Cerato-Platanins from Marine Fungi as Effective Protein Biosurfactants and Bioemulsifiers

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Fungal Growth in Oil as a Carbon Source and Protein Secretion
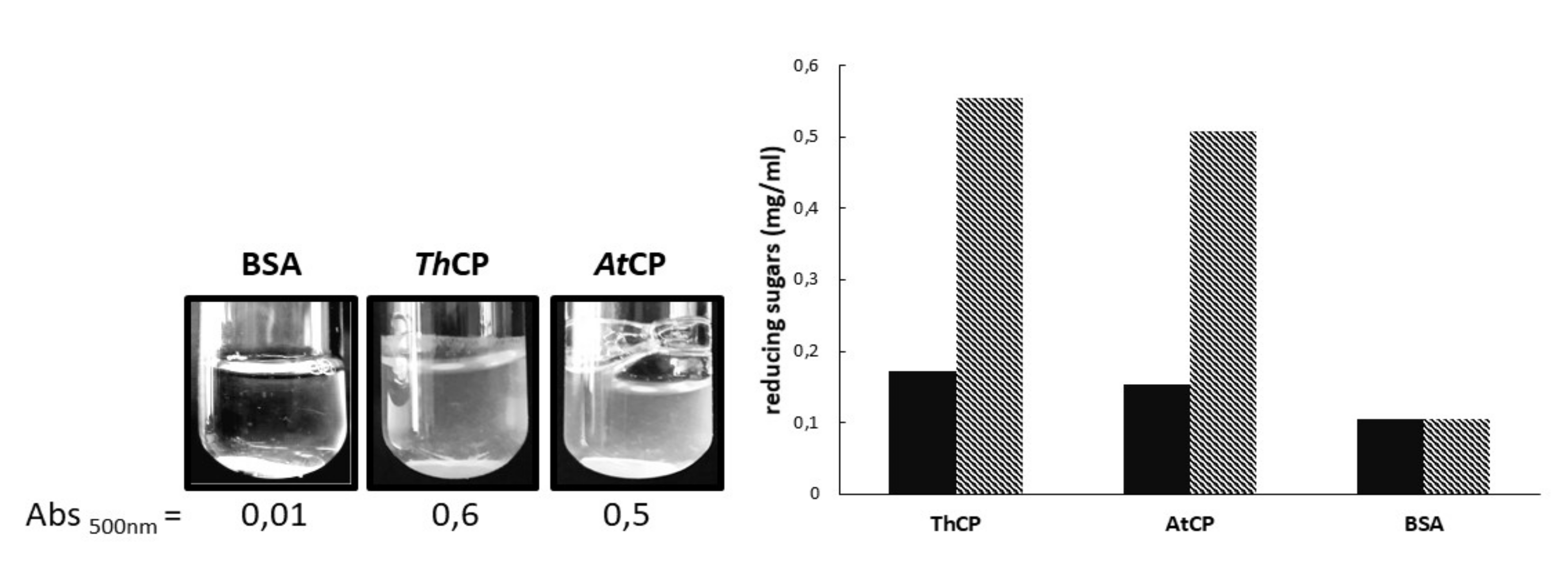
2.2. Emulsification Tests
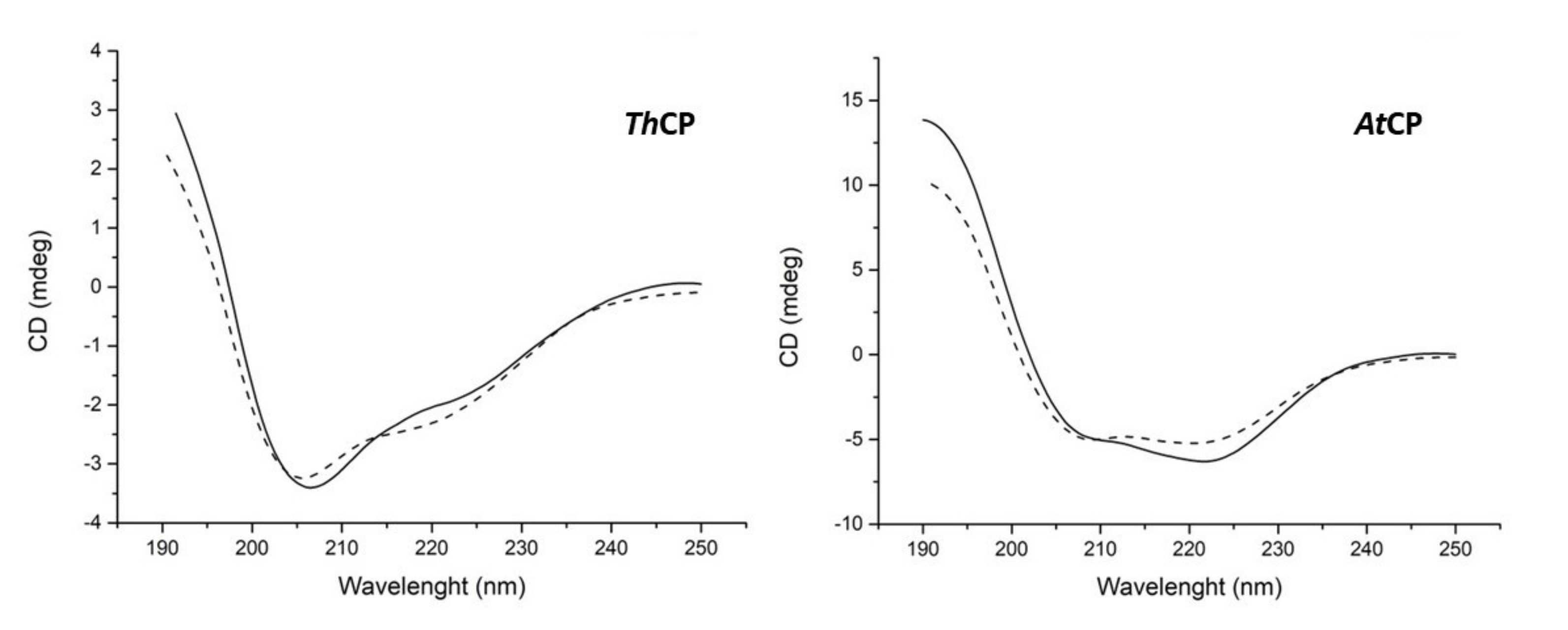
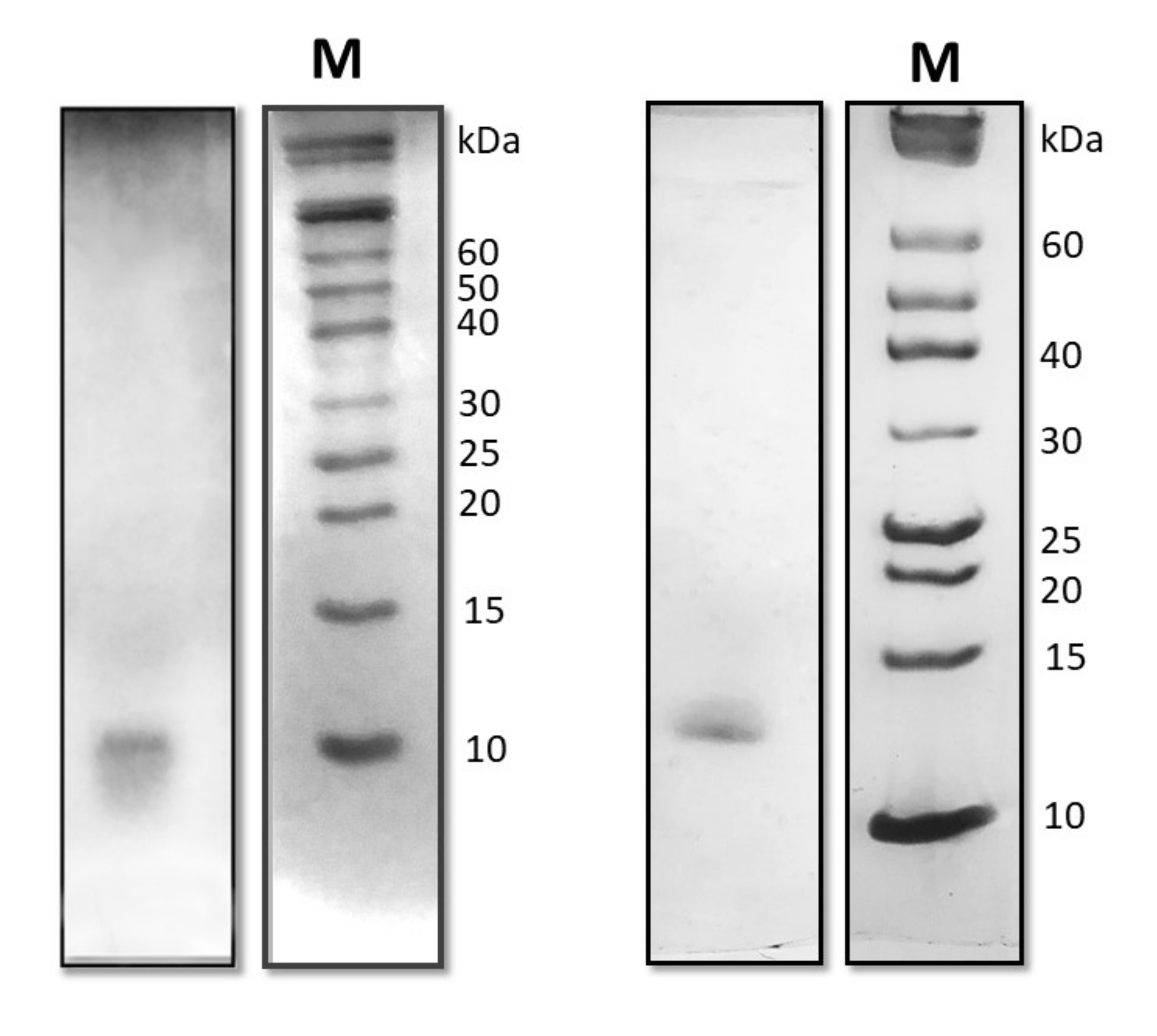
2.3. Protein Identification and Characterization
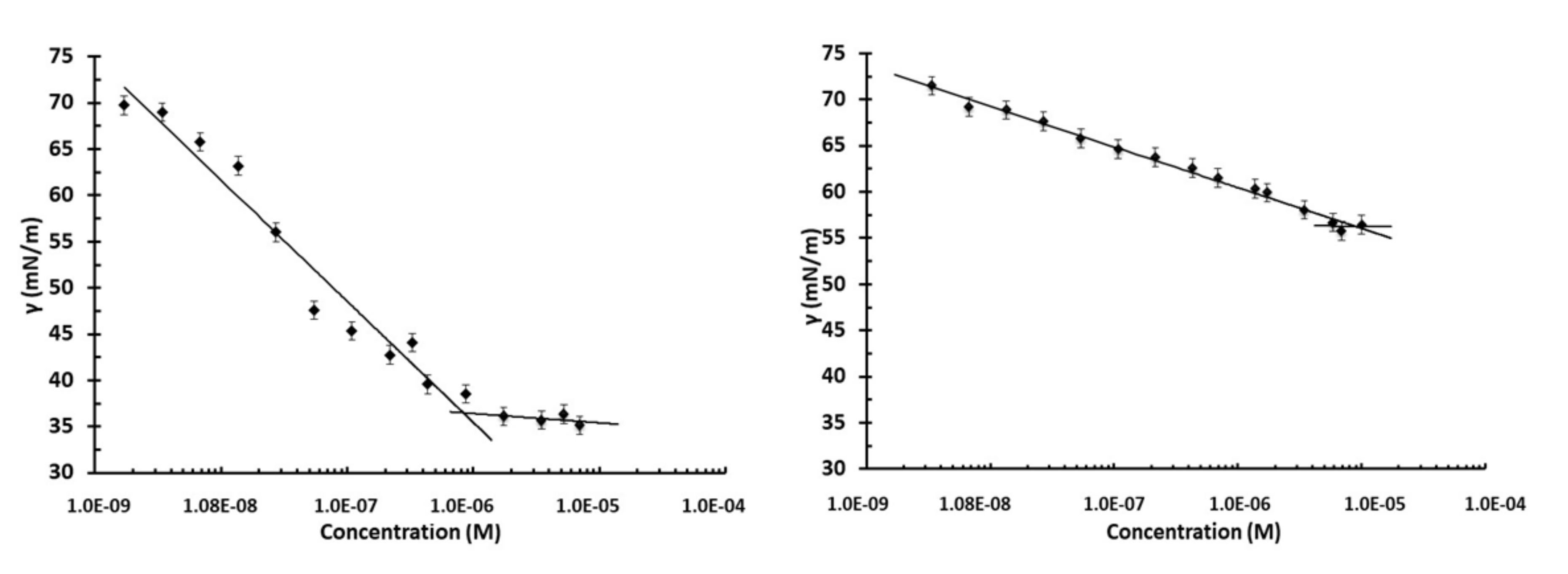
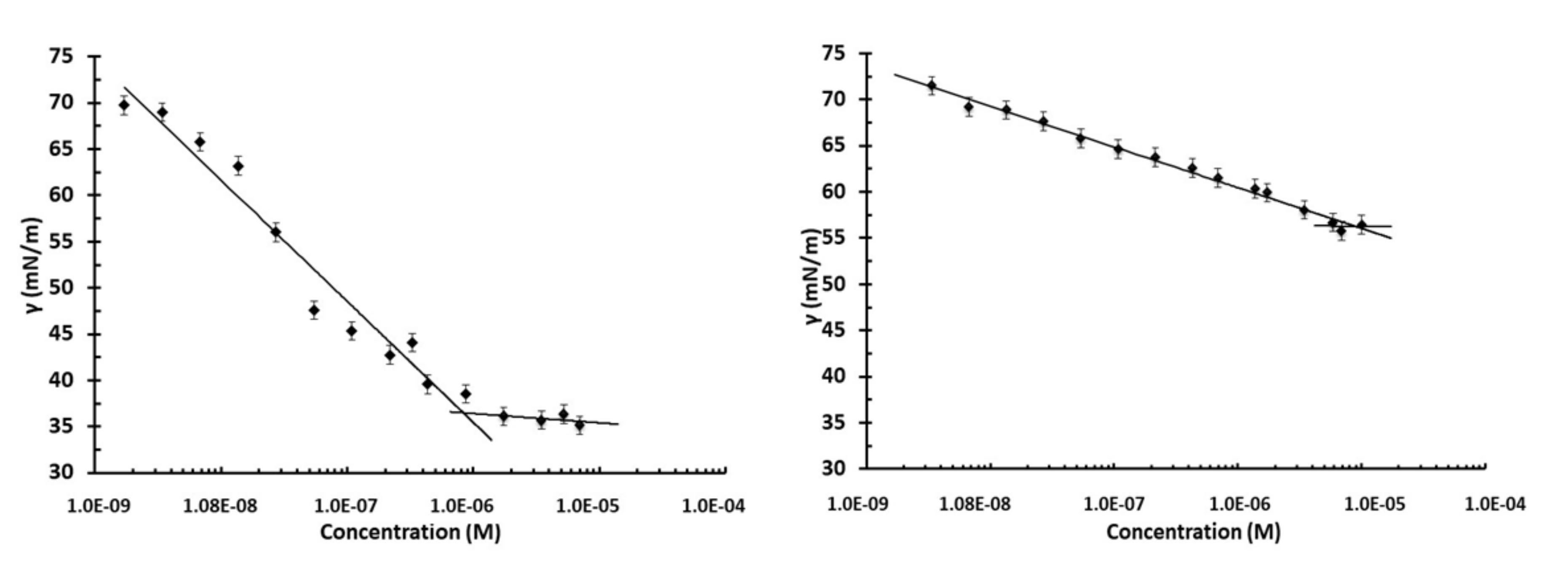
2.4. Surface Tension Measurements
2.5. Cellulose Loosening
2.6. Water Contact Angle (WCA) Measurement
3. Discussion
4. Materials and Methods
4.1. Fungal Growth
4.2. Purification of the Proteins
4.3. Emulsification Activity
4.4. Protein Identification by Mass Spectrometry
4.5. Circular Dichroism (CD) Spectroscopy
4.6. Surface Tension Measurement
4.7. Cellulose Loosening and Cellulolytic Activity Assay
4.8. Water Contact Angle Determination
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Raghukumar, C. Marine fungal biotechnology: An ecological perspective. Fungal Divers. 2008, 31, 19–36. [Google Scholar]
- Amend, A.; Heitman, J.; Hom, E.F.Y.; Ianiri, G.; Jones, A.C.; Kagami, M.; Picard, K.T.; Quandt, C.A.; Raghukumar, S.; Riquelme, M.; et al. Fungi in the Marine Environment: Open Questions and Unsolved Problems. Ecol. Evol. Sci. 2019, 10, 1–15. [Google Scholar]
