Genome-Wide Identification of M35 Family Metalloproteases in Rhizoctonia cerealis and Functional Analysis of RcMEP2 as a Virulence Factor during the Fungal Infection to Wheat

Abstract
:1. Introduction
2. Results
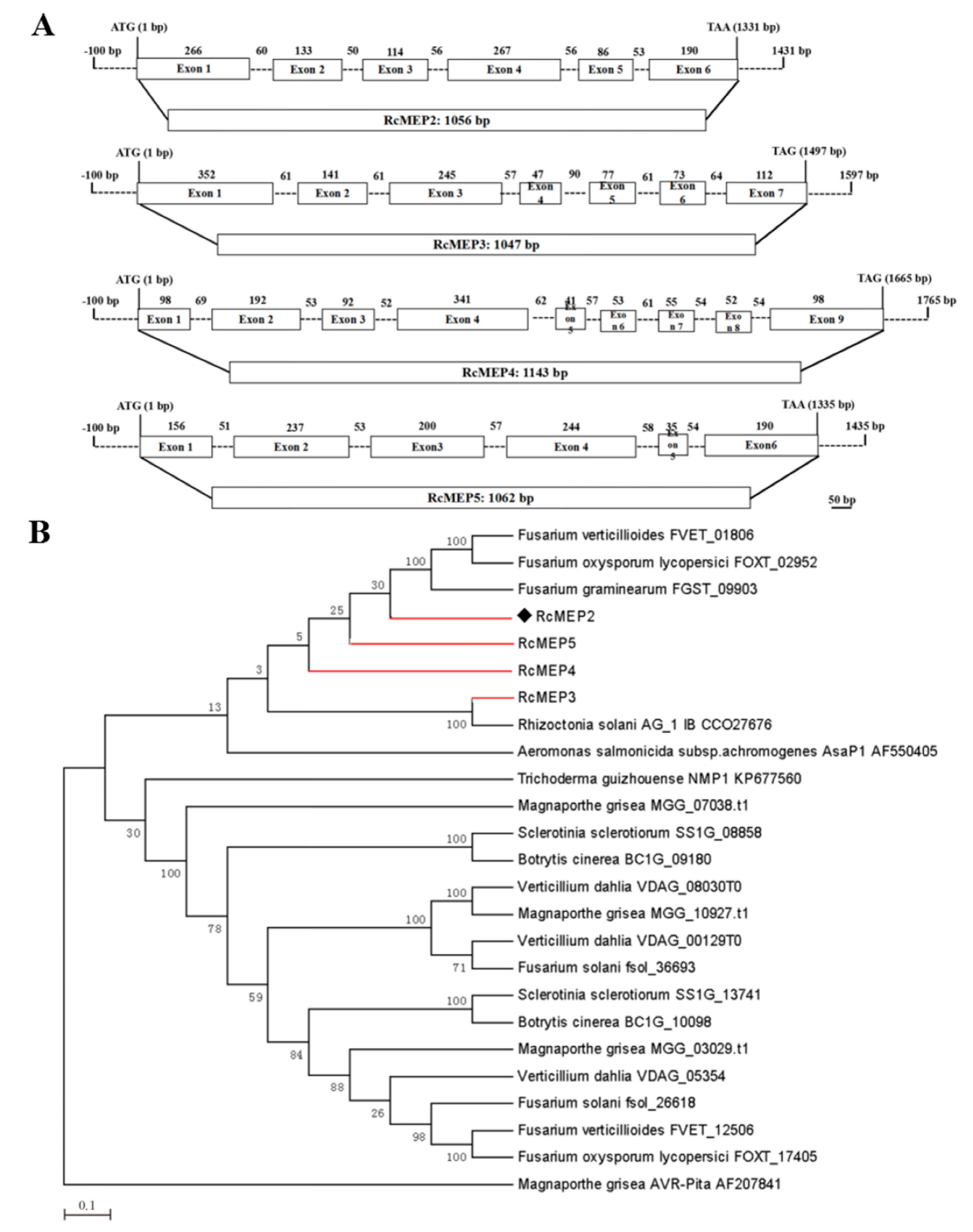
2.1. Global Characterization of the M35 Family Metalloproteases in R. cerealis Genome
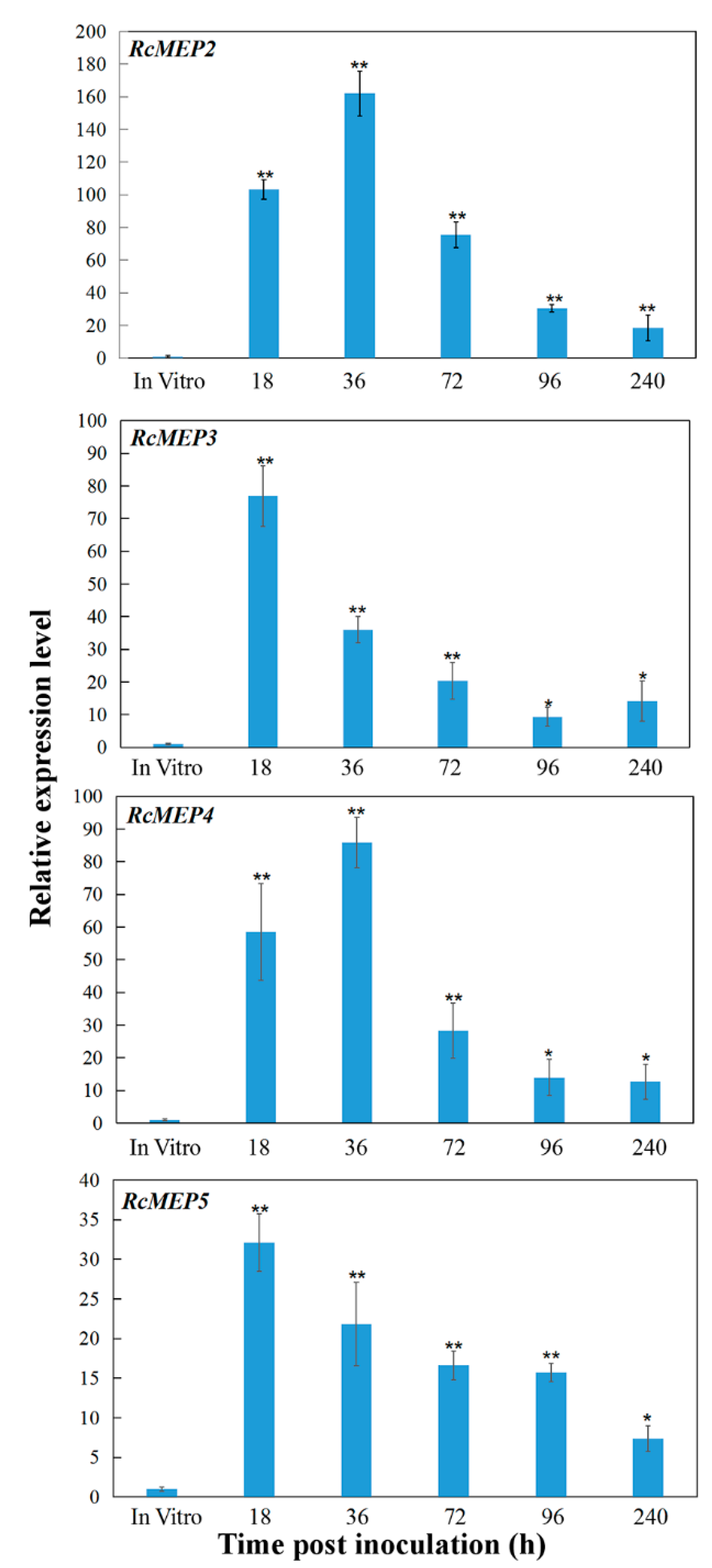
2.2. Transcriptional Profiles of RcMEP2-RcMEP5 during the Fungus Infection to Wheat
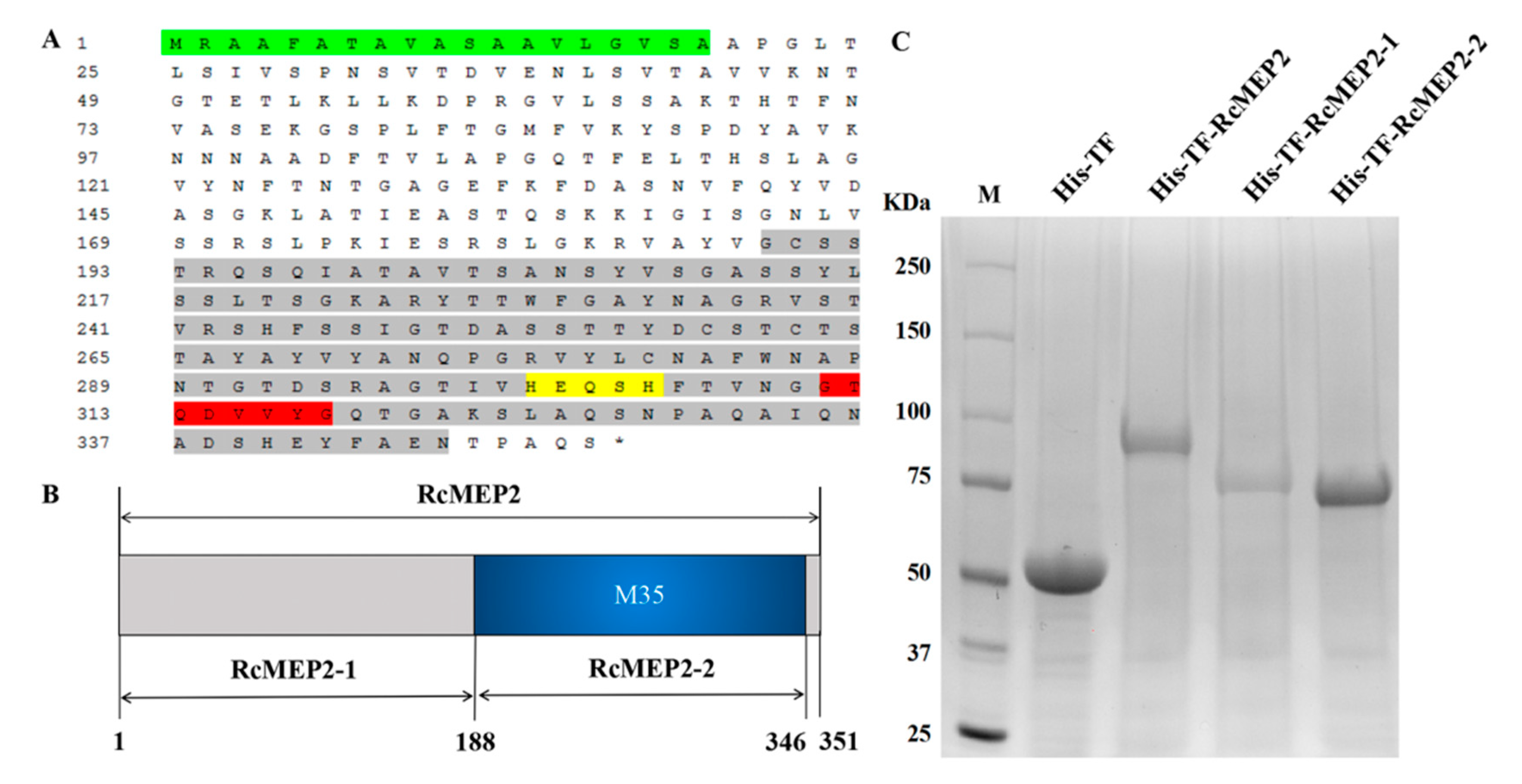
2.3. RcMEP2 and Its M35 Domain-Containing Peptides Induce Cell Death in Infiltrated Plant Leaves
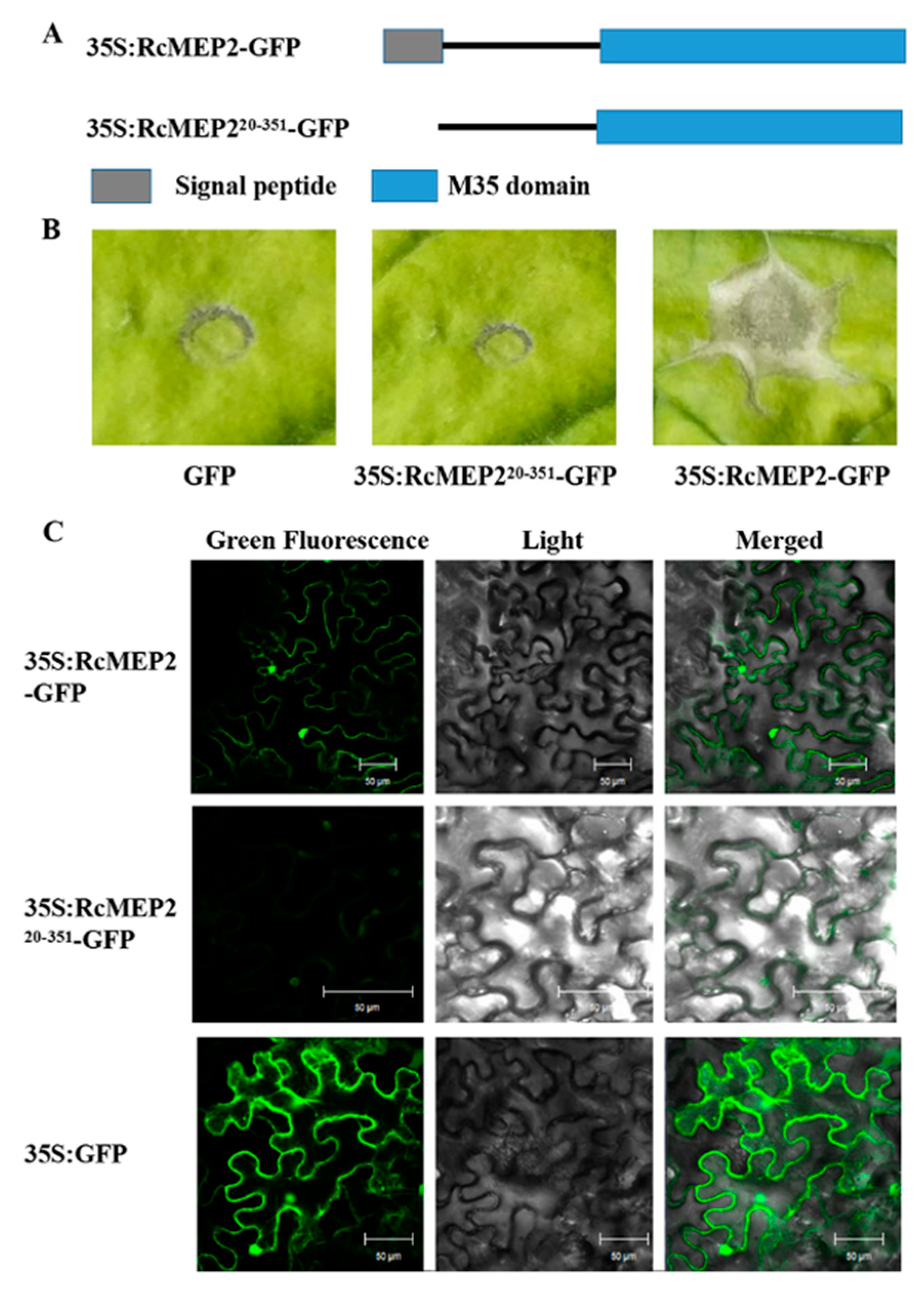
2.4. The Signal Peptide is Required for Secretory and Cell Death-Inducing Abilities of the Transiently-Expressing RcMEP2 in N. benthamiana
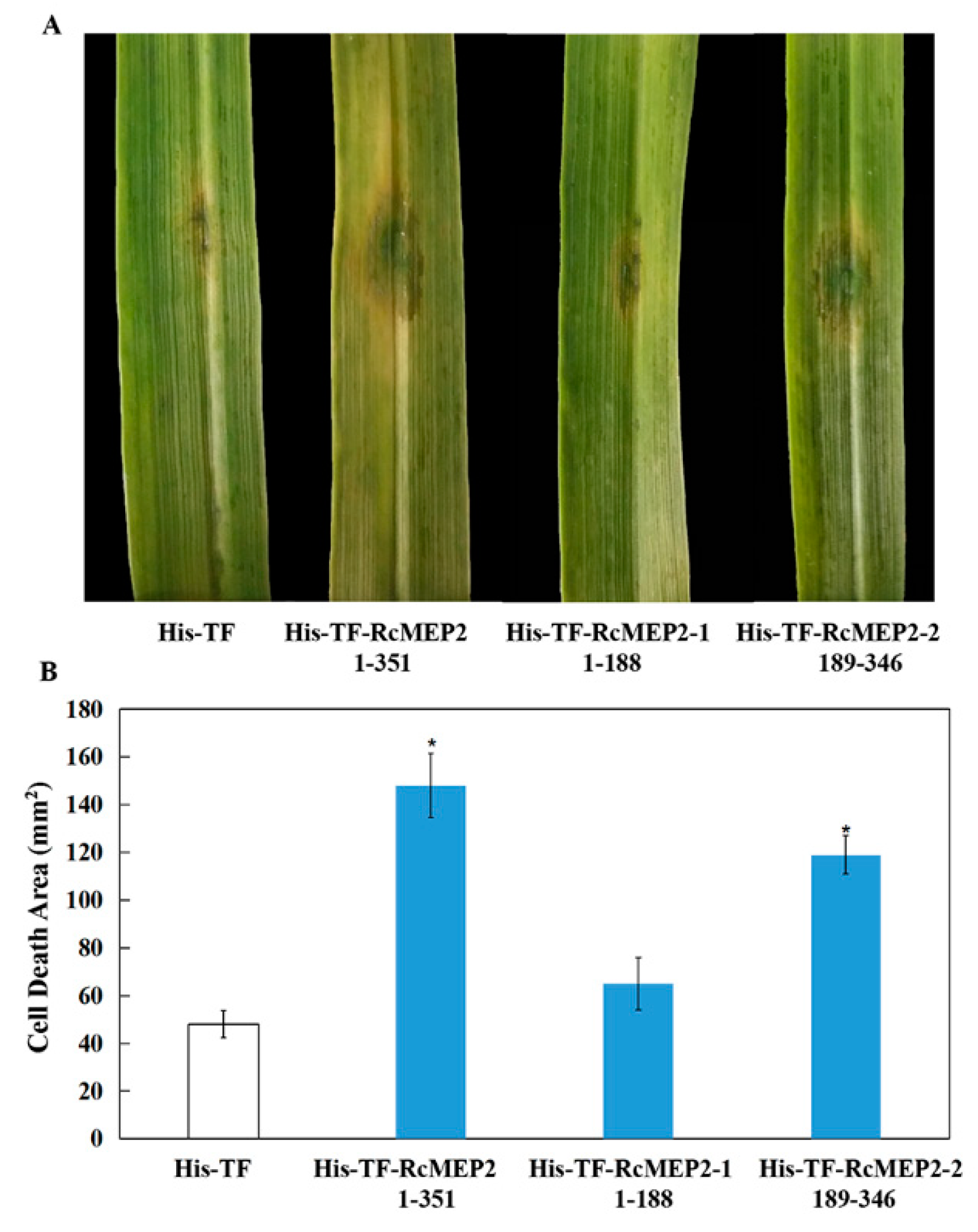
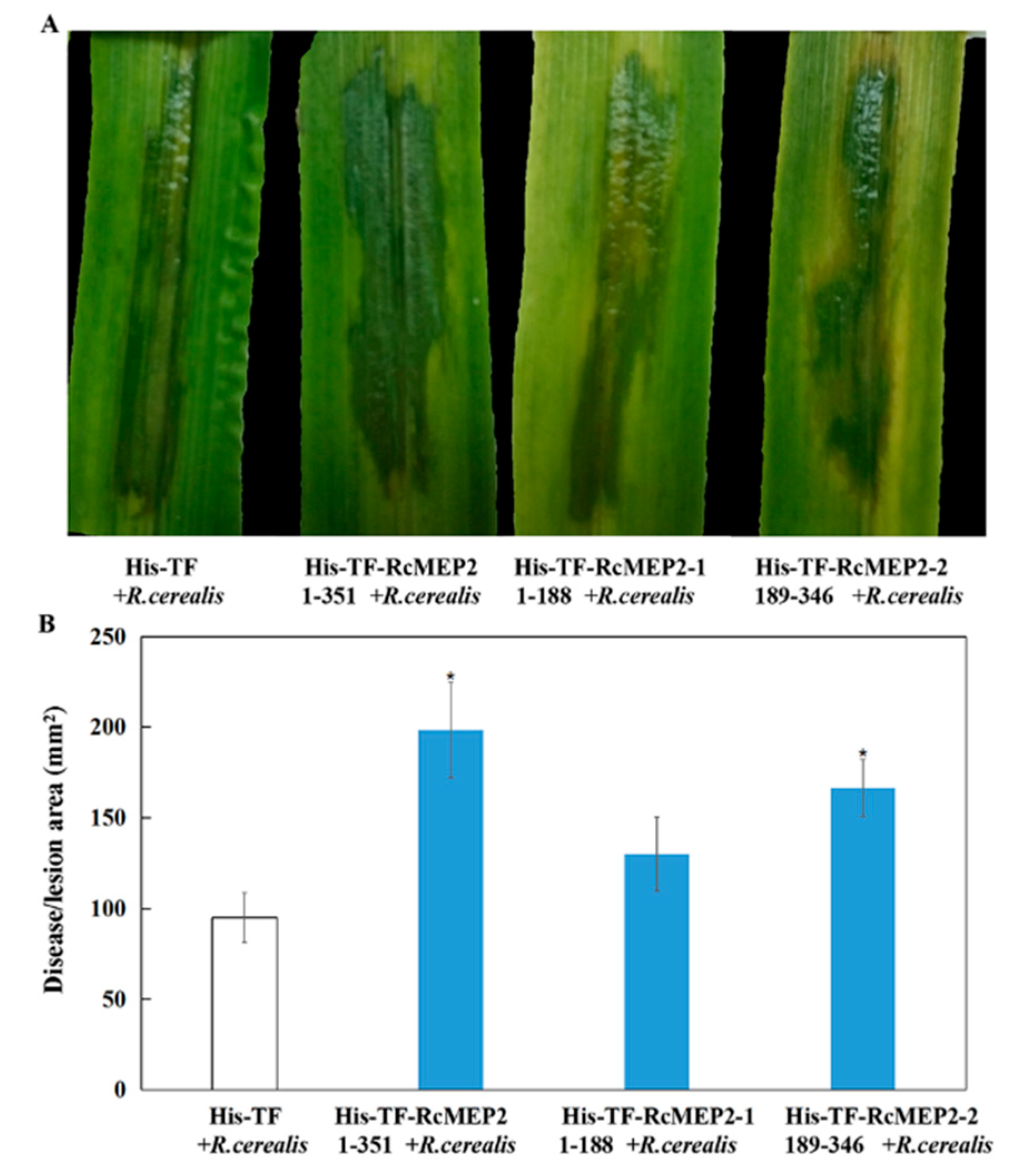
2.5. RcMEP2 and Its M35 Domain Peptides Contribute to Pathogenicity of R. cerealis to Wheat
2.6. RcMEP2 can Promote Accumulation of H2O2 in Infiltrated Plant Leaves
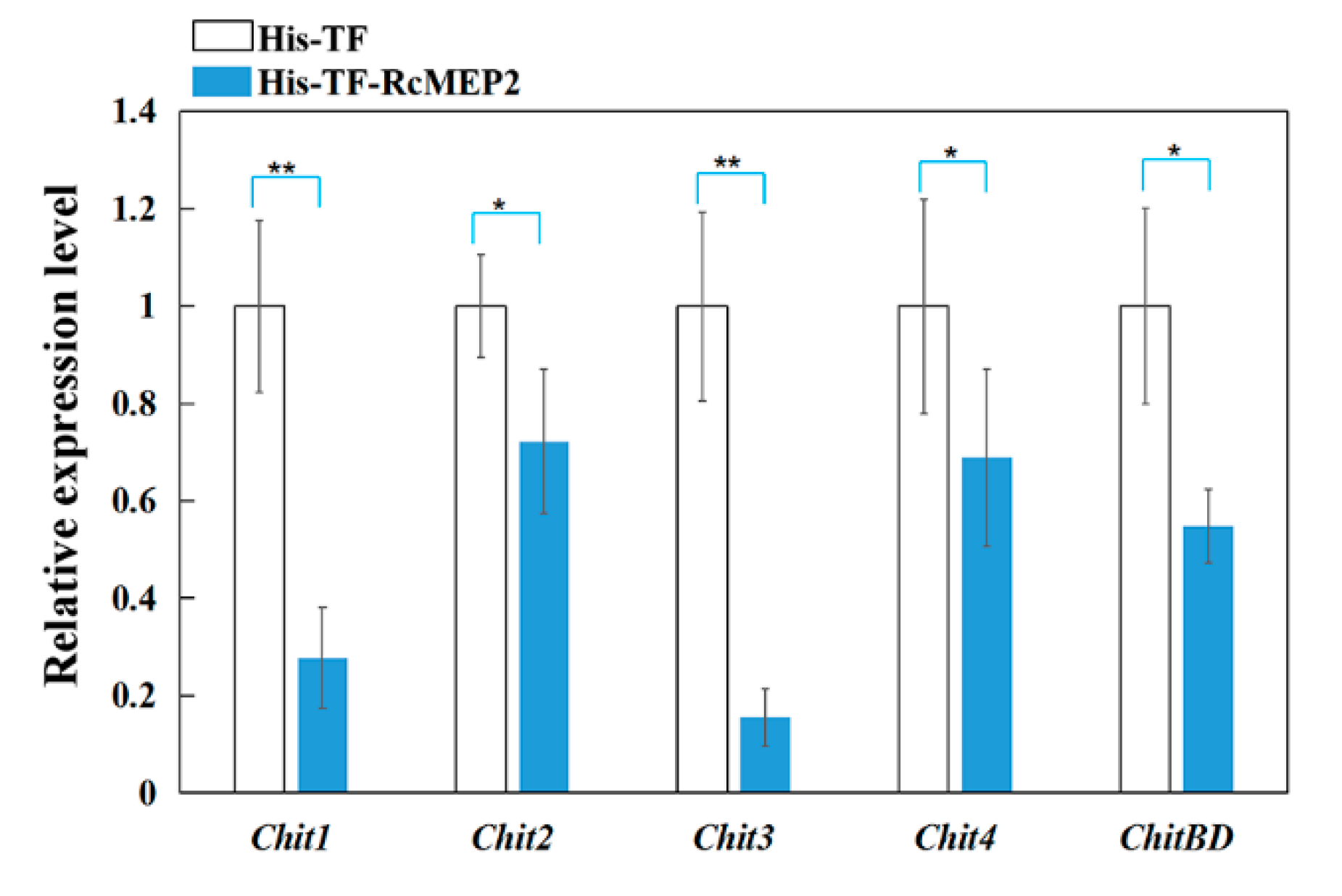
2.7. RcMEP2 Inhibits the Expression of Chitinases in Wheat
3. Discussion
4. Materials and Methods
4.1. Fungal Isolate, Plant Materials, and Treatments
4.2. Identification of M35 Family Metalloproteases in Rhizoctonia Cerealis
4.3. RNA Extraction, cDNA Synthesis, and All Primers
4.4. Plasmid Construction and Preparation of RcMEP2
4.5. Transient Expression of RcMEP2 or Its Signal Peptide-Deleting Proteins in N. benthamiana
4.6. qRT-PCR Analysis
4.7. Heterologous Expression and Purification of RcMEP2 and Its M35 Domain-Containing/Deleting Peptides
4.8. Cell Death-Inducing Activities of RcMEP2 Protein and Its M35-Containing Peptide
4.9. DAB Staining for Detection of H2O2
4.10. Disease Assay of the Purified RcMEP2 and Its M35-Containing Peptide in Wheat
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moore, J.W.; Herrerafoessel, S.; Lan, C.; Schnippenkoetter, W.; Ayliffe, M.; Huerta-Espino, J.; Lillemo, M.; Viccars, L.; Milne, R.; Periyannan, S.; et al. A recently evolved hexose transporter variant confers resistance to multiple pathogens in wheat. Nat. Genet. 2015, 47, 1494–1498. [Google Scholar] [CrossRef]
