Heterogeneity of Stemlike Circulating Tumor Cells in Invasive Breast Cancer

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
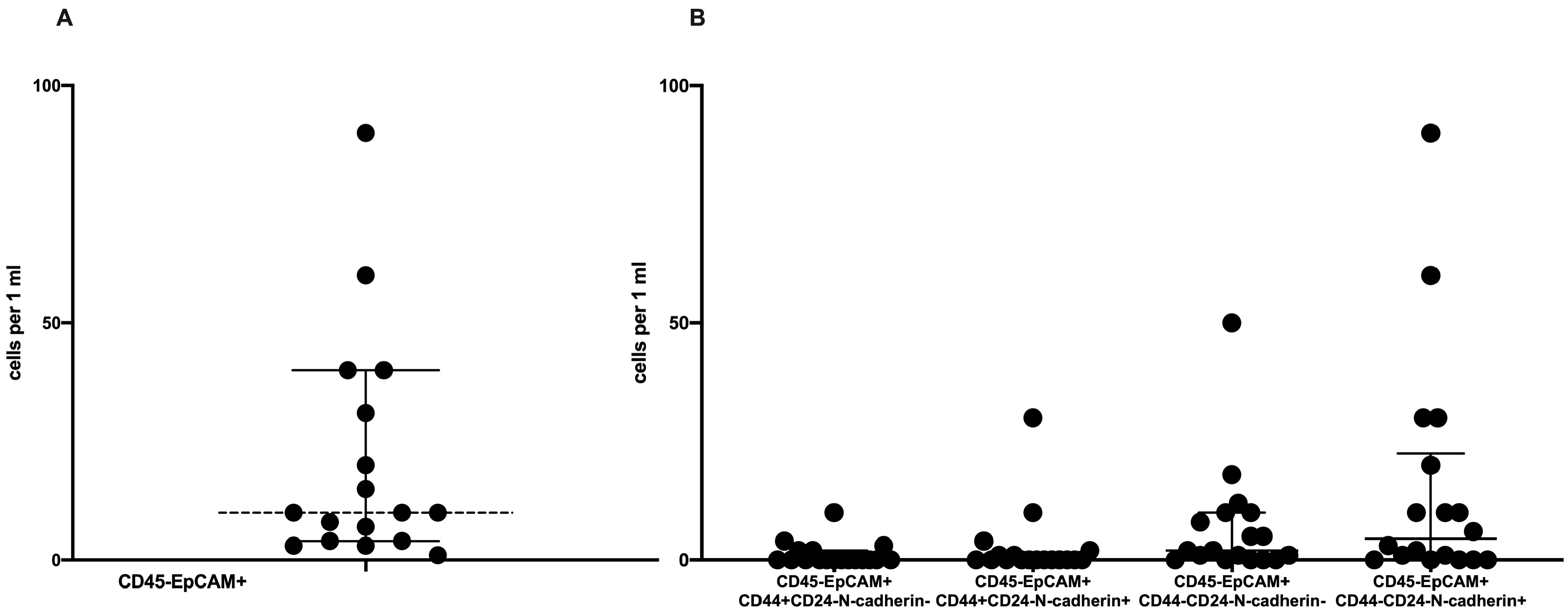
2.1. Phenotypic CTC Heterogeneity
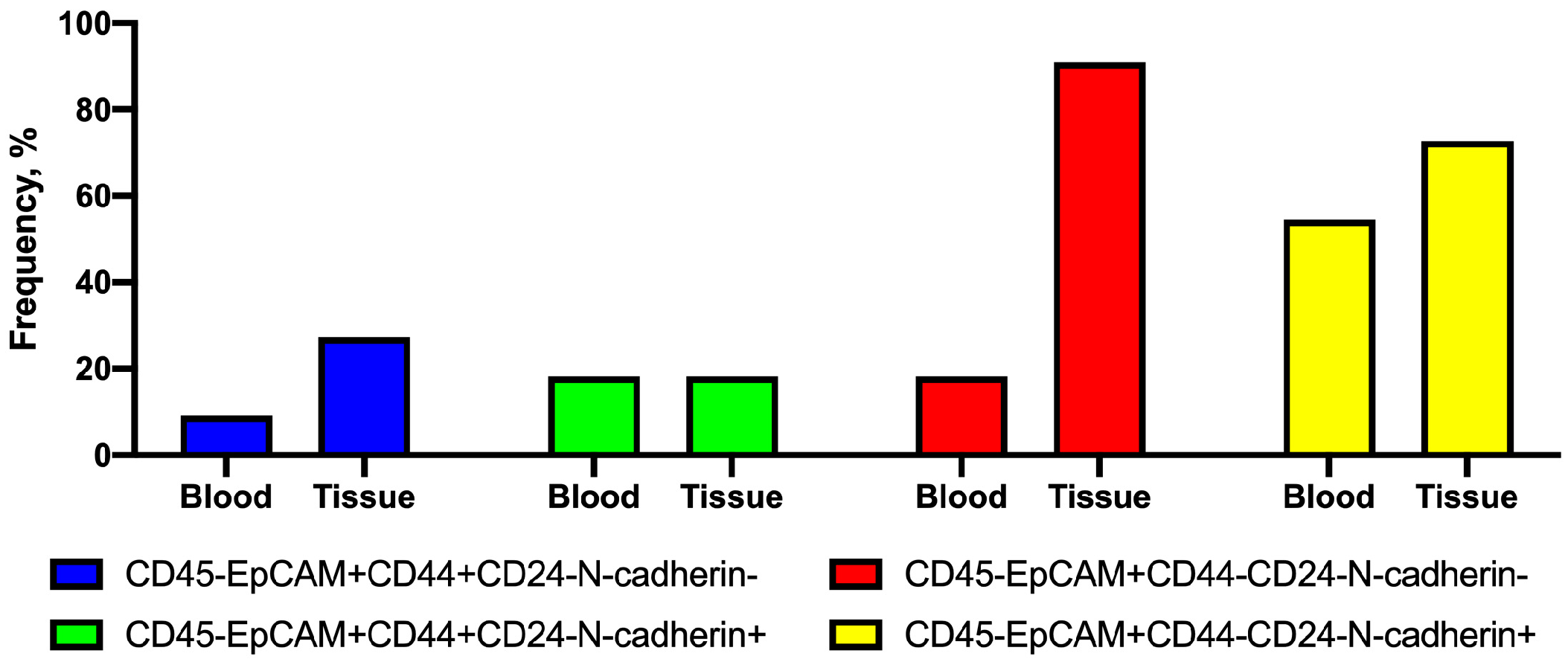
2.2. Frequency of Tumor Cells with Combinations of CD44 and N-Cadherin Expression in Primary Tumors and Peripheral Blood
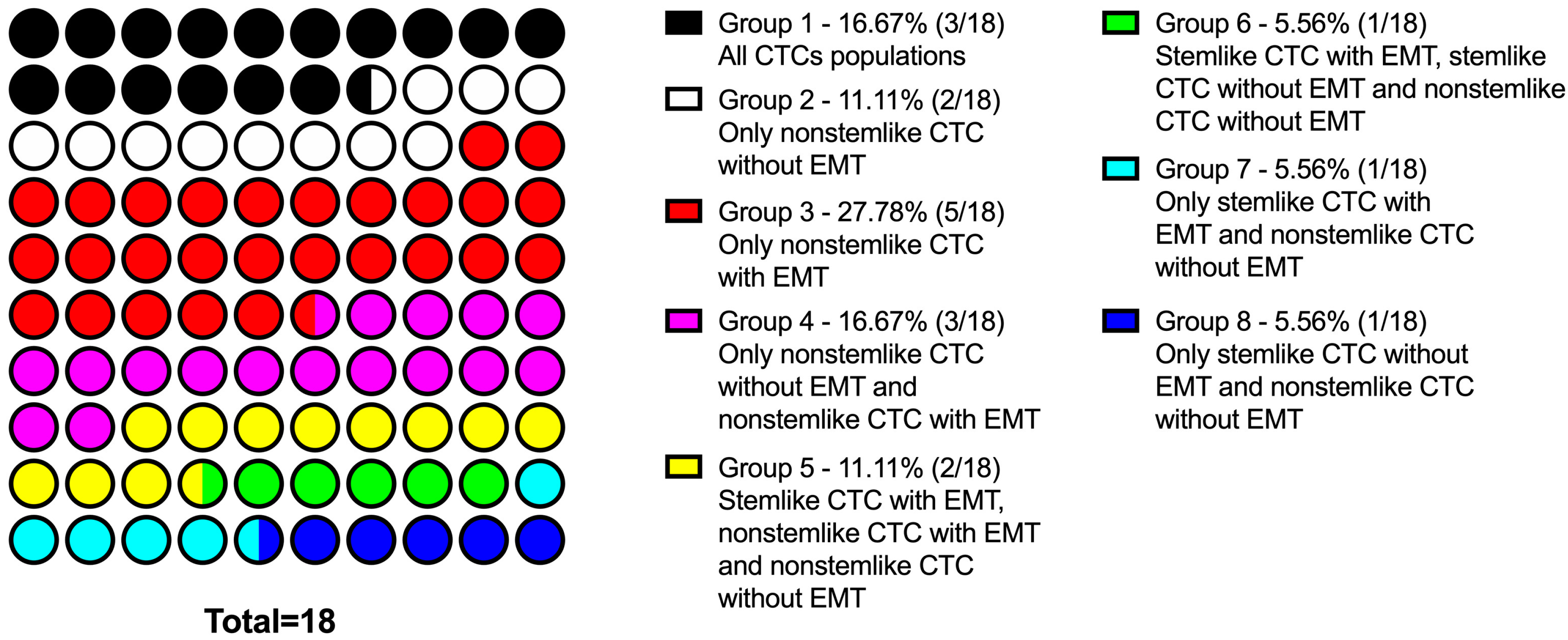
2.3. Combinations of Stem Markers in CTC Subsets
2.4. Association of CTC Subset Frequencies with Clinicopathological Parameters
3. Discussion
Summary
- CTCs were detected in 47% of patients with invasive carcinoma of no special type (NST).