- Mcgenity, T.J.; Folwell, B.D.; Mckew, B.A.; Sanni, G.O. Marine crude-oil biodegradation: A central role for interspecies interactions. Aquat. Biosyst. 2012, 8, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Bovio, E.; Gnavi, G.; Prigione, V.; Spina, F.; Denaro, R.; Yakimov, M.; Calogero, R.; Crisa, F.; Cristina, G. The culturable mycobiota of a Mediterranean marine site after an oil spill: Isolation, identification and potential application in bioremediation. Sci. Total Environ. 2017, 576, 310–318. [Google Scholar] [CrossRef]
- Perfumo, A.; Smyth, T.J.P.; Marchant, R.; Banat, I.M. Production and Roles of Biosurfactants and Bioemulsifiers in Accessing Hydrophobic Substrates. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer-Verlag: Heidelberg, Germany, 2010; pp. 1501–1512. [Google Scholar]
- Kubicki, S.; Bollinger, A.; Katzke, N.; Jaeger, K.; Loeschcke, A.; Thies, S. Marine Biosurfactants: Biosynthesis, Structural Diversity and Biotechnological Applications. Mar. Drugs 2019, 17, 408. [Google Scholar] [CrossRef] [Green Version]
- Uzoigwe, C.; Burgess, J.G.; Ennis, C.J.; Rahman, P.K.S.M. Bioemulsifiers are not biosurfactants and require different screening approaches. Front. Microbiol. 2015, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Lo, V.; Lai, J.I.; Sunde, M. Fungal Hydrophobins and Their Self-Assembly into Functional Nanomaterials. Adv. Exp. Med. Biol. 2019, 1174, 161–185. [Google Scholar]
- Cicatiello, P.; Sorrentino, I.; Piscitelli, A.; Giardina, P. Spotlight on Class I hydrophobins: Theri intriguing biochemical properties and industrial prospects. In Grand Challenges In Funga lBiotechnology; Nevalainen, H., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 333–348. [Google Scholar]
