Transcriptomic Profiling of Circular RNA in Different Brain Regions of Parkinson’s Disease in a Mouse Model

 , and
, and
Abstract
:1. Introduction
2. Results
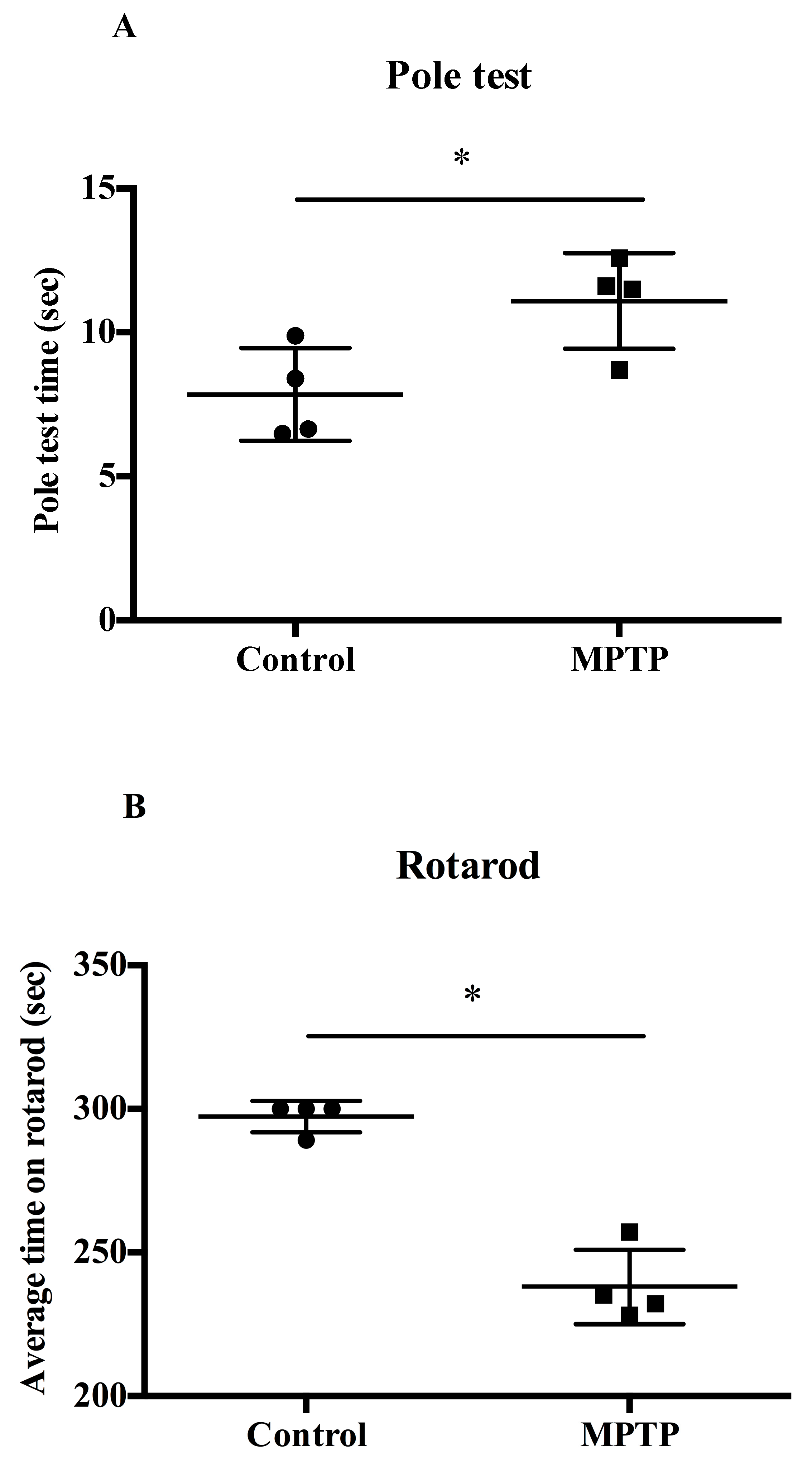
2.1. MPTP-Induced PD Motor Deficits
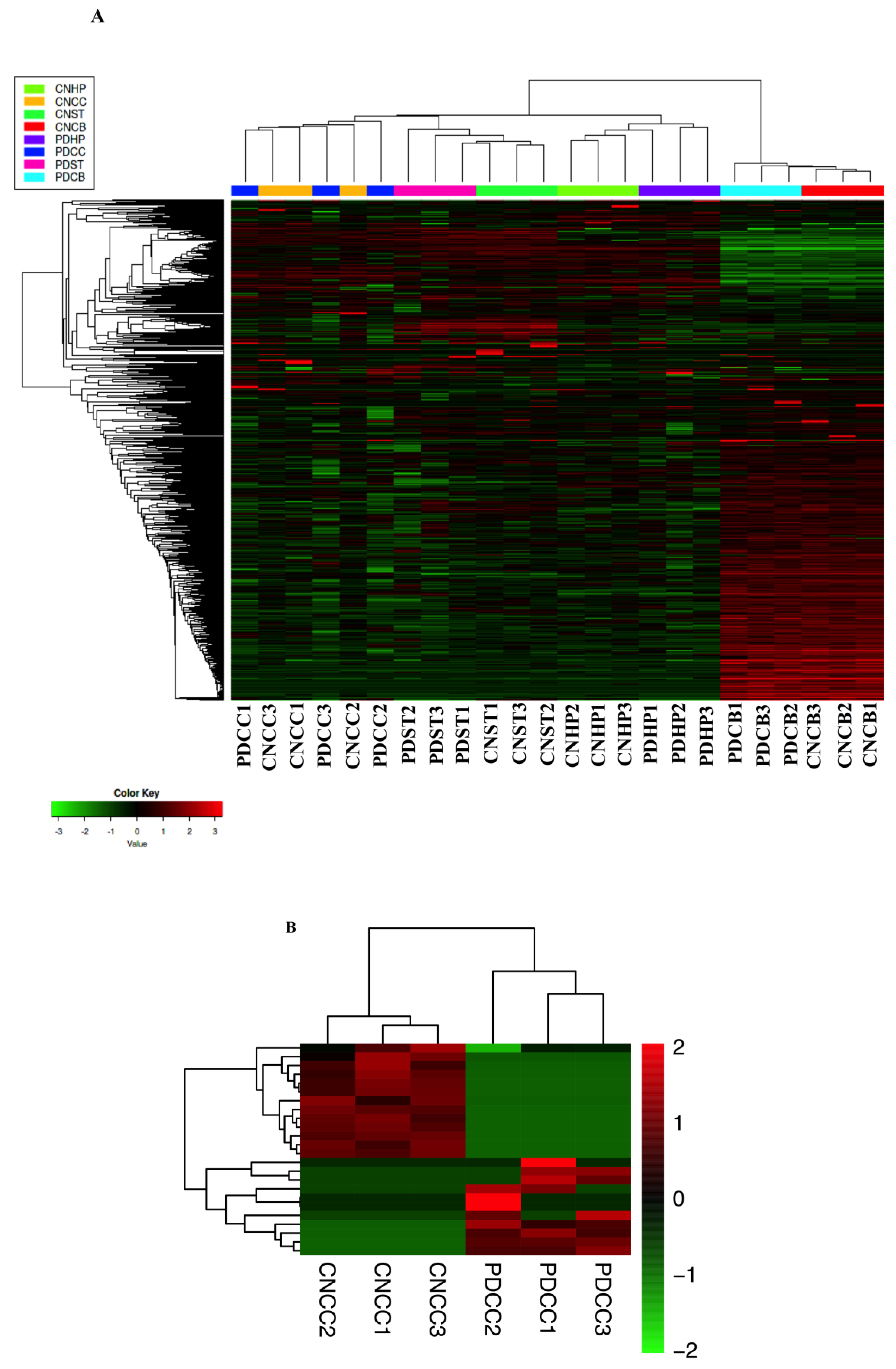
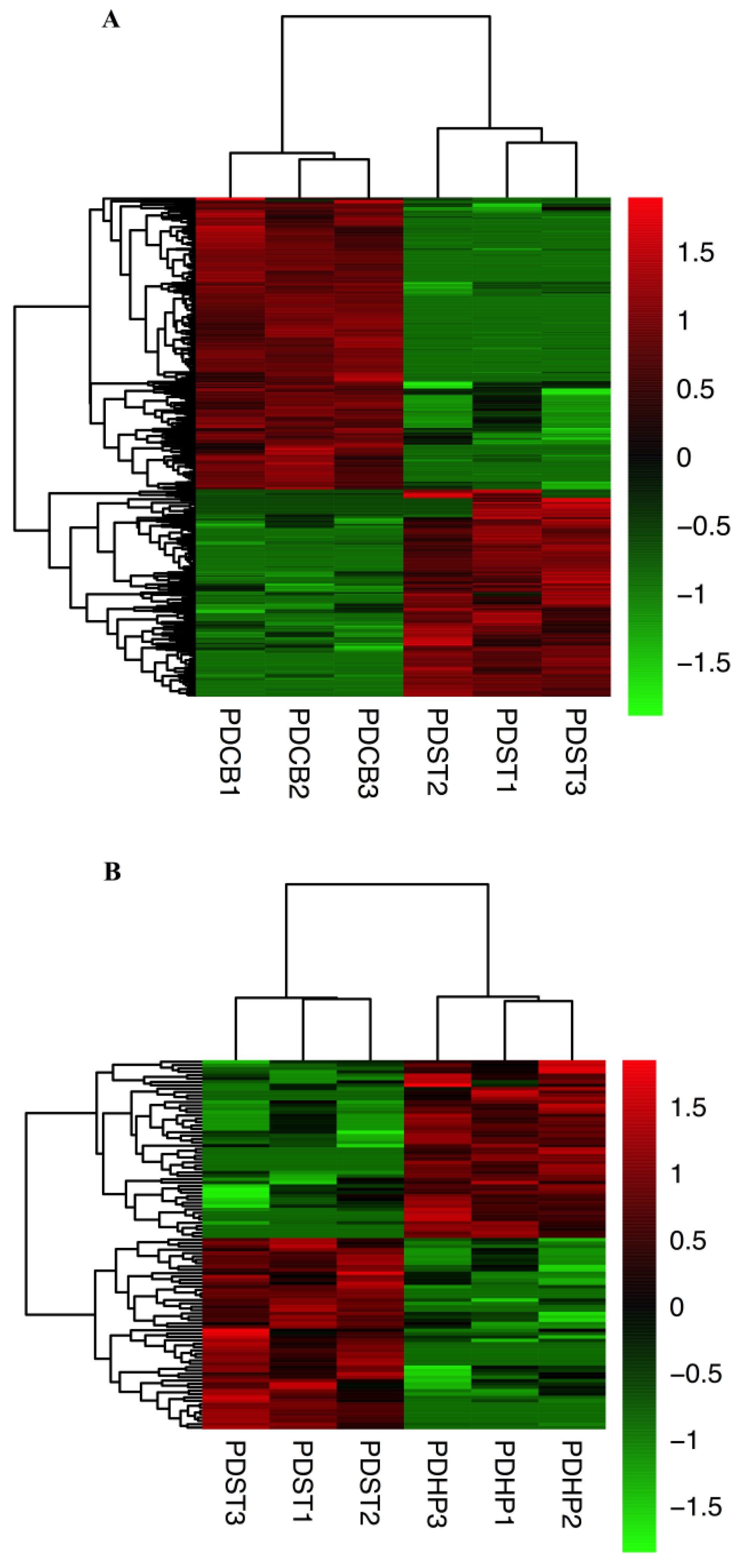
2.2. CircRNAs Expression Profiling in Different Brain Regions
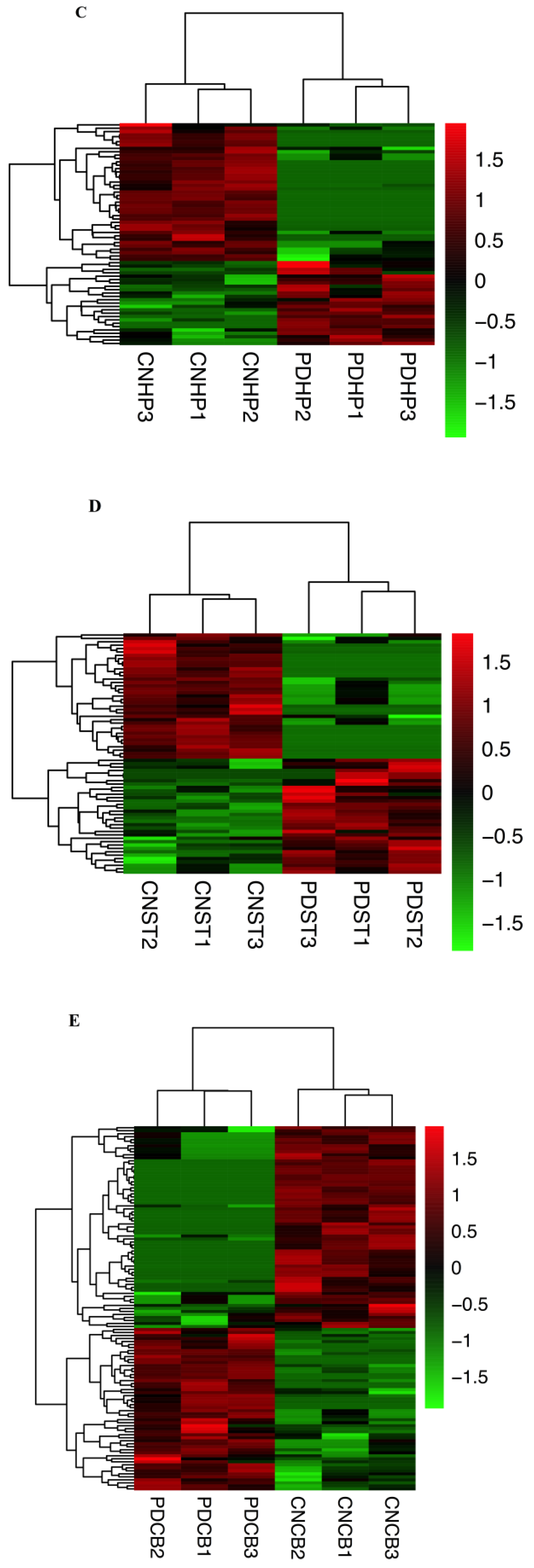
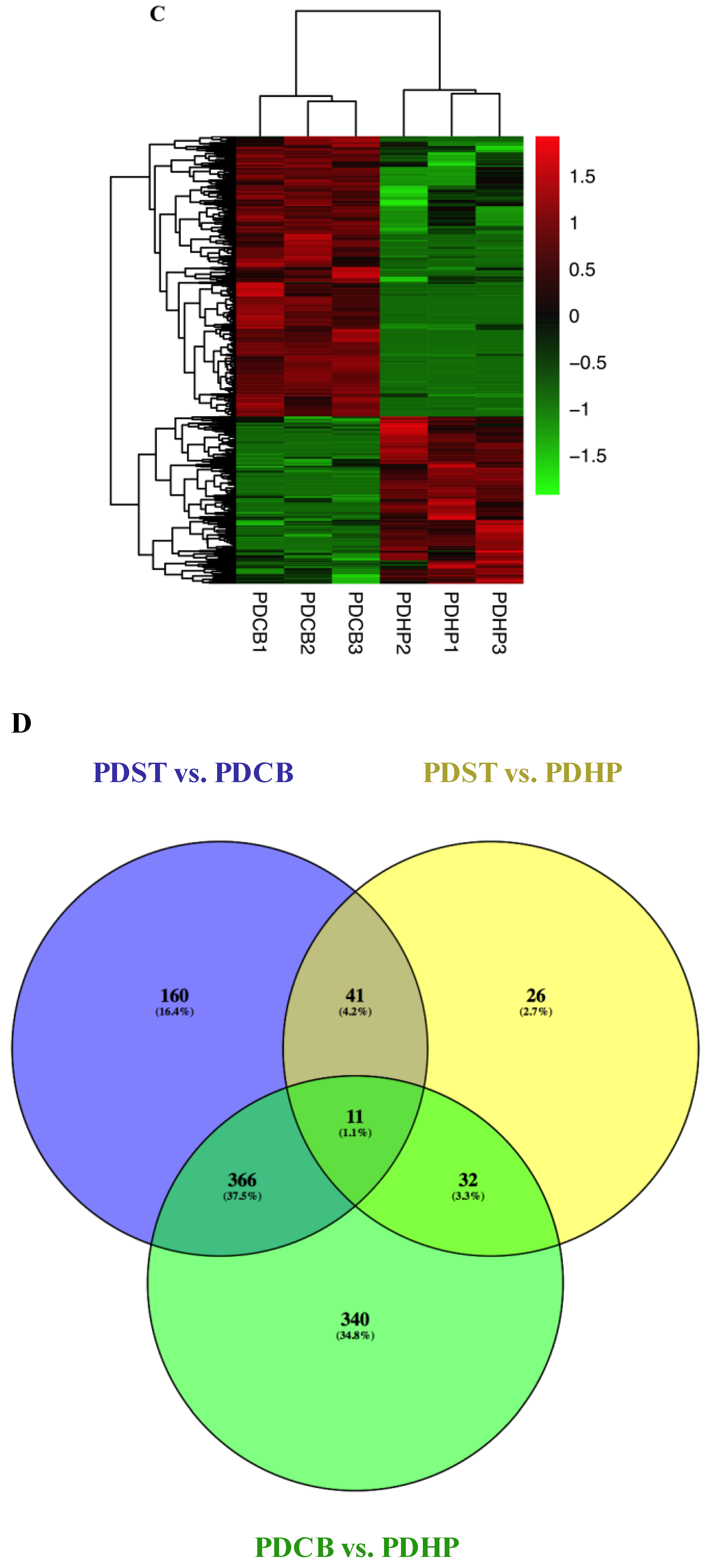
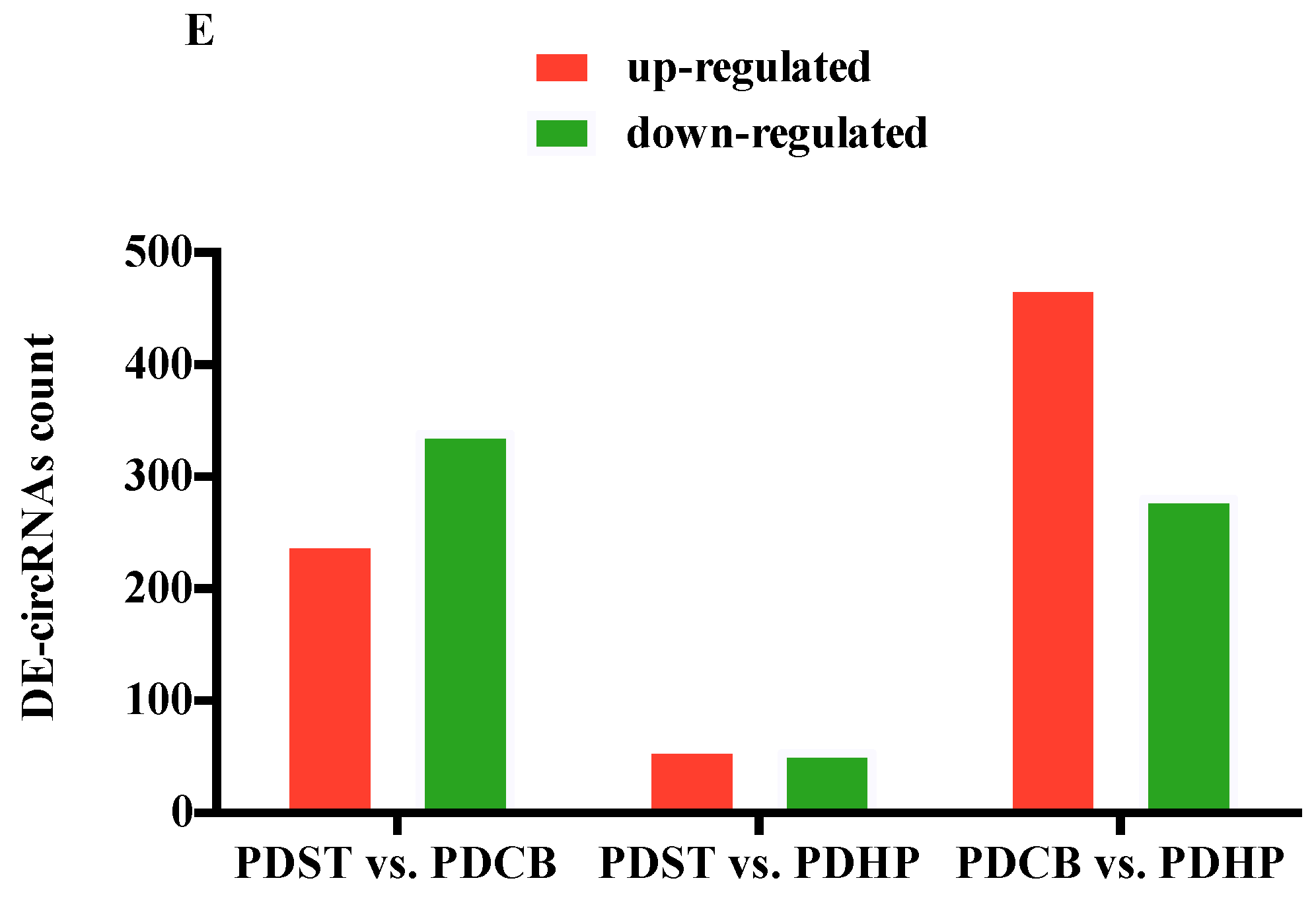
2.3. Comparison of Expression Profiles in Different Brain Regions of PD Mice Model
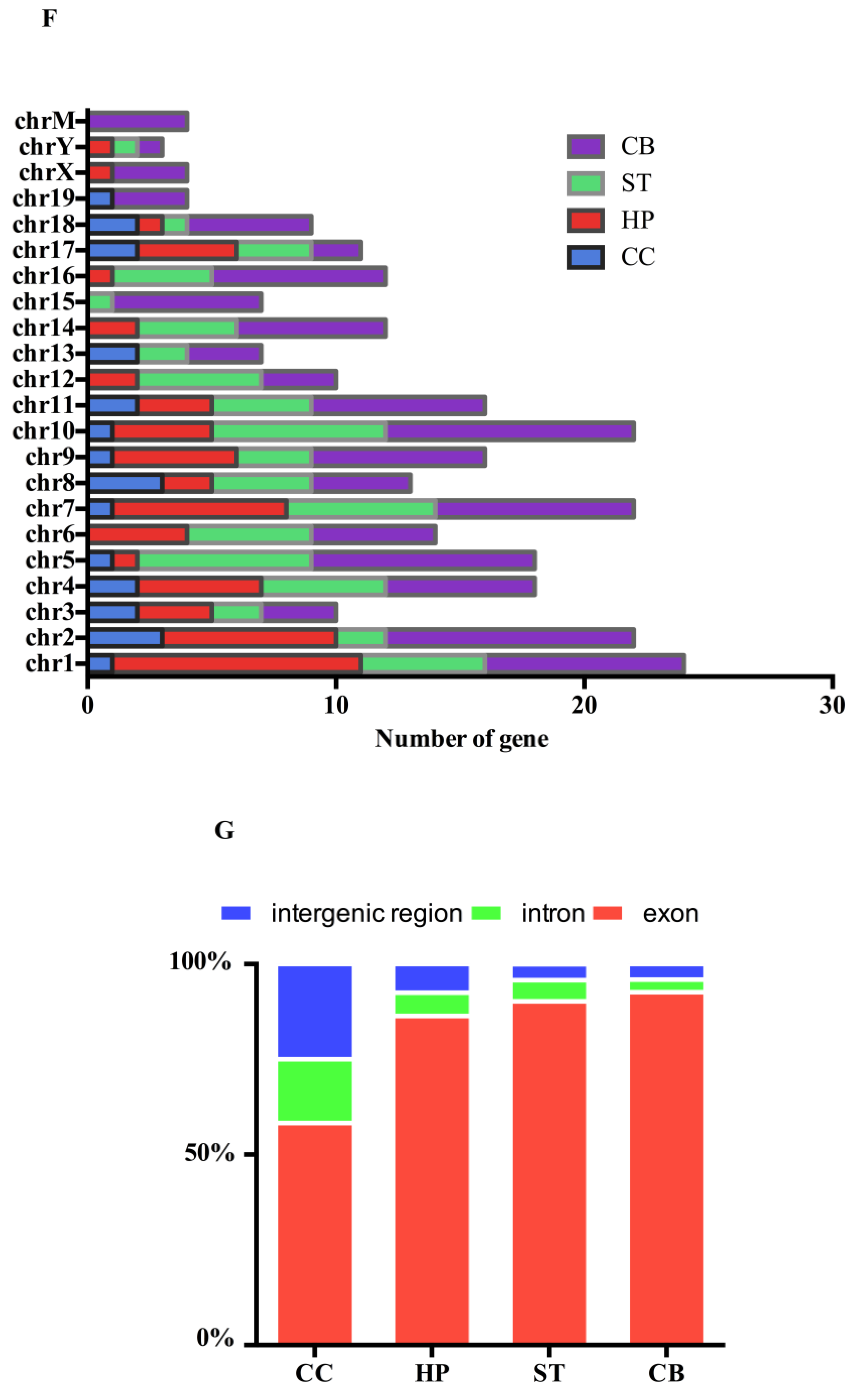
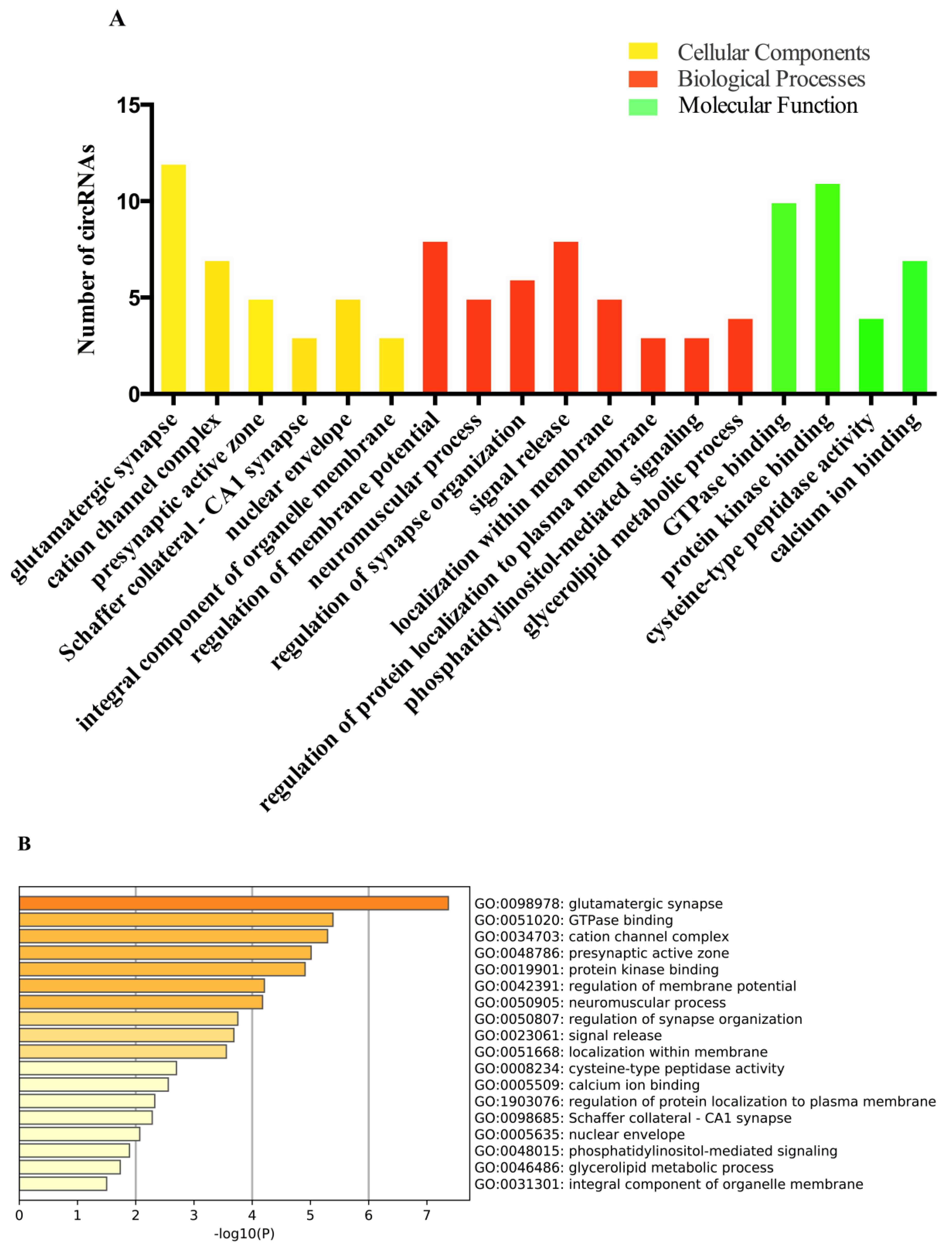
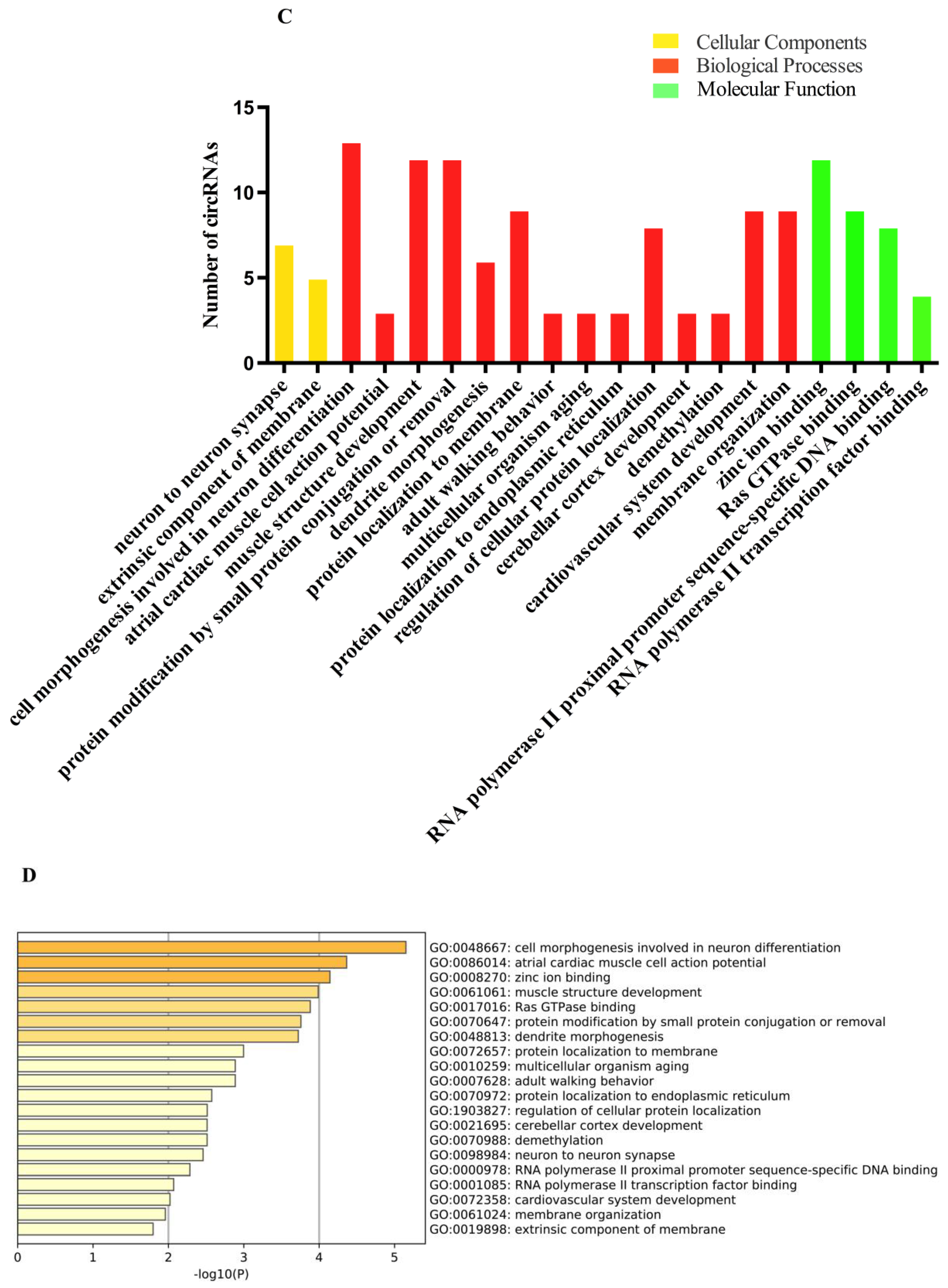
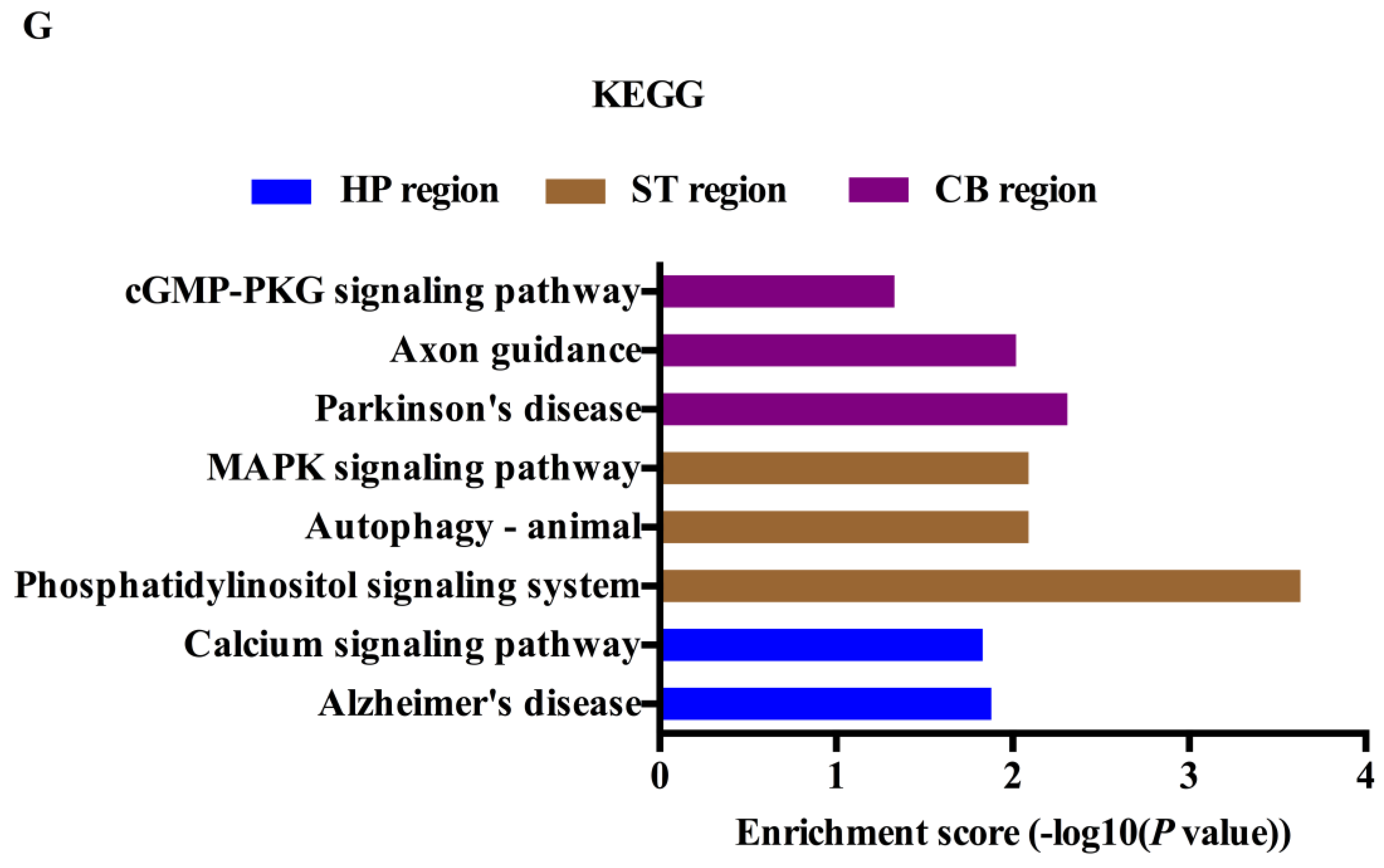
2.4. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Analyses of the circRNAs Parental Genes in Different Brain Regions
2.5. Validation by Quantitative Reverse Transcription PCR (qRT-PCR)
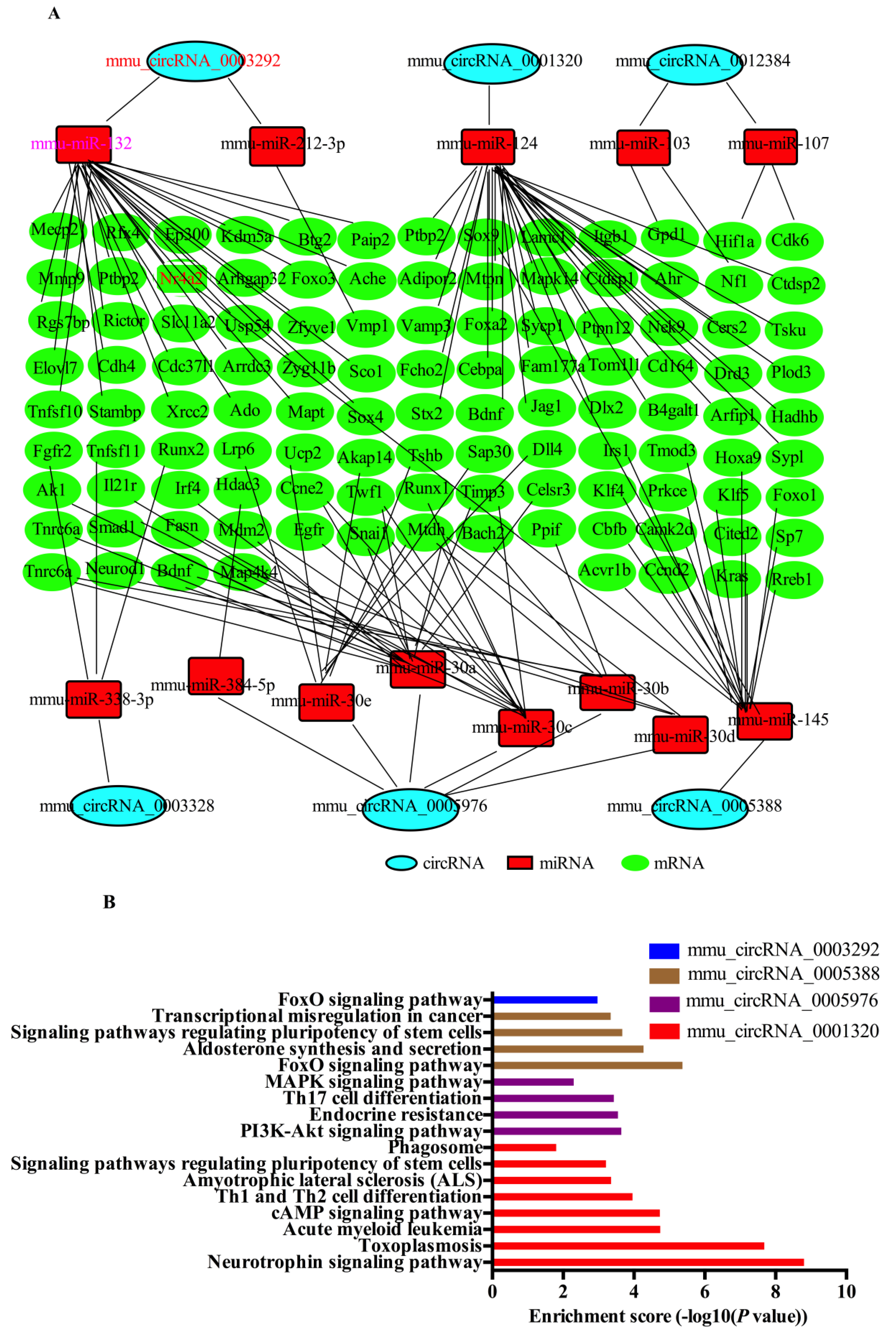
2.6. CircRNA-Targeted miRNA-mRNA Network Prediction and Annotation
3. Discussion
4. Materials and Methods
4.1. Animals and Establishment of the PD Mouse Model
4.2. Sample Collection in Different Brain Regions
4.3. Total RNA Isolation and cDNA Library Construction
4.4. CircRNA Transcriptome Analysis
4.5. Differentially Expressed circRNAs Enrichment Analysis
4.6. Quantitative Reverse Transcription PCR (qRT-PCR) Analysis
4.7. Construction of circRNA–miRNA–mRNA Interaction Network
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PD | Parkinson’s disease |
| CC | cerebral cortex |
| HP | hippocampus |
| ST | striatum |
| CB | cerebellum |
| DE-circRNAs | differentially expressed circRNAs |
| GO | gene ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| circRNA | circular RNA |
