Myocardium Metabolism in Physiological and Pathophysiological States: Implications of Epicardial Adipose Tissue and Potential Therapeutic Targets

Abstract
:1. The Metabolism of Cardiomyocytes in Physiological Condition
1.1. Excitation-Contraction of Cardiomyocytes
1.2. Energy and Substrates
1.2.1. Fatty Acids
1.2.2. Glucose
1.2.3. Ketone Bodies
1.3. Diet and Physical Activity
1.4. Circadian Rhythm
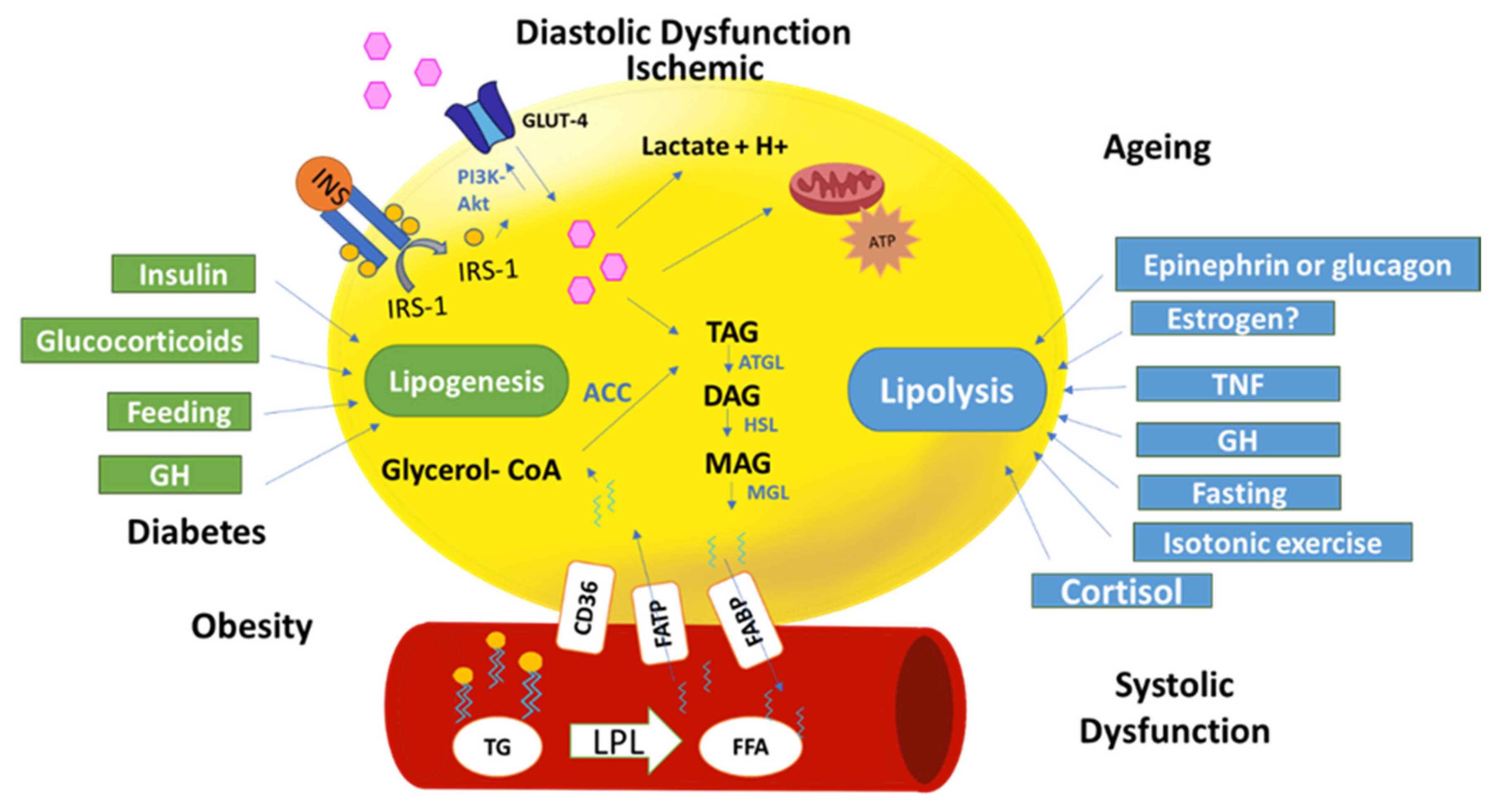
1.5. Hormonal Regulation
1.5.1. Insulin
1.5.2. Thyroid Hormones
1.5.3. Insulin Growth Factor 1 (IGF-1)
1.5.4. Growth Hormone
1.5.5. Estrogen
1.5.6. Cortisol
1.5.7. GLP-1
1.6. Ageing
2. The Metabolism of Cardiomyocytes in Pathological Conditions
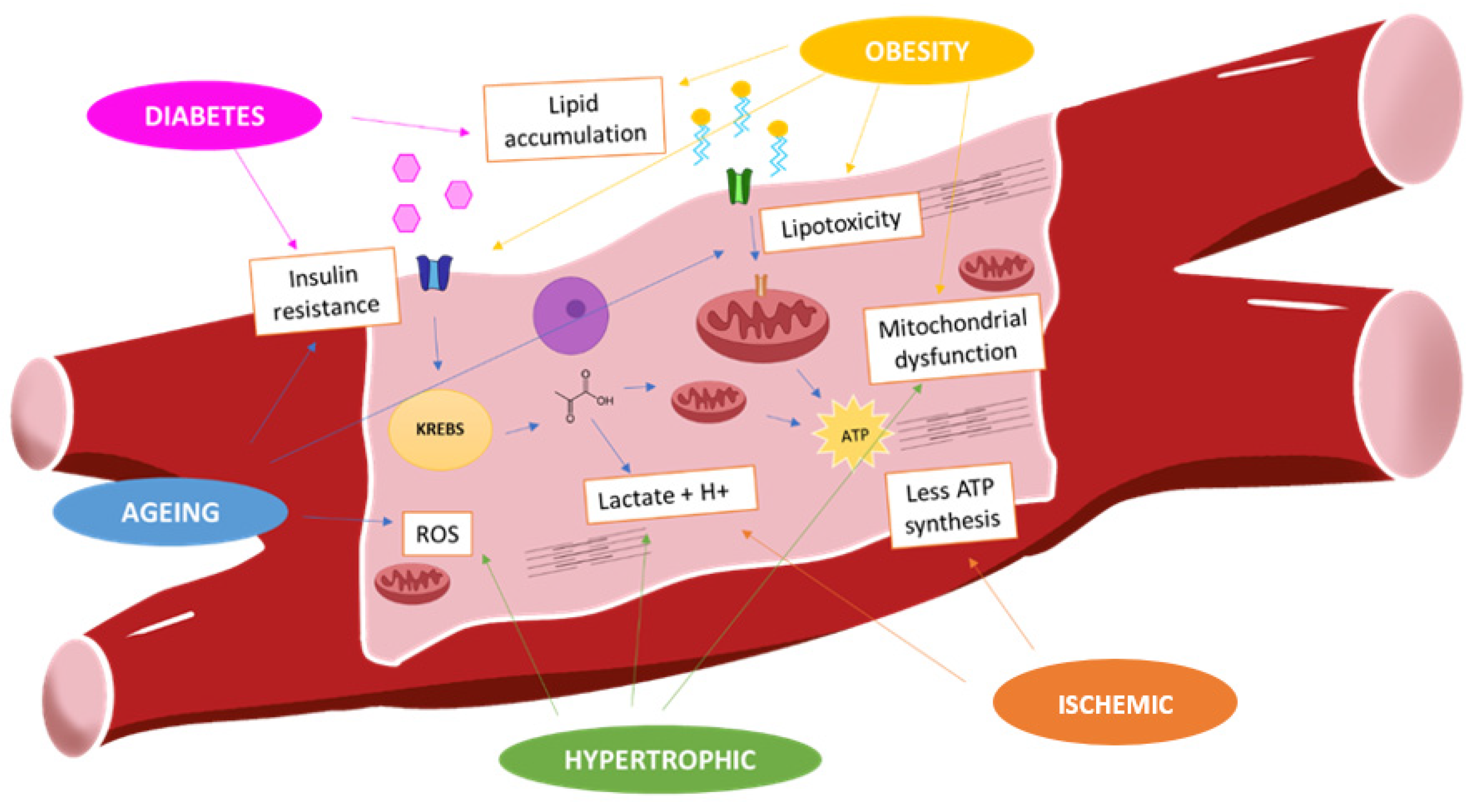
2.1. “Obesity” Cardiomyocytes and Metabolism
2.2. “Diabetes” Cardiomyocytes and Metabolism
2.3. Ischemic Cardiomyocytes and Metabolism
2.4. Hypertrophic Cardiomyocytes and Metabolism
3. Epicardial Adipose Tissue Metabolism
4. Hormonal Therapy on Cardiac Metabolism
4.1. Estrogen Therapy
4.2. GLP-1 Receptor Agonists
5. Solutions and Nutritional Therapies on Metabolic Signaling
5.1. Polarizing Solutions
5.2. Coenzyme Q10
5.3. L-carnitine
6. Others Metabolic Therapies
6.1. Trimetazidine
6.2. SGLT2 Inhibitors
6.3. Metformin
7. Conclusions
Funding
Conflicts of Interest
Abbreviations
| ATP | adenosine triphosphate |
| ATPase | adenosine triphosphatase |
| NCX | sarcolemmal Na+/Ca2+ exchanger |
| FFA | Free fatty acids |
| FADH | Flavin adenine dinucleotide |
| NADH | Nicotinamide adenine dinucleotide (NAD) + hydrogen (H) |
| GLUT | Glucose transporter |
| LDH | lactate dehydrogenase 4 |
| BMAL1 | Brain and muscle Arnt-like protein-1 |
| AMPK | adenosine monophosphate-activated protein kinase |
| IGF-1 | insulin growth factor |
| GLP1 | glucagon like peptide 1 |
| SGLT2 | sodium glucose transporter 2 |
References
- Niggli, E.; Lederer, W.J. Voltage-independent calcium release in heart muscle. Science 1990, 250, 565. [Google Scholar] [CrossRef] [PubMed]
- Bers, D.M. Cardiac excitation-contraction coupling. Nature 2002, 415, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Washio, T.; Shintani, S.A.; Higuchi, H.; Sugiura, S.; Hisada, T. Effect of myofibril passive elastic properties on the mechanical communication between motor proteins on adjacent sarcomeres. Sci. Rep. 2019, 9, 9355. [Google Scholar] [CrossRef] [PubMed]
- van der Vusse, G.J.; Glatz, J.F.; Stam, H.C.; Reneman, R.S. Fatty acid homeostasis in the normoxic and ischemic heart. Physiol. Rev. 1992, 72, 881–940. [Google Scholar] [CrossRef]
- Randle, P.J.; Garland, P.B.; Hales, C.N.; Newsholme, E.A. The glucose fatty-acid cycle its role in insulin sensitivity and the metabolic disturbances of diabetes mellttus. Lancet 1963, 281, 785–789. [Google Scholar] [CrossRef]
- Burkhoff, D.; Weiss, R.G.; Schulman, S.P.; Kalil-Filho, R.; Wannenburg, T.; Gerstenblith, G. Influence of metabolic substrate on rat heart function and metabolism at different coronary flows. Am. J. Physiol. Circ. Physiol. 1991, 261, H741–H750. [Google Scholar] [CrossRef] [Green Version]
- Korvald, C.; Elvenes, O.P.; Myrmel, T. Myocardial substrate metabolism influences left ventricular energetics in vivo. Am. J. Physiol. Hear. Circ. Physiol. 2000, 278, 1345–1351. [Google Scholar] [CrossRef] [Green Version]
- Sorrentino, D.; Stump, D.; Potter, B.J.; Robinson, R.B.; White, R.; Kiang, C.L.; Berk, P.D. Oleate uptake by cardiac myocytes is carrier mediated and involves a 40-kD plasma membrane fatty acid binding protein similar to that in liver, adipose tissue, and gut. J. Clin. Investig. 1988, 82, 928–935. [Google Scholar] [CrossRef] [Green Version]
- Turcotte, L.P.; Swenberger, J.R.; Tucker, M.Z.; Yee, A.J.; Trump, G.; Luiken, J.J.; Bonen, A. Muscle palmitate uptake and binding are saturable and inhibited by antibodies to FABPPM. Mol. Cell. Biochem. 2000, 210, 53–63. [Google Scholar] [CrossRef]
- Luiken, J.J.F.P.; Turcotte, L.P.; Bonen, A. Protein-mediated palmitate uptake and expression of fatty acid transport proteins in heart giant vesicles. J. Lipid Res. 1999, 40, 1007–1016. [Google Scholar]
- Nickerson, J.G.; Alkhateeb, H.; Benton, C.R.; Lally, J.; Nickerson, J.; Han, X.X.; Wilson, M.H.; Jain, S.S.; Snook, L.A.; Glatz, J.F.; et al. Greater transport efficiencies of the membrane fatty acid transporters FAT/CD36 and FATP4 compared with FABPpm and FATP1 and differential effects on fatty acid esterification and oxidation in rat skeletal muscle. J. Biol. Chem. 2009, 284, 16522–16530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmann, J.F.F.; Abumrad, N.A.; Ibrahimi, A.; van der Vusse, G.J.; Glatz, J.F.C. New insights into long-chain fatty acid uptake by heart muscle: A crucial role for fatty acid translocase/CD36. Biochem. J. 2002, 367, 561–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depre, C.; Vanoverschelde, J.L.J.; Taegtmeyer, H. Glucose for the Heart. Circulation 1999, 99, 578–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisneski, J.A.; Gertz, E.W.; Neese, R.A.; Gruenke, L.D.; Craig, J.C. Dual carbon-labeled isotope experiments using D-[6-14C] glucose and L-[1,2,3-13C3] lactate: A new approach for investigating human myocardial metabolism during ischemia. J. Am. Coll. Cardiol. 1985, 5, 1138–1146. [Google Scholar] [CrossRef] [Green Version]
- Wisneski, J.A.; Gertz, E.W.; Neese, R.A.; Gruenke, L.D.; Morris, D.L.; Craig, J.C. Metabolic fate of extracted glucose in normal human myocardium. J. Clin. Investig. 1985, 76, 1819–1827. [Google Scholar] [CrossRef] [PubMed]
- Wisneski, J.A.; Gertz, E.W.; Neese, R.A.; Mayr, M. Myocardial metabolism of free fatty acids. Studies with 14C-labeled substrates in humans. J. Clin. Investig. 1987, 79, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Kolwicz, S.C.J. An Exercise in Cardiac Metabolism. Front Cardiovasc Med. 2018, 5, 66. [Google Scholar] [CrossRef]
- Doenst, T.; Nguyen, T.D.; Abel, E.D. Cardiac metabolism in heart failure: Implications beyond atp production. Circ. Res. 2013, 113, 709–724. [Google Scholar] [CrossRef] [Green Version]
- Nakano, H.; Minami, I.; Braas, D.; Pappoe, H.; Wu, X.; Sagadevan, A.; Vergnes, L.; Fu, K.; Morselli, M.; Dunham, C.; et al. Glucose inhibits cardiac muscle maturation through nucleotide biosynthesis. Elife 2017, 6, e29330. [Google Scholar] [CrossRef]
- Galvez, A.S.; Ulloa, J.A.; Chiong, M.; Criollo, A.; Eisner, V.; Barros, L.F.; Lavandero, S. Aldose Reductase Induced by Hyperosmotic Stress Mediates Cardiomyocyte Apoptosis: Differential effects of sorbitol and mannitol. J. Biol. Chem. 2003, 278, 38484–38494. [Google Scholar] [CrossRef] [Green Version]
