Use of 3D Human Liver Organoids to Predict Drug-Induced Phospholipidosis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
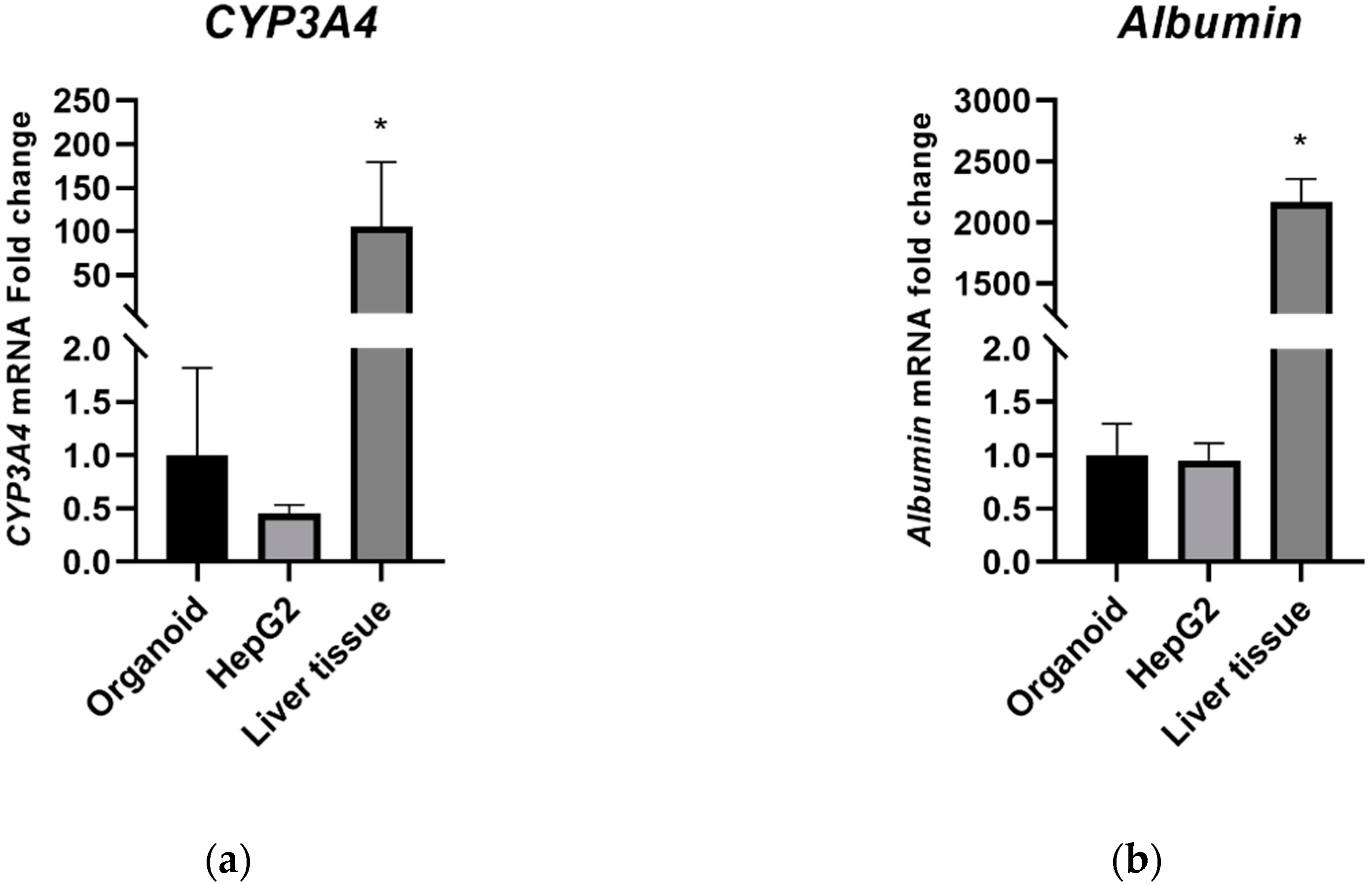
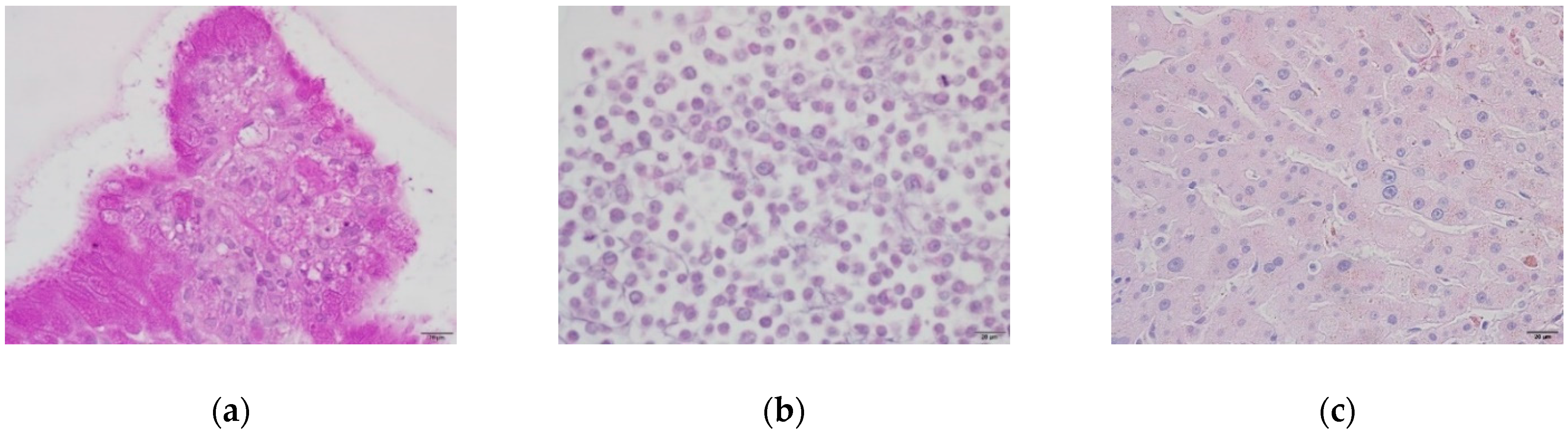
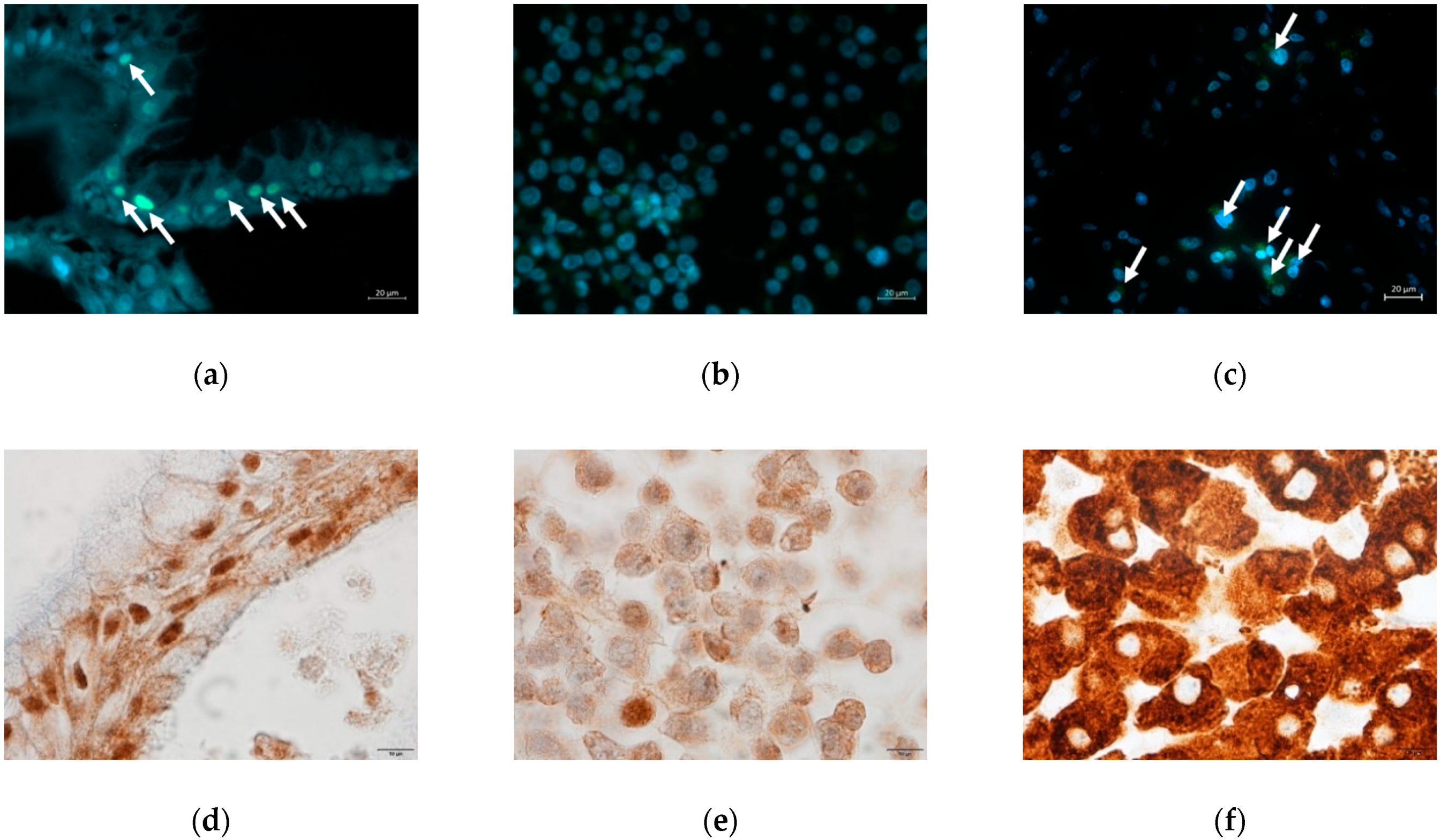
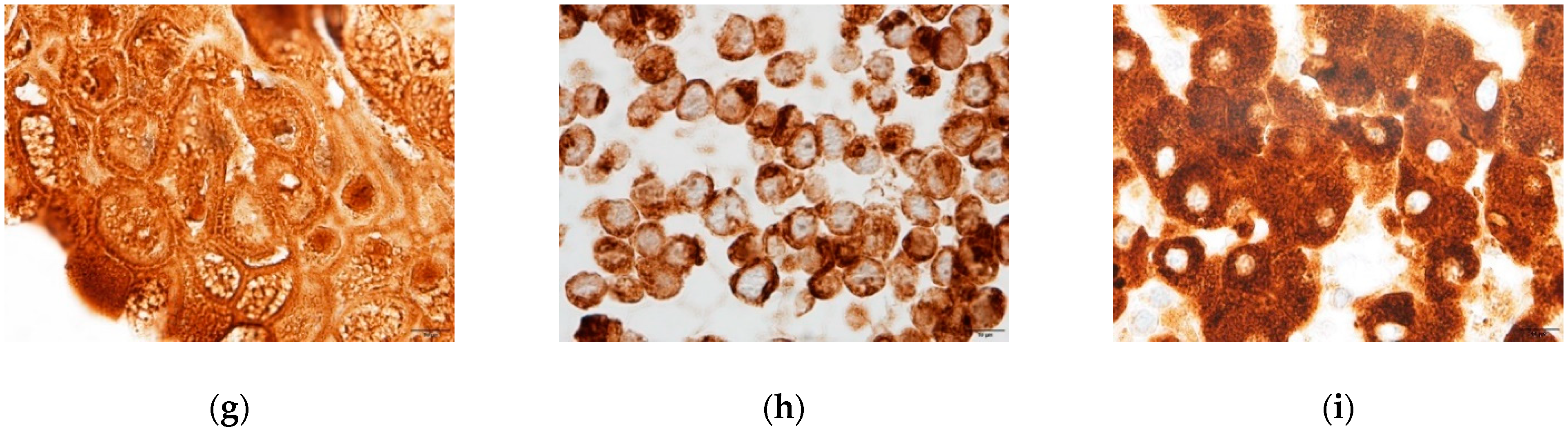
2.1. Liver Cell Characterization
2.1.1. Hepatic Marker Expression
2.1.2. Glycogen Storage
2.1.3. Hepatic Protein Expression
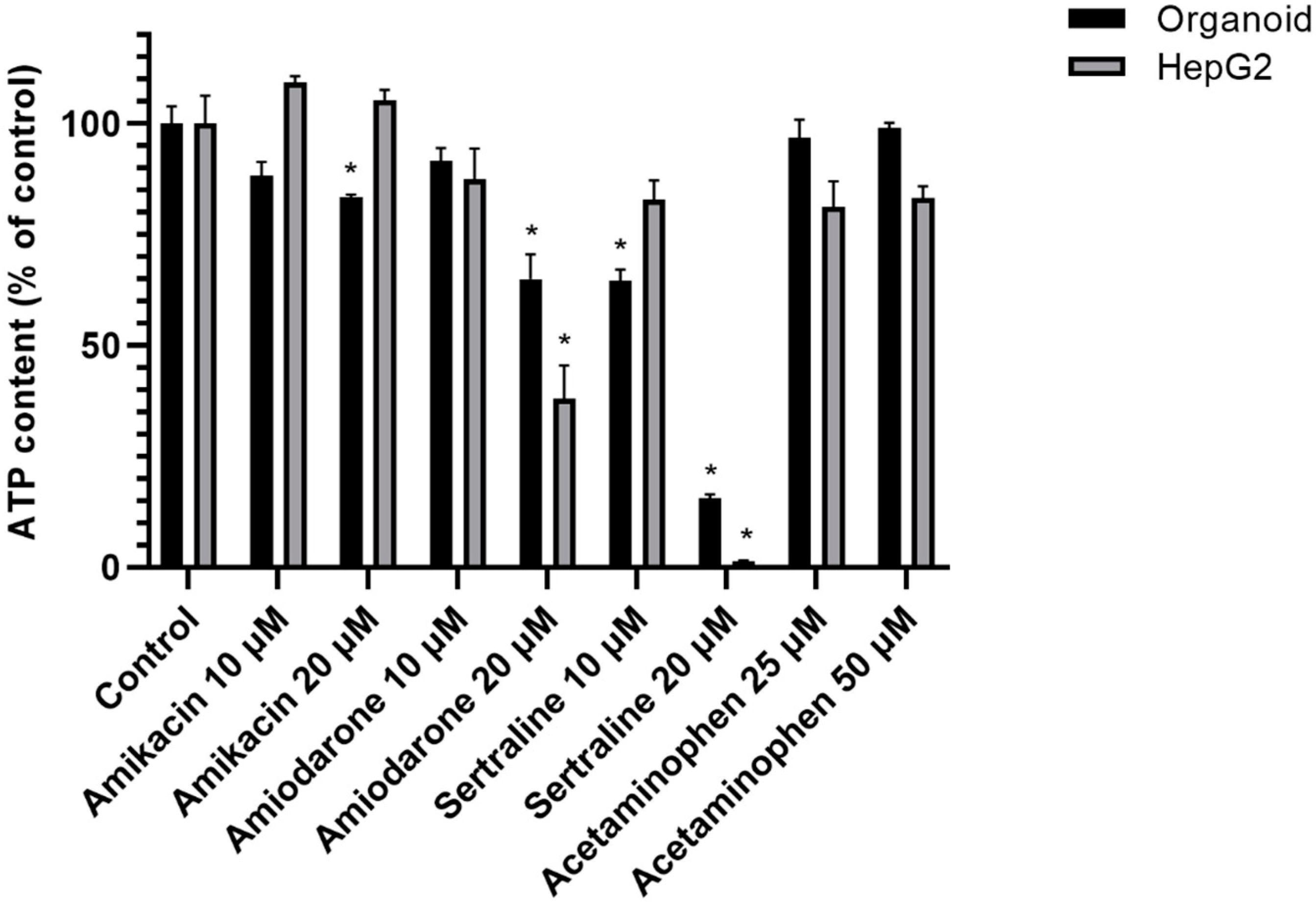
2.2. Cell Viability Changes
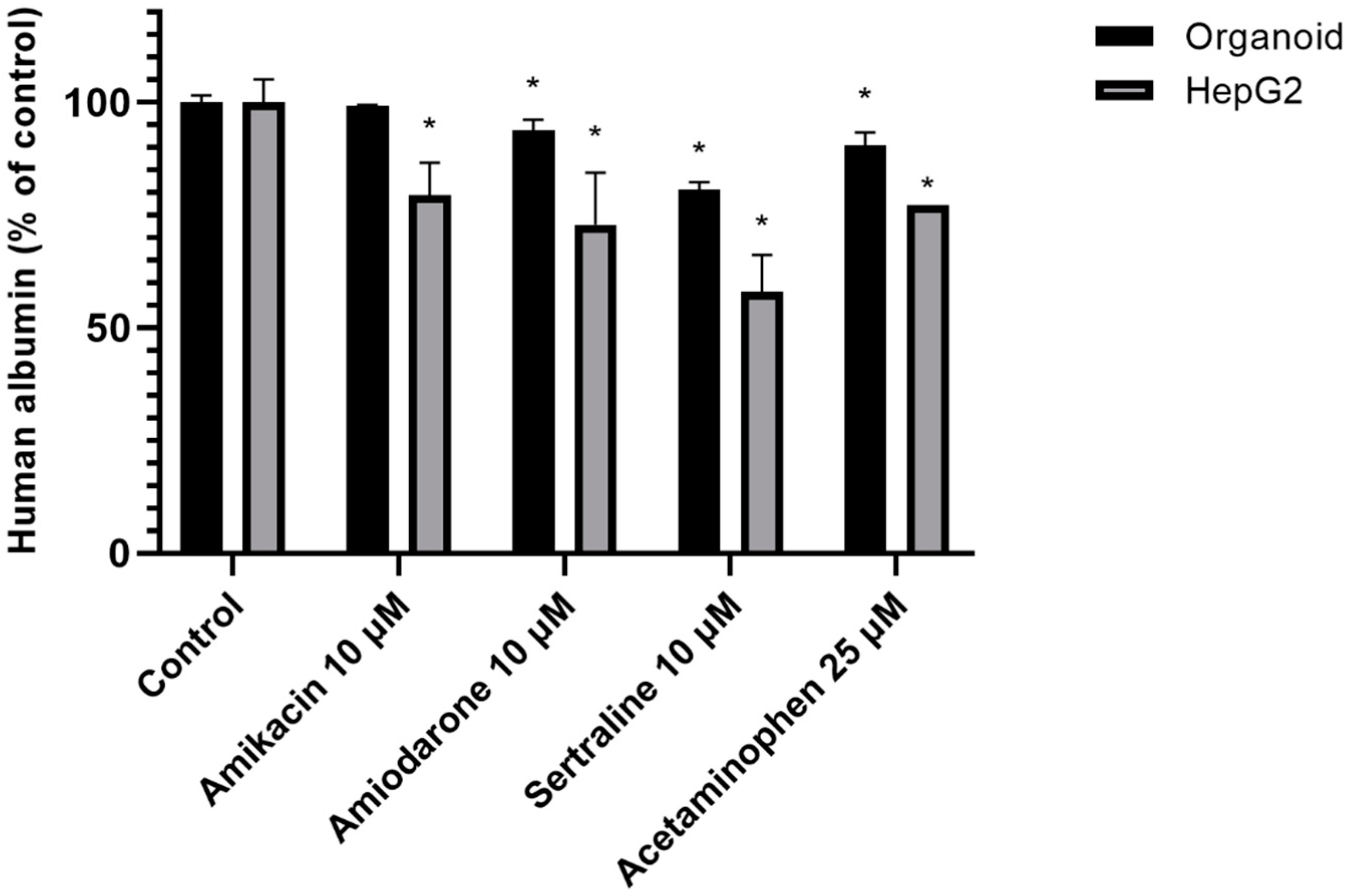
2.3. Albumin Secretion Content
2.4. Evaluation of Drug-Induced Phospholipidosis
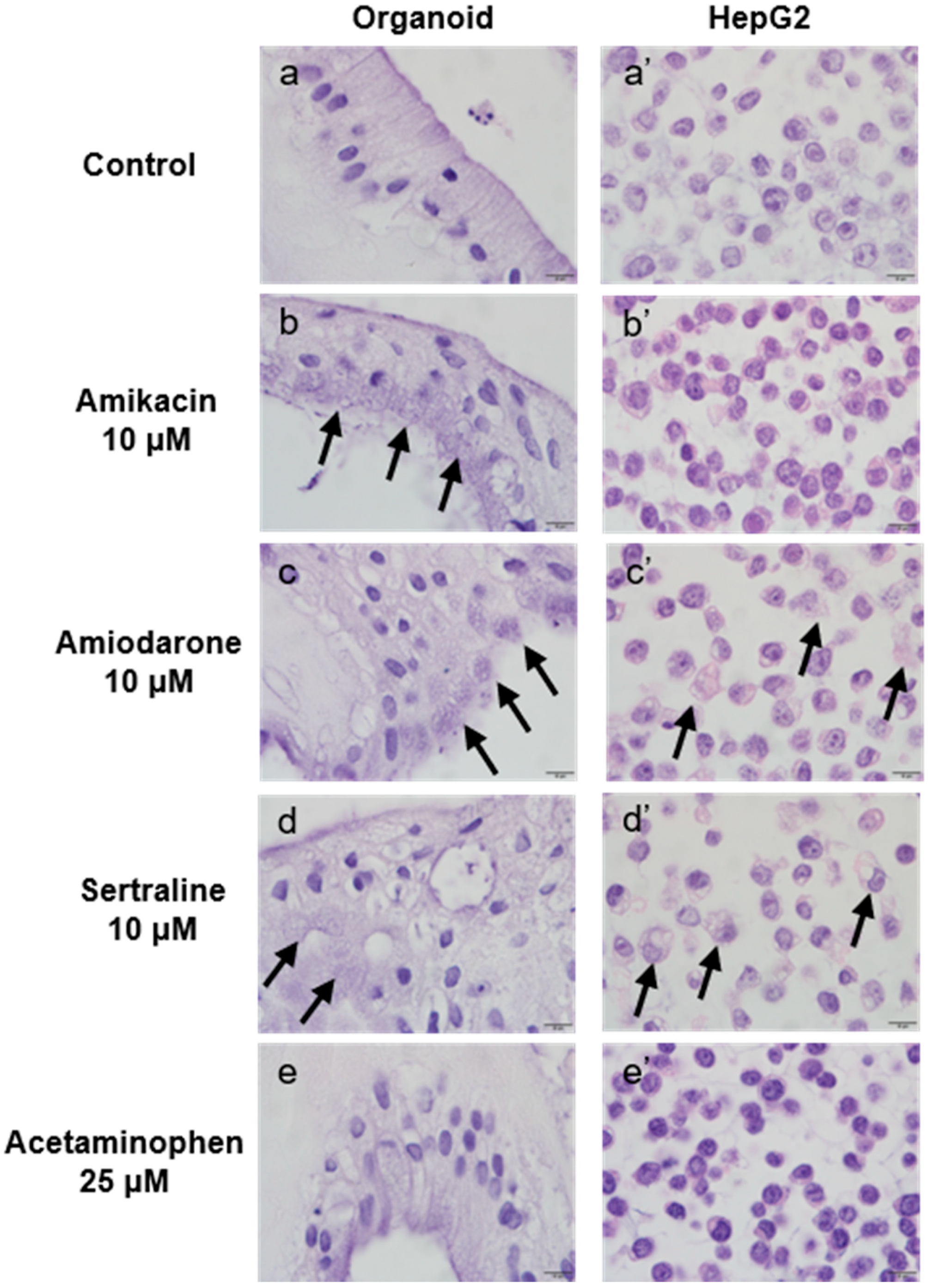
2.4.1. Morphological Changes
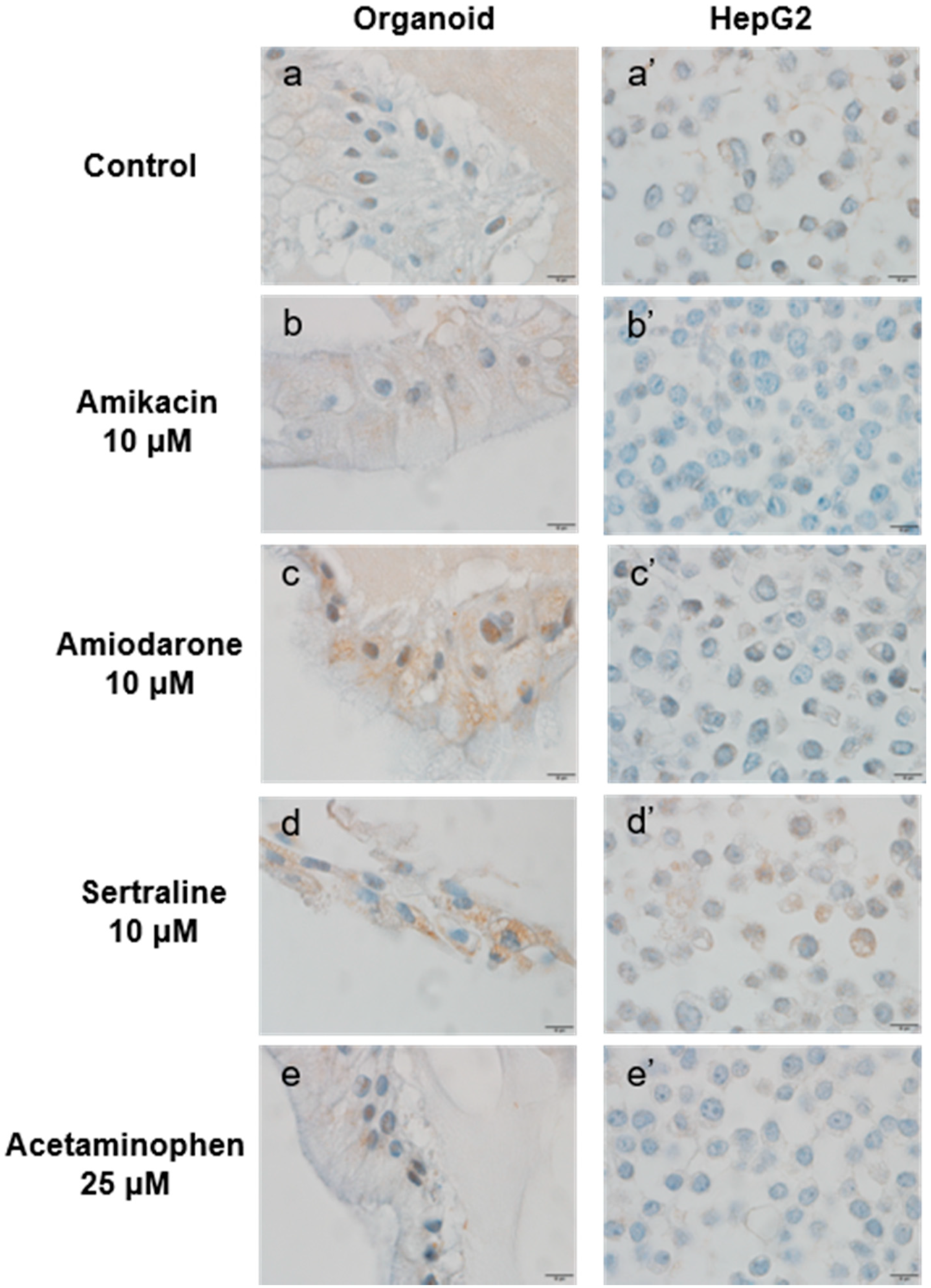
2.4.2. Comparison of LAMP-2 Expression
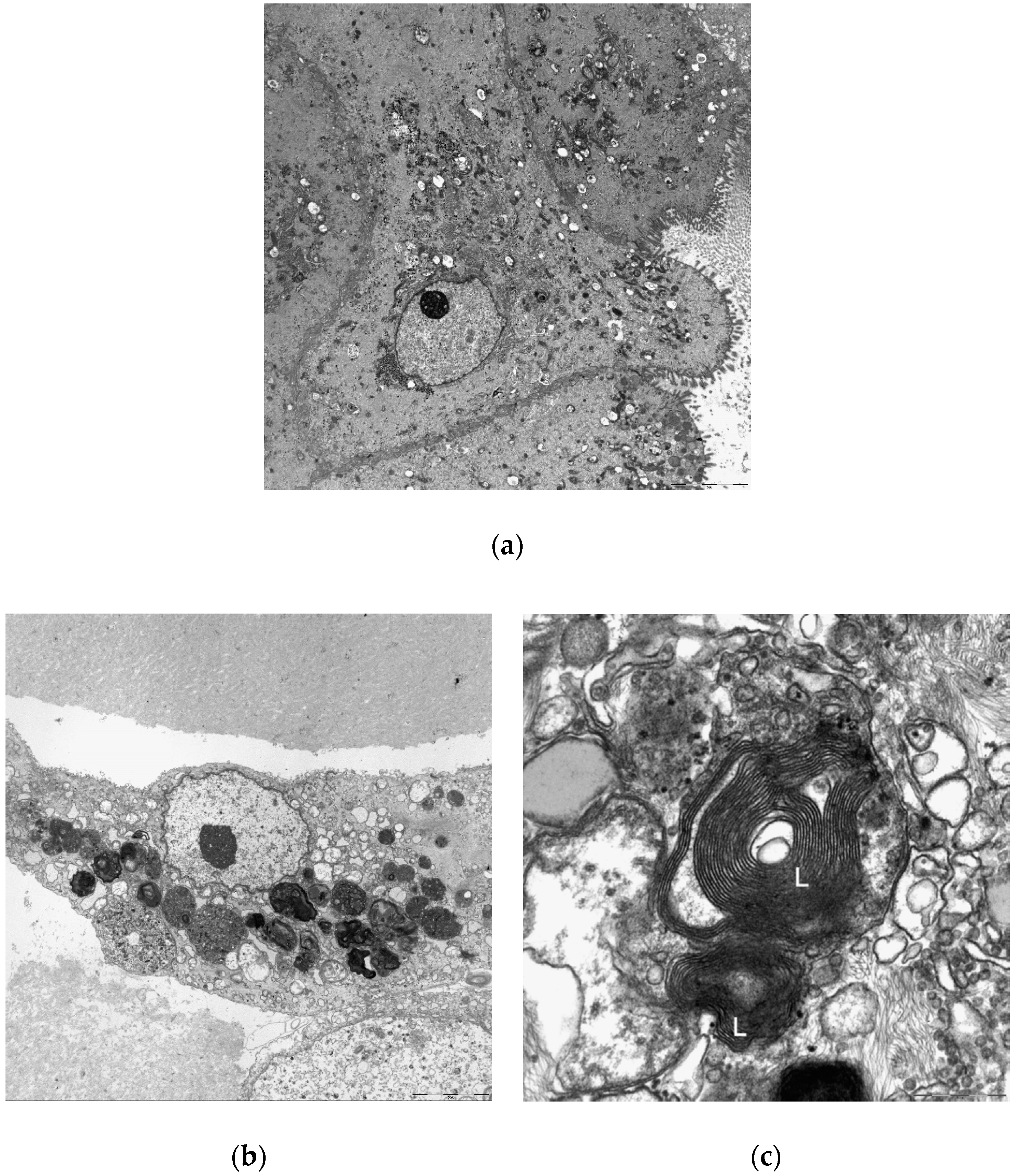
2.4.3. Confirmation of Drug-Induced Phospholipidosis
2.4.4. Gene Expression Changes under Conditions of Drug-Induced Phospholipidosis
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.2.1. Human Liver Organoids
4.2.2. HepG2 Cells
4.3. Treatment of Cells
4.4. RT-qPCR
4.5. PAS Staining
4.6. Immunostaining (Immunofluorescence and Immunohistochemistry)
4.7. Histological Examination
4.8. Detection of Lysosome-Associated Membrane Protein 2 (LAMP-2)
4.9. Cell Viability Assay
4.10. Albumin Content Measurement
4.11. Identification of Lamellar Bodies
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 3D | three-dimensional |
| 2D | two-dimensional |
| PL | phospholipidosis |
| HBSS | Hanks’ Balanced Salt Solution |
| EBSS | Earle’s Balanced Salt Solution |
| BME 2 | Basement Membrane Extract |
| EM | expansion medium |
| DM | differentiation medium |
| FBS | fetal bovine serum |
| PBS | phosphate-buffered saline |
| LAMP-2 | lysosome-associated membrane protein 2 |
