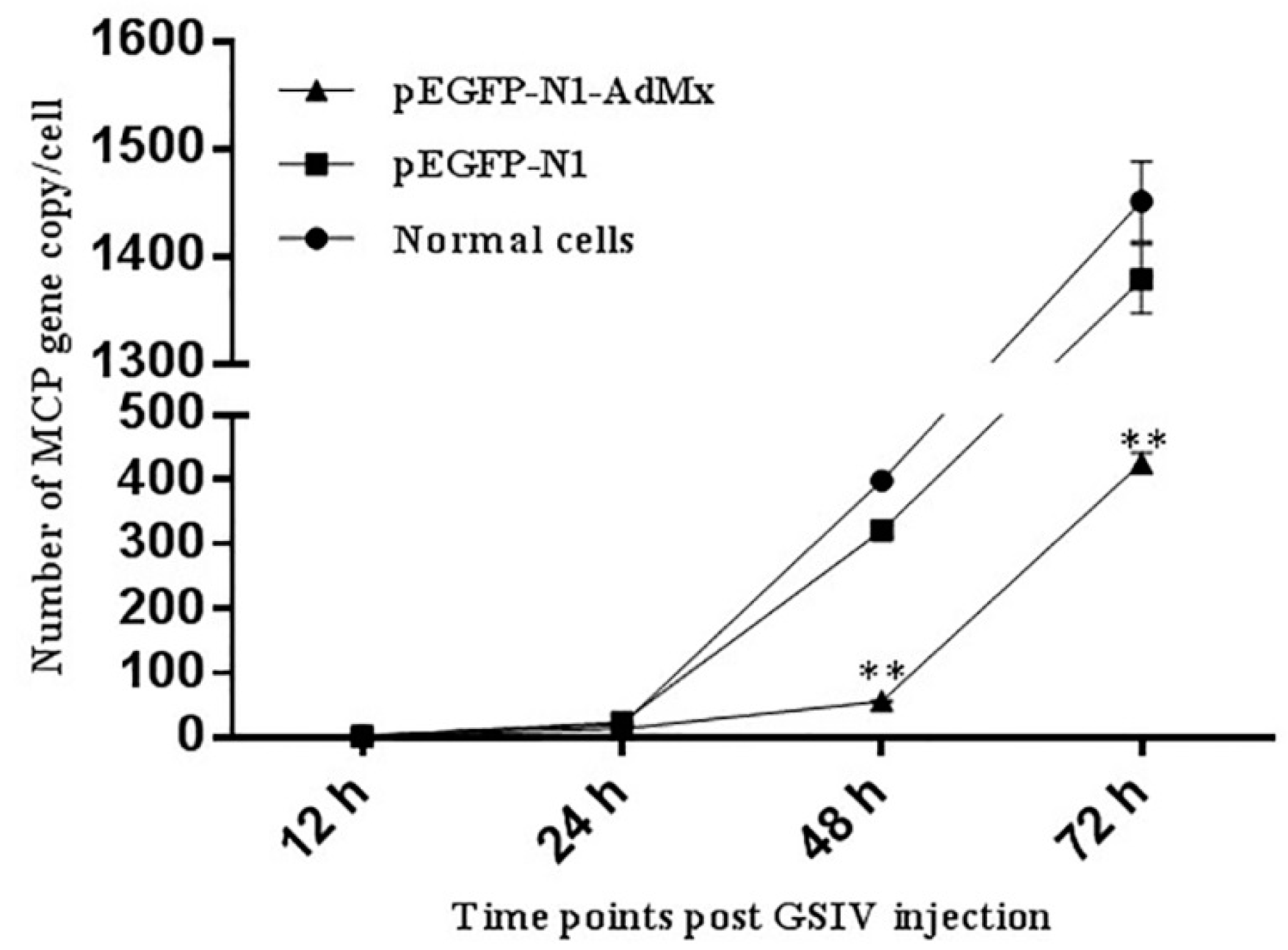
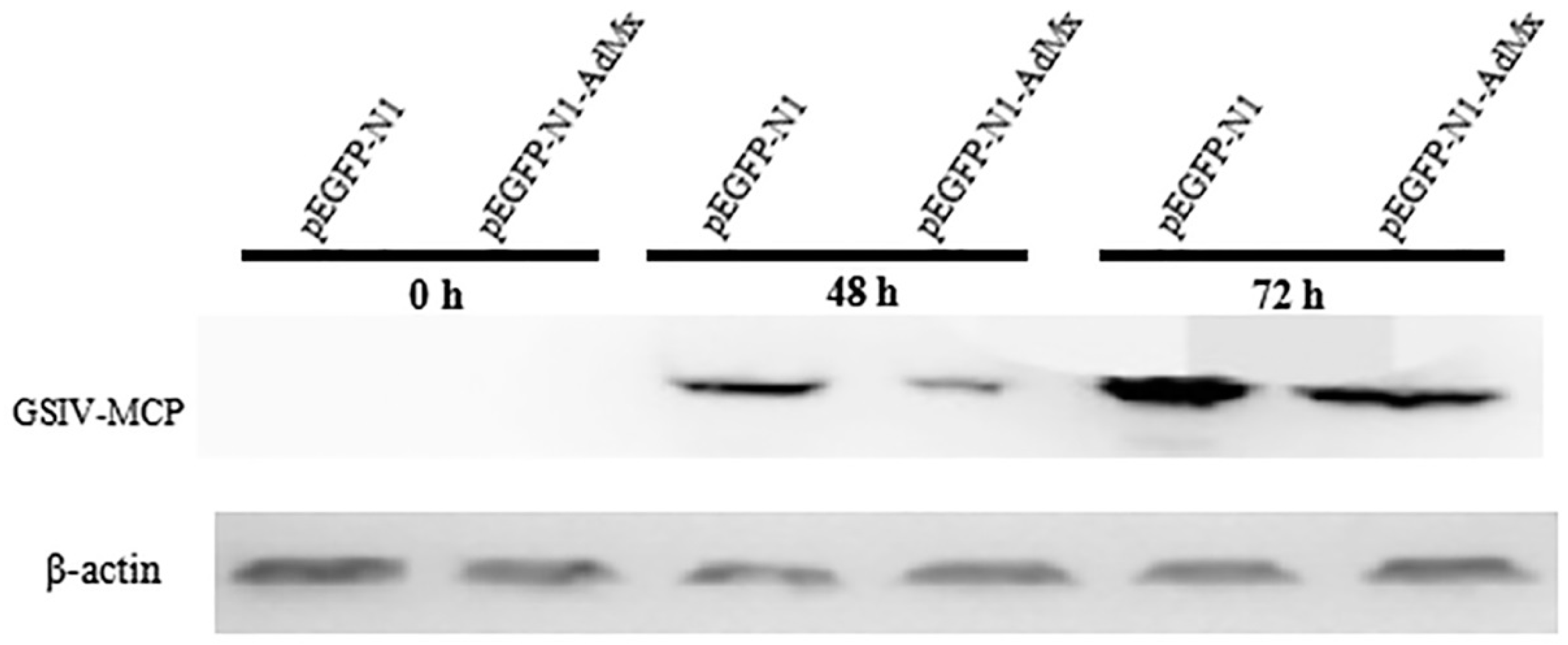
2.4. Antiviral effect of AdMx in vitro
3. Discussion
4. Materials and Methods
4.1. Animals, Cells, and Virus
4.2. Cloning of AdMx
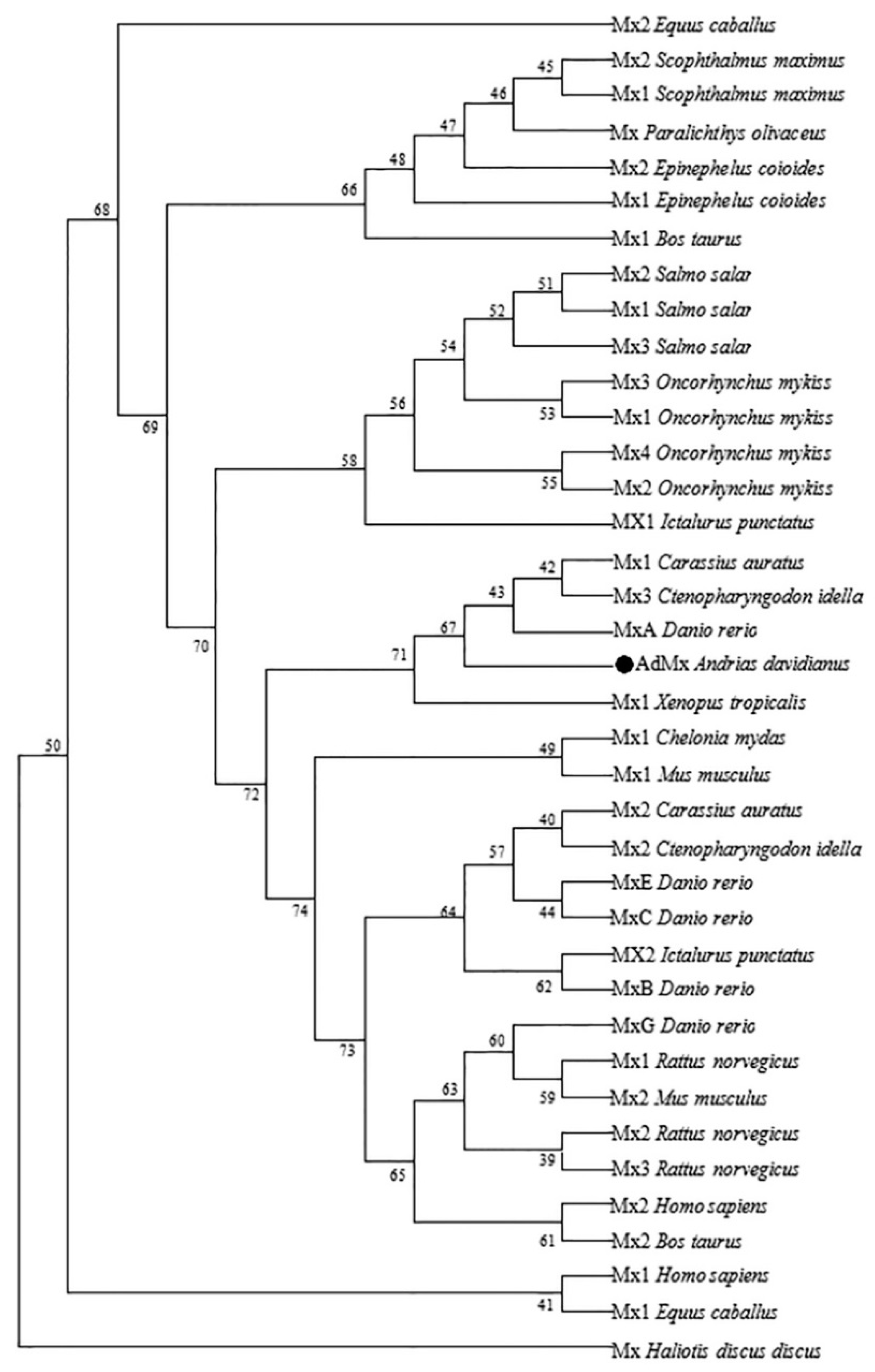
4.3. Sequence Alignment and Phylogenetic Analysis
4.4. Detection of AdMx Expression by Quantitative reAl-Time PCR (qRT-PCR)
4.5. Construction of AdMx Plasmid and Transfection
4.6. AdMx Subcellular Localization
4.7. Detection of GSIV Major Capsid Protein (MCP) Gene Copies by Droplet Digital PCR (ddPCR)
4.8. Western Blot
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| GSIV | Chinese giant salamander iridovirus |
| GSM | Chinese giant salamander muscle cell line |
| MOI | Multiplicity of infection |
References
- Jacobs, B.L.; Langland, J.O. When two strands are better than one: The mediators and modulators of the cellular responses to double-stranded RNA. Virology 1996, 219, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Haller, O.; Staeheli, P.; Kochs, G. Interferon-induced Mx proteins in antiviral host defense. Biochimie 2007, 89, 812–818. [Google Scholar] [CrossRef]
- Kochs, G.; Reichelt, M.; Danino, D.; Hinshaw, J.E.; Haller, O. Assay and Functional Analysis of Dynamin-Like Mx Proteins. Methods Enzymol. 2005, 404, 632–643. [Google Scholar]
- Lindenmann, J. Resistance of mice to mouse-adapted influenza A virus. Virology 1962, 16, 203–204. [Google Scholar] [CrossRef]
- Robertsen, B.; Trobridge, G.; Leongt, J.A. Molecular cloning of double-stranded RNA inducible Mx genes from Atlantic salmon (Salmo salar L.). Dev. Comp. Immunol. 1997, 21, 397–412. [Google Scholar] [CrossRef]
- Lee, J.Y.; Hirono, I.; Aoki, T. Cloning and analysis of expression of Mx cDNA in Japanese flounder, Paralichthys olivaceus. Dev. Comp. Immunol. 2000, 24, 407–415. [Google Scholar] [CrossRef]