- CTC population in NST patients is characterized by significant interpersonal heterogeneity: there are cells with only stem features, ones with only EMT features, with a combination of these properties, and with the absence of both features.
- Interpersonal heterogeneity of CTCs in patients with NST is characterized by different frequency of occurrence of each subsets and different combinations of subsets in each patient.
- Depending on the presence of different subsets of tumor cells in the primary tumor and among CTCs, the following patterns were revealed:
- ○
- stem cells, regardless of the expression of N-cadherin, were detected either only in the tumor or only among CTCs, and there were no cases of the simultaneous detection of such cells in the tumor and in the peripheral blood;
- ○
- nonstem cancer cells were simultaneously present in the tumor and in the peripheral blood, and more often, this combination was found in the presence of EMT features (positive N-cadherin expression).
- In patients who had CD44-CD24- CTCs, the subset of cells with the expression of other stem-cell markers (CD133 and ALDH1) was detected.
- The expression of CD133 or/and ALDH1 may be associated with the expression of N-cadherin:
- ○
- all populations of N-cadherin+ CTCs demonstrated stem features;
- ○
- in the absence of N-cadherin expression, true nonstem (CD44-CD24-CD133-ALDH1-) cells were found.
- The presence of CD45-EpCAM+CD44+CD24-N-cadherin- cells in NST patients is associated with lymph-node metastasis.
