Sugar Beet (Beta vulgaris) Guard Cells Responses to Salinity Stress: A Proteomic Analysis


Abstract
:1. Introduction
2. Results
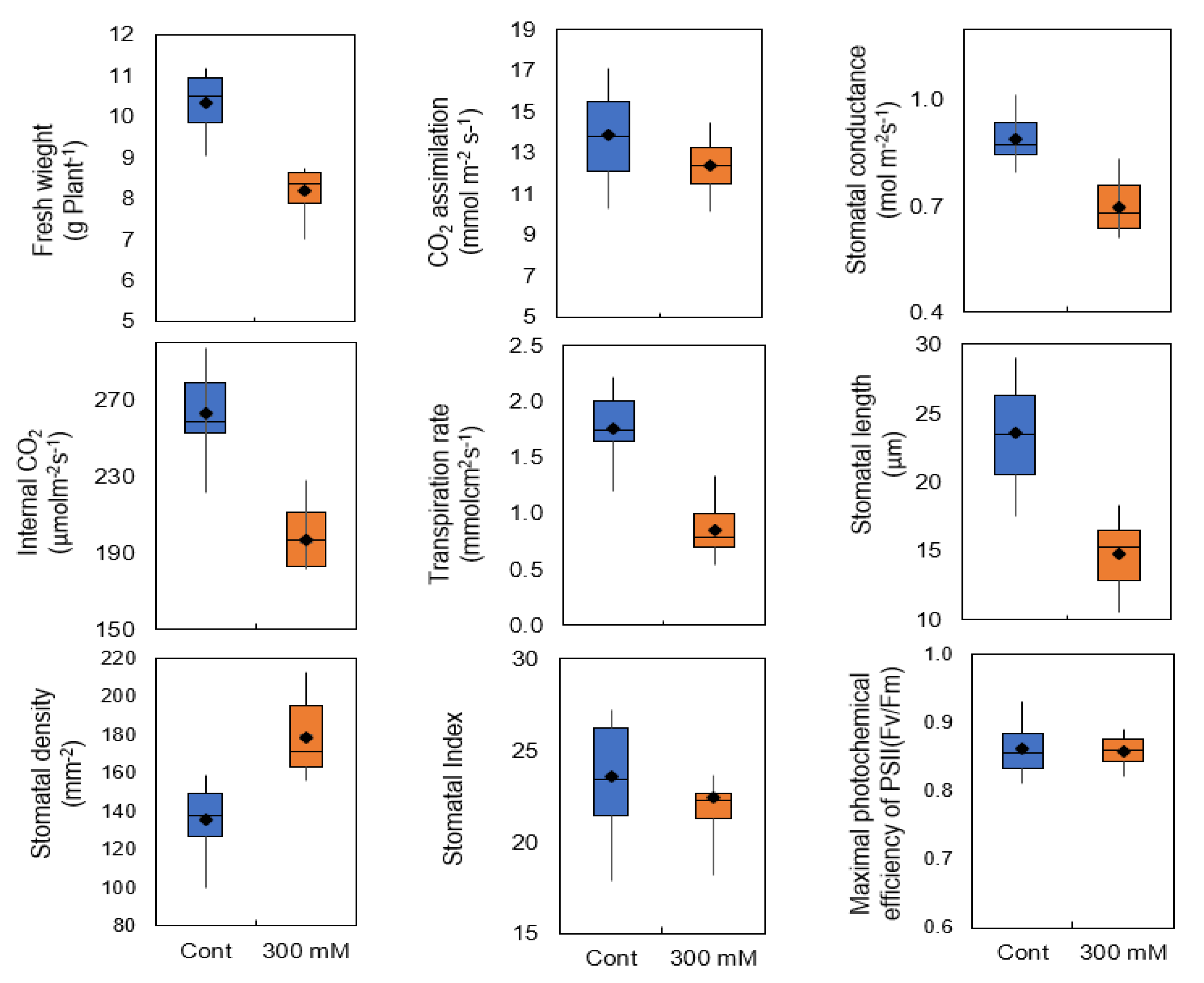
2.1. Stomatal Traits and Conductance in Sugar Beet
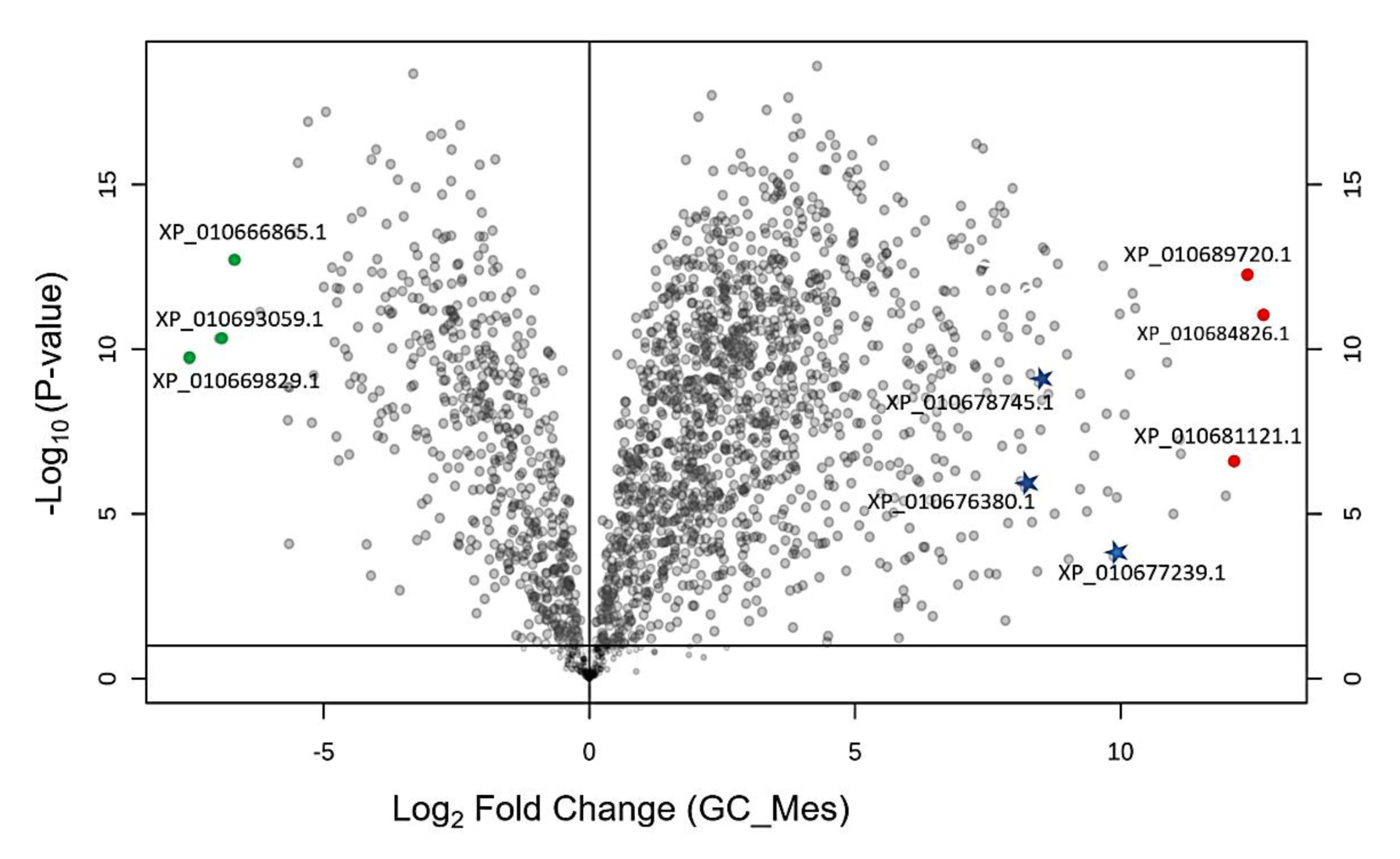
2.2. Comparison of Ground State Mesophyll and Guard Cells Proteome
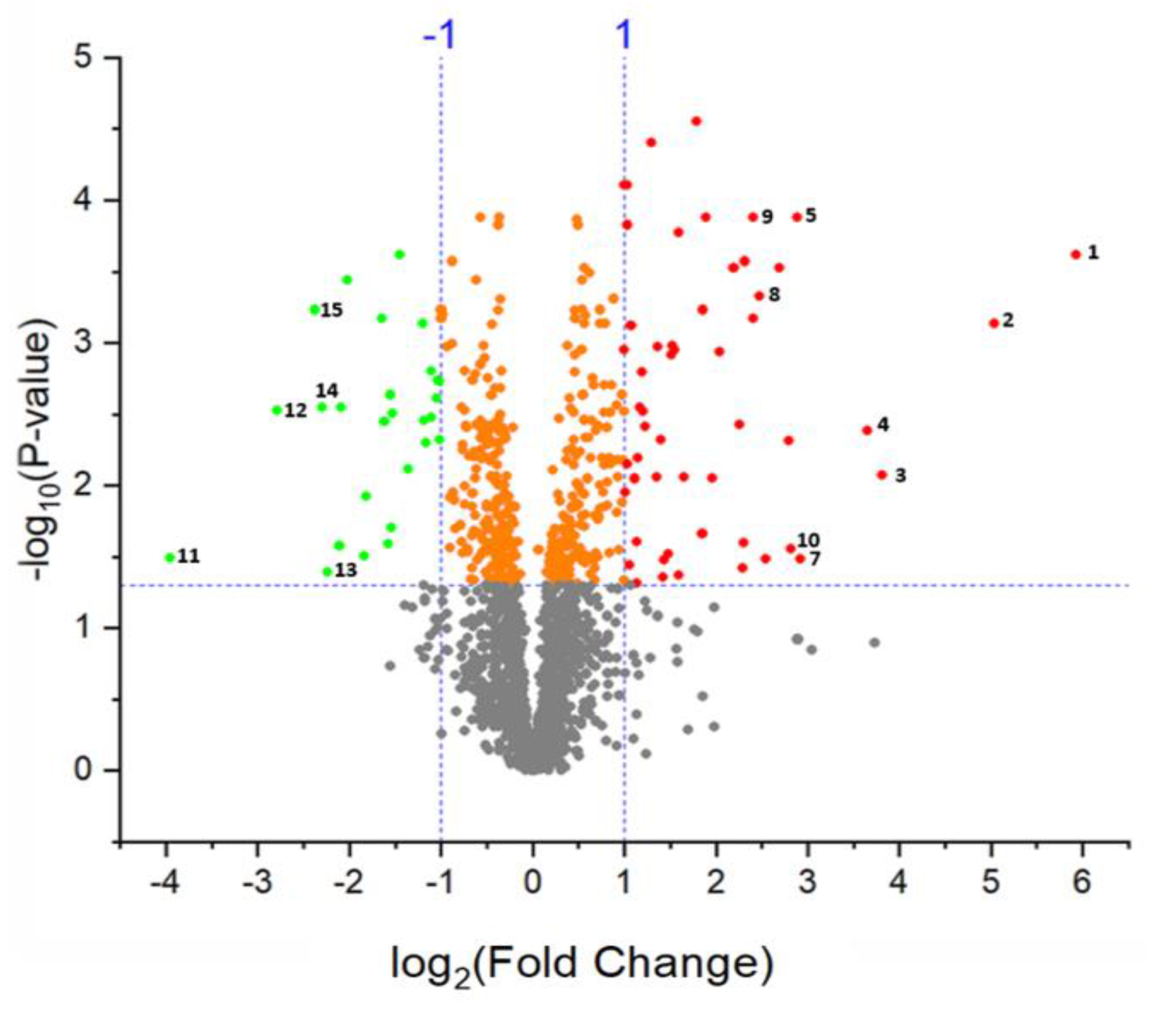
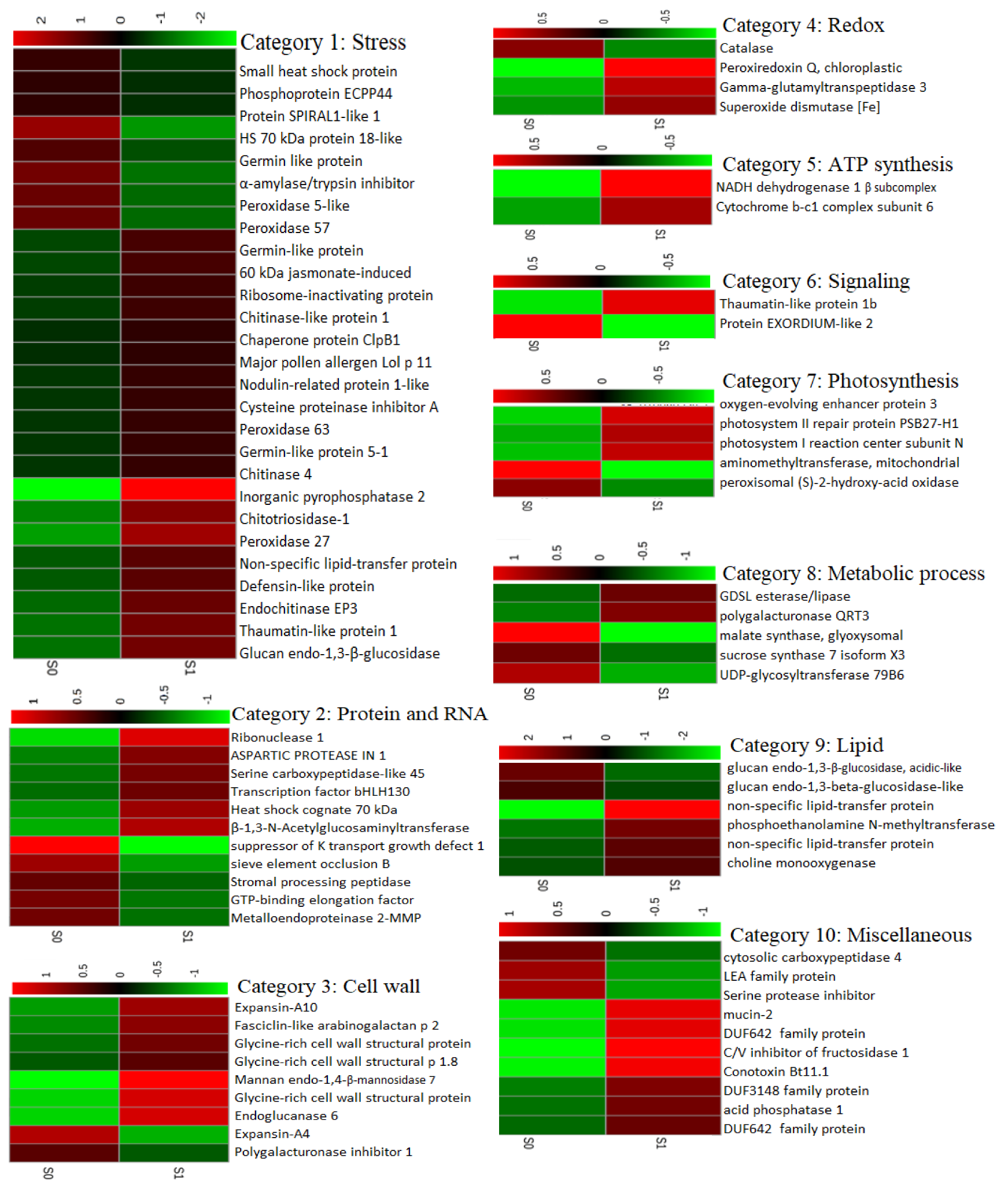
2.3. Differentially Abundant Proteins in Response to Salt Treatment in Guard Cells (GCs)
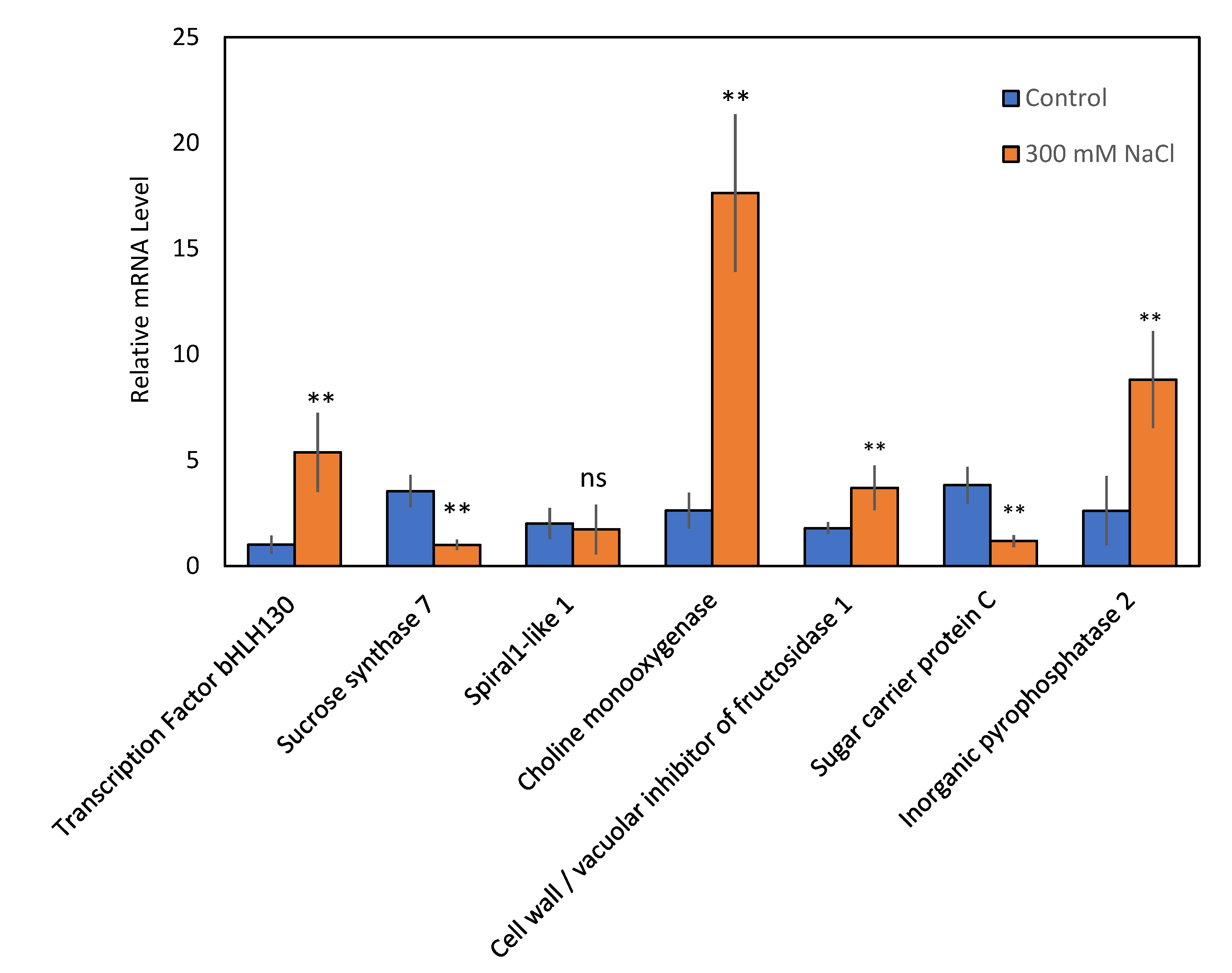
2.4. Transcription-Level Analysis of Genes Corresponding to Proteins Modulated in Salt-Stressed Guard Cells
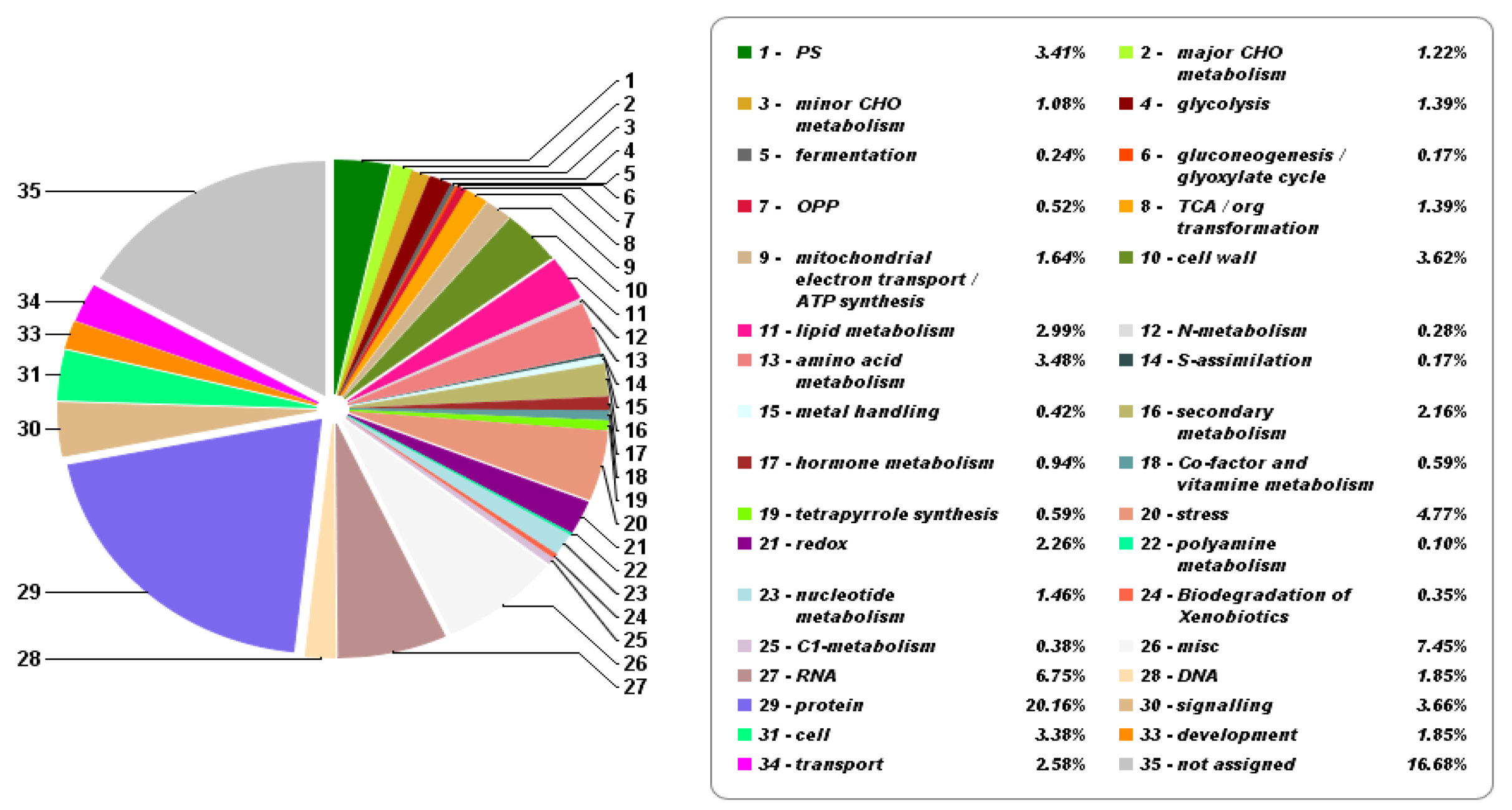
2.5. Differentially Abundant Proteins in Mesophyll Under Salinity Treatment
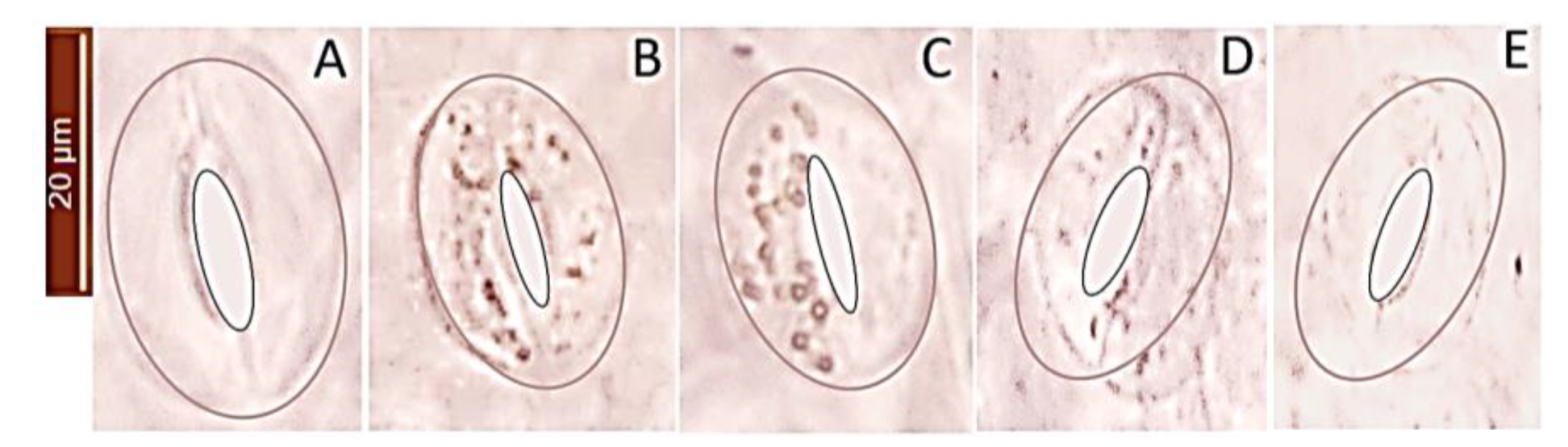
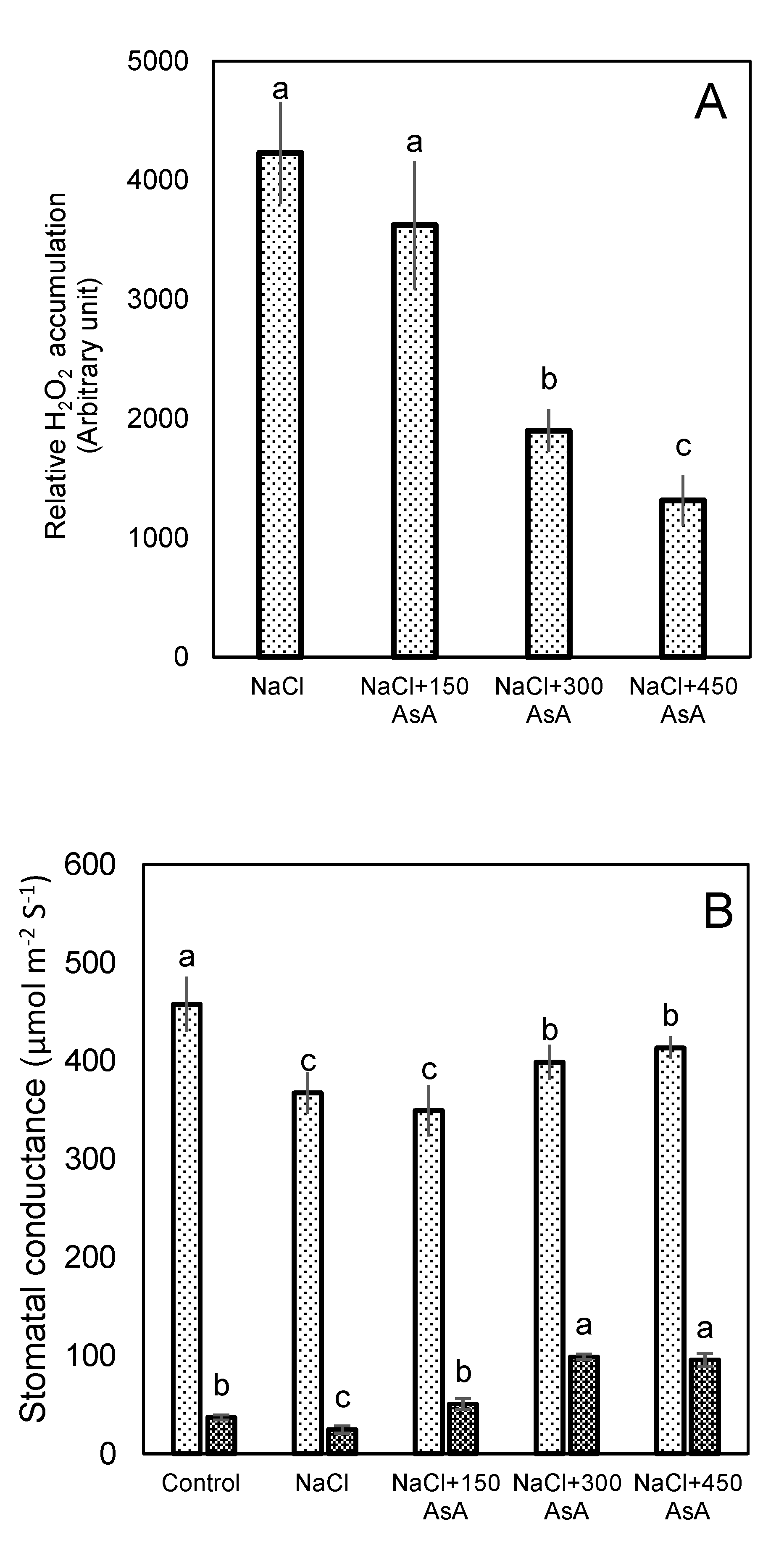
2.6. H2O2 Accumulation in Guard in Response to Salt Stress and Ascorbic Acid
3. Discussion
3.1. Most Abundant Proteins in GCs
3.2. Stress Related Proteins Differentially Abundant Proteins in GCs Under Salinity Treatment
3.3. Redox-Related Proteins
3.4. Photosynthesis and ATP Synthesis
3.5. Protein and RNA Biosynthesis
3.6. Cell Wall
3.7. Lipid Metabolism
3.8. Proteins Downregulated in Sugar Beet GCs in Response to Salinity
4. Materials and Methods
4.1. Growth conditions and physiological Parameters
4.2. In Situ Detection of H2O2 in Guard Cells and Ascorbic Acid Treatments
4.3. GC and Mesophyll Preparation
4.4. Protein Extraction
4.5. Enzymatic Digestion and Off-Line Desalting
4.6. Protein Identification by NanoLC–MS/MS
4.7. Database Searching and Protein Quantitation
4.8. Determination of Relative Protein Abundance and Statistical Analysis
4.9. Real-Time Quantitative Reverse Transcription PCR Analysis
4.10. Annotation and Gene Ontology (GO) Enrichment Analysis
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DAB | 3,3’-Diaminobenzidine |
| GC | Guard cell |
| GO | Gene Ontology |
| Mes | Mesophyll |
References
- Shabala, S.; Bose, J.; Hedrich, R. Salt bladders: Do they matter? Trends Plant Sci. 2014, 19, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.-Q.; Wang, J.-L.; Feng, R.-J.; Li, S.-J.; Wang, C.-M. iTRAQ-Based Comparative Proteomic Analysis Provides Insights into Molecular Mechanisms of Salt Tolerance in Sugar Beet (Beta vulgaris L.). Int. J. Mol. Sci. 2018, 19, 3866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, T. Guard cell photosynthesis and stomatal function. New Phytol. 2008, 181, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhang, W.; Stanley, B.; Assmann, S.M. Functional Proteomics of Arabidopsis thaliana Guard Cells Uncovers New Stomatal Signaling Pathways[W][OA]. Plant Cell 2008, 20, 3210–3226. [Google Scholar] [CrossRef] [Green Version]
