A Multiparametric Study of Internalization of Fullerenol C60(OH)36 Nanoparticles into Peripheral Blood Mononuclear Cells: Cytotoxicity in Oxidative Stress Induced by Ionizing Radiation


Abstract
:
1. Introduction
2. Results
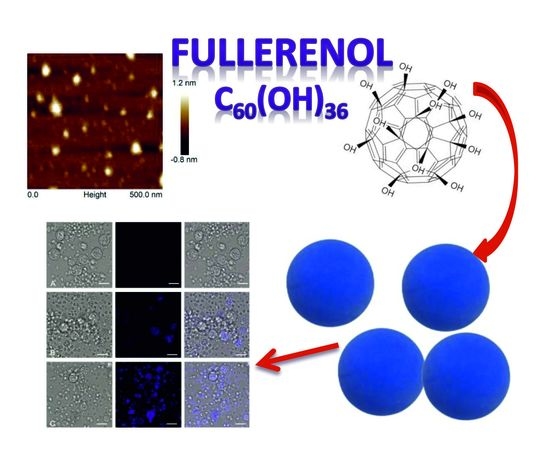
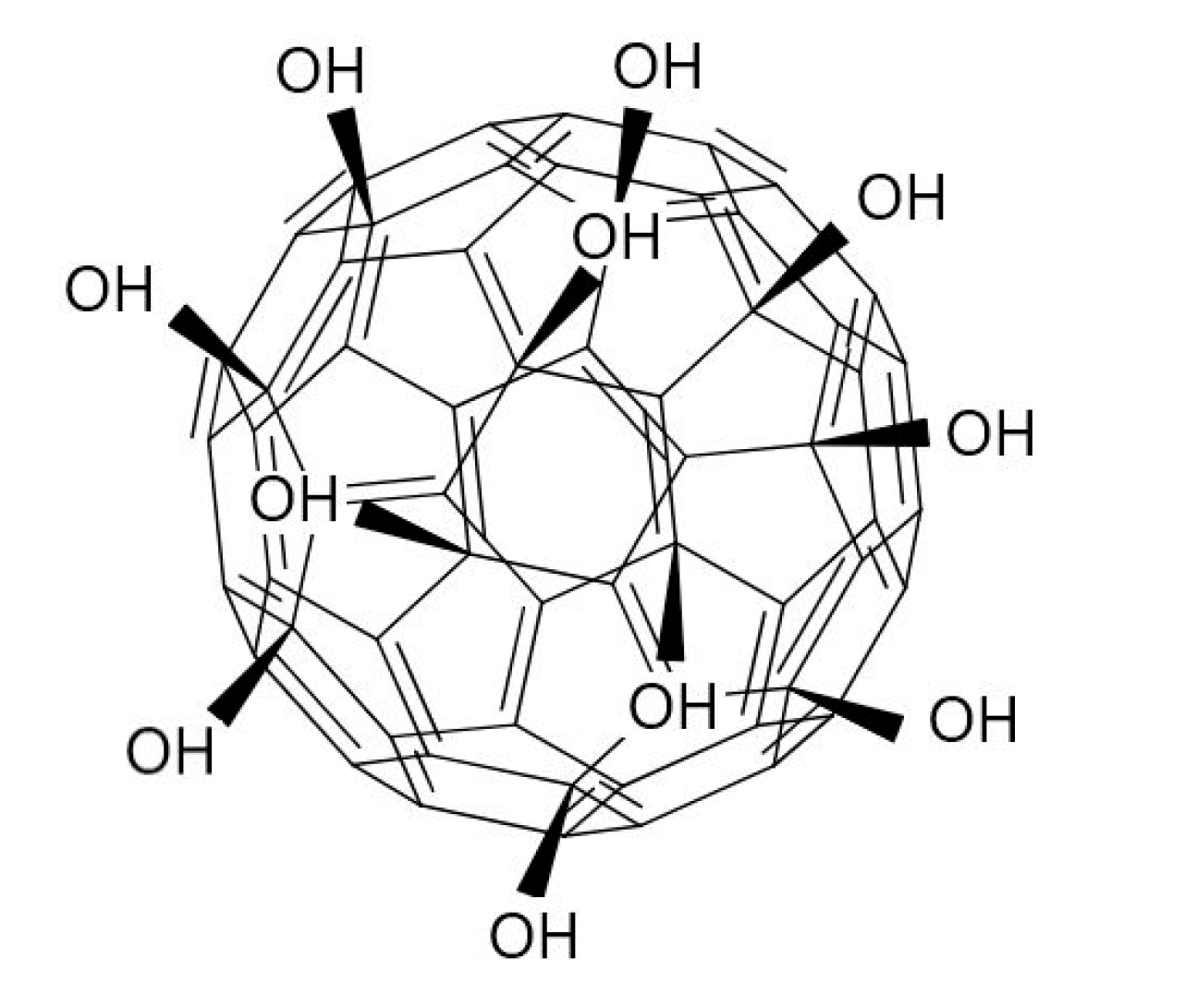
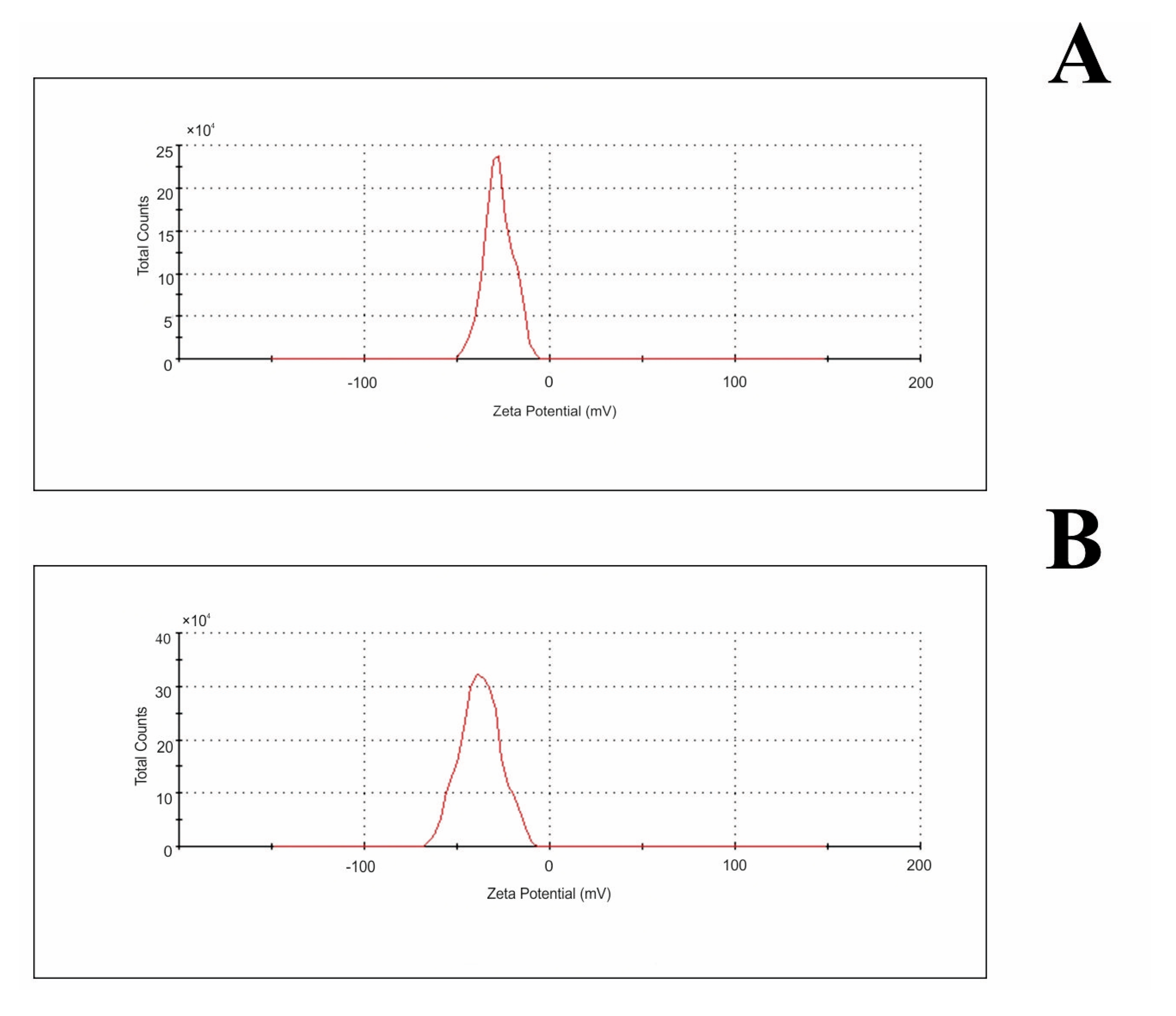
2.1. Characterization of Fullerenol Nanoparticles
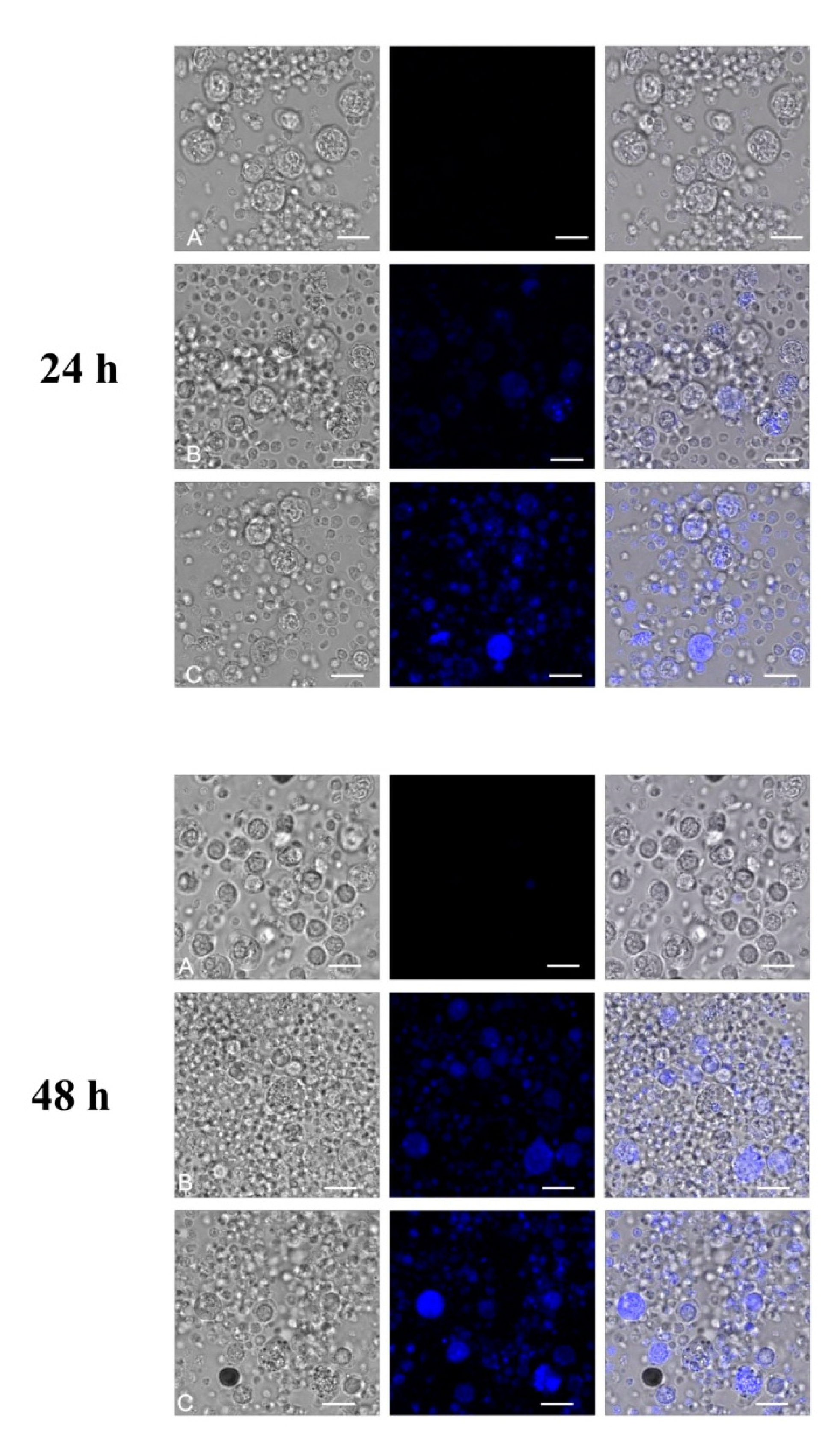
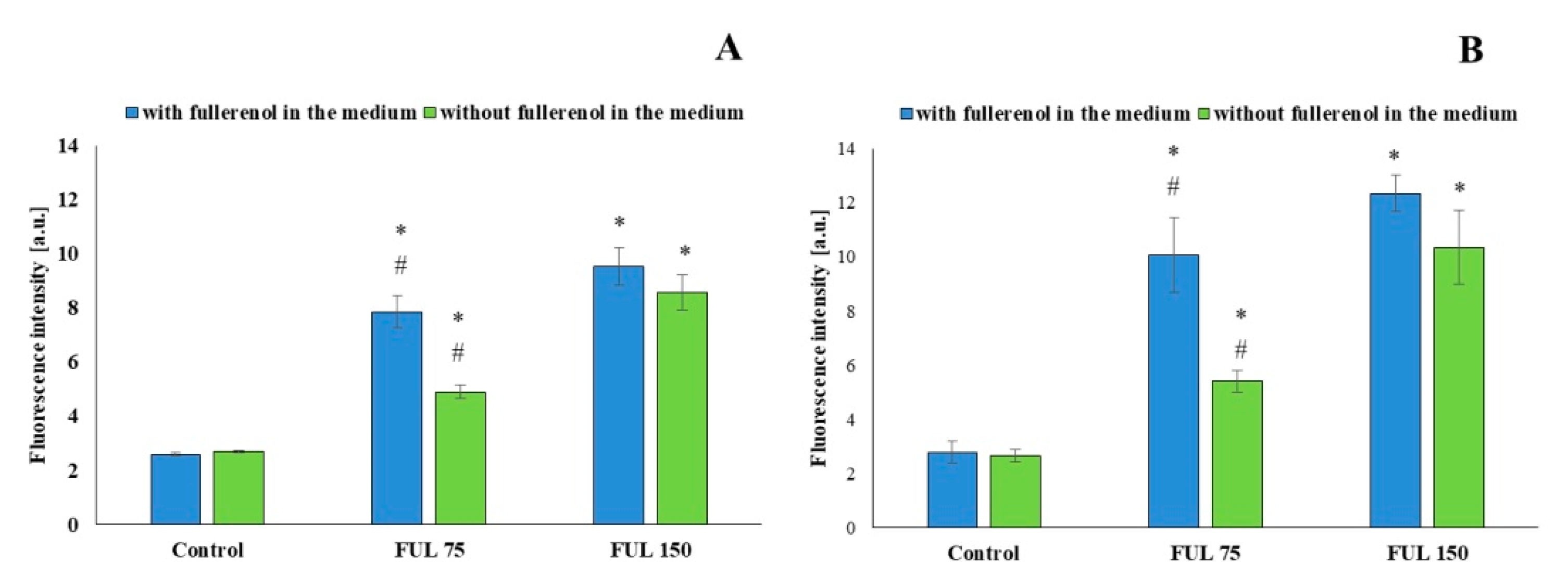
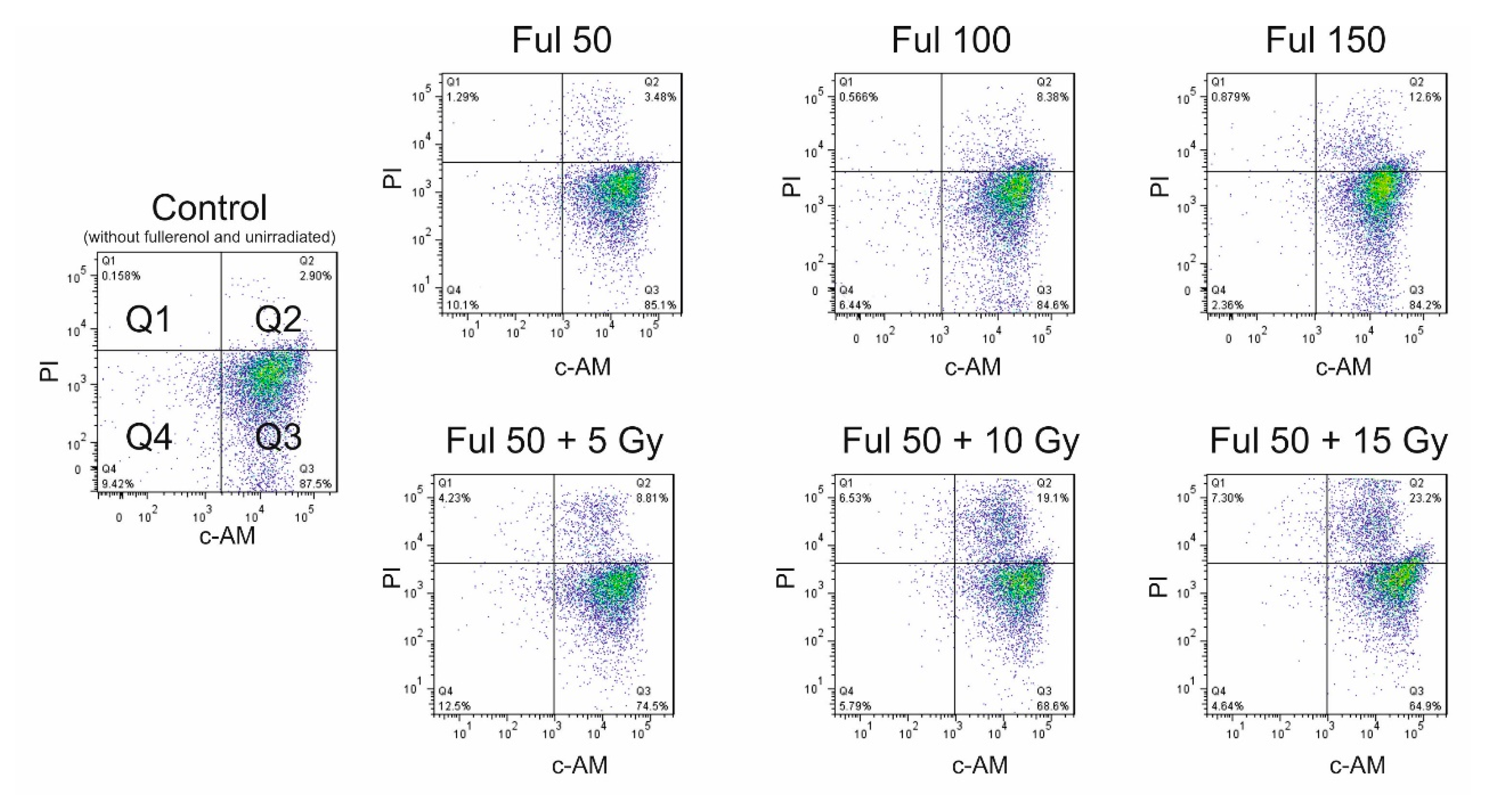
2.2. Direct Evidence of Fullerenol Internalization by Confocal Microscopy and Flow Cytofluorometry
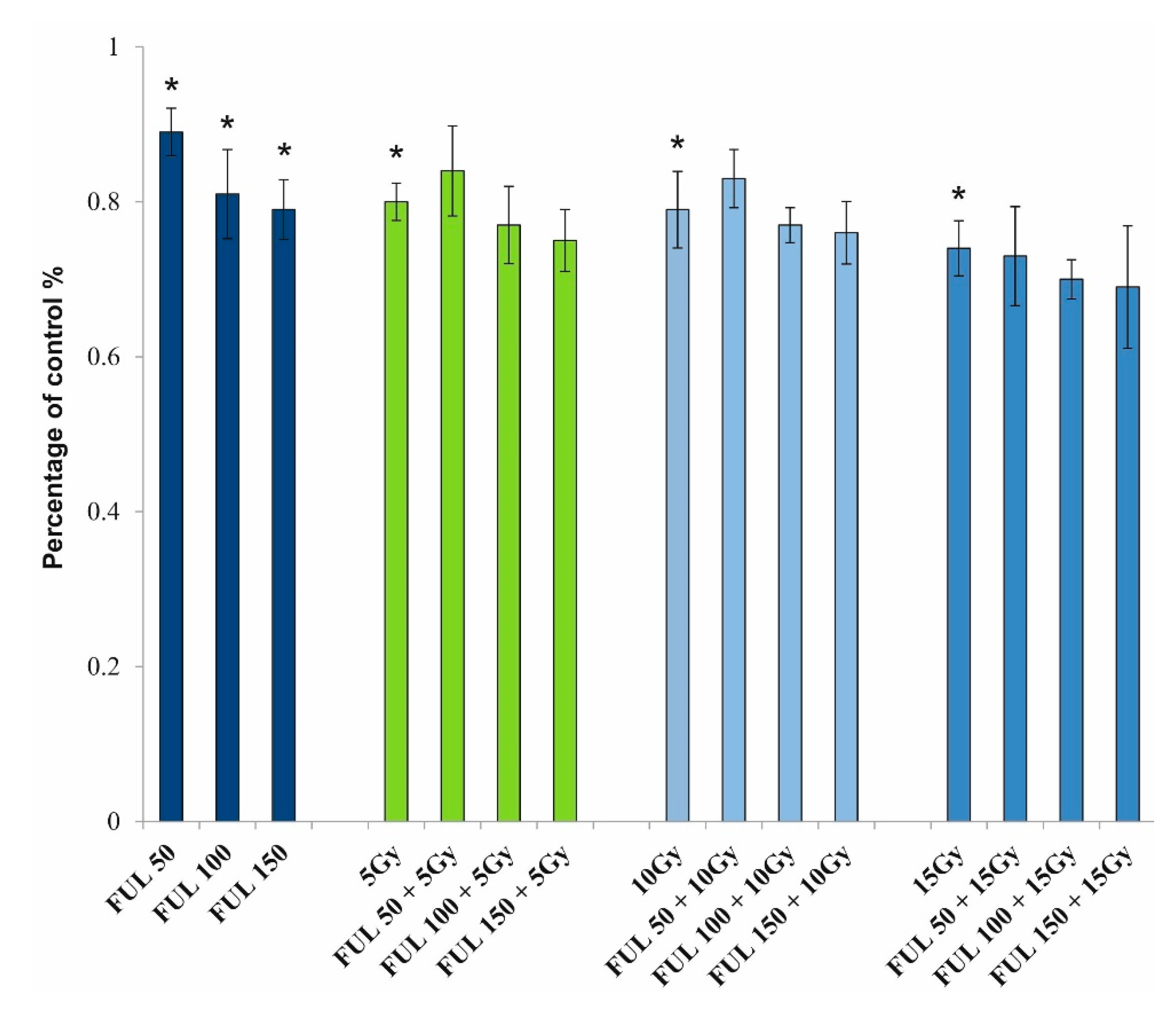
2.3. Effects of Fullerenol on the Viability of PBMCs
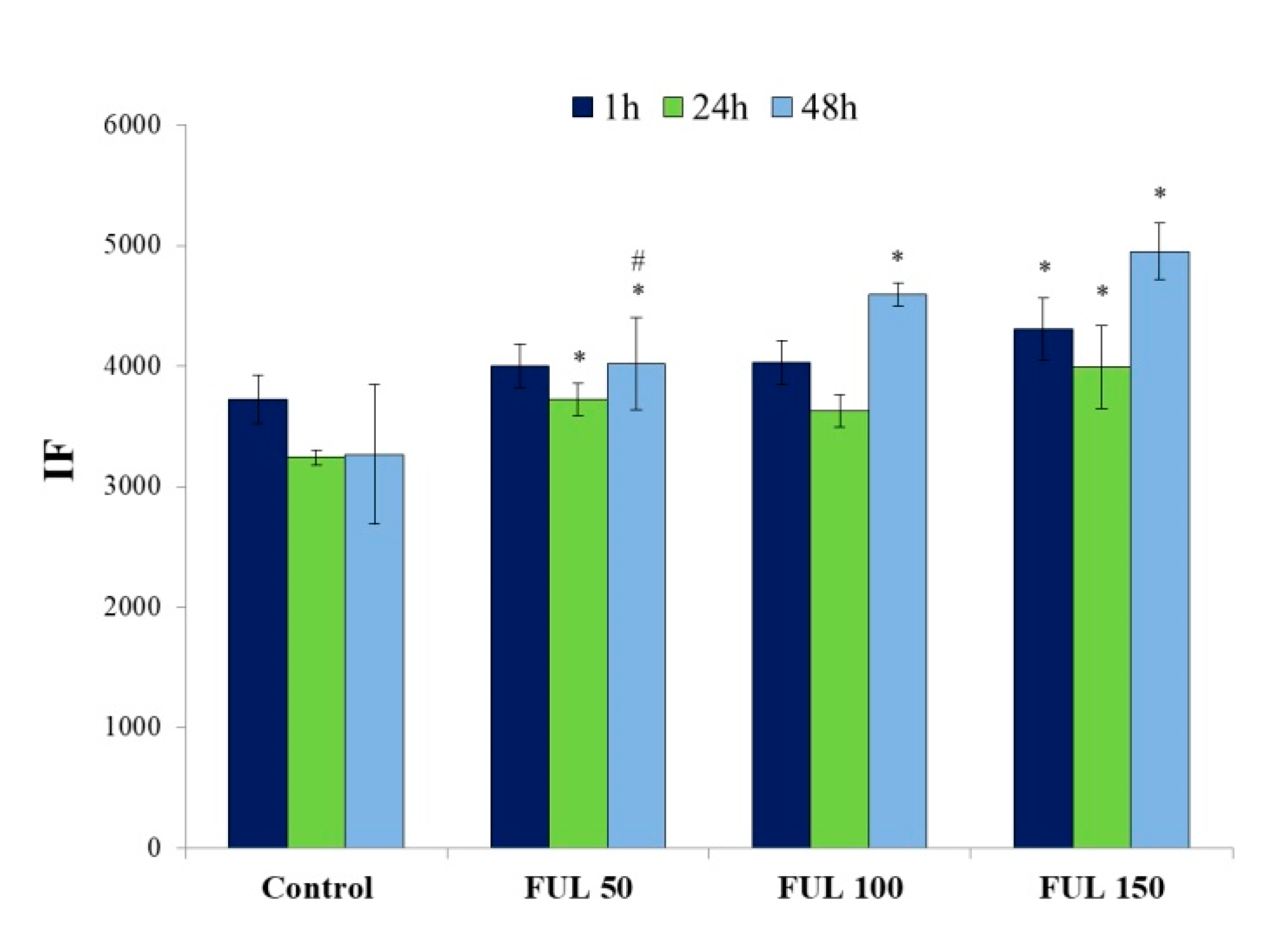
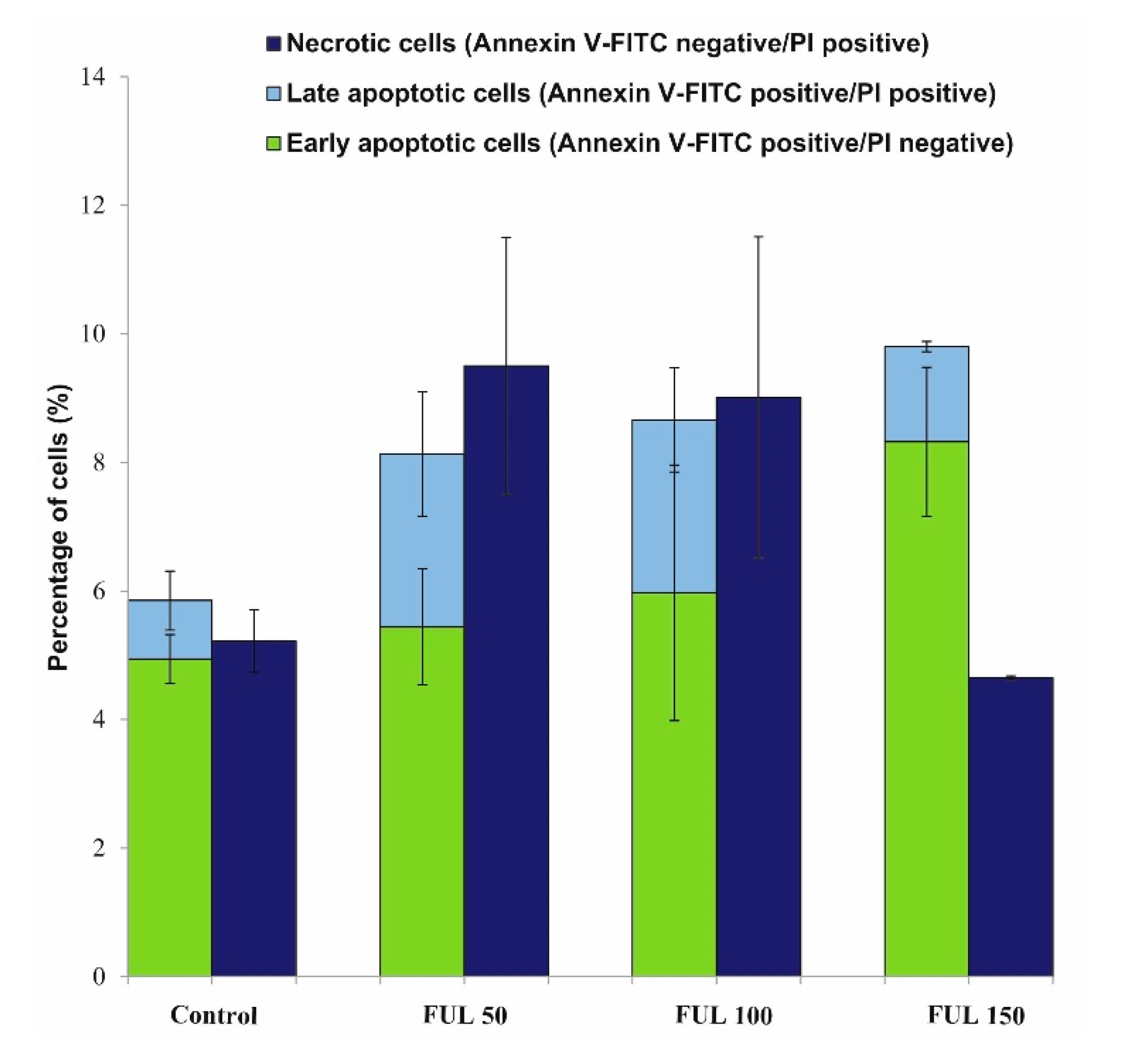
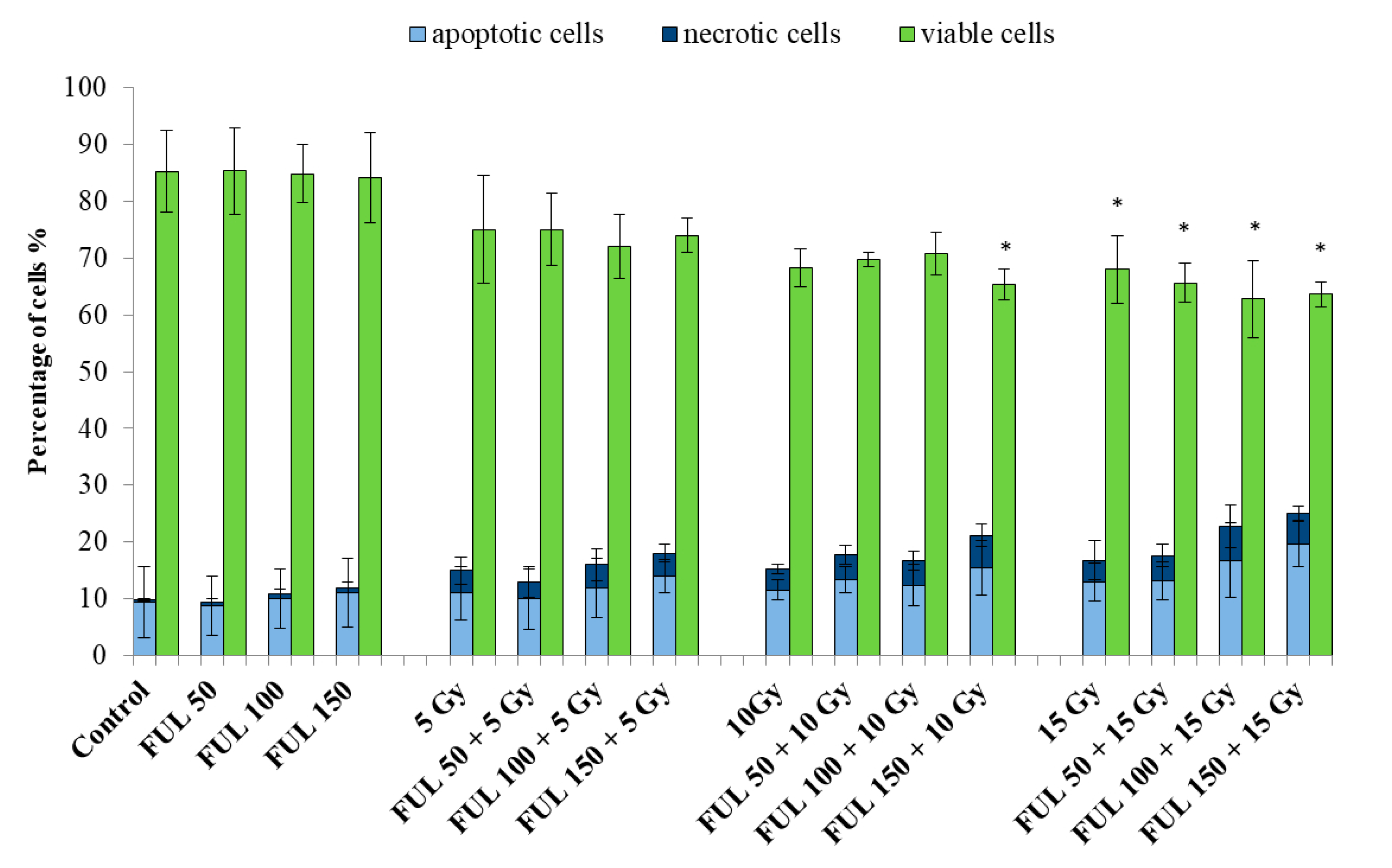
2.4. The Impact of Fullerenol on Human Peripheral Blood Mononuclear Cells under Radiation-Generated Oxidative Stress
2.5. Assessment of Mitochondrial Membrane Potential after Incubation with Fullerenol
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Methods
4.2.1. Synthesis of Fullerenol Nanoparticles
4.2.2. AFM Characterization of Fullerenol
4.2.3. Measurement of Size
4.2.4. Measurement of Zeta Potential
4.2.5. Preparation of Peripheral Blood Mononuclear Cell Suspensions
4.2.6. Fullerenol Treatment Conditions
4.2.7. Measurement of Fullerenol Internalization by Confocal Microscopy
4.2.8. Measurement of Fullerenol Internalization by Flow Cytofluorometry
4.2.9. Conditions of Irradiation
4.2.10. Cell Viability
4.2.11. Detection of Apoptosis
4.2.12. Detection of Mitochondrial Membrane Potential
4.2.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AFM | Atomic force microscopy |
| AO | Acridine orange |
| ATP | Adenosine triphosphate |
| c-AM | Calcein acetoxymethyl ester |
| DLS | Dynamic light scattering |
| DNA | Deoxyribonucleic acid |
| DOX | Doxorubicin |
| DSB | Double-strand breaks |
| EB | Ethidium bromide |
| ESR | Electron spin resonance spectroscopy |
| FBS | Fetal bovine serum |
| FITC | Fluorescein isothiocyanate |
| JC-1 | 5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimi-dazolylcarbocyanine iodide |
| MPT | Mitochondrial permeability transition |
| MRI | Magnetic resonance imaging |
| NMs | Nanomaterials |
| NPs | Nanoparticles |
| PBMCs | Peripheral blood mononuclear cells |
| PBS | Phosphate buffered saline |
| PI | Propidium iodide |
| PS | Phosphatydyl serine |
| ROS | Reactive oxygen species |
| TBARS | Thiobarbituric acid reactive substances |
| VDAC1 | Voltage-dependent anion channel 1 |