- Sunde, M.; Pham, C.L.L.; Kwan, A.H. Molecular Characteristics and Biological Functions of Surface-Active and Surfactant Proteins. Annu. Rev. Biochem. 2017, 86, 585–608. [Google Scholar] [CrossRef] [PubMed]
- Baccelli, I. Cerato-platanin family proteins: One function for multiple biological roles? Front. Plant Sci. 2015, 5, 2013–2016. [Google Scholar] [CrossRef] [PubMed]
- Frischmann, A.; Neudl, S.; Gaderer, R.; Bonazza, K.; Zach, S.; Gruber, S.; Spadiut, O.; Friedbacher, G.; Grothe, H.; Seidl-seiboth, V. Self-assembly at Air/Water Interfaces and Carbohydrate Binding Properties of the Small Secreted Protein EPL1 from the fungus Trichoderma atroviride. J. Biol. Chem. 2013, 288, 4278–4287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaderer, R.; Bonazza, K. Cerato-platanins: A fungal protein family with intriguing properties and application potential. Appl. Microbiol. Biotechnol. 2014, 98, 4795–4803. [Google Scholar] [CrossRef] [Green Version]
- Bonazza, K.; Gaderer, R.; Neudl, S.; Przylucka, A.; Allmaier, G.; Druzhinina, I.S. The fungal cerato-platanin protein EPL1 forms highly ordered layers at hydrophobic/hydrophilic interfaces. Soft Matter 2015, 11, 1723–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, A.L.; Gallo, M.; Pazzagli, L.; Benedetti, C.E.; Cappugi, G.; Scala, A.; Pantera, B.; Spisni, A.; Pertinhez, T.A.; Cicero, D.O. The Structure of the Elicitor Cerato-platanin (CP), the First Member of the CP Fungal Protein Family, Reveals a Double ψβ-Barrel Fold and