- Dai, S.; Chen, S. Single-cell-type proteomics: Toward a holistic understanding of plant function. Mol. Cell. Proteom. 2012, 11, 1622–1630. [Google Scholar] [CrossRef] [Green Version]
- Specht, H.; Slavov, N. Transformative Opportunities for Single-Cell Proteomics. J. Proteome Res. 2018, 17, 2565–2571. [Google Scholar] [CrossRef]
- Barkla, B.; Pantoja, O.; Vera-Estrella, R. Protein profiling of epidermal bladder cells from the halophyteMesembryanthemum crystallinum. Proteomics 2012, 12, 2862–2865. [Google Scholar] [CrossRef]
- Zhu, M.; Dai, S.; McClung, S.; Yan, X.; Chen, S. Functional differentiation of Brassica napus guard cells and mesophyll cells revealed by comparative proteomics. Mol. Cell. Proteom. 2008, 8, 752–766. [Google Scholar] [CrossRef] [Green Version]
- Geilfus, C.-M.; Lan, J.; Carpentier, S. Dawn regulates guard cell proteins in Arabidopsis thaliana that function in ATP production from fatty acid beta-oxidation. Plant Mol. Biol. 2018, 98, 525–543. [Google Scholar] [CrossRef]
- Zhao, Z.; Stanley, B.; Zhang, W.; Assmann, S.M. ABA-Regulated G Protein Signaling inArabidopsisGuard Cells: A Proteomic Perspective. J. Proteome Res. 2010, 9, 1637–1647. [Google Scholar] [CrossRef]
- Skorupa, M.; Gołębiewski, M.; Kurnik, K.; Niedojadło, J.; Kęsy, J.; Klamkowski, K.; Wójcik, K.; Treder, W.; Tretyn, A.; Tyburski, J. Salt stress vs. salt shock—The case of sugar beet and its halophytic ancestor. BMC Plant Biol. 2019, 19, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozema, J.; Cornelisse, D.; Zhang, Y.; Li, H.; Bruning, B.; Katschnig, D.; Broekman, R.; Ji, B.; Van Bodegom, P. Comparing salt tolerance of beet cultivars and their halophytic ancestor: Consequences of domestication and breeding programmes. AoB PLANTS 2015, 7, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Nan, J.; Yu, B. OMICS Technologies and Applications in Sugar Beet. Front. Plant Sci. 2016, 7, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, D.; Baskin, C.C.; Baskin, J.M.; Yang, F.; Huang, Z. Dormancy cycling and persistence of seeds in soil of a cold desert halophyte shrub. Ann. Bot. 2013, 113, 171–179. [Google Scholar] [CrossRef]