References
- Jeevanandam, J.; Barhoum, A.; Chan, Y.S.; Dufresne, A.; Danquah, M.K. Review on nanoparticles and nanostructured materials: History, sources, toxicity and regulations. Beilstein J. Nanotechnol. 2018, 9, 1050–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makvandi, P.; Wang, C.-Y.; Zare, E.N.; Borzacchiello, A.; Niu, L.-N.; Tay, F.R. Metal-based nanomaterials in biomedical applications: Antimicrobial activity and cytotoxicity aspects. Adv. Funct. Mater. 2020, 1910021. [Google Scholar] [CrossRef]
- Gupta, T.K.; Budarapu, P.R.; Chappidi, S.R.; YB, S.S.; Paggi, M.; Bordas, S.P. Advances in carbon based nanomaterials for bio-medical applications. Curr. Med. Chem. 2019, 26, 6851–6877. [Google Scholar] [CrossRef] [PubMed]
- Russ, K.A.; Elvati, P.; Parsonage, T.L.; Dews, A.; Jarvis, J.A.; Ray, M.; Schneider, B.; Smith, P.J.; Williamson, P.T.; Violi, A.; et al. C60 Fullerene localization and membrane interactions in RAW 264.7 immortalized mouse macrophages. Nanoscale 2016, 8, 4134–4144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrievsky, G.V.; Bruskov, V.I.; Tykhomyrov, A.A.; Gudkov, S.V. Peculiarities of the antioxidant and radioprotective effects of hydrated C60 fullerene nanostuctures in vitro and in vivo. Free Radic. Biol. Med. 2009, 47, 786–793. [Google Scholar] [CrossRef]
- Franskevych, D.; Palyvoda, K.; Petukhov, D.; Prylutska, S.; Grynyuk, I.; Schuetze, C.; Drobot, L.; Matyshevska, O.; Ritter, U. Fullerene C60 penetration into leukemic cells and its photoinduced cytotoxic effects. Nanoscale Res. Lett. 2017, 12, 40. [Google Scholar] [CrossRef] [Green Version]
- Panchuk, R.R.; Prylutska, S.V.; Chumakl, V.V.; Skorokhyd, N.R.; Lehka, L.V.; Evstigneev, M.P.; Prylutskyy, Y.I.; Berger, W.; Heffeter, P.; Scharff, P.; et al. Application of C60 fullerene-doxorubicin complex for tumor cell treatment in vitro and in vivo. J. Biomed. Nanotechnol. 2015, 11, 1139–1152. [Google Scholar] [CrossRef]
- Gonchar, O.O.; Maznychenko, A.V.; Bulgakova, N.V.; Vereshchaka, I.V.; Tomiak, T.; Ritter, U.; Prylutskyy, Y.I.; Mankovska, I.M.; Kostyukov, A.I. C60 Fullerene Prevents Restraint Stress-Induced Oxidative Disorders in Rat Tissues: Possible Involvement of the Nrf2/ARE-Antioxidant Pathway. Oxid. Med. Cell Longev. 2018, 2018, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Aly, F.M.; Othman, A.; Haridy, M.A.M. Protective Effects of Fullerene C60 Nanoparticles and Virgin Olive Oil against Genotoxicity Induced by Cyclophosphamide in Rats. Oxid. Med. Cell Longev. 2018, 2018, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Gao, N.; Wang, X.; Ren, J.; Qu, X. Near-Infrared Switchable Fullerene-Based Synergy Therapy for Alzheimer’s Disease. Small 2018, 14, 1801852. [Google Scholar] [CrossRef]