- Hamada, M.S.; Yin, Y.; Chen, H.; Ma, Z. The escalating threat of Rhizoctonia cerealis, the causal agent of sharp eyespot in wheat. Pest Manag. Sci. 2011, 67, 1411–1419. [Google Scholar] [CrossRef]
- Ji, L.; Liu, C.; Zhang, L.; Liu, A.; Yu, J. Variation of rDNA Internal Transcribed Spacer Sequences in Rhizoctonia cerealis. Curr. Microbiol. 2017, 74, 877–884. [Google Scholar] [CrossRef]
- Li, W.; Sun, H.; Deng, Y.; Zhang, A.; Chen, H. The heterogeneity of the rDNA-ITS sequence and its phylogeny in Rhizoctonia cerealis, the cause of sharp eyespot in wheat. Curr. Genet. 2014, 60, 1–9. [Google Scholar] [CrossRef]
- Van Der Hoeven, E.P.; Bollen, G.J. Effect of benomyl on soil fungi associated with rye. 1. Effect on the incidence of sharp eyespot caused by Rhizoctonia cerealis. Neth. J. Plant Pathol. 1980, 86, 163–180. [Google Scholar] [CrossRef]
- Lemańczyk, G.; Kwaśna, H. Effects of sharp eyespot (Rhizoctonia cerealis) on yield and grain quality of winter wheat. Eur. J. Plant Pathol. 2013, 135, 187–200. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, G.; Du, Z.; Quan, W.; Zhang, H.; Che, M.; Wang, Z.; Zhang, Z. Mapping of QTL conferring resistance to sharp eyespot (Rhizoctonia cerealis) in bread wheat at the adult plant growth stage. Theor. Appl. Genet. 2013, 126, 2865–2878. [Google Scholar] [CrossRef]
- Zhu, X.; Yang, K.; Wei, X.; Zhang, Q.; Rong, W.; Du, L.; Ye, X.; Qi, L.; Zhang, Z. The wheat AGC kinase TaAGC1 is a positive contributor to host resistance to the necrotrophic pathogen Rhizoctonia cerealis. J. Exp. Bot. 2015, 66, 6591–6603. [Google Scholar] [CrossRef] [Green Version]
- Burpee, L.L.; Sanders, P.L.; Cole, H., Jr.; Sherwood, R.T. Anastomosis groups among isolates of anastomosis groups among isolates of Ceratobasidium cornigerum and related fungi. Mycologia 1980, 72, 689–701. [Google Scholar] [CrossRef]