- Yu, B.; Li, J.; Koh, J.; Dufresne, C.; Yang, N.; Qi, S.; Zhang, Y.; Ma, C.; Duong, B.V.; Chen, S.; et al. Quantitative proteomics and phosphoproteomics of sugar beet monosomic addition line M14 in response to salt stress. J. Proteom. 2016, 143, 286–297. [Google Scholar] [CrossRef] [Green Version]
- Weyers, J.D.B.; Travis, A. Selection and Preparation of Leaf Epidermis for Experiments on Stomatal Physiology. J. Exp. Bot. 1981, 32, 837–850. [Google Scholar] [CrossRef]
- Rother, T.; Acker, G.; Scheibe, P.D.R. Immunogold Localization of Chloroplast Protein in Spinach Leaf Mesophyll, Epidermis, and Guard Cells. Bot. Acta 1988, 101, 311–320. [Google Scholar] [CrossRef]
- Nguyen, C.D.L.; Malchow, S.; Reich, S.; Steltgens, S.; Shuvaev, K.V.; Loroch, S.; Lorenz, C.; Sickmann, A.; Knobbe-Thomsen, C.B.; Tews, B.; et al. A sensitive and simple targeted proteomics approach to quantify transcription factor and membrane proteins of the unfolded protein response pathway in glioblastoma cells. Sci. Rep. 2019, 9, 8836. [Google Scholar] [CrossRef]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. agriGO v2.0: A GO analysis toolkit for the agricultural community, 2017 update. Nucleic Acids Research 2017, 45, W122–W129. [Google Scholar] [CrossRef]
- Szypulska, E.; Jankowski, K.; Weidner, S. ABA pretreatment can limit salinity-induced proteome changes in growing barley sprouts. Acta Physiol. Plant. 2017, 39, 8. [Google Scholar] [CrossRef] [Green Version]
- Asaoka, M.; Inoue, S.-I.; Gunji, S.; Kinoshita, T.; Maeshima, M.; Tsukaya, H.; Ferjani, A. Excess Pyrophosphate within Guard Cells Delays Stomatal Closure. Plant Cell Physiol. 2019, 60, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, T.; Slezina, M.P.; Istomina, E.A.; Korostyleva, T.V.; Kovtun, A.S.; Kasianov, A.S.; Shcherbakova, L.A.; Kudryavtsev, A.M. Non-Specific Lipid Transfer Proteins in Triticum kiharae Dorof. et Migush.: Identification, Characterization and Expression Profiling in Response to Pathogens and Resistance Inducers. Pathogens 2019, 8, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, F.-S.; Tang, L.; Li, Y.-Y.; Wang, W.-C.; Yang, Z.; Li, X.-G.; Zeng, C. Differential proteomic analysis reveals the mechanism of Musa paradisiaca responding to salt stress. Mol. Biol. Rep. 2018, 46, 1057–1068. [Google Scholar] [CrossRef] [Green Version]