4. Materials and Methods
4.1. Samples
4.2. CTC Enrichment and Flow Cytometry
4.3. CTC Spiking Experiment
4.4. TSA-Assistance Multiplex Immunofluorescence
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| EMT | Epithelial–mesenchymal transition |
| CTCs | Circulating tumor cells |
| ALDH1 | Aldehyde dehydrogenase 1 |
| DTCs | Disseminated tumor cells |
| NST | Invasive breast carcinoma of no special type |
| TMEM | Tumor microenvironment of metastasis |
References
- Tayoun, T.; Faugeroux, V.; Oulhen, M.; Aberlenc, A.; Pawlikowska, P.; Farace, F. CTC-Derived Models: A Window into the Seeding Capacity of Circulating Tumor Cells (CTCs). Cells 2019, 8, 1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psaila, B.; Lyden, D. The Metastatic Niche: Adapting the Foreign Soil. Nat. Rev. Cancer 2009, 9, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Barcellos-Hoff, M.H.; Lyden, D.; Wang, T.C. The evolution of the cancer niche during multistage carcinogenesis. Nat. Rev. Cancer 2013, 13, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Recasens, A.; Munoz, L. Targeting Cancer Cell Dormancy. Trends Pharmacol. Sci. 2019, 40, 128–141. [Google Scholar] [CrossRef]
- Agnoletto, C.; Corrà, F.; Minotti, L.; Baldassari, F.; Crudele, F.; Cook, W.J.J.; Di Leva, G.; d’Adamo, A.P.; Gasparini, P.; Volinia, S. Heterogeneity in Circulating Tumor Cells: The Relevance of the Stem-Cell Subset. Cancers 2019, 11, 483. [Google Scholar] [CrossRef] [Green Version]
- Shibue, T.; Weinberg, R.A. EMT, CSCs, and drug resistance: The mechanistic link and clinical implications. Nat. Rev. Clin. Oncol. 2018, 14, 611–629. [Google Scholar] [CrossRef] [Green Version]
- George, J.T.; Jolly, M.K.; Xu, S.; Somarelli, J.A.; Levine, H. Survival Outcomesin Cancer Patients Predicted by a Partial EMT Gene Expression Scoring Metric. Cancer Res. 2017, 77, 6415–6428. [Google Scholar] [CrossRef] [Green Version]
- Markiewicz, A.; Topa, J.; Nagel, A.; Skokowski, J.; Seroczynska, B.; Stokowy, T.; Welnicka-Jaskiewicz, M.; Zaczek, A.J. Spectrum of Epithelial-Mesenchymal Transition Phenotypes in Circulating Tumour Cells from Early Breast Cancer Patients. Cancers 2019, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- Jolly, M.K.; Jia, D.; Boareto, M.; Mani, S.A.; Pienta, K.J.; Ben-Jacob, E.; Levine, H. Coupling the modules of EMT and stemness: Atunable ‘stemness window’ model. Oncotarget 2015, 6, 25161–25174. [Google Scholar] [CrossRef] [Green Version]
- Loh, C.-Y.; Chai, J.Y.; Tang, T.F.; Wong, W.F.; Sethi, G.; Shanmugam, M.K.; Chong, P.P.; Looi, C.Y. The E-Cadherin and N-Cadherin Switch in Epithelial-to-Mesenchymal Transition: Signaling, Therapeutic Implications, and Challenges. Cells 2019, 8, 1118. [Google Scholar] [CrossRef] [Green Version]
- Paula, A.D.C.; Lopes, C. Implications of different cancer stem cell phenotypes in breast cancer. Anticancer Res. 2017, 37, 2173–2183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bane, A.; Viloria-Petit, A.; Pinnaduwage, D.; Mulligan, A.M.; O’Malley, F.P.; Andrulis, I.L. Clinical–pathologic significance of cancer stem cell marker expression in familial breast cancers. Breast Cancer Res. Treat. 2013, 140, 195–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginestier, C.; Hur, M.H.; Charafe-Jauffret, E.; Monville, F.; Dutcher, J.; Brown, M.; Jacquemier, J.; Viens, P.; Kleer, C.G.; Liu, S.; et al. ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cell Stem Cell 2007, 1, 555–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Lu, Y.; Jiang, X.; Li, X. Clinicopathological significancea nd prognostic value of CD133 expression in triple-negativeb reast carcinoma. Cancer Sci. 2011, 102, 1107–1111. [Google Scholar] [CrossRef]