- Nuutila, P.; Koivisto, V.A.; Knuuti, J.; Ruotsalainen, U.; Teräs, M.; Haaparanta, M.; Bergman, J.; Solin, O.; Voipio-Pulkki, L.M.; Wegelius, U. Glucose-free fatty acid cycle operates in human heart and skeletal muscle in vivo. J. Clin. Investig. 1992, 89, 1767–1774. [Google Scholar] [CrossRef] [PubMed]
- Taegtmeyer, H. Energy metabolism of the heart: From basic concepts to clinical applications applications. Curr. Probl. Cardiol. 1994, 19, 61–113. [Google Scholar] [CrossRef]
- Pessin, J.E.; Bell, G.I. Mammalian Facilitative Glucose Transporter Family: Structure and Molecular Regulation. Annu. Rev. Physiol. 1992, 54, 911–930. [Google Scholar] [CrossRef] [PubMed]
- Gould, G.W.; Holman, D.G. The glucose transporter family: Structure, function and tissue-specific expression. Biochem. J. 1993, 295, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Gertz, E.W.; Wisneski, J.A.; Stanley, W.C.; Neese, A.R. Myocardial substrate utilization during exercise in humans. Dual carbon-labeled carbohydrate isotope experiments. J. Clin. Investig. 1988, 82, 2017–2025. [Google Scholar] [CrossRef]
- Gay, R.J.; McComb, R.B.; Bowers, G.N., Jr. Optimum Reaction Conditions for Human Lactate Dehydrogenase Isoenzymes as They Affect Total Lactate Dehydrogenase Activity. Clin. Chem. 2020, 14, 740–753. [Google Scholar] [CrossRef]
- Burelle, Y.; Wambolt, R.B.; Grist, M.; Parsons, H.L.; Chow, J.C.; Antler, C.; Bonen, A.; Keller, A.; Dunaway, G.A.; Popov, K.M.; et al. Regular exercise is associated with a protective metabolic phenotype in the rat heart. Am. J. Physiol. Circ. Physiol. 2004, 287, H1055–H1063. [Google Scholar] [CrossRef]
- McMullen, J.R.; Jennings, G.L. Difference between pathological and physiological cardiac hypertrophy: Novel therapeutic startegies to treat heart failure. Clin. Exp. Pharmacol. Physiol. 2007, 34, 255–262. [Google Scholar] [CrossRef]
- Vega, R.B.; Konhilas, J.P.; Kelly, D.P.; Leinwand, L.A. Molecular Mechanisms Underlying Cardiac Adaptation to Exercise. Cell Metabol. 2017, 25, 1012–1026. [Google Scholar] [CrossRef]
- Rabinovich-Nikitin, I.; Lieberman, B.; Martino, T.A.; Kirshenbaum, L.A. Circadian-Regulated Cell Death in Cardiovascular Diseases. Circulation 2019, 139, 965–980. [Google Scholar] [CrossRef]
- Takeda, N.; Maemura, K. Circadian clock and cardiovascular disease. J. Cardiol. 2011, 57, 249–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Yang, G. Recent advances in circadian rhythms in cardiovascular system. Front. Pharmacol. 2015, 6, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durgan, D.J.; Hotze, M.A.; Tomlin, T.M.; Egbejimi, O.; Graveleau, C.; Abel, E.D.; Shaw, C.A.; Bray, M.S.; Hardin, P.E.; Young, M.E. The intrinsic circadian clock within the cardiomyocyte. Am. J. Physiol. Hear. Circ. Physiol. 2005, 289, 1530–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, M.E. Temporal partitioning of cardiac metabolism by the cardiomyocyte circadian clock. Exp. Physiol. 2016, 101, 1035–1039. [Google Scholar] [CrossRef] [Green Version]
- Lamia, K.A.; Sachdeva, U.M.; DiTacchio, L.; Williams, E.C.; Alvarez, J.G.; Egan, D.F.; Vasquez, D.S.; Juguilon, H.; Panda, S.; Shaw, R.J.; et al. AMPK Regulates the Circadian Clock by Cryptochrome Phosphorylation and Degradation. Science 2009, 326, 437–440. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Li, S.; Liu, T.; Borjigin, J.; Lin, J.D. Transcriptional coactivator PGC-1α integrates the mammalian clock and energy metabolism. Nature 2007, 447, 477–481. [Google Scholar] [CrossRef]
- Froy, O. The relationship between nutrition and circadian rhythms in mammals. Front. Neuroendocrinol. 2007, 28, 61–71. [Google Scholar] [CrossRef]
- Durgan, D.J.; Pat, B.M.; Laczy, B.; Bradley, J.A.; Tsai, J.Y.; Grenett, M.H.; Ratcliffe, W.F.; Brewer, R.A.; Nagendran, J.; Villegas-Montoya, C.; et al. O-GlcNAcylation, novel post-translational modification linking myocardial metabolism and cardiomyocyte circadian clock. J. Biol. Chem. 2011, 286, 44606–44619. [Google Scholar] [CrossRef] [Green Version]
- Evans, G. The glycogen content of the rat heart. J. Physiol. 1934, 82, 468–480. [Google Scholar] [CrossRef]
- Stowe, K.A.; Burgess, S.C.; Merritt, M.; Sherry, A.D.; Malloy, C.R. Storage and oxidation of long-chain fatty acids in the C57/BL6 mouse heart as measured by NMR spectroscopy. FEBS Lett. 2006, 580, 4282–4287. [Google Scholar] [CrossRef] [Green Version]
- McGinnis, G.R.; Young, M.E. Circadian regulation of metabolic homeostasis: Causes and consequences. Nat. Sci. Sleep 2016, 8, 163–180. [Google Scholar] [CrossRef] [Green Version]
- Young, M.E.; Peter, R.; Cedars, A.M.; Heinrich, T. Intrinsic Diurnal Variations in Cardiac Metabolism and Contractile Function. Circ. Res. 2001, 89, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- van Cauter, E.; Polonsky, K.S.; Scheen, A.J. Roles of Circadian Rhythmicity and Sleep in Human Glucose Regulation*. Endocr. Rev. 1997, 18, 716–738. [Google Scholar] [CrossRef] [Green Version]
- van Cauter, E.; Kerkhofs, M.; Caufriez, A.; van Onderbergen, A.; Thorner, M.O.; Copinschi, G. A quantitative estimation of growth hormone secretion in normal man: Reproducibility and relation to sleep and time of day. J. Clin. Endocrinol. Metab. 1992, 74, 1441–1450. [Google Scholar] [CrossRef]
- Hower, I.M.; Harper, S.A.; Buford, T.W. Circadian rhythms, exercise, and cardiovascular health. J. Circ. Rhythm. 2018, 16, 7. [Google Scholar] [CrossRef]
- Iliadis, F.; Kadoglou, N.; Didangelos, T. Insulin and the heart. Diabetes Res. Clin. Pract. 2011, 93, S86–S91. [Google Scholar] [CrossRef]
- Cahill, G.F. Physiology of Insulin in Man: The Banting Memorial Lecture 1971. Diabetes 1971, 20, 785–799. [Google Scholar] [CrossRef]
- Luiken, J.J.; Koonen, D.P.; Willems, J.; Zorzano, A.; Becker, C.; Fischer, Y.; Tandon, N.N.; Van Der Vusse, G.J.; Bonen, A.; Glatz, J.F. Insulin Stimulates Long-Chain Fatty Acid Utilization by Rat Cardiac Myocytes Through Cellular Redistribution of FAT/CD36. Diabetes 2002, 51, 3113–3119. [Google Scholar] [CrossRef] [Green Version]
- Randle, P.J.; Garland, P.B.; Hales, C.N.; Newsholme, E.A.; Denton, R.M.; Pogson, C.I. Interactions of Metabolism and the Physiological Role of Insulin11This work was supported by grants from the British Diabetic Association, The British Insulin Manufacturers, The Medical Research Council, and the Royal Society. Recent Prog. Horm. Res. 1966, 22, 1–48. [Google Scholar]
- Borensztajn, J.; Robinson, D.S. The effect of fasting on the utilization of chylomicron triglyceride fatty acids in relation to clearing factor lipase (lipoprotein lipase) releasable by heparin in the perfused rat heart. J. Lipid Res. 1970, 11, 111–117. [Google Scholar]