- Pitann, B.; Zörb, C.; Mühling, K. Comparative proteome analysis of maize (Zea maysL.) expansins under salinity. J. Plant Nutr. Soil Sci. 2009, 172, 75–77. [Google Scholar] [CrossRef]
- Medeiros, D.; Barros, J.A.; Fernie, A.R.; Araújo, W.L. Eating Away at ROS to Regulate Stomatal Opening. Trends Plant Sci. 2020, 25, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Santelia, D.; Lawson, T. Rethinking Guard Cell Metabolism. Plant Physiol. 2016, 172, 1371–1392. [Google Scholar] [CrossRef] [Green Version]
- Gharat, S.A.; Parmar, S.; Tambat, S.; Vasudevan, M.; Shaw, B.P. Transcriptome Analysis of the Response to NaCl in Suaeda maritima Provides an Insight into Salt Tolerance Mechanisms in Halophytes. PLoS ONE 2016, 11, e0163485. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, X.; Lu, C.; Zeng, X.; Li, Y.; Fu, D.; Wu, G. Non-specific lipid transfer proteins in plants: Presenting new advances and an integrated functional analysis. J. Exp. Bot. 2015, 66, 5663–5681. [Google Scholar] [CrossRef]
- Geng, S.; Yu, B.; Zhu, N.; Dufresne, C.; Chen, S. Metabolomics and Proteomics of Brassica napus Guard Cells in Response to Low CO2. Front. Mol. Biosci. 2017, 4, 51. [Google Scholar] [CrossRef] [Green Version]
- Zúñiga-Sánchez, E.; Soriano, D.; Martínez-Barajas, E.; Orozco-Segovia, A.; Gamboa-Debuen, A. BIIDXI, the At4g32460 DUF642 gene, is involved in pectin methyl esterase regulation during Arabidopsis thaliana seed germination and plant development. BMC Plant Biol. 2014, 14, 338. [Google Scholar] [CrossRef] [Green Version]
- Amsbury, S.; Hunt, L.; Elhaddad, N.; Baillie, A.; Lundgren, M.R.; Verhertbruggen, Y.; Scheller, H.V.; Knox, J.P.; Fleming, A.J.; Gray, J.E. Stomatal Function Requires Pectin De-methyl-esterification of the Guard Cell Wall. Curr. Biol. 2016, 26, 2899–2906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.S.; Kim, B.K.; Kwon, S.J.; Jin, H.C.; Park, O.K. Arabidopsis GDSL lipase 2 plays a role in pathogen defense via negative regulation of auxin signaling. Biochem. Biophys. Res. Commun. 2009, 379, 1038–1042. [Google Scholar] [CrossRef] [PubMed]
- Le, J.; Liu, X.; Yang, K.-Z.; Chen, X.-L.; Zou, J.-J.; Wang, H.-Z.; Wang, M.; Vanneste, S.; Morita, M.T.; Tasaka, M.; et al. Auxin transport and activity regulate stomatal patterning and development. Nat. Commun. 2014, 5, 3090. [Google Scholar] [CrossRef] [Green Version]
- Bauly, J.M. Overexpression of Auxin-Binding Protein Enhances the Sensitivity of Guard Cells to Auxin. Plant Physiol. 2000, 124, 1229–1238. [Google Scholar] [CrossRef] [Green Version]