- Johnston, H.J.; Hutchison, G.R.; Christensen, F.M.; Aschberger, K.; Stone, V. The biological mechanisms and physicochemical characteristics responsible for driving fullerene toxicity. Toxicol. Sci. 2010, 114, 162–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moussa, F. [60]Fullerene and derivatives for biomedical applications. In Nanobiomaterials: Nanostructured Materials for Biomedical Applications; Narayan, R., Ed.; Woodhead Publishing: Duxford, UK, 2018; pp. 113–136. [Google Scholar] [CrossRef]
- Ringwood, A.H.; Levi-Polyachenko, N.; Carroll, D.L. Fullerene Exposures with Oysters: Embryonic, Adult, and Cellular Responses. Environ. Sci. Technol. 2009, 43, 7136–7141. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.M.; Dinis, A.M.; Peixoto, F.; Ferreira, L.; Jurado, A.S.; Videira, R.A. Interaction of Fullerene Nanoparticles With Biomembranes: From the Partition in Lipid Membranes to Effects on Mitochondrial Bioenergetics. Toxicol. Sci. 2014, 138, 117–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, F.; Mackeyev, Y.; Watson, E.; Cheney, M.A.; Wilson, L.J.; Suh, J. Evidence for nuclear internalisation of biocompatible [60] fullerene. Eur. J. Nanomed. 2013, 5, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Castro, E.; Hernandez Garcia, A.; Zavala, G.; Echegoyen, L. Echegoyen Fullerenes in biology and medicine. J. Mater. Chem. 2017, B5, 6523–6535. [Google Scholar] [CrossRef] [PubMed]
- Sachkova, A.S.; Kovel, E.S.; Churilov, G.N.; Guseynov, O.A.; Bondar, A.A.; Dubinina, I.A.; Kudryasheva, N.S. On mechanism of antioxidant effect of fullerenols. Biochem. Biophys. Rep. 2017, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Vranes, M.; Borisev, I.; Tot, A.; Armakovic, S.; Armakovic, S.; Jovic, D.; Gadzuric, S.; Djordjevic, A. Self-assembling, reactivity and molecular dynamics of fullerenol nanoparticles. Phys. Chem. Chem. Phys. 2017, 19, 135–144. [Google Scholar] [CrossRef]
- Lao, F.; Li, W.; Han, D.; Qu, Y.; Liu, Y.; Zhao, Y.; Chen, C. Fullerene derivatives protect endothelial cells against NO-induced damage. Nanotechnology 2009, 20, 225103. [Google Scholar] [CrossRef]
- Zemanova, E.; Danihelka, P.; Klouda, K.; Zeman, K. C60 Fullerene Derivative: Radioprotective Properties and a Toxicity Test in Vivo & in Vitro. J. Mater. Sci. Eng. A 2012, 2, 1–11. [Google Scholar]