| ceRNA | competing endogenous RNA |
| miRNA | microRNA |
| lncRNA | long-chain noncoding RNA |
References
- Elbassuoni, E.A.; Ahmed, R.F. Mechanism of the neuroprotective effect of GLP-1 in a rat model of Parkinson’s with pre-existing diabetes. Neurochem. Int. 2019, 131, 104583. [Google Scholar] [CrossRef] [PubMed]
- Niedzielska, E.; Smaga, I.; Gawlik, M.; Moniczewski, A.; Stankowicz, P.; Pera, J.; Filip, M. Oxidative stress in neurodegenerative diseases. Neural Regen. Res. 2016, 53, 4094–4125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duty, S.; Jenner, P. Animal models of Parkinson’s disease: A source of novel treatments and clues to the cause of the disease. Br. J. Pharmacol. 2011, 164, 1357–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cookson, M.R.; Hardy, J.; Lewis, P.A. Genetic neuropathology of Parkinson’s disease. Int. J. Clin. Exp. Pathol. 2008, 1, 217–231. [Google Scholar]
- Forno, L.S.; Delanney, L.E.; Irwin, I.; Langston, J.W. Similarities and differences between MPTP-induced parkinsonsim and Parkinson’s disease. Neuropathologic considerations. Adv. Neurol. 1993, 60, 600–608. [Google Scholar] [PubMed]
- Lu, C.; Zhang, J.; Shi, X.; Miao, S.; Bi, L.; Zhang, S.; Yang, Q.; Zhou, X.; Zhang, M.; Xie, Y. Neuroprotective effects of tetramethylpyrazine against dopaminergic neuron injury in a rat model of Parkinson’s disease induced by MPTP. Int. J. Biol. Sci. 2014, 10, 350–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lees, A.J.; Hardy, J.; Revesz, T. Parkinson’s disease[J]. Lancet 2009, 373, 2055–2066. [Google Scholar] [CrossRef]