- Dib, L.; Hayek, P.; Sadek, H.; Beyrouthy, B.; Khalaf, R.A. The Candida albicans Ddr48 protein is essential for filamentation, stress response, and confers partial antifungal drug resistance. Med. Sci. Monit. 2008, 14, 113–121. [Google Scholar]
- Gao, F.; Duan, X.; Yue, G.; Yang, A.; Zhang, J.-R. Cloning of an H+-PPase gene from Thellungiella halophila and its heterologous expression to improve tobacco salt tolerance. J. Exp. Bot. 2006, 57, 3259–3270. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Gao, F.; Zhao, L.; Li, K.; Zhang, J.-R. Identification of a new 130 bp cis-acting element in the TsVP1 promoter involved in the salt stress response from Thellungiella halophila. BMC Plant Biol. 2010, 10, 90. [Google Scholar] [CrossRef] [Green Version]
- Maeshima, M. Vacuolar H+-pyrophosphatase. Biochim. Biophys. Acta Biomembr. 2000, 1465, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Li, S.; Yu, D. Identification of an Arabidopsis Nodulin-related protein in heat stress. Mol. Cells 2009, 29, 77–84. [Google Scholar] [CrossRef]
- Arondel, V.; Vergnolle, C.; Cantrel, C.; Kader, J.-C. Lipid transfer proteins are encoded by a small multigene family in Arabidopsis thaliana. Plant Sci. 2000, 157, 1–12. [Google Scholar] [CrossRef]
- Hairat, S.; Baranwal, V.K.; Khurana, P. Identification of Triticum aestivum nsLTPs and functional validation of two members in development and stress mitigation roles. Plant Physiol. Biochem. 2018, 130, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Li, J.; Jiao, L.; Li, C.; Zhu, D.; Yu, J. A Non-specific Setaria italica Lipid Transfer Protein Gene Plays a Critical Role under Abiotic Stress. Front. Plant Sci. 2016, 7, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnuma, T.; Numata, T.; Osawa, T.; Mizuhara, M.; Lampela, O.; Juffer, A.; Skriver, K.; Fukamizo, T. A class V chitinase from Arabidopsis thaliana: Gene responses, enzymatic properties, and crystallographic analysis. Planta 2011, 234, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.; Kim, S.-H.; Jung, M.-S.; Kim, M.-S.; Oh, J.-E.; Ju, H.-W.; Kim, K.-I.; Vierling, E.; Lee, H.; Hong, S.-W. Arabidopsis hot2 encodes an endochitinase-like protein that is essential for tolerance to heat, salt and drought stresses. Plant J. 2006, 49, 184–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y. Oligogalacturonic Acid and Chitosan Reduce Stomatal Aperture by Inducing the Evolution of Reactive Oxygen Species from Guard Cells of Tomato and Commelina communis. Plant Physiol. 