- Zhu, G.Z.; Liu, Y.; Hashikawa, Y.; Zhang, Q.F.; Murata, Y.; Wang, L.S. Probing the interaction between the encapsulated water molecule and the fullerene cages in H2O@C60− and H2O@C59N−. Chem. Sci. 2018, 9, 5666–5671. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.J.; Lao, F.; Fu, P.P.; Wamer, W.G.; Zhao, Y.; Wang, P.C.; Qiu, Y.; Sun, B.; Xing, G.; Dong, J.; et al. The scavenging of reactive oxygen species and the potential for cell protection by functionalized fullerene materials. Biomaterials 2009, 30, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Jacevic, V.; Djordjevic, A.; Srdjenovic, B.; Milic-Tores, V.; Segrt, Z.; Dragojevic-Simic, V.; Kuca, K. Fullerenol nanoparticles prevents doxorubicin-induced acute hepatotoxicity in rats. Exp. Mol. Pathol. 2017, 102, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Geng, H.; Chang, Y.N.; Bai, X.; Liu, S.; Yuan, Q.; Gu, W.; Li, J.; Chen, K.; Xing, G.; Xing, G. Fullerenol nanoparticles suppress RANKL-induced osteoclastogenesis by inhibiting differentiation and maturation. Nanoscale 2017, 9, 12516–12523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.Y.; Hua, S.Y.; Zhou, Z.Q.; Wang, G.C.; Jiang, F.L.; Liu, Y. Characterization of fullerenol-protein interactions and an extended investigation on cytotoxicity. Colloids Surf. B Biointerfaces 2017, 157, 261–267. [Google Scholar] [CrossRef]
- Rokitskaya, T.I.; Antonenko, Y.N. Fullerenol C60(OH)24 increases ion permeability of lipid membranes in a pH-dependent manner. Biochim. Biophys. Acta 2016, 1858, 1165–1174. [Google Scholar] [CrossRef]
- Krokosz, A.; Lichota, A.; Nowak, K.E.; Grebowski, J. Carbon nanoparticles as possible radioprotectors in biological systems. Radiat. Phys. Chem. 2016, 128, 143–150. [Google Scholar] [CrossRef]
- Islam, M.T. Radiation interactions with biological systems. Int. J. Radiat. Biol. 2017, 93, 487–493. [Google Scholar] [CrossRef]
- Vavrova, J.; Rezacova, M.; Pejchal, J. Fullerene nanoparticles and their anti-oxidative effects: A comparison to other radioprotective agents. J. Appl. Biomed. 2012, 10, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Xie, J.; Dong, X.; Mei, L.; Zhao, M.; Leng, Z.; Hu, H.; Li, L.; Gu, Z.; Zhao, Y. Clinically Approved Carbon Nanoparticles with Oral Administration for Intestinal Radioprotection via Protecting the Small Intestinal Crypt Stem Cells and Maintaining the Balance of Intestinal Flora. Small 2020, 1906915. [Google Scholar] [CrossRef]
- Tyurin, D.P.; Kolmogorov, F.S.; Cherepkova, I.A.; Charykov, N.A.; Semenov, K.; Keskinov, V.A.; Safyannikov, N.M.; Pukharenko, Y.V.; Letenko, D.G.; Segeda, T.A.; et al. Antioxidant properties of fullerenol-d. Nanosyst. Phys. Chem. Math. 2018, 9, 798–810. [Google Scholar] [CrossRef] [Green Version]