- Yap, W.H.; Tay, A.; Brenner, S.; Venkatesh, B. Molecular cloning of the pufferfish (Takifugu rubripes) Mx gene and functional characterization of its promoter. Immunogenetics 2003, 54, 705–713. [Google Scholar] [CrossRef]
- Plant, K.P.; Thune, R.L. Cloning and characterisation of a channel catfish (Ictalurus punctatus) Mx gene. Fish. Shellfish Immunol. 2004, 16, 0–405. [Google Scholar] [CrossRef]
- Zoysa, M.D.; Kang, H.-S.; Song, Y.-B.; Jee, Y.; Lee, Y.-D.; Lee, J. First report of invertebrate Mx: Cloning, characterization and expression analysis of Mx cDNA in disk abalone (Haliotis discus discus). Fish. Shellfish Immunol. 2007, 23, 0–96. [Google Scholar] [CrossRef]
- Mittelman, M.; Gardyn, J.; Carmel, M.; Malovani, H.; Barak, Y.; Nir, U. Analysis of the erythropoietin receptor gene in patients with myeloproliferative and myelodysplastic syndromes. Leuk Res. 1996, 20, 459–466. [Google Scholar] [CrossRef]
- Meier, E.; Kunz, G.; Haller, O.; Arnheiter, H.A. Activity of rat Mx proteins against a rhabdovirus. J. Virol. 1991, 64, 6263–6269. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.K.; Yoshimatsu, K.; Takada, A.; Ogino, M.; Asano, A.; Arikawa, J.; Watanabe, T. Mouse Mx2 protein inhibits hantavirus but not influenza virus replication. Arch. Virol. 2001, 146, 41–49. [Google Scholar] [CrossRef]
- González-Mariscal, J.A.; Fernández-Trujillo, M.A.; Alonso, M.C.; García-Rosado, E.; Álvarez, M.C.; Béjar, J. Gilthead seabream (Sparus aurata) Mx gene promoters respond differentially to IPNV and VHSV infections in RTG-2 cells. Vet. Immunol. Immunopathol. 2016, 171, 73–80. [Google Scholar] [CrossRef]