- Tomaso-Peterson, M.; Trevathan, L.E. Characterization of Rhizoctonia-like fungi isolated from agronomic crops and turfgrasses in Mississippi. Plant Dis. 2007, 91, 260–265. [Google Scholar] [CrossRef]
- Lu, L.; Rong, W.; Massart, S.; Zhang, Z.Y. Genome-wide identification and expression analysis of cutinase gene family in Rhizoctonia cerealis and functional study of an active cutinase RcCUT1 in the fungal-wheat interaction. Front. Microbiol. 2018, 9, 1813. [Google Scholar] [CrossRef]
- Lu, L.; Liu, Y.; Zhang, Z. Global characterization of GH10 family xylanase genes in Rhizoctonia cerealis and functional analysis of xylanase RcXYN1 during fungus infection in wheat. Int. J. Mol. Sci. 2020, 21, 1812. [Google Scholar] [CrossRef] [Green Version]
- Stergiopoulos, I.; de Wit, P.J.G.M. Fungal effector proteins. Annu. Rev. Phytopathol. 2009, 47, 233–263. [Google Scholar] [CrossRef] [Green Version]
- Balbi-Pena, M.I.; Schwan-Estrada, K.R.F.; Stangarlin, J.R. Differential occurrence of the oxidative burst and the activity of defence related enzymes in compatible and incompatible tomato–Oidium neolycopersici interactions. Australas. Plant Pathol. 2012, 41, 573–586. [Google Scholar] [CrossRef]
- Horbach, R.; Navarro-Quesada, A.R.; Knogge, W.; Deising, H.B. When and how to kill a plant cell: Infection strategies of plant pathogenic fungi. J. Plant Physiol. 2011, 168, 51–62. [Google Scholar] [CrossRef]
- Temme, N.; Tudzynski, P. Does Botrytis cinerea ignore H2O2-induced oxidative stress during infection? Characterization of Botrytis Activator Protein 1. Mol. Plant Microbe Interact. 2009, 22, 987–998. [Google Scholar] [CrossRef] [Green Version]
- Tallant, C.; García-Castellanos, R.; Seco, J.; Baumann, U.; Gomis-Rüth, F.X. Molecular analysis of ulilysin, the structural prototype of a new family of metzincin metalloproteases. J. Biol. Chem. 2006, 281, 17920–17928. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Zhou, X.; Fan, A.; Wang, Z.; Huang, B. Differential functions of two metalloproteases, Mrmep1 and Mrmep2, in growth, sporulation, cell wall integrity, and virulence in the filamentous fungus Metarhizium robertsii. Front. Microbiol. 2018, 9, 1528. [Google Scholar] [CrossRef]