| qPCR | quantitative real-time polymerase chain reaction |
| RT | reverse transcription |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| PAS | Periodic Acid Schiff |
References
- Asaoka, Y.; Togashi, Y.; Imura, N.; Sai, T.; Miyoshi, T.; Miyamoto, Y. Immunohistochemistry of LAMP-2 and adipophilin for phospholipidosis in liver and kidney in ketoconazole-treated mice. Exp. Toxicol. Pathol. 2013, 65, 817–823. [Google Scholar] [CrossRef]
- Gräbner, R. Influence of cationic amphiphilic drugs on the phosphatidylcholine hydrolysis by phospholipase A2. Biochem. Pharmacol. 1987, 36, 1063–1067. [Google Scholar] [CrossRef]
- Halliwell, W.H. Cationic Amphiphilic Drug-Induced Phospholipidosis. Toxicol. Pathol. 1997, 25, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Reasor, M.J.; Kacew, S. Drug-induced phospholipidosis: Are there functional consequences? Exp. Biol. Med. 2001, 226, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, A.M.; Krise, J. Lysosomal Sequestration of Amine-Containing Drugs: Analysis and Therapeutic Implications. J. Pharm. Sci. 2007, 96, 729–746. [Google Scholar] [CrossRef]
- Lenz, B.; Braendli-Baiocco, A.; Engelhardt, J.; Fant, P.; Fischer, H.; Francke, S.; Fukuda, R.; Groters, S.; Harada, T.; Harleman, H.; et al. Characterizing Adversity of Lysosomal Accumulation in Nonclinical Toxicity Studies: Results from the 5th ESTP International Expert Workshop; SAGE Publications Sage CA: Los Angeles, CA, USA, 2018. [Google Scholar]
- Reasor, M.J.; Hastings, K.L.; Ulrich, R.G. Drug-induced phospholipidosis: Issues and future directions. Expert Opin. Drug Saf. 2006, 5, 567–583. [Google Scholar] [CrossRef] [PubMed]