- Nadal, R.; Ortega, F.G.; Salido, M.; Lorente, J.A.; Rodriguez-Rivera, M.; Delgado-Rodriguez, M.; Macià, M.; Fernández, A.; Corominas, J.M.; Garcia-Puche, J.L.; et al. CD133 expression in circulating tumor cells from breast cancer patients: Potential role in resistance to chemotherapy. Int. J. Cancer 2013, 133, 2398–2407. [Google Scholar] [CrossRef] [Green Version]
- Wirtz, D.; Konstantopoulos, K.; Searson, P.C. The physics of cancer: The role of physical interactions and mechanical forces in metastasis. Nat. Rev. Cancer 2011, 11, 512–522. [Google Scholar] [CrossRef] [Green Version]
- Rejniak, K.A. Circulating tumor cells: When a solid tumor meets a fluid microenvironment. Adv. Exp. Med. Biol. 2016, 936, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Grosse-Wilde, A.; d’Hérouël, A.F.; McIntosh, E.; Ertaylan, G.; Skupin, A.; Kuestner, R.E.; del Sol, A.; Walters, K.-A.; Huang, S. Stemness of the hybrid Epithelial/Mesenchymal Statein Breast Cancer and Its Association with Poor Survival. PLoS ONE 2015, 10, e0126522. [Google Scholar] [CrossRef]
- Jolly, M.K.; Somarelli, J.A.; Sheth, M.; Biddle, A.; Tripathi, S.C.; Armstrong, A.J.; Hanash, S.M.; Bapat, S.A.; Rangarajan, A.; Levine, H.; et al. Hybrid epithelial/mesenchymal phenotypes promote metastasis and therapy resistance across carcinomas. Pharmacol. Ther. 2019, 194, 161–184. [Google Scholar] [CrossRef]
- Eaton, E.N.; Reinhardt, F.; Khodor, Y.L.; Thiru, P.; Bierie, B.; Ye, X.; Burge, C.B.; Weinberg, R.A. Acquisition of a hybrid E/M state is essential for tumorigenicity of basal breast cancer cells. Proc. Natl. Acad. Sci. USA 2019, 116, 7353–7362. [Google Scholar] [CrossRef] [Green Version]
- Zavyalova, M.V.; Denisov, E.V.; Tashireva, L.A.; Savelieva, O.E.; Kaigorodova, E.V.; Krakhmal, N.V.; Perelmuter, V.M. Intravasation as a Key Step in Cancer Metastasis. Biochemistry 2019, 84, 762–772. [Google Scholar] [CrossRef] [PubMed]
- Harney, A.S.; Arwert, E.N.; Entenberg, D.; Wang, Y.; Guo, P.; Qian, B.Z.; Oktay, M.H.; Pollard, J.W.; Jones, J.G.; Condeelis, J.S. Realtime imaging reveals local, transient vascular permeability, and tumor cell intravasation stimulated by TIE2hi macrophagederived VEGFA. Cancer Discov. 2015, 5, 932–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, J.W. Macrophages define the invasive microenvironment in breast cancer. J. Leukoc. Biol. 2008, 84, 623–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, S.P.H.; Cabrera, R.M.; Segall, J.E. Tumor cell intravasation. Am. J. Physiol. Cell Physiol. 2016, 311, C1–C14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deryugina, E.I.; Kiosses, W.B. Intratumoral cancer cell intravasation can occur independent of invasion in to the adjacent stroma. Cell Rep. 2017, 19, 601–616. [Google Scholar] [CrossRef] [Green Version]
- Meng, S.; Tripathy, D.; Frenkel, E.P.; Shete, S.; Naftalis, E.Z.; Huth, J.F.; Beitsch, P.D.; Leitch, M.; Hoover, S.; Euhus, D.; et al. Circulating tumor cells in patients with breast cancer dormancy. Clin. Cancer Res. 2004, 10, 8152–8162. [Google Scholar] [CrossRef] [Green Version]
- Al-Othman, N.; Alhendi, A.; Ihbaisha, M.; Barahmeh, M.; Alqaraleh, M.; Al-Momany, B.Z. Role of CD44 in breast cancer. Breast Dis. 2020, 39, 1–13. [Google Scholar] [CrossRef]