- Ullah, H.; Khan, H. Anti-Parkinson Potential of Silymarin: Mechanistic Insight and Therapeutic Standing. Front. Pharmacol. 2018, 9, 422. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xue, W.; Li, X.; Zhang, J.; Chen, S.; Zhang, J.L.; Yang, L.; Chen, L.L. The Biogenesis of Nascent Circular RNAs. Cell Rep. 2016, 15, 611–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nat. 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Han, B.; Zhang, Y.; Zhang, Y.; Bai, Y.; Chen, X.; Huang, R.; Wu, F.; Leng, S.; Chao, J.; Zhang, J.H.; et al. Novel insight into circular RNA HECTD1 in astrocyte activation via autophagy by targeting MIR142-TIPARP: Implications for cerebral ischemic stroke. Autophagy 2018, 14, 1164–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.P.; Ye, S.; Long, Y.; Fan, Y.; Mao, H.F.; Chen, M.T.; Ma, Q.J. Circular RNA expression alterations are involved in OGD/R-induced neuron injury. Biochem. Biophys. Res. Commun. 2016, 471, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Yang, X.; Li, X.; Wang, J.; Li, H. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015, 365, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Huiqin, Z.; Mao, X.; Linpei, W.; Hao, X.; Jigui, P.; Jingjing, H.; Lingyun, L.; Jianchun, C. Regulatory network of circRNA-miRNA-mRNA contributes to the histological classification and disease progression in gastric cancer. J. Transl. Med. 2018, 16, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.B.; Niu, Y.L.; Huang, G.X.; Lu, J.J.; Chen, A.; Zhu, L. Silencing of circRNA.2837 Plays a Protective Role in Sciatic Nerve Injury by Sponging the miR-34 Family via Regulating Neuronal Autophagy. Mol. Ther. Nucleic Acids 2018, 12, 718–729. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.; Pan, J.; Ye, X.; Yu, B.; Zhang, W.; Wan, J. Whole-Transcriptome Analysis of APP/PS1 Mouse Brain and Identification of circRNA-miRNA-mRNA Networks to Investigate AD Pathogenesis. Mol. Ther. Nucleic Acids 2019, 18, 1049–1062. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Shi, Q.; Wang, Q.; Tan, X.; Pang, K.; Liu, X.; Zhu, S.; Xi, K.; Zhang, J.; Gao, Q.; et al. Profiling circular RNA in methamphetamine-treated primary cortical neurons identified novel circRNAs related to methamphetamine addiction. Neurosci. Lett. 2019, 701, 146–153. [Google Scholar] [CrossRef]
- You, X.; Vlatkovic, I.; Babic, A.; Will, T.; Epstein, I.; Tushev, G.; Akbalik, G.; Wang, M.; Glock, C.; Quedenau, C.; et al. Neural circular RNAs are derived from synaptic genes and regulated by development and plasticity. Nat. Neurosci. 2015, 18, 603–610. [Google Scholar] [CrossRef] [Green Version]
- Hanan, M.; Soreq, H.; Kadener, S. CircRNAs in the brain. RNA Biol. 2017, 14, 1028–1034. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, J.L.; Day, M.; Peterson, J.D.; Xie, Z.; Kress, G.J.; Rafalovich, I.; Kondapalli, J.; Gertler, T.S.; Flajolet, M.; Greengard, P.; et al. Impaired TrkB receptor signaling underlies corticostriatal dysfunction in Huntington’s disease. Neuron 2014, 83, 178–188. [Google Scholar] [CrossRef] [Green Version]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J.R. Natural RNA circles function as efficient microRNA sponges. Nat. 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Van, R.D.; Verheijen, B.M.; Pasterkamp, R.J. Circular RNAs: Novel Regulators of Neuronal Development. Front. Mol. Neurosci. 2016, 9, 74. [Google Scholar]
- Kumar, L.; Shamsuzzama, J.P.; Haque, R.; Shukla, S.; Nazir, A. Functional Characterization of Novel Circular RNA Molecule, circzip-2 and Its Synthesizing Gene zip-2 in C. elegans Model of Parkinson’s Disease. Mol. Neurobiol. 2018, 55, 6914–6926. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Li, L.; Yu, B.; Jiao, L.; Han, Z.; Zhao, H.; Li, G.; Ma, Y.; Luo, Y. Circular RNA profiling of neutrophil transcriptome provides insights into asymptomatic Moyamoya disease. Brain Res. 2019, 1719, 104–112. [Google Scholar] [CrossRef]