- Grayfer, L.; De Jesus Andino, F.; Robert, J. The Amphibian (Xenopus laevis) Type I Interferon Response to Frog Virus 3: New Insight into Ranavirus Pathogenicity. J. Virol. 2014, 88, 5766–5777. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Ma, J.; Fan, Y.; Meng, Y.; Xu, J.; Zhou, Y.; Liu, W.; Zeng, X.; Zeng, L. Identification of Type I IFN in Chinese giant salamander (Andrias davidianus) and the response to an iridovirus infection. Mol. Immunol. 2015, 65, 350–359. [Google Scholar] [CrossRef]
- Meng, Y.; Ma, J.; Jiang, N.; Zeng, L.-B.; Xiao, H.-B. Pathological and microbiological findings from mortality of the Chinese giant salamander (Andrias davidianus). Arch. Virol. 2014, 159, 1403–1412. [Google Scholar] [CrossRef]
- Kochs, G.; Haller, O. GTP-bound human MxA protein interacts with the nucleocapsids of Thogoto virus (Orthomyxoviridae). J. Biol. Chem. 1999, 274, 4370–4376. [Google Scholar] [CrossRef] [Green Version]
- Melén, K.; Julkunen, I. Mutational analysis of murine Mx1 protein: GTP binding core domain is essential for anti-influenza A activity. Virology 1994, 205, 269–279. [Google Scholar] [CrossRef]
- Asano, A.; KO, J.H.; Morozumi, T.; Hamashima, N.; Watanabe, T. Polymorphisms and the Antiviral Property of Porcine Mx1 Protein. J. Vet. Med. Sci. 2002, 64, 1085–1089. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.H.; Jin, H.K.; Asano, A.; Takada, A.; Ninomiya, A.; Kida, H.; Hokiyama, H.; Ohara, M.; Tsuzuki, M.; Nishibori, M. Polymorphisms and the differential antiviral activity of the chicken Mx gene. Genome Res. 2002, 14, 595–601. [Google Scholar] [CrossRef] [Green Version]