1999, 121, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidangmayum, A.; Dwivedi, P.; Katiyar, D.; Hemantaranjan, A. Application of chitosan on plant responses with special reference to abiotic stress. Physiol. Mol. Biol. Plants 2019, 25, 313–326. [Google Scholar] [CrossRef]
- De Tullio, M.; Guether, M.; Balestrini, R. Ascorbate oxidase is the potential conductor of a symphony of signaling pathways. Plant Signal. Behav. 2013, 8, 23213. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Chi, Y.; Jiang, Z.; Xu, Y.; Xie, L.; Huang, F.; Wan, D.; Ni, J.; Yuan, F.; Wu, X.; et al. Hydrogen peroxide sensor HPCA1 is an LRR receptor kinase in Arabidopsis. Nature 2020. [Google Scholar] [CrossRef]
- Iwai, S.; Ogata, S.; Yamada, N.; Onjo, M.; Sonoike, K.; Shimazaki, K.-I. Guard cell photosynthesis is crucial in abscisic acid-induced stomatal closure. Plant Direct. 2019, 3, e00137. [Google Scholar] [CrossRef] [Green Version]
- Tyerman, S.D.; Munns, R.; Fricke, W.; Arsova, B.; Barkla, B.J.; Bose, J.; Bramley, H.; Byrt, C.; Chen, Z.; Colmer, T.D.; et al. Energy costs of salinity tolerance in crop plants. New Phytol. 2019, 221, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, Y.; Ebisu, Y.; Kinoshita, T.; Doi, M.; Okuma, E.; Murata, Y.; Shimazaki, K.-I. bHLH Transcription Factors That Facilitate K+ Uptake During Stomatal Opening Are Repressed by Abscisic Acid Through Phosphorylation. Sci. Signal 2013, 6, ra48. [Google Scholar] [CrossRef] [PubMed]
- Leonhardt, N.; Kwak, J.M.; Robert, N.; Waner, D.; Leonhardt, G.; Schroeder, J.I. Microarray Expression Analyses of Arabidopsis Guard Cells and Isolation of a Recessive Abscisic Acid Hypersensitive Protein Phosphatase 2C Mutant. Plant Cell 2004, 16, 596–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, X.; Xiong, W.; Ye, T.; Wu, Y. Overexpression of the aspartic protease ASPG1 gene confers drought avoidance in Arabidopsis. J. Exp. Bot. 2012, 63, 2579–2593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rui, Y.; Chen, Y.; Kandemir, B.; Yi, H.; Wang, J.Z.; Puri, V.M.; Anderson, C.T. Balancing Strength and Flexibility: How the Synthesis, Organization, and Modification of Guard Cell Walls Govern Stomatal Development and Dynamics. Front. Plant Sci. 2018, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 5, 771. [Google Scholar] [CrossRef] [Green Version]