- Huang, J.L.; Xu, Z.H.; Yang, S.M.; Yu, C.; Zhang, F.; Qin, M.C.; Zhou, Y.; Zhong, Z.G.; Wu, D.P. Identification of Differentially Expressed Profiles of Alzheimer’s Disease Associated Circular RNAs in a Panax Notoginseng Saponins-Treated Alzheimer’s Disease Mouse Model. Comput. Struct. Biotechnol. J. 2018, 16, 523–531. [Google Scholar] [CrossRef]
- Zhu, L.; Zhao, R.; Huang, L.; Mo, S.; Yu, Z.; Jiang, L.; Qiao, L. Circular RNA Expression in the Brain of a Neonatal Rat Model of Periventricular White Matter Damage. Cell Physiol. Biochem. 2018, 49, 2264–2276. [Google Scholar] [CrossRef]
- Bai, Y.; Zhang, Y.; Han, B.; Yang, L.; Chen, X.; Huang, R.; Wu, F.; Chao, J.; Liu, P.; Hu, G. Circular RNA DLGAP4 Ameliorates Ischemic Stroke Outcomes by Targeting miR-143 to Regulate Endothelial-Mesenchymal Transition Associated with Blood-Brain Barrier Integrity. J. Neurosci. 2018, 38, 32–50. [Google Scholar]
- Shen, J.; Yang, X.; Xie, B.; Chen, Y.; Swaim, M.; Hackett, S.F.; Campochiaro, P.A. MicroRNAs regulate ocular neovascularization. Mol. Ther. 2008, 16, 1208–1216. [Google Scholar] [CrossRef] [Green Version]
- Miñones-Moyano, E.; Porta, S.; Escaramís, G.; Rabionet, R.; Iraola, S.; Kagerbauer, B.; Espinosa-Parrilla, Y.; Ferrer, I.; Estivill, X.; Martí, E. MicroRNA profiling of Parkinson’s disease brains identifies early downregulation of miR-34b/c which modulate mitochondrial function. Hum. Mol. Genet. 2011, 20, 3067–3078. [Google Scholar] [CrossRef]
- Knowland, D.; Lilascharoen, V.; Pacia, C.P.; Shin, S.; Wang, E.H.; Lim, B.K. Distinct Ventral Pallidal Neural Populations Mediate Separate Symptoms of Depression. Cell 2017, 170, 284–297. [Google Scholar] [CrossRef] [Green Version]