- Laurent, G.; Kishore, B.; Tulkens, P.M. Aminoglycoside-induced renal phospholipidosis and nephrotoxicity. Biochem. Pharmacol. 1990, 40, 2383–2392. [Google Scholar] [CrossRef]
- Terry, R.D.; Weiss, M. Studies in Tay-Sachs disease. II. Ultrastructure of the cerebrum. J. Neuropathol. Exp. Neurol. 1963, 22, 18–55. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, S.S.; Vethamany, V.G.; Schneck, L.; Volk, B.W. Fine structure and histochemistry of peripheral blood cells in Niemann-Pick disease. Lab. Investig. 1967, 17, 155–170. [Google Scholar]
- Rigas, B.; E Rosenfeld, L.; Barwick, K.W.; Enriquez, R.; Helzberg, J.; Batsford, W.P.; E Josephson, M.; Riely, C.A. Amiodarone hepatotoxicity: A clinicopathologic study of five patients. Ann. Intern. Med. 1986, 104, 348–351. [Google Scholar] [CrossRef]
- Shepherd, N.A.; Dawson, A.M.; Crocker, P.R.; Levison, D.A. Granular cells as a marker of early amiodarone hepatotoxicity: A pathological and analytical study. J. Clin. Pathol. 1987, 40, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Xia, M.; Shahane, S.A.; Jadhav, A.; Austin, C.P.; Huang, R. Are hERG channel blockers also phospholipidosis inducers? Bioorg. Med. Chem. Lett. 2013, 23, 4587–4590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slavov, S.; Stoyanova-Slavova, I.; Li, S.; Zhao, J.; Huang, R.; Beger, R.D.; Xia, M. Why are most phospholipidosis inducers also hERG blockers? Arch. Toxicol. 2017, 91, 3885–3895. [Google Scholar] [CrossRef] [PubMed]
- Kacew, S.; Bergeron, M.G. Pathogenic factors in aminoglycoside-induced nephrotoxicity. Toxicol. Lett. 1990, 51, 241–259. [Google Scholar] [CrossRef]
- Mingeot-Leclercq, M.-P.; Tulkens, P.M. Aminoglycosides: Nephrotoxicity. Antimicrob. Agents Chemother. 1999, 43, 1003–1012. [Google Scholar] [CrossRef] [Green Version]