- Laursen, L.S.; Overgaard, M.T.; Nielsen, C.G.; Boldt, H.B.; Hopmann, K.H.; Conover, C.A.; Oxvig, C. Substrate specificity of the metalloproteinase pregnancy- associated plasma protein-A (PAPP-A) assessed by mutagenesis and analysis of synthetic peptides: Substrate residues distant from the scissile bond are critical for proteolysis. Biochem. J. 2002, 367, 31–40. [Google Scholar] [CrossRef]
- Glerup, S.; Boldt, H.B.; Overgaard, M.T.; Sottrup-Jensen, L.; Giudice, L.C.; Oxvig, C. Proteinase inhibition by proform of eosinophil major basic protein (pro-MBP) is a multistep process of intra-and intermolecular disulfide rearrangements. J. Biol. Chem. 2005, 280, 9823–9832. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, N.D.; Morton, F.R.; Barrett, A.J. MEROPS: The peptidase database. Nucleic. Acids. Res. 2006, 34, D270–D272. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhang, K. Independent expansion of zincin metalloproteinases in onygenales fungi may be associated with their pathogenicity. PLoS ONE 2014, 2, e90225. [Google Scholar] [CrossRef] [Green Version]
- Hori, T.; Kumasaka, T.; Yamamoto, M.; Nonaka, T.; Tanaka, N. Structure of a new ‘aspzincin’ metalloendopeptidase from Grifola frondosa implications for the catalytic mechanism and substrate specificity based on several different crystal forms. Acta Crystallogr. D 2001, 57, 361–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnadottir, H.; Hvanndal, I.; Andresdottir, V.; Burr, S.; Frey, J.; Gudmundsdottir, B. The AsaP1 peptidase of Aeromonas salmonicida subsp. achromogenes is a highly conserved deuterolysin metalloprotease (family M35) and a major virulence factor. J. Bacteriol. 2009, 191, 403–410. [Google Scholar]