- Pederson, T.M.; Kramer, D.L.; Rondinone, C.M. Serine/Threonine Phosphorylation of IRS-1 Triggers Its Degradation. Diabetes 2001, 50, 24–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odegaard, J.I.; Chawla, A. Pleiotropic actions of insulin resistance and inflammation in metabolic homeostasis. Science 2013, 339, 172–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sears, B.; Perry, M. The role of fatty acids in insulin resistance. Lipids Health Dis. 2015, 14, 121. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, L.; Horman, S.; Beauloye, C.; Vanoverschelde, J.L. Insulin signalling in the heart. Cardiovasc. Res. 2008, 79, 238–248. [Google Scholar] [CrossRef] [Green Version]
- Rutigliano, G.; Zucchi, R. Cardiac actions of thyroid hormone metabolites. Mol. Cell. Endocrinol. 2017, 458, 76–81. [Google Scholar] [CrossRef]
- Kahaly, G.J.; Dillmann, W.H. Thyroid hormone action in the heart. Endocr. Rev. 2005, 26, 704–728. [Google Scholar] [CrossRef] [Green Version]
- Cumero, S.; Fogolari, F.; Domenis, R.; Zucchi, R.; Mavelli, I.; Contessi, S. Mitochondrial F0F1-ATP synthase is a molecular target of 3-iodothyronamine, an endogenous metabolite of thyroid hormone. Br. J. Pharmacol. 2012, 166, 2331–2347. [Google Scholar] [CrossRef] [Green Version]
- Cook, G.A.; Edwards, T.L.; Jansen, M.S.; Bahouth, S.W.; Wilcox, H.G.; Park, E.A. Differential Regulation of Carnitine Palmitoyltransferase-I Gene Isoforms (CPT-I α and CPT-Iβ) in the Rat Heart. J. Mol. Cell. Cardiol. 2001, 33, 317–329. [Google Scholar] [CrossRef]
- Orfali, K.A.; Fryer, L.G.D.; Holness, M.J.; Sugden, M.C. Interactive effects of insulin and triiodothyronine on pyruvate dehydrogenase kinase activity in cardiac myocytes. J. Mol. Cell. Cardiol. 1995, 27, 901–908. [Google Scholar] [CrossRef]
- Schwartz, V.; Lue, H.; Kraemer, S.; Korbiel, J.; Krohn, R.; Ohl, K.; Bucala, R.; Weber, C.; Bernhagen, J. A functional heteromeric MIF receptor formed by CD74 and CXCR4. FEBS Lett. 2009, 583, 2749–2757. [Google Scholar] [CrossRef] [Green Version]
- Portman, M.A. Thyroid Hormone Regulation of Heart Metabolism. Thyroid 2008, 18, 217–225. [Google Scholar] [CrossRef]
- de Meyts, P.; Whittaker, J. Structural biology of insulin and IGF1 receptors: Implications for drug design. Nat. Rev. Drug Discov. 2002, 1, 769–783. [Google Scholar] [CrossRef]
- DeBosch, B.J.; Muslin, A.J. Insulin signaling pathways and cardiac growth. J. Mol. Cell. Cardiol. 2008, 44, 855–864. [Google Scholar] [CrossRef] [Green Version]
- Clemmons, D.R. The relative roles of growth hormone and IGF-1 in controlling insulin sensitivity. J. Clin. Investig. 2004, 113, 25–27. [Google Scholar] [CrossRef] [Green Version]
- Yakar, S.; Setser, J.; Zhao, H.; Stannard, B.; Haluzik, M.; Glatt, V.; Bouxsein, M.L.; Kopchick, J.J.; LeRoith, D. Inhibition of growth hormone action improves insulin sensitivity in liver IGF-1-deficient mice. J. Clin. Investig. 2004, 113, 96–105. [Google Scholar] [CrossRef]
- Pires, K.M.; Buffolo, M.; Schaaf, C.; David Symons, J.; Cox, J.; Abel, E.D.; Selzman, C.H.; Boudina, S. Activation of IGF-1 receptors and Akt signaling by systemic hyperinsulinemia contributes to cardiac hypertrophy but does not regulate cardiac autophagy in obese diabetic mice. J. Mol. Cell. Cardiol. 2017, 113, 39–50. [Google Scholar] [CrossRef]
- Colao, A.; Marzullo, P.; di Somma, C.; Lombardi, G. Growth hormone and the heart. Clin. Endocrinol. 2001, 54, 137–154. [Google Scholar] [CrossRef]
- Piccioli, L.; Arcopinto, M.; Salzano, A.; D’Assante, R.; Schiavo, A.; Stagnaro, F.M.; Lombardi, A.; Panicara, V.; Valente, P.; Vitale, G.; et al. The impairment of the Growth Hormone/Insulin-like growth factor 1 (IGF-1) axis in heart failure: A possible target for future therapy. Mon. Arch. Chest Dis. 2018, 88, 20–25. [Google Scholar] [CrossRef]
- Lu, C.; Schwartzbauer, G.; Sperling, M.A.; Devaskar, S.U.; Thamotharan, S.; Robbins, P.D.; McTiernan, C.F.; Liu, J.L.; Jiang, J.; Frank, S.J.; et al. Demonstration of Direct Effects of Growth Hormone on Neonatal Cardiomyocytes. J. Biol. Chem. 2001, 276, 22892–22900. [Google Scholar] [CrossRef] [Green Version]
- Barros, R.P.A.; Gustafsson, J.Å. Estrogen receptors and the metabolic network. Cell Metabol. 2011, 14, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Knowlton, A.A.; Korzick, D.H. Estrogen and the female heart. Mol. Cell. Endocrinol. 2014, 389, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Sbert-Roig, M.; Bauzá-Thorbrügge, M.; Galmés-Pascual, B.M.; Capllonch-Amer, G.; García-Palmer, F.J.; Lladó, I.; Proenza, A.M.; Gianotti, M. GPER mediates the effects of 17β-estradiol in cardiac mitochondrial biogenesis and function. Mol. Cell. Endocrinol. 2016, 420, 116–124. [Google Scholar] [CrossRef]
- Deschamps, A.M.; Murphy, E. Activation of a novel estrogen receptor, GPER, is cardioprotective in male and female rats. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1806–H1813. [Google Scholar] [CrossRef] [Green Version]
- Clegg, D.J.; Brown, L.M.; Woods, S.C.; Benoit, S.C. Gonadal Hormones Determine Sensitivity to Central Leptin and Insulin. Diabetes 2006, 55, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Jeong, I.-K. The role of cortisol in the pathogenesis of the metabolic syndrome. Diabetes Metab. J. 2012, 36, 207–210. [Google Scholar] [CrossRef]
- Fraser, R.; Ingram, M.C.; Anderson, N.H.; Morrison, C.; Davies, E.; Connell, J.M.C. Cortisol Effects on Body Mass, Blood Pressure, and Cholesterol in the General Population Data Analyses. Hypertension 1999, 33, 1364–1368. [Google Scholar] [CrossRef] [Green Version]
- Walejko, J.M.; Antolic, A.; Koelmel, J.P.; Garrett, T.J.; Edison, A.S.; Keller-Wood, M. Chronic maternal cortisol excess during late gestation leads to metabolic alterations in the newborn heart. Am. J. Physiol. Metab. 2019, 316, E546–E556. [Google Scholar] [CrossRef]
- Pedersen, S.B.; Jønler, M.; Richelsen, B. Characterization of regional and gender differences in glucocorticoid receptors and lipoprotein lipase activity in human adipose tissue. J. Clin. Endocrinol. Metab. 1994, 78, 1354–1359. [Google Scholar] [CrossRef]
- Fain, J.N.; Kovacev, V.P.; Scow, R.O. Effect of Growth Hormone and Dexamethasone on Lipolysis and Metabolism in Isolated Fat Cells of the Rat. J. Biol. Chem. 1965, 240, 3522–3529. [Google Scholar]