- Al-Shammari, B.; Khalifa, M.; Bakheet, S.A.; Yasser, M. A Mechanistic Study on the Amiodarone-Induced Pulmonary Toxicity. Oxidative Med. Cell. Longev. 2016, 2016, 6265853. [Google Scholar] [CrossRef] [Green Version]
- Martin, W.J.; Standing, J.E. Amiodarone pulmonary toxicity: Biochemical evidence for a cellular phospholipidosis in the bronchoalveolar lavage of human subjects. J. Pharmacol. Exp. Ther. 1988, 244, 774–779. [Google Scholar]
- D’Amico, D.J.; Kenyon, K.R.; Ruskin, J.N. Amiodarone keratopathy: Drug-induced lipid storage disease. Arch. Ophthalmol. 1981, 99, 257–261. [Google Scholar] [CrossRef]
- Jacobs, J.M.; Costa-Jussà, F.R. The pathology of amiodarone neurotoxicity: II. Peripheral neuropathy in man. Brain 1985, 108, 753–769. [Google Scholar] [CrossRef]
- Rappersberger, K.; Hönigsmann, H.; Ortel, B.; Tanew, A.; Konrad, K.; Wolff, K. Photosensitivity and Hyperpigmentation in Amiodarone-Treated Patients: Incidence, Time Course, and Recovery. J. Investig. Dermatol. 1989, 93, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Pirovino, M.; Müller, O.; Zysset, T.; Honegger, U. Amiodarone-induced hepatic phospholipidosis: Correlation of morphological and biochemical findings in an animal model. Hepatology 1988, 8, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Maekawa, K.; Ishikawa, M.; Senoo, Y.; Urata, M.; Murayama, M.; Nakatsu, N.; Yamada, H.; Saito, Y. Glucosylceramide and Lysophosphatidylcholines as Potential Blood Biomarkers for Drug-Induced Hepatic Phospholipidosis. Toxicol. Sci. 2014, 141, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecommandeur, E.; Baker, D.; Cox, T.M.; Nicholls, A.W.; Griffin, J.L. Alterations in endo-lysosomal function induce similar hepatic lipid profiles in rodent models of