- Dow, M.; Newman, M.A.; Von Roepenack, E. The induction and modulation of plant defense responses by bacterial lipopolysaccharides. Annu. Rev. Phytopathol. 2000, 38, 241–261. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; McAdams, S.A.; Bryan, G.T.; Hershey, H.P.; Valent, B. Direct interaction of resistance gene and avirulence gene products confers rice blast resistance. EMBO J. 2000, 19, 4004–4014. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Martín, J.M.; Pacheco-Arjona, J.R.; Bello-Rico, V.; Vargas, W.A.; Monod, M.; Díaz-Mínguez, J.M.; Sukno, S.A. A highly conserved metalloprotease effector enhances virulence in the maize anthracnose fungus Colletotrichum graminicola. Mol. Plant Pathol. 2016, 17, 1048–1062. [Google Scholar] [CrossRef] [Green Version]
- Vargas, W.A.; Martín, J.M.S.; Rech, G.E.; Rivera, L.P.; Benito, E.P.; Díaz-Mínguez, J.M.; Sukno, S.A. Plant defense mechanisms are activated during biotrophic and necrotrophic development of Colletotricum graminicola in maize. Plant Physiol. 2012, 158, 1342–1358. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Bak, D.D.; Heid, H.; Geider, K. Molecular characterization of a protease secreted by Erwinia amylovora. J. Mol. Biol. 1999, 289, 1239–1251. [Google Scholar] [CrossRef]
- Jashni, M.K.; Dols, I.H.; Iida, Y.; Boeren, S.; Beenen, H.G.; Mehrabi, R.; de Wit, P.J. Synergistic action of a metalloprotease and a serine protease from Fusarium oxysporum f. sp. lycopersici cleaves chitin-binding tomato chitinases, reduces their antifungal activity, and enhances fungal virulence. Mol. Plant Microbe Interact. 2015, 28, 996–1008. [Google Scholar]
- Saitou, N.; Nei, N. The neighbour joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Naumann, T.A.; Wicklow, D.T.; Price, N.P. Identification of a chitinase-modifying protein from Fusarium verticillioides truncation of a host resistance protein by a fungalysin metalloprotease. J. Biol. Chem. 2011, 286, 35358–35366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naumann, T.A.; Wicklow, D.T. Chitinase modifying proteins from phylogenetically distinct lineages of Brassica pathogens. Physiol. Mol. Plant P 2013, 82, 1–9. [Google Scholar] [CrossRef]