Carbohydrate Binding. J. Biol. Chem. 2011, 286, 17560–17568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baccelli, I.; Luti, S.; Bernardi, R. Cerato-platanin shows expansin-like activity on cellulosic materials. Appl. Microbiol. Biotechnol. 2014, 98, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Arantes, V.; Saddler, J.N. Access to cellulose limits the efficiency of enzymatic hydrolysis: The role of amorphogenesis. Biotechnol. Biofuels 2010, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Djonovic, S.; Vargas, W.A.; Kolomiets, M.V.; Horndeski, M. A Proteinaceous Elicitor Sm1 from the Beneficial Fungus Trichoderma virens Is Required for Induced Systemic Resistance in Maize. Plant Physiol. 2007, 145, 875–889. [Google Scholar] [CrossRef] [Green Version]
- Blesic, M.; Dichiarante, V.; Milani, R.; Linder, M. Evaluating the potential of natural surfactants in the petroleum industry: The case of hydrophobins. Pure Appl. Chem. 2017, 90, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Kovalchuk, A.; Keriö, S.; Asiegbu, F.O.; Chen, H.; Kovalchuk, A.; Keriö, S.; Asiegbu, F.O. Distribution and bioinformatic analysis of the cerato-platanin protein family in Dikarya. Mycologia 2017, 5514, 1479–1488. [Google Scholar] [CrossRef]
- Cicatiello, P.; Gravagnuolo, A.M.; Gnavi, G.; Varese, G.C.; Giardina, P. Marine fungi as source of new hydrophobins. Int. J. Biol. Macromol. 2016, 92, 1229–1233. [Google Scholar] [CrossRef]