- Grebowski, J.; Krokosz, A.; Puchala, M. Membrane fluidity and activity of membrane ATPases in human erythrocytes under the influence of polyhydroxylated fullerene. Biochim. Biophys. Acta 2013, 1828, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grebowski, J.; Kazmierska, P.; Litwinienko, G.; Lankoff, A.; Wolszczak, M.; Krokosz, A. Fullerenol C60(OH)36 protects human erythrocyte membrane against high-energy electrons. Biochim. Biophys. Acta 2018, 1860, 1528–1536. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Iijima, R.; Ako, J.; Shinke, T.; Okada, H.; Ito, Y.; Ando, K.; Anzai, H.; Tanaka, H.; Ueda, Y.; et al. NIPPON Investigators. Dual antiplatelet therapy for 6 versus 18 months after biodegradable polymer drug-eluting stent implantation. JACC Cardiovasc. Interv. 2017, 10, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Clarke, R.; Bennett, D.; Guo, Y.; Walters, R.G.; Hill, M.; Parish, S.; Millwood, I.Y.; Bian, Z.; Chen, Y.; et al. China Kadoorie Biobank Collaborative Group; International Steering Committee; International Co-ordinating Centre, Oxford; National Co-ordinating Centre, Beijing; Regional Co-ordinating Centres. Causal associations of blood lipids with risk of ischemic stroke and intracerebral hemorrhage in Chinese adults. Nat. Med. 2019, 25, 569–574. [Google Scholar] [CrossRef]
- Li, Y.; Luo, H.B.; Zhang, H.Y.; Guo, Q.; Yao, H.C.; Li, J.Q.; Chang, Q.; Yang, J.G.; Wang, F.; Wang, C.D.; et al. Potential hepatoprotective effects of fullerenol nanoparticles on alcohol-induced oxidative stress by ROS. RSC Adv. 2016, 6, 31122–31130. [Google Scholar] [CrossRef]
- Lee, J.; Lee, C.; Osawa, E.; Lee, K.M. Dynamic Behavior of C60 Fullerene in Carbon Nanopeapods: Tight-Binding Molecular Dynamics Simulation. Bull. Korean Chem. Soc. 2019. [Google Scholar] [CrossRef]
- Mottier, A.; Mouchet, F.; Pinelli, E.; Gauthier, L.; Flahaut, E. Environmental impact of engineered carbon nanoparticles: From releases to effects on the aquatic biota. Curr. Opin. Biotechnol. 2017, 46, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bundschuh, M.; Filser, J.; Luderwald, S.; McKee, M.S.; Metreveli, G.; Schaumann, G.E.; Schulz, R.; Wagner, S. Nanoparticles in the environment: Where do we come from, where do we go to? Environ. Sci. Eur. 2018, 30. [Google Scholar] [CrossRef] [Green Version]