drug-induced phospholipidosis and Sandhoff disease. J. Lipid Res. 2017, 58, 1306–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatman, L.A.; Morton, D.; Johnson, T.O.; Anway, S.D. A Strategy for Risk Management of Drug-Induced Phospholipidosis. Toxicol. Pathol. 2009, 37, 997–1005. [Google Scholar] [CrossRef] [Green Version]
- Lecureux, L.; Cheng, C.S.; Herbst, J.; Reilly, T.P.; Lehman-McKeeman, L.; Otieno, M. Evaluation and validation of multiple cell lines and primary mouse macrophages to predict phospholipidosis potential. Toxicol. In Vitro 2011, 25, 1934–1943. [Google Scholar] [CrossRef]
- Bhandari, N.; Figueroa, D.J.; Lawrence, J.W.; Gerhold, D.L. Phospholipidosis Assay in HepG2 Cells and Rat or Rhesus Hepatocytes Using Phospholipid Probe NBD-PE. Assay Drug Dev. Technol. 2008, 6, 407–419. [Google Scholar] [CrossRef]
- Shahane, S.A.; Huang, R.; Gerhold, D.; Baxa, U.; Austin, C.P.; Xia, M. Detection of phospholipidosis induction: A cell-based assay in high-throughput and high-content format. J. Biomol. Screen. 2013, 19, 66–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van De Water, F.; Havinga, J.; Ravesloot, W.; Horbach, G.; Schoonen, W. High content screening analysis of phospholipidosis: Validation of a 96-well assay with CHO-K1 and HepG2 cells for the prediction of in vivo based phospholipidosis. Toxicol. In Vitro 2011, 25, 1870–1882. [Google Scholar] [CrossRef]
- Broutier, L.; Andersson-Rolf, A.; Hindley, C.J.; Boj, S.F.; Clevers, H.; Koo, B.-K.; Huch, M. Culture and establishment of self-renewing human and mouse adult liver and pancreas 3D organoids and their genetic manipulation. Nat. Protoc. 2016, 11, 1724–1743. [Google Scholar] [CrossRef]