- Ringli, C.; Keller, B.; Ryser, U. Glycine-rich proteins as structural components of plant cell walls. Cell. Mol. Life Sci. 2001, 58, 1430–1441. [Google Scholar] [CrossRef]
- Shabala, S.; Mackay, A.; Tian, Y.; Jacobsen, S.-E.; Zhou, D.; Shabala, S. Oxidative stress protection and stomatal patterning as components of salinity tolerance mechanism in quinoa (Chenopodium quinoa). Physiol. Plant 2012, 146, 26–38. [Google Scholar] [CrossRef]
- Lambou, K.; Pennati, A.; Valsecchi, I.; Tada, R.; Sherman, S.; Sato, H.; Beau, R.; Gadda, G.; Latgé, J.-P. Pathway of Glycine Betaine Biosynthesis in Aspergillus fumigatus. Eukaryot. Cell 2013, 12, 853–863. [Google Scholar] [CrossRef] [Green Version]
- Catusse, J.; Strub, J.-M.; Job, C.; Van Dorsselaer, A.; Job, D. Proteome-wide characterization of sugarbeet seed vigor and its tissue specific expression. Proc. Natl. Acad. Sci. USA 2008, 105, 10262–10267. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Kurepa, J.; Hashimoto, T.; Smalle, J.A. Salt Stress–Induced Disassembly of Arabidopsis Cortical Microtubule Arrays Involves 26S Proteasome–Dependent Degradation of SPIRAL1. Plant Cell 2011, 23, 3412–3427. [Google Scholar] [CrossRef] [Green Version]
- Ho, L.-W.; Yang, T.-T.; Shieh, S.-S.; Edwards, G.E.; Yen, H.E. Reduced expression of a vesicle trafficking-related ATPase SKD1 decreases salt tolerance in Arabidopsis. Funct. Plant Biol. 2010, 37, 962–973. [Google Scholar] [CrossRef]
- Ritte, G.; Rosenfeld, J.; Rohrig, K.; Raschke, K. Rates of Sugar Uptake by Guard Cell Protoplasts of Pisum sativum L. Related to the Solute Requirement for Stomatal Opening. Plant Physiol. 1999, 121, 647–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol Cell Proteomics 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.S.; ElSayed, A.I.; Moore, M.; Dietz, K.J. Redox and reactive oxygen species network in acclimation for salinity tolerance in sugar beet. J. Exp. Bot. 2017, 68, 1283–1298. [Google Scholar] [CrossRef] [Green Version]
- Schwacke, R.; Ponce-Soto, G.Y.; Krause, K.; Bolger, A.M.; Arsova, B.; Hallab, A.; Gruden, K.; Stitt, M.; Bolger, M.E.; Usadel, B. MapMan4: A Refined Protein Classification and Annotation Framework Applicable to Multi-Omics Data Analysis. Mol. Plant 2019, 12, 879–892. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Accession Number | Log2 LFQ Intensity (Control) | Log2 LFQ Intensity (Salt Stress) | Unique Peptides | Fold Change | t-Test q Value | Annotation |