- Gathercole, L.L.; Morgan, S.A.; Bujalska, I.J.; Hauton, D.; Stewart, P.M.; Tomlinson, J.W. Regulation of lipogenesis by glucocorticoids and insulin in human adipose tissue. PLoS ONE 2011, 6, e26223. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-J.; Gong, D.-W.; Burkey, B.F.; Fried, S.K. Pathways regulated by glucocorticoids in omental and subcutaneous human adipose tissues: a microarray study. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E571–E580. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Shin, S.; Shigihara, T.; Hahm, E.; Liu, M.J.; Han, J.; Yoon, J.W.; Jun, H.S. Glucagon-Like Peptide-1 Gene Therapy in Obese Diabetic Mice Results in Long-Term Cure of Diabetes by Improving Insulin Sensitivity and Reducing Hepatic Gluconeogenesis. Diabetes 2007, 56, 1671–1679. [Google Scholar] [CrossRef] [Green Version]
- Drucker, D.J. The biology of incretin hormones. Cell Metab. 2006, 3, 153–165. [Google Scholar] [CrossRef] [Green Version]
- Baggio, L.L.; Yusta, B.; Mulvihill, E.E.; Cao, X.; Streutker, C.J.; Butany, J.; Cappola, T.P.; Margulies, K.B.; Drucker, D.J. GLP-1 Receptor Expression within the Human Heart. Endocrinology 2018, 159, 1570–1584. [Google Scholar] [CrossRef] [Green Version]
- Xiaolin, W. Glucagon-Like Peptide-1 Increases cAMP but Fails to Augment Contraction in Adult Rat Cardiac Myocytes. Circ. Res. 2001, 89, 445–452. [Google Scholar] [CrossRef] [Green Version]
- Tao, L.; Wang, L.; Yang, X.; Jiang, X.; Hua, F. Recombinant human glucagon-like peptide-1 protects against chronic intermittent hypoxia by improving myocardial energy metabolism and mitochondrial biogenesis. Mol. Cell. Endocrinol. 2019, 481, 95–103. [Google Scholar] [CrossRef]
- Decara, J.; Rivera, P.; Arrabal, S.; Vargas, A.; Serrano, A.; Pavón, F.J.; Dieguez, C.; Nogueiras, R.; Rodríguez de Fonseca, F.; Suárez, J. Cooperative role of the glucagon-like peptide-1 receptor and β3-adrenergic-mediated signalling on fat mass reduction through the downregulation of PKA/AKT/AMPK signalling in the adipose tissue and muscle of rats. Acta Physiol. 2018, 222, e13008. [Google Scholar] [CrossRef]
- Nikolaidis, L.A.; Elahi, D.; Hentosz, T.; Doverspike, A.; Huerbin, R.; Zourelias, L.; Stolarski, C.; Shen, Y.T.; Shannon, R.P. Recombinant Glucagon-Like Peptide-1 Increases Myocardial Glucose Uptake and Improves Left Ventricular Performance in Conscious Dogs with Pacing-Induced Dilated Cardiomyopathy. Circulation 2004, 110, 955–961. [Google Scholar] [CrossRef] [Green Version]
- Lesnefsky, E.J.; Chen, Q.; Hoppel, C.L. Mitochondrial Metabolism in Aging Heart. Circul. Res. 2016, 118, 1593–1611. [Google Scholar] [CrossRef] [Green Version]
- Koonen, D.P.; Febbraio, M.; Bonnet, S.; Nagendran, J.; Young, M.E.; Michelakis, E.D.; Dyck, J.R. CD36 Expression Contributes to Age-Induced Cardiomyopathy in Mice. Circulation 2007, 116, 2139–2147. [Google Scholar] [CrossRef] [Green Version]
- Drosatos, K.; Schulze, P.C. Cardiac lipotoxicity: Molecular pathways and therapeutic implications. Curr. Heart Fail. Rep. 2013, 10, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Hyyti, O.M.; Ledee, D.; Ning, X.-H.; Ge, M.; Portman, M.A. Aging impairs myocardial fatty acid and ketone oxidation and modifies cardiac functional and metabolic responses to insulin in mice. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H868–H875. [Google Scholar] [CrossRef] [Green Version]
- Chiao, Y.A.; Kolwicz, S.C.; Basisty, N.; Gagnidze, A.; Zhang, J.; Gu, H.; Djukovic, D.; Beyer, R.P.; Raftery, D.; MacCoss, M.; et al. Rapamycin transiently induces mitochondrial remodeling to reprogram energy metabolism in old hearts. Aging (Albany. NY). 2016, 8, 314–327. [Google Scholar] [CrossRef] [Green Version]
- Chuffa, L.G.; Seiva, F. Combined effects of age and diet-induced obesity on biochemical parameters and cardiac energy metabolism in rats. Indian J. Biochem. Biophys. 2013, 50, 40–47. [Google Scholar]
- Schüll, S.; Günther, S.D.; Brodesser, S.; Seeger, J.M.; Tosetti, B.; Wiegmann, K.; Pongratz, C.; Diaz, F.; Witt, A.; Andree, M.; et al. Cytochrome c oxidase deficiency accelerates mitochondrial apoptosis by activating ceramide synthase 6. Cell Death Dis. 2015, 6, e1691. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Asfour, H.; Bursac, N. Age-dependent functional crosstalk between cardiac fibroblasts and cardiomyocytes in a 3D engineered cardiac tissue. Acta Biomater. 2017, 55, 120–130. [Google Scholar] [CrossRef]
- Lönnqvist, F.; Nyberg, B.; Wahrenberg, H.; Arner, P. Catecholamine-induced lipolysis in adipose tissue of the elderly. J. Clin. Investig. 1990, 85, 1614–1621. [Google Scholar] [CrossRef] [Green Version]
- Toth, M.J.; Tchernof, A. Lipid metabolism in the elderly. Eur. J. Clin. Nutr. 2000, 54, S121–S125. [Google Scholar] [CrossRef]
- Silaghi, A.; Piercecchi-Marti, M.D.; Grino, M.; Leonetti, G.; Alessi, M.C.; Clement, K.; Dadoun, F.; Dutour, A. Epicardial Adipose Tissue Extent: Relationship with Age, Body Fat Distribution, and Coronaropathy. Obesity 2008, 16, 2424–2430. [Google Scholar] [CrossRef]
- Poehlman, E.T.; Horton, E.S. Regulation of Energy Expenditure in Aging Humans. Annu. Rev. Nutr. 1990, 10, 255–275. [Google Scholar] [CrossRef]
- Fanelli, C.; Calderone, S.; Epifano, L.; De Vincenzo, A.; Modarelli, F.; Pampanelli, S.; Perriello, G.; De Feo, P.; Brunetti, P.; Gerich, J.E. Demonstration of a critical role for free fatty acids in mediating counterregulatory stimulation of gluconeogenesis and suppression of glucose utilization in humans. J. Clin. Investig. 1993, 92, 1617–1622. [Google Scholar] [CrossRef] [Green Version]
- Boden, G.; Chen, X.; Ruiz, J.; White, J.V.; Rossetti, L. Mechanisms of fatty acid-induced inhibition of glucose uptake. J. Clin. Investig. 1994, 93, 2438–2446. [Google Scholar] [CrossRef]
- Havel, R.J.; Kane, J.P.; Balasse, E.O.; Segel, N.; Basso, L.V. Splanchnic metabolism of free fatty acids and production of triglycerides of very low density lipoproteins in normotriglyceridemic and hypertriglyceridemic humans. J. Clin. Investig. 1970, 49, 2017–2035. [Google Scholar] [CrossRef]
- Alpert, M.A.; Karthikeyan, K.; Abdullah, O.; Ghadban, R. Obesity and Cardiac Remodeling in Adults: Mechanisms and Clinical Implications. Prog. Cardiovasc. Dis. 2018, 61, 114–123. [Google Scholar] [CrossRef]
- Nishida, K.; Otsu, K. Inflammation and metabolic cardiomyopathy. Cardiovasc. Res. 2017, 113, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Vos, A.G. Obesity and the heart: The impact of obesity beyond the body mass index. Eur. J. Prev. Cardiol. 2020, 2047487319897170. [Google Scholar] [CrossRef] [Green Version]
- López-Jiménez, F.; Cortés-Bergoderi, M. Obesidad y corazón. Rev. Esp. Cardiol. 2011, 64, 140–149. [Google Scholar] [CrossRef]
- Peterson, L.R.; Soto, P.F.; Herrero, P.; Mohammed, B.S.; Avidan, M.S.; Schechtman, K.B.; Dence, C.; Gropler, R.J. Impact of gender on the myocardial metabolic response to obesity. JACC Cardiovasc. Imaging 2008, 1, 424–433. [Google Scholar] [CrossRef] [Green Version]