- Xu, H.; Jiao, Y.; Qin, S.; Zhao, W.; Chu, Q.; Wu, K. Organoid technology in disease modelling, drug development, personalized treatment and regeneration medicine. Exp. Hematol. 2018, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Meng, Q. Three-dimensional culture of hepatocytes for prediction of drug-induced hepatotoxicity. Expert Opin. Drug Metab. Toxicol. 2010, 6, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Kostadinova, R.; Boess, F.; Applegate, D.; Suter, L.; Weiser, T.; Singer, T.; Naughton, B.; Roth, A. A long-term three dimensional liver co-culture system for improved prediction of clinically relevant drug-induced hepatotoxicity. Toxicol. Appl. Pharmacol. 2013, 268, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Donato, M.T.; Gomez-Lechon, M.J. Drug-induced liver steatosis and phospholipidosis: Cell-based assays for early screening of drug candidates. Curr. Drug Metab. 2012, 13, 1160–1173. [Google Scholar] [CrossRef] [PubMed]
- Nioi, P.; Perry, B.K.; Wang, E.-J.; Gu, Y.-Z.; Snyder, R.D. In Vitro Detection of Drug-Induced Phospholipidosis Using Gene Expression and Fluorescent Phospholipid Based Methodologies. Toxicol. Sci. 2007, 99, 162–173. [Google Scholar] [CrossRef] [Green Version]
- Sawada, H.; Takami, K.; Asahi, S. A Toxicogenomic Approach to Drug-Induced Phospholipidosis: Analysis of Its Induction Mechanism and Establishment of a Novel in Vitro Screening System. Toxicol. Sci. 2004, 83, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Tengstrand, E.A.; Miwa, G.T.; Hsieh, F.Y. Bis (monoacylglycerol) phosphate as a non-invasive biomarker to monitor the onset and time-course of phospholipidosis with drug-induced toxicities. Expert Opin. Drug Metab. Toxicol. 2010, 6, 555–570. [Google Scholar] [CrossRef]