|---|---|---|---|---|---|---|---|
| 1 | XP_010683515.1 | 23.1 | 29.0 | 4 | 60.9 | 2.4 × 10−4 | non-specific lipid-transfer protein |
| 2 | XP_010690044.1 | 23.1 | 28.6 | 8 | 45.0 | 7.3 × 10−4 | inorganic pyrophosphatase 2 |
| 3 | XP_010680759.1 | 23.9 | 27.3 | 7 | 10.3 | 8.5 × 10−3 | chitotriosidase-1 |
| 4 | XP_010679746.1 | 25.2 | 28.5 | 14 | 10.2 | 4.2 × 10−3 | peroxidase 27 |
| 5 | XP_010679767.1 | 26.0 | 28.9 | 14 | 7.5 | 1.3 × 10−4 | mannan endo-1,4-beta-mannosidase 7 |
| 6 | XP_010686587.1 | 28.0 | 30.7 | 24 | 6.6 | 3.0 × 10−4 | phosphoethanolamine N-methyltransferase |
| 7 | XP_010676511.1 | 22.6 | 25.1 | 4 | 5.7 | 3.3 × 10−2 | thaumatin-like protein 1 |
| 8 | XP_010679501.1 | 26.9 | 29.4 | 2 | 5.3 | 4.7 × 10−4 | glycine-rich cell wall structural protein |
| 9 | KMT11925.1 | 27.0 | 29.4 | 16 | 5.3 | 1.3 × 10−4 | endoglucanase 6 |
| 10 | XP_010687451.1 | 23.0 | 25.4 | 2 | 5.2 | 2.8 × 10−2 | endochitinase EP3 |
| 11 | XP_010678753.1 | 26.5 | 23.1 | 3 | −10.25 | 3.2 × 10−2 | heat shock 70 kDa protein 18-like |
| 12 | XP_010677928.1 | 26.5 | 23.8 | 8 | −6.51 | 3.0 × 10−3 | malate synthase, glyoxysomal |
| 13 | XP_010686038.1 | 27.1 | 24.6 | 2 | -5.72 | 3.1 × 10−2 | protein suppressor of K+ transport growth defect1 |
| 14 | KMT14555.1 | 30.2 | 27.8 | 6 | -5.39 | 2.8 × 10−3 | glucan endo-1,3-beta-glucosidase, acidic-like |
| 15 | XP_010676519.1 | 30.3 | 27.8 | 8 | -5.32 | 5.9 × 10−4 | alpha-amylase/trypsin inhibitor |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasouli, F.; Kiani-Pouya, A.; Li, L.; Zhang, H.; Chen, Z.; Hedrich, R.; Wilson, R.; Shabala, S. Sugar Beet (Beta vulgaris) Guard Cells Responses to Salinity Stress: A Proteomic Analysis. Int. J. Mol. Sci. 2020, 21, 2331. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072331
Rasouli F, Kiani-Pouya A, Li L, Zhang H, Chen Z, Hedrich R, Wilson R, Shabala S. Sugar Beet (Beta vulgaris) Guard Cells Responses to Salinity Stress: A Proteomic Analysis. International Journal of Molecular Sciences. 2020; 21(7):2331. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072331
Chicago/Turabian StyleRasouli, Fatemeh, Ali Kiani-Pouya, Leiting Li, Heng Zhang, Zhonghua Chen, Rainer Hedrich, Richard Wilson, and Sergey Shabala. 2020. "Sugar Beet (Beta vulgaris) Guard Cells Responses to Salinity Stress: A Proteomic Analysis" International Journal of Molecular Sciences 21, no. 7: 2331. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072331