- Murray, A.J.; Anderson, R.E.; Watson, G.C.; Radda, G.K.; Clarke, K. Uncoupling proteins in human heart. Lancet 2004, 364, 1786–1788. [Google Scholar] [CrossRef]
- Peterson, L.R.; Waggoner, A.D.; Schechtman, K.B.; Meyer, T.; Gropler, R.J.; Barzilai, B.; Dávila-Román, V.G. Alterations in left ventricular structure and function in young healthy obese women: Assessment by echocardiography and tissue Doppler imaging. J. Am. Coll. Cardiol. 2004, 43, 1399–1404. [Google Scholar] [CrossRef] [Green Version]
- Randle, P.J.; Kerbey, A.L.; Espinal, J. Mechanisms decreasing glucose oxidation in diabetes and starvation: Role of lipid fuels and hormones. Diabetes Metab. Rev. 1988, 4, 623–638. [Google Scholar] [CrossRef]
- Chiu, H.C.; Kovacs, A.; Ford, D.A.; Hsu, F.F.; Garcia, R.; Herrero, P.; Saffitz, J.E.; Schaffer, J.E. A novel mouse model of lipotoxic cardiomyopathy. J. Clin. Investig. 2001, 107, 813–822. [Google Scholar] [CrossRef] [Green Version]
- Horwich, T.B.; Fonarow, G.C.; Clark, A.L. Obesity and the Obesity Paradox in Heart Failure. Prog. Cardiovasc. Dis. 2018, 61, 151–156. [Google Scholar] [CrossRef]
- Feuvray, D.; Darmellah, A. Diabetes-related metabolic perturbations in cardiac myocyte. Diabetes Metab. 2008, 34, 3–9. [Google Scholar] [CrossRef]
- Son, N.H.; Park, T.S.; Yamashita, H.; Yokoyama, M.; Huggins, L.A.; Okajima, K.; Homma, S.; Szabolcs, M.J.; Huang, L.S.; Goldberg, I.J. Cardiomyocyte expression of PPARgamma leads to cardiac dysfunction in mice. J. Clin. Investig. 2007, 117, 2791–2801. [Google Scholar] [CrossRef] [Green Version]
- Finck, B.N.; Lehman, J.J.; Leone, T.C.; Welch, M.J.; Bennett, M.J.; Kovacs, A.; Han, X.; Gross, R.W.; Kozak, R.; Lopaschuk, G.D.; et al. The cardiac phenotype induced by PPARalpha overexpression mimics that caused by diabetes mellitus. J. Clin. Investig. 2002, 109, 121–130. [Google Scholar] [CrossRef]
- Wong, R.H.F.; Sul, H.S. Insulin signaling in fatty acid and fat synthesis: a transcriptional perspective. Curr. Opin. Pharmacol. 2010, 10, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Marfella, R.; Di Filippo, C.; Portoghese, M.; Barbieri, M.; Ferraraccio, F.; Siniscalchi, M.; Cacciapuoti, F.; Rossi, F.; D’Amico, M.; Paolisso, G. Myocardial lipid accumulation in patients with pressure-overloaded heart and metabolic syndrome. J. Lipid Res. 2009, 50, 2314–2323. [Google Scholar] [CrossRef] [Green Version]
- Noyes, A.M. Cardiac adipose tissue and its relationship to diabetes mellitus and cardiovascular disease. World J. Diabetes 2014, 5, 868. [Google Scholar] [CrossRef]
- Atsumi, T.; Nishio, T.; Niwa, H.; Takeuchi, J.; Bando, H.; Shimizu, C.; Yoshioka, N.; Bucala, R.; Koike, T. Expression of Inducible 6-Phosphofructo-2-Kinase/Fructose-2,6-Bisphosphatase/PFKFB3 Isoforms in Adipocytes and Their Potential Role in Glycolytic Regulation. Diabetes 2005, 54, 3349–3357. [Google Scholar] [CrossRef] [Green Version]
- Stanley, W.C.; Lopaschuk, G.D.; Hall, J.L.; McCormack, J.G. Regulation of myocardial carbohydrate metabolism under normal and ischaemic conditions: Potential for pharmacological interventions. Cardiovasc. Res. 1997, 33, 243–257. [Google Scholar] [CrossRef]
- Fabiato, A.; Fabiato, F. Effects of pH on the myofilaments and the sarcoplasmic reticulum of skinned cells from cardiace and skeletal muscles. J. Physiol. 1978, 276, 233–255. [Google Scholar] [CrossRef]
- Zimmer, H.-G. Regulation of and intervention into the oxidative pentose phosphate pathway and adenine nucleotide metabolism in the heart. Mol. Cell. Biochem. 1996, 160–161, 101–109. [Google Scholar] [CrossRef]
- Lopaschuk, G.D.; Collins-Nakai, R.; Olley, P.M.; Montague, T.J.; McNeil, G.; Gayle, M.; Penkoske, P.; Finegan, B.A. Plasma fatty acid levels in infants and adults after myocardial ischemia. Am. Heart J. 1994, 128, 61–67. [Google Scholar] [CrossRef]
- Dalla-Volta, S.; Maraglino, G.; Della-Valentina, P.; Viena, P.; Desideri, A. Comparison of trimetazidine with nifedipine in effort angina: A double-blind, crossover study. Cardiovasc. Drugs Ther. 1990, 4, 853–859. [Google Scholar] [CrossRef]
- McCarthy, C.P.; Mullins, K.V.; Kerins, D.M. The role of trimetazidine in cardiovascular disease: Beyond an anti-anginal agent. Eur. Hear. J. Cardiovasc. Pharmacother. 2015, 2, 266–272. [Google Scholar] [CrossRef] [Green Version]
- Arnaud, L.; Raymond, K. The Antianginal Drug Trimetazidine Shifts Cardiac Energy Metabolism from Fatty Acid Oxidation to Glucose Oxidation by Inhibiting Mitochondrial Long-Chain 3-Ketoacyl Coenzyme A Thiolase. Circ. Res. 2000, 86, 580–588. [Google Scholar] [CrossRef] [Green Version]
- Leong, H.S.; Brownsey, R.W.; Kulpa, J.E.; Allard, M.F. Glycolysis and pyruvate oxidation in cardiac hypertrophy—Why so unbalanced? Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 135, 499–513. [Google Scholar] [CrossRef]
- Baba, S.P.; Zhang, D.; Singh, M.; Dassanayaka, S.; Xie, Z.; Jagatheesan, G.; Zhao, J.; Schmidtke, V.K.; Brittian, K.R.; Merchant, M.L.; et al. Deficiency of aldose reductase exacerbates early pressure overload-induced cardiac dysfunction and autophagy in mice. J. Mol. Cell. Cardiol. 2018, 118, 183–192. [Google Scholar] [CrossRef]
- McCommis, K.S.; Douglas, D.L.; Krenz, M.; Baines, C.P. Cardiac-specific hexokinase 2 overexpression attenuates hypertrophy by increasing pentose phosphate pathway flux. J. Am. Heart Assoc. 2013, 2, e000355. [Google Scholar] [CrossRef]
- Markan, K.R.; Jurczak, M.J.; Brady, M.J. Stranger in a strange land: Roles of glycogen turnover in adipose tissue metabolism. Mol. Cell. Endocrinol. 2010, 318, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frayn, K.N.; Humphreys, S.M. Metabolic characteristics of human subcutaneous abdominal adipose tissue after overnight fast. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E468–E475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabkin, S.W. Epicardial fat: Properties, function and relationship to obesity. Obes. Rev. 2007, 8, 253–261. [Google Scholar] [CrossRef]
- Liang, K.W.; Tsai, I.C.; Lee, W.J.; Lee, I.T.; Lee, W.L.; Lin, S.Y.; Wan, C.J.; Fu, C.P.; Ting, C.T.; Sheu, W.H. MRI Measured Epicardial Adipose Tissue Thickness at the Right AV Groove Differentiates Inflammatory Status in Obese Men with Metabolic Syndrome. Obesity 2012, 20, 525–532. [Google Scholar] [CrossRef]
- Nieto-Vazquez, I.; Fernández-Veledo, S.; Krämer, D.K.; Vila-Bedmar, R.; Garcia-Guerra, L.; Lorenzo, M. Insulin resistance associated to obesity: The link TNF-alpha. Arch. Physiol. Biochem. 2008, 114, 183–194. [Google Scholar] [CrossRef]
- Han, L.; Liu, J.; Zhu, L.; Tan, F.; Qin, Y.; Huang, H.; Yu, Y. Free fatty acid can induce cardiac dysfunction and alter insulin signaling pathways in the heart. Lipids Health Dis. 2018, 17, 185. [Google Scholar] [CrossRef] [Green Version]