- Nonoyama, T.; Fukuda, R. Drug-induced Phospholipidosis -Pathological Aspects and Its Prediction. J. Toxicol. Pathol. 2008, 21, 9–24. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Shen, C.; Zhang, G.; Hu, W. Rifampicin exacerbates isoniazid-induced toxicity in human but not in rat hepatocytes in tissue-like cultures. Br. J. Pharmacol. 2007, 153, 784–791. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Zhang, G.; Qiu, H.; Shen, C. Acetaminophen-induced hepatotoxicity of gel entrapped rat hepatocytes in hollow fibers. Chem. Interact. 2006, 162, 53–61. [Google Scholar] [CrossRef]
- Kikkawa, R.; Fujikawa, M.; Yamamoto, T.; Hamada, Y.; Yamada, H.; Horii, I. In vivo hepatotoxicity study of rats in comparison with in vitro hepatotoxicity screening system. J. Toxicol. Sci. 2006, 31, 23–34. [Google Scholar] [CrossRef] [Green Version]
- O′brien, P.J.; Chan, K.; Silber, P.M. Human and animal hepatocytes in vitro with extrapolation in vivo. Chem. Interact. 2004, 150, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Neal, J.; Li, X.; Zhu, J.; Giangarra, V.; Grzeskowiak, C.L.; Ju, J.; Liu, I.H.; Chiou, S.-H.; Salahudeen, A.A.; Smith, A.R.; et al. Organoid Modeling of the Tumor Immune Microenvironment. Cell 2018, 175, 1972–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettinato, G.; Lehoux, S.; Ramanathan, R.; Salem, M.M.; He, L.-X.; Muse, O.; Flaumenhaft, R.; Thompson, M.T.; Rouse, E.A.; Cummings, R.D.; et al. Generation of fully functional hepatocyte-like organoids from human induced pluripotent stem cells mixed with Endothelial Cells. Sci. Rep. 2019, 9, 8920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrell, R.E. Chapter 2—RNA Isolation Strategies. In RNA Methodologies, 4th ed.; Farrell, R.E., Ed.; Academic Press: San Diego, CA, USA, 2010; pp. 45–80. [Google Scholar] [CrossRef]