- Caipang, C.M.A.; Hirono, I.; Aoki, T. In vitro inhibition of fish rhabdoviruses by Japanese flounder, Paralichthys olivaceus Mx. Virology 2003, 317, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Jensen, V.; Robertsen, B.R. Cloning of an Mx cDNA from Atlantic Halibut (Hippoglossus hippoglossus) and Characterization of Mx mRNA Expression in Response to Double-Stranded RNA or Infectious Pancreatic Necrosis Virus. J. Interferon Cytokine Res. 2000, 20, 701–710. [Google Scholar] [CrossRef]
- Zhi-guang, X.; Xiao-min, L.; Li, W.; Wei, L.; Wei, W.; Wei-jie, H.; Jian-chao, W.; Qing-wen, M. Cloning and Tissue Expression of Mx1 Gene in ICR Mouse. Prog. Vet. Med. 2011, 32, 23–28. [Google Scholar]
- Peng, L.; Chunrong, Y.; Jianguo, S.; Zhanjiang, L. Protective Roles of Grass Carp Ctenopharyngodon idella Mx Isoforms against Grass Carp Reovirus. PLoS ONE 2012, 7, e52142. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Trujillo, M.A.; García-Rosado, E.; Alonso, M.C.; Castro, D.; Álvarez, M.C.; Béjar, J. Mx1, Mx2 and Mx3 proteins from the gilthead seabream (Sparus aurata) show in vitro antiviral activity against RNA and DNA viruses. Mol. Immunol. 2013, 56, 630–636. [Google Scholar] [CrossRef]
- Roy, P.; Panda, S.P.; Pal, A.; Mishra, S.S.; Jayasankar, P.; Das, B.K. Expression of Mx Gene in Cirrhinus mrigala (Hamilton, 1822) to OmpC Protein of Aeromonas hydrophila and Bacterial Infection. Appl. Biochem. Biotechnol. 2016, 178, 640–653. [Google Scholar] [CrossRef]
- Zhang, S.H.; Lv, X.Y.; Deng, D.; Zeng, L.; Huang, W.; Tang, D.D.; Zhang, L.F.; Xu, Q.Q.; Wei, Q.W. Gene characterization and expression pattern of Mx and Viperin genes in Dabry’s sturgeon Acipenser dabryanus. J. Appl. Ichthyol. 2019, 35, 408–419. [Google Scholar] [CrossRef]
- Zhu, Q.; Li, C.; Yu, Z.-X.; Zou, P.-F.; Meng, Q.-X.; Yao, C.-L. Molecular and immune response characterizations of IL-6 in large yellow croaker (Larimichthys crocea). Fish. Shellfish Immunol. 2016, 53, 78. [Google Scholar] [CrossRef]