- Iacobellis, G.; Leonetti, F. Epicardial Adipose Tissue and Insulin Resistance in Obese Subjects. J. Clin. Endocrinol. Metab. 2005, 90, 6300–6302. [Google Scholar] [CrossRef]
- Iacobellis, G.; Barbaro, G.; Gerstein, H.C. Relationship of epicardial fat thickness and fasting glucose. Int. J. Cardiol. 2008, 128, 424–426. [Google Scholar] [CrossRef]
- Ferder, L.; Inserra, F.; Martínez-Maldonado, M. Inflammation and the metabolic syndrome: Role of angiotensin II and oxidative stress. Curr. Hypertens. Rep. 2006, 8, 191–198. [Google Scholar] [CrossRef]
- Roubícek, T.; Dolinková, M.; Bláha, J.; Haluzíková, D.; Bosanská, L.; Mráz, M.; Kremen, J.; Haluzík, M. Increased angiotensinogen production in epicardial adipose tissue during cardiac surgery: Possible role in a postoperative insulin resistance. Physiol. Res. 2008, 57, 1–18. [Google Scholar]
- Kotulák, T.; Drápalová, J.; Kopecký, P.; Lacinová, Z.; Kramář, P.; Říha, H.; Netuka, I.; Malý, J.; Housa, D.; Bláha, J.; et al. Increased circulating and epicardial adipose tissue mrna expression of fibroblast growth factor-21 after cardiac surgery: Possible role in postoperative inflammatory response and insulin resistance. Physiol. Res. 2011, 60, 757–767. [Google Scholar] [CrossRef]
- Kremen, J.; Dolinkova, M.; Krajickova, J.; Blaha, J.; Anderlova, K.; Lacinova, Z.; Haluzikova, D.; Bosanska, L.; Vokurka, M.; Svacina, S.; et al. Increased Subcutaneous and Epicardial Adipose Tissue Production of Proinflammatory Cytokines in Cardiac Surgery Patients: Possible Role in Postoperative Insulin Resistance. J. Clin. Endocrinol. Metab. 2006, 91, 4620–4627. [Google Scholar] [CrossRef] [Green Version]
- Nasarre, L.; Juan-Babot, O.; Gastelurrutia, P.; Llucia-Valldeperas, A.; Badimon, L.; Bayes-Genis, A.; Llorente-Cortés, V. Low density lipoprotein receptor–related protein 1 is upregulated in epicardial fat from type 2 diabetes mellitus patients and correlates with glucose and triglyceride plasma levels. Acta Diabetol. 2014, 51, 23–30. [Google Scholar] [CrossRef]
- Fernández-Trasancos, A.; Fandiño-Vaquero, R.; Agra, R.M.; Fernández, A.L.; Viñuela, J.E.; González-Juanatey, J.R.; Eiras, S. Impaired Adipogenesis and Insulin Resistance in Epicardial Fat-Mesenchymal Cells From Patients With Cardiovascular Disease. J. Cell. Physiol. 2014, 229, 1722–1730. [Google Scholar] [CrossRef]
- Fernández-Trasancos, Á.; Guerola-Segura, R.; Paradela-Dobarro, B.; Álvarez, E.; García-Acuña, J.M.; Fernández, Á.L.; González-Juanatey, J.R.; Eiras, S. Glucose and Inflammatory Cells Decrease Adiponectin in Epicardial Adipose Tissue Cells: Paracrine Consequences on Vascular Endothelium. J. Cell. Physiol. 2016, 231, 1015–1023. [Google Scholar] [CrossRef]
- Salgado-Somoza, A.; Teijeira-Fernández, E.; Rubio, J.; Couso, E.; González-Juanatey, J.R.; Eiras, S. Coronary artery disease is associated with higher epicardial Retinol-binding protein 4 (RBP4) and lower glucose transporter (GLUT) 4 levels in epicardial and subcutaneous adipose tissue. Clin. Endocrinol. (Oxf.) 2012, 76, 51–58. [Google Scholar] [CrossRef]
- Greulich, S.; de Wiza, D.H.; Preilowski, S.; Ding, Z.; Mueller, H.; Langin, D.; Jaquet, K.; Ouwens, D.M.; Eckel, J. Secretory products of guinea pig epicardial fat induce insulin resistance and impair primary adult rat cardiomyocyte function. J. Cell. Mol. Med. 2011, 15, 2399–2410. [Google Scholar] [CrossRef] [Green Version]
- Obokata, M.; Reddy, Y.N.V.; Pislaru, S.V.; Melenovsky, V.; Borlaug, B.A. Evidence Supporting the Existence of a Distinct Obese Phenotype of Heart Failure with Preserved Ejection Fraction. Circulation 2017, 136, 6–19. [Google Scholar] [CrossRef]
- Doehner, W.; Frenneaux, M.; Anker, S.D. Metabolic Impairment in Heart Failure: The Myocardial and Systemic Perspective. J. Am. Coll. Cardiol. 2014, 64, 1388–1400. [Google Scholar] [CrossRef]
- Rydén, M.; Arner, P. Fat loss in cachexia—Is there a role for adipocyte lipolysis? Clin. Nutr. 2007, 26, 1–6. [Google Scholar] [CrossRef]
- Szabó, T.; Postrach, E.; Mähler, A.; Kung, T.; Turhan, G.; von Haehling, S.; Anker, S.D.; Boschmann, M.; Doehner, W. Increased catabolic activity in adipose tissue of patients with chronic heart failure. Eur. J. Heart Fail. 2013, 15, 1131–1137. [Google Scholar] [CrossRef]
- Khawaja, T.; Greer, C.; Chokshi, A.; Chavarria, N.; Thadani, S.; Jones, M.; Schaefle, K.; Bhatia, K.; Collado, J.E.; Shimbo, D.; et al. Epicardial Fat Volume in Patients With Left Ventricular Systolic Dysfunction. Am. J. Cardiol. 2011, 108, 397–401. [Google Scholar] [CrossRef]
- Fosshaug, L.E.; Dahl, C.P.; Risnes, I.; Bohov, P.; Berge, R.K.; Nymo, S.; Geiran, O.; Yndestad, A.; Gullestad, L.; Aukrust, P.; et al. Altered Levels of Fatty Acids and Inflammatory and Metabolic Mediators in Epicardial Adipose Tissue in Patients with Systolic Heart Failure. J. Card. Fail. 2015, 21, 916–923. [Google Scholar] [CrossRef]
- Burgeiro, A.; Fuhrmann, A.; Cherian, S.; Espinoza, D.; Jarak, I.; Carvalho, R.A.; Loureiro, M.; Patrício, M.; Antunes, M.; Carvalho, E. Glucose uptake and lipid metabolism are impaired in epicardial adipose tissue from heart failure patients with or without diabetes. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E550–E564. [Google Scholar] [CrossRef] [Green Version]
- Flüchter, S.; Haghi, D.; Dinter, D.; Heberlein, W.; Kühl, H.P.; Neff, W.; Sueselbeck, T.; Borggrefe, M.; Papavassiliu, T. Volumetric Assessment of Epicardial Adipose Tissue with Cardiovascular Magnetic Resonance Imaging. Obesity 2007, 15, 870–878. [Google Scholar] [CrossRef]
- Purohit, A.; Reed, M.J. Regulation of estrogen synthesis in postmenopausal women. Steroids 2002, 67, 979–983. [Google Scholar] [CrossRef]
- El Khoudary, S.R.; Zhao, Q.; Venugopal, V.; Manson, J.E.; Brooks, M.M.; Santoro, N.; Black, D.M.; Harman, S.M.; Cedars, M.I.; Hopkins, P.N.; et al. Effects of Hormone Therapy on Heart Fat and Coronary Artery Calcification Progression: Secondary Analysis from the KEEPS Trial. J. Am. Heart Assoc. 2019, 8, e012763. [Google Scholar] [CrossRef] [Green Version]
- Homma, H.; Kurachi, H.; Nishio, Y.; Takeda, T.; Yamamoto, T.; Adachi, K.; Morishige, K.; Ohmichi, M.; Matsuzawa, Y.; Murata, Y. Estrogen Suppresses Transcription of Lipoprotein Lipase Gene: Existence of a unique estrogen response element on the lipoprotein lipase promoter. J. Biol. Chem. 2000, 275, 11404–11411. [Google Scholar] [CrossRef] [Green Version]
- Palin, S.L.; McTernan, P.G.; Anderson, L.A.; Sturdee, D.W.; Barnett, A.H.; Kumar, S. 17β-estradiol and anti-estrogen ICI:Compound 182,780 regulate expression of lipoprotein lipase and hormone-sensitive lipase in isolated subcutaneous abdominal adipocytes. Metabolism 2003, 52, 383–388. [Google Scholar] [CrossRef]