- Han, L.B.; Li, Y.B.; Wang, F.X.; Wang, W.Y.; Liu, J.; Wu, J.H.; Xia, G.X. The Cotton apoplastic protein CRR1 stabilizes chitinase 28 to facilitate defense against the fungal pathogen verticillium dahliae. Plant Cell 2019, 31, 520–536. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Bayram, A.G.; Atanasova, L.; Rahimi, M.J.; Przylucka, A.; Yang, D.; Druzhinina, I.S. Neutral metallopeptidase NMP1 of Trichoderma guizhouense is required for mycotrophy and self–defence. Environ. Microbiol. 2016, 18, 580–597. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Jiang, N.; Liu, J.L.; Liu, W.D.; Wang, G.L. The role of effectors and host immunity in plant-necrotrophic fungal interactions. Virulence 2014, 5, 722–732. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.B.; Ren, L.J.; Yan, W.; Wu, J.Z.; Chen, H.G.; Wu, X.Y.; Zhang, X.Y. Germplasm development and mapping of resistance to sharp eyespot (Rhizoctonia cerealis) in wheat. Sci. Agric. Sin. 2006, 39, 928–934. [Google Scholar]
- Ma, Z.C.; Song, T.; Zhu, L.; Ye, W.; Wang, Y.; Shao, Y.; Tyler, B.M. A Phytophthora sojae glycoside hydrolase 12 protein is a major virulence factor during soybean infection and is recognized as a PAMP. Plant Cell 2015, 27, 2057–2072. [Google Scholar] [CrossRef] [Green Version]
- James, K.H.; Jonathan, P.A.; Angela, H.W.; Jana, S.; Karam, B.S. Genome sequencing and comparative genomics of the broad host-range pathogen Rhizoctonia solani AG8. PLoS Genet. 2014, 10, e1004281. [Google Scholar]
- Petersen, T.N.; Søren Brunak Heijne, G.V.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Kumar, S.; Dudley, J.; Nei, M.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, P.; Parry, D.W. Development and use of a PCR assay to detect Rhizoctonia cerealis, the cause of sharp eyespot in wheat. Plant Pathol. 1996, 45, 872–883. [Google Scholar] [CrossRef]