- Bravo, J.; Real, F.; Padilla, D.; Olveira, J.G.; Grasso, V.; Román, L.; Acosta, F. Effect of lipopolysaccharides from Vibrio alginolyticus on the Mx gene expression and virus recovery from gilthead sea bream (Sparus aurata L.) experimentally infected with Nodavirus. Fish. Shellfish Immunol. 2013, 34, 383–386. [Google Scholar] [CrossRef]
- Van Wijngaarden, S.E.; Said-Bouyeri, S.B.; Ninaber, M.K.; Huizinga, T.W.; Schalij, M.J.; Bax, J.J.; Delgado, V.; de Vries-Bouwstra, J.K.; Marsan, N.A. Progression of Left Ventricular Myocardial Dysfunction in Systemic Sclerosis: A Speckle-tracking Strain Echocardiography Study. J. Rheumatol. 2019, 46, 405–415. [Google Scholar] [CrossRef]
- Lee, S.H.; Vidal, S.M. Functional diversity of Mx proteins: Variations on a theme of host resistance to infection. Genome Res. 2002, 12, 527–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zürcher, T.; Pavlovic, J.; Staeheli, P. Mouse mx2 protein inhibits vesicular stomatitis vreus. Virology 1992. [Google Scholar] [CrossRef]
- Kibenge, M.J.; Munir, K.; Kibenge, F.S. Constitutive expression of Atlantic salmon Mx1 protein in CHSE-214 cells confers resistance to Infectious Salmon Anaemia virus. Virol. J. 2005, 2, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, R.; Rokenes, T.P.; Robertsen, B. Inhibition of Infectious Pancreatic Necrosis Virus Replication by Atlantic Salmon Mx1 Protein. J. Virol. 2004, 78, 7938–7944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haller, O.; Kochs, G. Human MxA Protein: An Interferon-Induced Dynamin-Like GTPase with Broad Antiviral Activity. J. Interferon Cytokine Res. 2011, 31, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Yang, C.; Zhu, Z.; Wang, Y.; Jang, S.; Liao, L. Enhanced grass carp reovirus resistance of Mx-transgenic rare minnow (Gobiocypris rarus). Fish. Shellfish Immunol. 2009, 26, 828–835. [Google Scholar] [CrossRef]