- Sigmund, G.; Jiang, C.; Hofmann, T.; Chen, W. Environmental transformation of natural and engineered carbon nanoparticles and implications for the fate of organic contaminants. Environ. Sci-Nano 2018. [Google Scholar] [CrossRef] [Green Version]
- Johnson-Lyles, D.N.; Peifley, K.; Lockett, S.; Neun, B.W.; Hansen, M.; Clogston, J.; Stern, S.T.; McNeil, S.E. Fullerenol cytotoxicity in kidney cells is associated with cytoskeleton disruption, autophagic vacuole accumulation, and mitochondrial dysfunction. Toxicol. Appl. Pharmacol. 2010, 248, 249–258. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Xu, J.Y.; Shen, P.; Li, J.; Wang, L.; Li, Q.; Li, W.; Xu, G.T.; Fan, C.; Huang, Q. Cellular uptake and cytotoxic evaluation of fullerenol in different cell lines. Toxicology 2010, 269, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Qiao, R.; Roberts, A.P.; Mount, A.S.; Klaine, S.J.; Ke, P.C. Translocation of C60 and its derivatives across a lipid bilayer. Nano Lett. 2007, 7, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Unfried, K.; Albrecht, C.; Klotz, L.-O.; Von Mikecz, A.; Grether-Beck, A.; Schins, R.P.F. Cellular responses to nanoparticles: Target structures and mechanisms. Nanotoxicology 2007, 1, 52–71. [Google Scholar] [CrossRef]
- Foroozandeh, P.; Aziz, A.A. Insight into Cellular Uptake and Intracellular Trafficking of Nanoparticles. Nanoscale Res. Lett. 2018, 13, 339. [Google Scholar] [CrossRef]
- Albanese, A.; Tang, P.S.; Chan, W.C. The effect of nanoparticle size, shape, and surface chemistry on biological systems. Annu. Rev. Biomed. Eng. 2012, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Eom, H.-J.; Choi, J. Clathrin-mediated endocytosis is involved in uptake and toxicity of silica nanoparticles in Caenohabditis elegans. Chem. Biol. Interact. 2019, 311, 108774. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Suzuki, T.; Ishii, H.; Nakae, D.; Ogata, A. Cytotoxic effects of hydroxylated fullerenes on isolated rat hepatocytes via mitochondrial dysfunction. Arch. Toxicol. 2011, 85, 1429–1440. [Google Scholar] [CrossRef]
- Shimizu, K.; Kubota, R.; Kobayashi, N.; Tahara, M.; Sugimoto, N.; Nishimura, T.; Ikarashi, Y. Cytotoxic Effects of Hydroxylated Fullerenes in Three Types of Liver Cells. Materials 2013, 6, 2713–2722. [Google Scholar] [CrossRef] [Green Version]
- Chaban, V.V.; Fileti, E.E. Which fullerenols are water soluble? Systematic atomistic investigation. New J. Chem. 2017, 41, 184–189. [Google Scholar] [CrossRef]
- Bednarikova, Z.; Huy, P.D.; Mocanu, M.M.; Fedunova, D.; Li, M.S.; Gazova, Z. Fullerenol C60(OH)16 prevents amyloid fibrillization of Aβ40 – in vitro and in silico approach. Phys. Chem. Chem. Phys. 2016, 18, 18855–18867. [Google Scholar] [CrossRef]