- Baaten, B.J.; Li, C.R.; Deiro, M.F.; Lin, M.M.; Linton, P.J.; Bradley, L.M. CD44 regulates survival and memory development in Th1 cells. Immunity 2010, 32, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Abraham, B.K.; Fritz, P.; McClellan, M.; Hauptvogel, P.; Athelogou, M.; Brauch, H. Prevalence of CD44+/CD24−/low cells in breast cancer may not be associated with clinical outcome but may favor distant metastasis. Clin. Cancer Res. 2005, 11, 1154–1159. [Google Scholar]
- Tomita, H.; Tanaka, K.; Tanaka, T.; Hara, A. Aldehyde dehydrogenase 1A1 in stem cells and cancer. Oncotarget 2016, 7, 11018–11032. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ma, H.; Zhang, J.; Zhu, L.; Wang, C.; Yang, Y. Unraveling the roles of CD44/CD24 and ALDH1 as cancer stem cell markers in tumorigenesis and metastasis. Sci. Rep. 2017, 7, 13856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulaiman, A.; McGarry, S.; Han, X.; Liu, S.; Wang, L. CSCs in Breast Cancer-One Size Does Not Fit All: Therapeutic Advancesin Targeting Heterogeneous Epithelial and Mesenchymal CSCs. Cancers 2019, 11, 1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aktas, B.; Tewes, M.; Fehm, T.; Hauch, S.; Kimmig, R.; Kasimir-Bauer, S. Stem cell and epithelial–mesenchymal transition markers are frequently overexpressed in circulating tumor cells of metastatic breast cancer patients. Breast Cancer Res. 2009, 11, R46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- May, C.D.; Sphyris, N.; Evans, K.W.; Werden, S.J.; Guo, W.; Mani, S.A. Epithelial-mesenchymal transition and cancer stem cells: A dangerously dynamic duo in breast cancer progression. Breast Cancer Res. 2011, 13, 202. [Google Scholar] [CrossRef] [Green Version]
- Paula, A.D.C.; Marques, O.; Sampaio, R.; Rosa, A.M.; Garcia, J.; Rêma, A.; Faria, M.D.F.; Silva, P.; Vizcaíno, R.; Lopes, C. Characterization of CD44+/ALDH1+/Ki-67− cells in nonmalignant and neoplastic lesions of the breast. Anticancer Res. 2016, 36, 4629–4638. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Cong, Y.; Wang, D.; Sun, Y.; Deng, L.; Liu, Y.; Martin-Trevino, R.; Shang, L.; McDermott, S.P.; Landis, M.D.; et al. Breast cancer stem cells transition between epithelial and mesenchymal states reflective of their normal counterparts. Stem. Cell Rep. 2014, 2, 78–91. [Google Scholar] [CrossRef]
- Colacino, J.A.; Azizi, E.; Brooks, M.D.; Harouaka, R.; Fouladdel, S.; McDermott, S.P.; Lee, M.; Hill, D.; Madden, J.; Boerner, J.; et al. Heterogeneity of Human Breast Stem and Progenitor Cells as Revealed by Transcriptional Profiling. Stem. Cell Rep. 2018, 10, 1596–1609. [Google Scholar] [CrossRef] [Green Version]
- Liou, G.Y. CD133 as a regulator of cancer metastasis through the cancer stem cells. Int. J. Biochem. Cell Biol. 2019, 106, 1–7. [Google Scholar] [CrossRef]
- Brugnoli, F.; Grassilli, S.; Al-Qassab, Y.; Capitani, S.; Bertagnolo, V. CD133 in Breast Cancer Cells: More than a Stem Cell Marker. J. Oncol. 2019, 2019, 7512632. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Guo, S.; Liu, C.; Zhao, Y.; Feng, C.; Liu, Y.; Wang, T.; Li, C. Overexpression of SDF-1 activates the NF-κB pathway to induce epithelial to mesenchymal transition and cancer stem cell-like phenotypes of breast cancer cells. Int. J. Oncol. 2016, 48, 1085–1094. [Google Scholar] [CrossRef] [Green Version]
- Pospelova, R.A. Leuko-Concentration in Clinical Practice; Moscow Medical: Moscow, ID, USA, 1973; p. 22, (In Russ). Available online: https://sinref.ru/000_uchebniki/03200medecina/993_04_labor-metod-issledov-klinik-menshikov-1987/031.htm (accessed on 20 February 2020).