- Wu, Y.C.; Chi, S.C. Cloning and analysis of antiviral activity of a barramundi (Lates calcarifer) Mx gene. Immunology 2007, 23, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.-C.; Lu, Y.-F.; Chi, S.-C. Anti-viral mechanism of barramundi Mx against betanodavirus involves the inhibition of viral RNA synthesis through the interference of RdRp. Fish. Shellfish Immunol. 2010, 28, 467–475. [Google Scholar] [CrossRef]
- Chen, Y.-M.; Su, Y.-L.; Shie, P.-S.; Huang, S.-L.; Yang, H.-L.; Chen, T.-Y. Grouper Mx confers resistance to nodavirus and interacts with coat protein. Dev. Comp. Immunol. 2008, 32, 825–836. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2000, 25, 402–408. [Google Scholar] [CrossRef]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Li, Y.; Zhou, Y.; Jiang, N.; Fan, Y.; Zeng, L. Characterization, Expression Pattern and Antiviral Activities of Mx Gene in Chinese Giant Salamander, Andrias davidianus. Int. J. Mol. Sci. 2020, 21, 2246. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062246
Liu Y, Li Y, Zhou Y, Jiang N, Fan Y, Zeng L. Characterization, Expression Pattern and Antiviral Activities of Mx Gene in Chinese Giant Salamander, Andrias davidianus. International Journal of Molecular Sciences. 2020; 21(6):2246. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062246
Chicago/Turabian StyleLiu, Yanan, Yiqun Li, Yongze Zhou, Nan Jiang, Yuding Fan, and Lingbing Zeng. 2020. "Characterization, Expression Pattern and Antiviral Activities of Mx Gene in Chinese Giant Salamander, Andrias davidianus" International Journal of Molecular Sciences 21, no. 6: 2246. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062246