- Poovaiah, N.; Davoudi, Z.; Peng, H.; Schlichtmann, B.; Mallapragada, S.; Narasimhan, B.; Wang, Q. Treatment of neurodegenerative disorders through the blood-brain barrier using nanocarriers. Nanoscale 2018, 10, 16962–16983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seke, M.; Petrovic, D.; Labudovic Borovic, M.; Borisev, I.; Novakovic, M.; Rakocevic, Z.; Djordjevic, A. Fullerenol/iron nanocomposite diminishes doxorubicin-induced toxicity. J. Nanopart Res. 2019, 21, 239. [Google Scholar] [CrossRef]
- Heimann, J.; Morrow, L.; Anderson, R.E.; Barron, A.R. Understanding the relative binding ability of hydroxyfullerene to divalent and trivalent metals. Dalton Trans. 2015, 44, 4380–4388. [Google Scholar] [CrossRef] [PubMed]
- Lichota, A.; Krokosz, A. Fullerenols in therapy and diagnosis of cancer. Med. Pr. 2016, 67, 817–831. [Google Scholar] [CrossRef]
- Acquah, S.F.A.; Penkova, A.V.; Markelov, D.A.; Semisalova, A.S.; Leonhardt, B.E.; Magi, J.M. Review—The Beautiful Molecule: 30 Years of C60 and Its Derivatives. ECS J. Solid State Sci. Technol. 2017, 6, M3155–M3162. [Google Scholar] [CrossRef]
- Bacchetta, R.; Santo, N.; Valenti, I.; Maggioni, D.; Longhi, M.; Tremolada, P. Comparative toxicity of three differently shaped carbon nanomaterials on Daphnia magna: Does a shape effect exist? Nanotoxicology 2018, 12, 201–223. [Google Scholar] [CrossRef]
- Bayal, M.; Janardhanan, P.; Tom, E.; Chandran, N.; Devadathan, S.; Ranjeet, D.; Unniyampurath, U.; Pilankatta, R.; Nair, S.S. Cytotoxicity of nanoparticles - Are the size and shape only matters? Or the media parameters too? A study on band engineered ZnS nanoparticles and calculations based on equivolume stress model. Nanotoxicology 2019, 1–16. [Google Scholar] [CrossRef]
- Yang, L.Y.; Gao, J.L.; Gao, T.; Dong, P.; Ma, L.; Jiang, F.L.; Liu, Y. Toxicity of polyhydroxylated fullerene to mitochondria. J. Hazard. Mater. 2016, 301, 119–126. [Google Scholar] [CrossRef]
- Walsh, D.; Siebenwirth, C.; Greubel, C.; Ilicic, K.; Reindl, J.; Girst, S.; Muggiolu, G.; Simon, M.; Barberet, P.; Seznec, H.; et al. Live cell imaging of mitochondria following targeted irradiation in situ reveals rapid and highly localized loss of membrane potential. Sci. Rep. 2017, 7, 46684. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Jia, H.; Liu, Z.; Hou, B.; Luo, C.; Feng, Z.; Li, W.; Liu, J. Polyhydroxylated fullerene derivative C60(OH)24prevents mitochondrial dysfunction and oxidative damage in an MPP+-induced cellular model of Parkinson’s disease. J. Neurosci. Res. 2008, 86, 3622–3634. [Google Scholar] [CrossRef]