- Gormsen, L.C.; Høst, C.; Hjerrild, B.E.; Pedersen, S.B.; Nielsen, S.; Christiansen, J.S.; Gravholt, C.H. Estradiol acutely inhibits whole body lipid oxidation and attenuates lipolysis in subcutaneous adipose tissue: A randomized, placebo-controlled study in postmenopausal women. Eur. J. Endocrinol. 2012, 167, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Lafontan, M.; Berlan, M. Fat cell adrenergic receptors and the control of white and brown fat cell function. J. Lipid Res. 1993, 34, 1057–1091. [Google Scholar]
- Barrett-Connor, E.; Mosca, L.; Collins, P.; Geiger, M.J.; Grady, D.; Kornitzer, M.; McNabb, M.A.; Wenger, N.K. Effects of Raloxifene on Cardiovascular Events and Breast Cancer in Postmenopausal Women. N. Engl. J. Med. 2006, 355, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Manson, J.E.; Hsia, J.; Johnson, K.C.; Rossouw, J.E.; Assaf, A.R.; Lasser, N.L.; Trevisan, M.; Black, H.R.; Heckbert, S.R.; Detrano, R.; et al. Estrogen plus Progestin and the Risk of Coronary Heart Disease. N. Engl. J. Med. 2003, 349, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Sokos, G.G.; Bolukoglu, H.; German, J.; Hentosz, T.; Magovern, G.J., Jr.; Maher, T.D.; Dean, D.A.; Bailey, S.H.; Marrone, G.; Benckart, D.H.; et al. Effect of glucagon-like peptide-1 (GLP-1) on glycemic control and left ventricular function in patients undergoing coronary artery bypass grafting. Am. J. Cardiol. 2007, 100, 824–829. [Google Scholar] [CrossRef]
- McCormick, L.M.; Hoole, S.P.; White, P.A.; Read, P.A.; Axell, R.G.; Clarke, S.J.; O’Sullivan, M.; West, N.; Dutka, D.P. Pre-treatment with glucagon-like peptide-1 protects against ischemic left ventricular dysfunction and stunning without a detected difference in myocardial substrate utilization. JACC Cardiovasc Interv. 2015, 8, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Marso, S.P.; Bain, S.C.; Consoli, A.; Eliaschewitz, F.G.; Jódar, E.; Leiter, L.A.; Lingvay, I.; Rosenstock, J.; Seufert, J.; Warren, M.L.; et al. Investigators, semaglutide and cardiovascular outcomes in patients with type 2 diabetes. N. Engl. J. Med. 2016, 375, 1834–1844. [Google Scholar] [CrossRef] [Green Version]
- Drucker, D.J. The Cardiovascular Biology of Glucagon-like Peptide-1. Cell Metabol. 2016, 24, 15–30. [Google Scholar] [CrossRef]
- Nyström, T.; Gonon, A.T.; Sjöholm, Å.; Pernow, J. Glucagon-like peptide-1 relaxes rat conduit arteries via an endothelium-independent mechanism. Regul. Pept. 2005, 125, 173–177. [Google Scholar] [CrossRef]
- Iacobellis, G.; Camarena, V.; Sant, D.W.; Wang, G. Human Epicardial Fat Expresses Glucagon-Like Peptide 1 and 2 Receptors Genes. Horm. Metab. Res. 2017, 49, 625–630. [Google Scholar] [CrossRef]
- Dozio, E.; Vianello, E.; Malavazos, A.E.; Tacchini, L.; Schmitz, G.; Iacobellis, G.; Corsi Romanelli, M.M. Epicardial adipose tissue GLP-1 receptor is associated with genes involved in fatty acid oxidation and white-to-brown fat differentiation: A target to modulate cardiovascular risk? Int. J. Cardiol. 2019, 292, 218–224. [Google Scholar] [CrossRef] [Green Version]
- Batran, R.; Almutairi, M.; Ussher, J.R. Glucagon-like peptide-1 receptor mediated control of cardiac energy metabolism. Peptides 2018, 100, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Dutour, A.; Abdesselam, I.; Ancel, P.; Kober, F.; Mrad, G.; Darmon, P.; Ronsin, O.; Pradel, V.; Lesavre, N.; Martin, J.C.; et al. Exenatide decreases liver fat content and epicardial adipose tissue in patients with obesity and type 2 diabetes: A prospective randomized clinical trial using magnetic resonance imaging and spectroscopy. Diabetes Obes. Metab. 2016, 18, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Iacobellis, G.; Mohseni, M.; Bianco, S.D.; Banga, P.K. Liraglutide causes large and rapid epicardial fat reduction. Obesity 2017, 25, 311–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bizino, M.B.; Jazet, I.M.; de Heer, P.; van Eyk, H.J.; Dekkers, I.A.; Rensen, P.; Paiman, E.; Lamb, H.J.; Smit, J.W. Placebo-controlled randomised trial with liraglutide on magnetic resonance endpoints in individuals with type 2 diabetes: a pre-specified secondary study on ectopic fat accumulation. Diabetologia 2020, 63, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Erdogdu, O.; Nathanson, D.; Sjöholm, A.; Nyström, T.; Zhang, Q. Exendin-4 stimulates proliferation of human coronary artery endothelial cells through eNOS-, PKA- and PI3K/Akt-dependent pathways and requires GLP-1 receptor. Mol. Cell Endocrinol. 2010, 325, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Opie, L.H. Proof that glucose-insulin-potassium provides metabolic protection of ischaemic myocardium? Lancet 1999, 353, 768–769. [Google Scholar] [CrossRef]
- Carbó, R.; Nava, P.; Guarner, V. Effects of polarizing solution on glucose uptake of rat oxygenated or hypoxic ventricular myocytes. Clin. Exp. Pharmacol. Physiol. 2003, 30, 64–71. [Google Scholar] [CrossRef]
- Lazar, H.L.; Chipkin, S.; Philippides, G.; Bao, Y.; Apstein, C. Glucose-insulin-potassium solutions improve outcomes in diabetics who have coronary artery operations. Ann. Thorac. Surg. 2000, 70, 145–150. [Google Scholar] [CrossRef]
- Coleman, G.M.; Gradinac, S.; Taegtmeyer, H.; Sweeney, M.; Frazier, O.H. Efficacy of metabolic support with glucose-insulin-potassium for left ventricular pump failure after aortocoronary bypass surgery. Circulation 1989, 80, 1916–1989. [Google Scholar]
- Zabala, R.C.; Guarner, V. Cambios en el metabolismo cardíaco y su posible aprovechamiento en la terapéutica (Parte II). Arch. Cardiol. Mex. 2004, 74, 68–79. [Google Scholar]
- Yang, Y.K.; Wang, L.P.; Chen, L.; Yao, X.P.; Yang, K.Q.; Gao, L.G.; Zhou, X.L. Coenzyme Q10 treatment of cardiovascular disorders of ageing including heart failure, hypertension and endothelial dysfunction. Clin. Chim. Acta 2015, 450, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Jafari, M.; Mousavi, S.M.; Asgharzadeh, A.; Yazdani, N. Coenzyme Q10 in the treatment of heart failure: A systematic review of systematic reviews. Indian Heart J. 2018, 70, S111–S117. [Google Scholar] [CrossRef] [PubMed]
- Alehagen, U.; Aaseth, J.; Johansson, P. Reduced Cardiovascular Mortality 10 Years after Supplementation with Selenium and Coenzyme Q10 for Four Years: Follow-Up Results of a Prospective Randomized Double-Blind Placebo-Controlled Trial in Elderly Citizens. PLoS ONE 2015, 10, e0141641. [Google Scholar] [CrossRef]