- Mujumdar, S.; Joshi, P.; Karve, N. Production, characterization, and applications of bioemulsifiers (BE) and biosurfactants (BS) produced by Acinetobacter spp.: A review. J. Basic. Microbiol. 2019, 59, 277–287. [Google Scholar] [CrossRef]
- Cicatiello, P.; Stanzione, I.; Dardano, P.; De Stefano, L.; Birolo, L.; De Chiaro, A.; Monti, D.M.; Petruk, G.; D’Errico, G.; Giardina, P. Characterization of a Surface-Active Protein Extracted from a Marine Strain of Penicillium chrysogenum. Int. J. Biol. Macromol. 2019, 20, 3242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pazzagli, L.; Cappugi, G.; Manao, G.; Camici, G.; Santini, A.; Scala, A. Purification, Characterization, and Amino Acid Sequence of Cerato-platanin, a New Phytotoxic Protein from Ceratocystis fimbriata f. sp. platani. J. Biol. Chem. 1999, 274, 24959–24964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martellini, F.; Faoro, F.; Carresi, L.; Pantera, B. Cerato-Populin and Cerato-Platanin, Two Non-Catalytic Proteins from Phytopathogenic Fungi, Interact with Hydrophobic Inanimate Surfaces and Leaves. Mol. Biotechnol. 2013, 55, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Pazzagli, L.; Zoppi, C.; Carresi, L.; Tiribilli, B.; Sbrana, F.; Schiff, S.; Pertinhez, T.A.; Scala, A.; Cappugi, G. Characterization of ordered aggregates of cerato-platanin and their involvement in fungus–host interactions. BBA-Gen. Subj. 2009, 1790, 1334–1344. [Google Scholar] [CrossRef]