- Rybak-Wolf, A.; Stottmeister, C.; Glažar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinform. 2018, 19, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Li, T.; Wang, Y.; Tang, Y.; Cui, H.; Tang, Y.; Zhang, X.; Chen, D.; Shen, N.; Le, W. miR-132 regulates the differentiation of dopamine neurons by directly targeting Nurr1 expression. J. Cell Sci. 2012, 125, 1673–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lungu, G.; Stoica, G.; Ambrus, A. MicroRNA profiling and the role of microRNA-132 in neurodegeneration using a rat model. Neurosci. Lett. 2013, 553, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, C.; Paiva, J.; Santos, T.; Ferreira, L.; Bernardino, L. MicroRNA-124 loaded nanoparticles enhance brain repair in Parkinson’s disease. J. Control. Release 2016, 235, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Tomiyama, H. A commentary on Axon guidance pathway genes and Parkinson’s disease. J. Hum. Genet. 2011, 56, 102–103. [Google Scholar] [CrossRef] [PubMed]
- Livesey, F.J.; Hunt, S.P. Netrin and netrin receptor expression in the embryonic mammalian nervous system suggests roles in retinal, striatal, nigral, and cerebellar development. Mol. Cell Neurosci. 1997, 8, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Valet, C.; Chicanne, G.; Severac, C.; Chaussade, C.; Whitehead, M.A.; Cabou, C.; Gratacap, M.P.; Gaits-Iacovoni, F.; Vanhaesebroeck, B.; Payrastre, B. Essential role of class II PI3K-C2α in platelet membrane morphology. Blood 2015, 126, 1128–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Liu, T.; Wang, X.; He, A. Circles reshaping the RNA world: From waste to treasure. Mol. Cancer 2017, 16, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Wang, J.; Zhao, F. CIRI: An