- Ma, J.Y.; Snook, S.; Garrovillo, S.; Johnson, C.; La, D. An Immunohistochemical Investigation of Renal Phospholipidosis and Toxicity in Rats. Int. J. Toxicol. 2017, 36, 386–394. [Google Scholar] [CrossRef]
- Obert, L.; Sobocinski, G.P.; Bobrowski, W.F.; Metz, A.L.; Rolsma, M.D.; Altrogge, D.M.; Dunstan, R.W. An Immunohistochemical Approach to Differentiate Hepatic Lipidosis from Hepatic Phospholipidosis in Rats. Toxicol. Pathol. 2007, 35, 728–734. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.M.; Gombart, Z.J.; Chen, J.W. Chloroquine treatment of ARPE-19 cells leads to lysosome dilation and intracellular lipid accumulation: Possible implications of lysosomal dysfunction in macular degeneration. Cell Biosci. 2011, 1, 10. [Google Scholar] [CrossRef] [Green Version]











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-Y.; Han, H.-J.; Lee, S.-J.; Cho, E.-H.; Lee, H.-B.; Seok, J.-H.; Lim, H.S.; Son, W.-C. Use of 3D Human Liver Organoids to Predict Drug-Induced Phospholipidosis. Int. J. Mol. Sci. 2020, 21, 2982. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082982
Lee J-Y, Han H-J, Lee S-J, Cho E-H, Lee H-B, Seok J-H, Lim HS, Son W-C. Use of 3D Human Liver Organoids to Predict Drug-Induced Phospholipidosis. International Journal of Molecular Sciences. 2020; 21(8):2982. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082982
Chicago/Turabian StyleLee, Ji-Young, Hyo-Jeong Han, Sang-Joon Lee, Eun-Ho Cho, Han-Byul Lee, Ju-Hyung Seok, Hee Seon Lim, and Woo-Chan Son. 2020. "Use of 3D Human Liver Organoids to Predict Drug-Induced Phospholipidosis" International Journal of Molecular Sciences 21, no. 8: 2982. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082982