- Parra, E.R.; Uraoka, N.; Jiang, M.; Cook, P.; Gibbons, D.; Forget, M.-A.; Bernatchez, C.; Haymaker, C.; Wistuba, I.I.; Rodriguez-Canales, J. Validation of multiplex immunofluorescence panels using multispectral microscopy for immune-profiling of formalin-fixed and paraffin-embedded human tumor tissues. Sci. Rep. 2017, 7, 13380. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Parameters | General Group | CTC– | CTC+ | |
|---|---|---|---|---|
| Age (year) | 35–50 | 14/38 (37%) | 6/38 (16%) | 8/38 (21%) |
| >50 | 24/38 (63%) | 14/38 (37%) | 10/38 (26%) | |
| Molecular type of breast cancer | Luminal A | 13/38 (34%) | 9/38 (24%) | 4/38 (10%) |
| Luminal B | 22/38 (58%) | 10/38 (26%) | 12/38 (31%) | |
| HER2-positive | 1/38 (3%) | 0/38 (0%) | 1/38 (3%) | |
| Triple-negative | 2/38 (5%) | 1/38 (3%) | 1/38 (3%) | |
| Menstrual function | Premenopausal | 13/38 (34%) | 7/38 (19%) | 6/38 (16%) |
| Postmenopausal | 25/38 (66%) | 13/38 (34%) | 12/38 (31%) | |
| Tumor size | <20 mm | 15/38 (39%) | 7/38 (19%) | 8/38 (21%) |
| 20–50 mm | 23/38 (61%) | 13/38 (34%) | 10/38 (26%) | |
| Stage | I | 13/38 (34%) | 6/38 (16%) | 7/38 (18%) |
| IIA | 19/38 (50%) | 11/38 (29%) | 8/38 (21%) | |
| IIB | 5/38 (13%) | 2/38 (5%) | 3/38 (8%) | |
| No data | 1/38 (3%) | 1/38 (3%) | 0/38 (0%) | |
| Tumor grade | 1 | 5/38 (13%) | 2/38 (5%) | 3/38 (8%) |
| 2 | 25/38 (66%) | 13/38 (34%) | 12/38 (32%) | |
| 3 | 3/38 (8%) | 2/38 (5%) | 1/38 (3%) | |
| No data | 5/38 (13%) | 3/38 (8%) | 2/38 (5%) | |
| Estrogen receptor | Positive | 35/38 (92%) | 19/38 (50%) | 16/38 (42%) |
| Negative | 3/38 (8%) | 1/38 (3%) | 2/38 (5%) | |
| Progesterone receptor | Positive | 28/38 (74%) | 14/38 (37%) | 14/38 (37%) |
| Negative | 10/38 (26%) | 6/38 (16%) | 4/38 (10%) | |
| HER2/neu | Positive | 7/38 (18%) | 3/38 (8%) | 4/38 (10%) |
| Negative | 31/38 (82%) | 17/38 (45%) | 14/38 (37%) | |
| Ki67 expression | <20% | 13/38 (34%) | 9/38 (24%) | 4/38 (10%) |
| >20% | 25/38 (66%) | 11/38 (29%) | 14/38 (37%) | |
| Lymph-node status | Yes | 28/38 (74%) | 14/38 (37%) | 14/38 (37%) |
| No | 7/38 (18%) | 3/38 (8%) | 4/38 (10%) | |
| No surgery | 3/38 (8%) | 3/38 (8%) | 0/38 (0%) | |
| Distant metastasis | Yes | 0/38 (0%) | 0/38 (0%) | 0/38 (0%) |
| No | 38/38 (100%) | 20/38 (53%) | 18/38 (47%) | |
| Intraoperative radiotherapy | Yes | 22/38 (58%) | 11/38 (29%) | 11/38 (29%) |
| No | 11/38 (29%) | 5/38 (13%) | 6/38 (16%) | |
| No data | 5/38 (13%) | 4/38 (10%) | 1/38 (3%) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savelieva, O.E.; Tashireva, L.A.; Kaigorodova, E.V.; Buzenkova, A.V.; Mukhamedzhanov, R.K.; Grigoryeva, E.S.; Zavyalova, M.V.; Tarabanovskaya, N.A.; Cherdyntseva, N.V.; Perelmuter, V.M. Heterogeneity of Stemlike Circulating Tumor Cells in Invasive Breast Cancer. Int. J. Mol. Sci. 2020, 21, 2780. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082780
Savelieva OE, Tashireva LA, Kaigorodova EV, Buzenkova AV, Mukhamedzhanov RK, Grigoryeva ES, Zavyalova MV, Tarabanovskaya NA, Cherdyntseva NV, Perelmuter VM. Heterogeneity of Stemlike Circulating Tumor Cells in Invasive Breast Cancer. International Journal of Molecular Sciences. 2020; 21(8):2780. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082780
Chicago/Turabian StyleSavelieva, Olga E., Liubov A. Tashireva, Evgeniya V. Kaigorodova, Angelina V. Buzenkova, Rustam Kh. Mukhamedzhanov, Evgeniya S. Grigoryeva, Marina V. Zavyalova, Natalia A. Tarabanovskaya, Nadezhda V. Cherdyntseva, and Vladimir M. Perelmuter. 2020. "Heterogeneity of Stemlike Circulating Tumor Cells in Invasive Breast Cancer" International Journal of Molecular Sciences 21, no. 8: 2780. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082780