efficient and unbiased algorithm for de novo circular RNA identification. Genome Biol. 2015, 16, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wang, H.; Jin, L.; Wang, L.; Huang, X.; Chen, W.; Yan, M.; Liu, G. Profile analysis of circRNAs induced by porcine endemic diarrhea virus infection in porcine intestinal epithelial cells. Virol. 2019, 527, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zheng, Y.C.; Kayani, M.U.R.; Xu, W.; Wang, G.Q.; Sun, P.; Ao, N.; Zhang, L.N.; Gu, Z.Q.; Wu, L.C.; et al. Comprehensive analysis of circRNA expression profiles in humans by RAISE. Int. J. Oncol. 2017, 51, 1625–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.; Tu, L.; Li, T.; Yang, X.; Ren, Y.; Gu, R.; Zhang, Q.; Yao, H.; Qu, X.; Wang, Q.; et al. Downregulation of lncRNA UCA1 ameliorates the damage of dopaminergic neurons, reduces oxidative stress and inflammation in Parkinson’s disease through the inhibition of the PI3K/Akt signaling pathway. Int. Immunopharmacol. 2019, 75, 105734. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Duan, M.; Peng, N. Knockdown of TRB3 improved the MPP+/MPTP-induced Parkinson’s disease through the MAPK and AKT signaling pathways. Neurosci. Lett. 2019, 709, 134352. [Google Scholar] [CrossRef]
- Jiao, F.; Wang, Q.; Zhang, P.; Bu, L.; Yan, J.; Tian, B. Expression signatures of long non-coding RNA in the substantia nigra of pre-symptomatic mouse model of Parkinson’s disease. Behav. Brain Res. 2017, 331, 123–130. [Google Scholar] [CrossRef]
- Warre, R.; Thiele, S.; Talwar, S.; Kamal, M.; Johnston, T.H.; Wang, S.; Lam, D.; Lo, C.; Khademullah, C.S.; Perera, G.; et al. Altered function of glutamatergic cortico-striatal synapses causes output pathway abnormalities in a chronic model of parkinsonism. Neurobiol. Dis. 2011, 41, 591–604. [Google Scholar] [CrossRef]