- Barsottini, M.R.D.O.; Oliveira, J.F.; De Adamoski, D.; Teixeira, P.J.P.L.; Prado, P.F.V.; Tiezzi, H.O.; Sforça, M.L.; Cassago, A.; Oliveira, P.S.L.; De Zeri, A.C.D.M.; et al. Functional Diversification of Cerato-Platanins in Moniliophthora perniciosa as Seen by Differential Expression and Protein Function Specialization. MPMI 2013, 26, 1281–1293. [Google Scholar] [CrossRef] [Green Version]
- Gravagnuolo, A.M.; Longobardi, S.; Luchini, A.; Appavou, M.; Stefano, L.; De Notomista, E.; Paduano, L.; Giardina, P. Class I Hydrophobin Vmh2 Adopts Atypical Mechanisms to Self-Assemble into Functional Amyloid Fibrils. Biomacromolecules 2016, 17, 954–964. [Google Scholar] [CrossRef] [Green Version]
- Cicatiello, P.; Dardano, P.; Pirozzi, M.; Gravagnuolo, A.M.; Stefano, L.; De Giardina, P. Self-Assembly of Two Hydrophobins from Marine Fungi Affected by Interaction With Surfaces. Biotechnol. Bioeng. 2017, 114, 2173–2186. [Google Scholar] [CrossRef]
- Dyksterhouse, S.E.; Gray, J.P.; Herwig, R.P.; Lara, J.C.; Staley, J.T. Hydrocarbon-Degrading Bacterium from Marine Sediments. Int. J. Syst. Bacteriol. 1995, 45, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Whitmore, L.; Wallace, B.A. Protein secondary structure analyses from circular dichroism spectroscopy: Methods and reference databases. Biopolymers 2007, 89, 392–400. [Google Scholar] [CrossRef]
- Russo, I.; Imperatore, R.; Santis, A.; De Luchini, A.; Paduano, L.; Errico, G.D. Structure and dynamics of cetyltrimethylammonium chloride-sodium dodecylsulfate (CTAC-SDS ) catanionic vesicles: High-value nano-vehicles from low-cost surfactants. J. Colloid Interface Sci. 2017, 501, 112–122. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Protein namea,b (Entry Code) | Family | Peptides (Ion Score) | Sequence Coverage (Number of Peptides) |