- Nowak, K.; Krokosz, A.; Rodacka, A.; Puchala, M. Study on the effect of polyhydroxylated fullerene, C60(OH)36, on X-ray irradiated human peripheral blood mononuclear cells. Radiat. Phys. Chem. 2014, 97, 325–331. [Google Scholar] [CrossRef]
- Zamzami, N.; Larochette, N.; Kroemer, G. Mitochondrial permeability transition in apoptosis and necrosis. Cell Death Differ. 2005, 12, 1478–1480. [Google Scholar] [CrossRef] [PubMed]
- Kam, W.W.; Banati, R.B. Effects of ionizing radiation on mitochondria. Free Radic. Biol. Med. 2013, 65, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Shoshan-Barmatz, V.; Krelin, Y.; Shteinfer-Kuzmine, A. VDAC1 functions in Ca2+ homeostasis and cell life and death in health and disease. Cell Calcium 2018, 69, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gao, X.; Zhao, Y. Mechanisms of Antioxidant Activities of Fullerenols from First Principles Calculation. J. Phys. Chem. A 2018, 122, 8183–8190. [Google Scholar] [CrossRef]
- Krokosz, A.; Grebowski, J.; Rodacka, A.; Pasternak, B.; Puchala, M. The effect of fullerenol C60(OH)~30 on the alcohol dehydrogenase activity irradiated with X-rays. Radiat. Phys. Chem. 2014, 97, 102–106. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water | 0.02 M Phosphate Buffer pH 7.4 | |
|---|---|---|
| zeta potential (mV) ± SD | −27.5 ± 1.0 | −37.4 ± 3.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lichota, A.; Piwoński, I.; Michlewska, S.; Krokosz, A. A Multiparametric Study of Internalization of Fullerenol C60(OH)36 Nanoparticles into Peripheral Blood Mononuclear Cells: Cytotoxicity in Oxidative Stress Induced by Ionizing Radiation. Int. J. Mol. Sci. 2020, 21, 2281. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072281
Lichota A, Piwoński I, Michlewska S, Krokosz A. A Multiparametric Study of Internalization of Fullerenol C60(OH)36 Nanoparticles into Peripheral Blood Mononuclear Cells: Cytotoxicity in Oxidative Stress Induced by Ionizing Radiation. International Journal of Molecular Sciences. 2020; 21(7):2281. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072281
Chicago/Turabian StyleLichota, Anna, Ireneusz Piwoński, Sylwia Michlewska, and Anita Krokosz. 2020. "A Multiparametric Study of Internalization of Fullerenol C60(OH)36 Nanoparticles into Peripheral Blood Mononuclear Cells: Cytotoxicity in Oxidative Stress Induced by Ionizing Radiation" International Journal of Molecular Sciences 21, no. 7: 2281. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072281