- Sobirin, M.A.; Herry, Y.; Sofia, S.N.; Uddin, I.; Rifqi, S.; Tsutsui, H. Effects of coenzyme Q10 supplementation on diastolic function in patients with heart failure with preserved ejection fraction. Drug Discov. Ther. 2019, 13, 38–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholte, H.R.; Luyt-Houwen, I.E.M.; Vaandrager-Verduin, M.H. The role of the carnitine system in myocardial fatty acid oxidation: Carnitine deficiency, failing mitochondria and cardiomyopathy. Basic Res. Cardiol. 1987, 82, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Pekala, J.; Patkowska-Sokola, B.; Bodkowski, R.; Jamroz, D.; Nowakowski, P.; Lochynski, S.; Librowski, T. L-Carnitine Metabolic Functions and Meaning in Humans Life. Curr. Drug Metabol. 2011, 12, 667–678. [Google Scholar] [CrossRef]
- Hoppel, C. The role of carnitine in normal and altered fatty acid metabolism. Am. J. Kidney Dis. 2003, 41, S4–S12. [Google Scholar] [CrossRef]
- Yancy, C.W.; Jessup, M.; Bozkurt, B.; Butler, J.; Casey, D.E., Jr.; Drazner, M.H.; Fonarow, G.C.; Geraci, S.A.; Horwich, T.; Januzzi, J.L.; et al. 2013 ACCF/AHA Guideline for the Management of Heart Failure: Executive Summary. Circulation 2013, 128, 1810–1852. [Google Scholar] [CrossRef]
- Marzilli, M.; Vinereanu, D.; Lopaschuk, G.; Chen, Y.; Dalal, J.J.; Danchin, N.; Etriby, E.; Ferrari, R.; Gowdak, L.H.; Lopatin, Y.; et al. Trimetazidine in cardiovascular medicine. Int. J. Cardiol. 2019, 293, 39–44. [Google Scholar] [CrossRef]
- Lopatin, Y. Metabolic cardiac protection is beneficial in patients undergoing coronary revascularization: Is it necessary afterwards? Heart. Metab. 2013, 58, 25–30. [Google Scholar]
- Zhang, N.; Lei, J.; Liu, Q.; Huang, W.; Xiao, H.; Lei, H. The Effectiveness of Preoperative Trimetazidine on Myocardial Preservation in Coronary Artery Bypass Graft Patients: A Systematic Review and Meta-Analysis. Cardiology 2015, 131, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Lommi, J.; Kupari, M.; Yki-Järvinen, H. Free Fatty Acid Kinetics and Oxidation in Congestive Heart Failure. Am. J. Cardiol. 1998, 81, 45–50. [Google Scholar] [CrossRef]
- Fragasso, G.; Rosano, G.; Baek, S.H.; Sisakian, H.; Di Napoli, P.; Alberti, L.; Calori, G.; Kang, S.M.; Sahakyan, L.; Sanosyan, A.; et al. Effect of partial fatty acid oxidation inhibition with trimetazidine on mortality and morbidity in heart failure: Results from an international multicentre retrospective cohort study. Int. J. Cardiol. 2013, 163, 320–325. [Google Scholar] [CrossRef]
- Wiviott, S.D.; Raz, I.; Bonaca, M.P.; Mosenzon, O.; Kato, E.T.; Cahn, A.; Silverman, M.G.; Zelniker, T.A.; Kuder, J.F.; Murphy, S.A.; et al. Dapagliflozin and Cardiovascular Outcomes in Type 2 Diabetes. N. Engl. J. Med. 2018, 380, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Zinman, B.; Lachin, J.M.; Inzucchi, S.E. Empagliflozin, Cardiovascular Outcomes, and Mortality in Type 2 Diabetes. N. Engl. J. Med. 2015, 373, 2117–2128. [Google Scholar] [CrossRef]
- Neal, B.; Perkovic, V.; Mahaffey, K.W.; de Zeeuw, D.; Fulcher, G.; Erondu, N.; Shaw, W.; Law, G.; Desai, M.; Matthews, D.R.; et al. Canagliflozin and Cardiovascular and Renal Events in Type 2 Diabetes. N. Engl. J. Med. 2017, 377, 644–657. [Google Scholar] [CrossRef]
- Yancy, C.W.; Januzzi, J.L., Jr.; Allen, L.A.; Butler, J.; Davis, L.L.; Fonarow, G.C.; Ibrahim, N.E.; Jessup, M.; Lindenfeld, J.; Maddox, T.M.; et al. 2017 ACC Expert Consensus Decision Pathway for Optimization of Heart Failure Treatment: Answers to 10 Pivotal Issues About Heart Failure With Reduced Ejection Fraction: A Report of the American College of Cardiology Task Force on Expert Consensus Decision. J. Am. Coll. Cardiol. 2018, 71, 201–230. [Google Scholar] [CrossRef]
- Couselo-Seijas, M.; Agra-Bermejo, R.M.; Fernández, A.L.; Martínez-Cereijo, J.M.; Sierra, J.; Soto-Pérez, M.; Rozados-Luis, A.; González-Juanatey, J.R.; Eiras, S. High released lactate by epicardial fat from coronary artery disease patients is reduced by dapagliflozin treatment. Atherosclerosis 2020, 292, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Rodríguez, E.; Agra, R.M.; Fernández, Á.L.; Adrio, B.; García-Caballero, T.; González-Juanatey, J.R.; Eiras, S. Effects of dapagliflozin on human epicardial adipose tissue: modulation of insulin resistance, inflammatory chemokine production, and differentiation ability. Cardiovasc. Res. 2017, 114, 336–346. [Google Scholar] [CrossRef] [Green Version]
- Driver, C.; Bamitale, K.D.S.; Kazi, A.; Olla, M.; Nyane, N.A.; Owira, P.M.O. Cardioprotective Effects of Metformin. J. Cardiovasc. Pharmacol. 2018, 72, 121–127. [Google Scholar] [CrossRef]
- Bairwa, S.C.; Parajuli, N.; Dyck, J.R. The role of AMPK in cardiomyocyte health and survival. Biochim. Biophys. Acta. 2016, 1862, 2199–2210. [Google Scholar] [CrossRef] [PubMed]
- Kahn, B.B.; Alquier, T.; Carling, D.; Hardie, D.G. AMP-activated protein kinase: Ancient energy gauge provides clues to modern understanding of metabolism. Cell Metab. 2015, 1, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Ziyrek, M.; Kahraman, S.; Ozdemir, E.; Dogan, A. Metformin monotherapy significantly decreases epicardial adipose tissue thickness in newly diagnosed type 2 diabetes patients. Rev. Port. Cardiol. 2019, 38, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Xie, H.; Liu, Y.; Gao, P.; Yang, X.; Shen, Z. Effect of metformin on all-cause and cardiovascular mortality in patients with coronary artery diseases: A systematic review and an updated meta-analysis. Cardiovasc. Diabetol. 2019, 18, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Pathological Conditions | Energy Substrate |
|---|---|
| Obesity | Fatty acids |
| Diabetes | Fatty acids |
| Ischemic cardiomyopathy | Glucose, lactate |
| Hypertrophic cardiomyopathy | Glucose |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gandoy-Fieiras, N.; Gonzalez-Juanatey, J.R.; Eiras, S. Myocardium Metabolism in Physiological and Pathophysiological States: Implications of Epicardial Adipose Tissue and Potential Therapeutic Targets. Int. J. Mol. Sci. 2020, 21, 2641. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072641
Gandoy-Fieiras N, Gonzalez-Juanatey JR, Eiras S. Myocardium Metabolism in Physiological and Pathophysiological States: Implications of Epicardial Adipose Tissue and Potential Therapeutic Targets. International Journal of Molecular Sciences. 2020; 21(7):2641. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072641
Chicago/Turabian StyleGandoy-Fieiras, Nerea, Jose Ramon Gonzalez-Juanatey, and Sonia Eiras. 2020. "Myocardium Metabolism in Physiological and Pathophysiological States: Implications of Epicardial Adipose Tissue and Potential Therapeutic Targets" International Journal of Molecular Sciences 21, no. 7: 2641. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072641