|---|---|---|---|---|
| T. harzianum | ThCPa SnodProt1b (gi|818166392) | Cerato-platanins | R.YHWSTQGQIPR.F (25) | 45% (4) |
| R.SLNVVSCSDGPNGLETR.Y (70) | ||||
| R.FPYIGGVQAVAGWNSASCGTCWK.L (28) | ||||
| R.VSATASQVAVK.N (57) | ||||
| A. terreus | AtCPa allergen Asp f 15b (gi|115384120) | Cerato-platanins | K.LTYGGK.S (29) | 31% (4) |
| K.WPTFGSVPK.F (26) | ||||
| R.VQATYQEVAK.S (54) | ||||
| K.FPHIGGSPTIPGWNSPNCGK.C (35) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pitocchi, R.; Cicatiello, P.; Birolo, L.; Piscitelli, A.; Bovio, E.; Varese, G.C.; Giardina, P. Cerato-Platanins from Marine Fungi as Effective Protein Biosurfactants and Bioemulsifiers. Int. J. Mol. Sci. 2020, 21, 2913. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082913
Pitocchi R, Cicatiello P, Birolo L, Piscitelli A, Bovio E, Varese GC, Giardina P. Cerato-Platanins from Marine Fungi as Effective Protein Biosurfactants and Bioemulsifiers. International Journal of Molecular Sciences. 2020; 21(8):2913. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082913
Chicago/Turabian StylePitocchi, Rossana, Paola Cicatiello, Leila Birolo, Alessandra Piscitelli, Elena Bovio, Giovanna Cristina Varese, and Paola Giardina. 2020. "Cerato-Platanins from Marine Fungi as Effective Protein Biosurfactants and Bioemulsifiers" International Journal of Molecular Sciences 21, no. 8: 2913. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082913