- Zhang, Z.; Yao, W.; Dong, N.; Liang, H.; Liu, H.; Huang, R. A novel ERF transcription activator in wheat and its induction kinetics after pathogen and hormone treatments. J. Exp. Bot. 2007, 58, 2993–3003. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, J.; Yuan, J.; Wang, X.L.; Zhao, Q.P.; Kong, P.T.; Zhang, X. NITRIC OXIDE-ASSOCIATED PROTEIN1 (At NOA 1) is essential for salicylic acid-induced root waving in Arabidopsis thaliana. New Phytol. 2015, 207, 211–224. [Google Scholar] [CrossRef]
- Dong, N.; Liu, X.; Liu, Y.; Du, L.; Xu, H.; Liu, H.; Xin, Z.; Zhang, Z. Overexpression of TaPIEP1, a pathogen-induced ERF gene of wheat, confers host-enhanced resistance to fungal pathogen Bipolarissoro kiniana. Funct. Integr. Genom. 2010, 10, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zheng, A.; Lin, R.; Zhang, D.; Qin, P.; Xu, L.; Ai, P.; Ding, L.; Wang, Y.; Chen, Y.; Liu, Y.; et al. The evolution and pathogenic mechanisms of the rice sheath blight pathogen. Nat. Commun. 2013, 4, 1424. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Han, C.; Chen, J.; Li, H.; He, K.; Liu, A.; Li, D. Fungal cellulase is an elicitor but its enzymatic activity is not required for its elicitor activity. Mol. Plant Pathol. 2015, 16, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Lee, B.H.; Lee, H.; Xiong, L.; Zhu, J. A mitochondrial complex I defect impairs cold-regulated nuclear gene expression. Plant Cell 2002, 14, 1235–1251. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5’-3’) |
|---|---|
| RcMEP2-F | 5’-TCGGCTTGGTTACCCTCTTCTAC-3’ |
| RcMEP2-R | 5’-CATCCGGGTAGTAGTTACAAGTAAT-3’ |
| His-RcMEP2-F | 5’-GGTACCCTCGAGGGATCCATGTTGTTCTCTGCTCTT-3’ |
| His-RcMEP2-R | 5’-AAGCTTGAATTCGGATCCAGATTGAGCGGGGGTGTT-3’ |
| His-RcMEP2-1-F | 5’-GGTACCCTCGAGGGATCCATGCGCGCTGCTTTCGCTAC-3’ |
| His-RcMEP2-1-R | 5’-AAGCTTGAATTCGGATCCAACATAGGCGACACGTTTGC-3’ |
| His-RcMEP2-2-F | 5’-GGTACCCTCGAGGGATCCGGATGCAGCTCAACTCGCCA-3’ |
| His-RcMEP2-2-R | 5’-AAGCTTGAATTCGGATCCGTTCTCCGCAAAGTATTCAT-3’ |
| TaActin-F | 5’-CACTGGAATGGTCAAGGCTG-3’ |
| TaActin-R | 5’-CTCCATGTCATCCCAGTTG-3’ |
| RcActin-F | 5’- GCATCCACGAGACCACTTAC-3’ |
| RcActin-R | 5’- GCGTCCCGCTGCTCAAGAT-3’ |
| RcMEP2-QF | 5’-CAGCGGAAACCTCGTCTCAT-3’ |
| RcMEP2-QR | 5’-GGTCCCGATCGACGAAAAGT-3’ |
| RcMEP3-QF | 5’-TACGATTGTTACGCGACCCC-3’ |
| RcMEP3-QR | 5’-ATGCGCCAATACCAGTCGAA-3’ |
| RcMEP4-QF | 5’-GGTTCGGAGCTTGGGATCAA-3’ |
| RcMEP4-QR | 5’-GGGCAAGATCTCGACAACCA-3’ |
| RcMEP5-QF | 5’-TGAGCACCGTCAAGTCTCAC-3’ |
| RcMEP5-QR | 5’-GGAGCACTCCAGAAGACACC-3’ |
| Chit1-QF | 5’-GGTAGCACCGACGTCAAGAA-3’ |
| Chit1-QR | 5’-CGGCCCGTAGTTGTAGTTGT-3’ |
| Chit2-QF | 5’-AAGAACTACTGCGACCCGAC-3’ |
| Chit2-QR | 5’-ACGCTGTTCATCCAGTACCA-3’ |
| Chit3-QF | 5’-GACGTGGACTACGAGCACTT-3’ |
| Chit3-QR | 5’-GTACTGGACGACGGTGTTGG-3’ |
| Chit4-QF | 5’-GCAAGTACGGTTATTGCGGG-3’ |
| Chit4-QR | 5’-GGAAAAACTGGCGCGTGTAA-3’ |
| ChitBD-QF | 5’-CGACGGCAAGAGGGAGATAG-3’ |
| ChitBD-QR | 5’-CGCCCCGTAGTTGTAGTTCC-3’ |
| GFP-RcMEP2-F | 5’-ACAGCCCAGATCACTAGTATGCGCGCTGCTTTCGCT-3’ |
| GFP-RcMEP2-R | 5’-CTTGCTCACCATGGATCCAGATTGAGCGGGGGTGTT-3’ |
| GFP-RcMEP220-351-F | 5’-ACCGGTCCCGGGGGATCCGCTCCTGGCCTCACTCTG-3’ |
| GFP-RcMEP220-351-R | 5’-CTTGCTCACCATGGATCCAGATTGAGCGGGGGTGTT-3’ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, L.; Wen, S.; Yu, J.; Lu, L.; Zhu, X.; Zhang, Z. Genome-Wide Identification of M35 Family Metalloproteases in Rhizoctonia cerealis and Functional Analysis of RcMEP2 as a Virulence Factor during the Fungal Infection to Wheat. Int. J. Mol. Sci. 2020, 21, 2984. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082984
Pan L, Wen S, Yu J, Lu L, Zhu X, Zhang Z. Genome-Wide Identification of M35 Family Metalloproteases in Rhizoctonia cerealis and Functional Analysis of RcMEP2 as a Virulence Factor during the Fungal Infection to Wheat. International Journal of Molecular Sciences. 2020; 21(8):2984. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082984
Chicago/Turabian StylePan, Lijun, Shengxian Wen, Jinfeng Yu, Lin Lu, Xiuliang Zhu, and Zengyan Zhang. 2020. "Genome-Wide Identification of M35 Family Metalloproteases in Rhizoctonia cerealis and Functional Analysis of RcMEP2 as a Virulence Factor during the Fungal Infection to Wheat" International Journal of Molecular Sciences 21, no. 8: 2984. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082984