- Feng, Z.; Zhang, L.; Wang, S.; Hong, Q. Circular RNA circDLGAP4 exerts neuroprotective effects via modulating miR-134-5p/CREB pathway in Parkinson’s disease. Biochem. Biophys. Res. Commun. 2019, 522, 388–394. [Google Scholar] [CrossRef]
- Junn, E.; Mouradian, M.M. MicroRNAs in neurodegenerative diseases and their therapeutic potential. Pharmacol. Ther. 2012, 133, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Hansen, K.F.; Sakamoto, K.; Wayman, G.A.; Impey, S.; Obrietan, K. Transgenic miR132 alters neuronal spine density and impairs novel object recognition memory. PLoS ONE 2010, 5, 15497. [Google Scholar] [CrossRef] [Green Version]
- Jankovic, J.; Chen, S.; Le, W.D. The role of Nurr1 in the development of dopaminergic neurons and Parkinson’s disease. Prog. Neurobiol. 2005, 77, 128–138. [Google Scholar] [CrossRef]
- Wang, H.; Ye, Y.; Zhu, Z.; Mo, L.; Lin, C.; Wang, Q.; Wang, H.; Gong, X.; He, X.; Lu, G.; et al. MiR-124 Regulates Apoptosis and Autophagy Process in MPTP Model of Parkinson’s Disease by Targeting to Bim. Brain Pathol. 2016, 26, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Kanagaraj, N.; Beiping, H.; Dheen, S.T.; Tay, S.S. Downregulation of miR-124 in MPTP-treated mouse model of Parkinson’s disease and MPP iodide-treated MN9D cells modulates the expression of the calpain/cdk5 pathway proteins. Neurosci. 2014, 272, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Li, S.D.; Liu, Y.; Yang, M.H. Effects of bushen huoxue yin on brain NF-kB and NO content in the Parkinson’s disease model mouse. J. Tradit. Chin. Med. 2012, 32, 67–70. [Google Scholar] [CrossRef] [Green Version]
- Bulteau, A.L.; Mena, N.P.; Auchère, F.; Lee, I.; Prigent, A.; Lobsiger, C.S.; Camadro, J.M.; Hirsch, E.C. Dysfunction of mitochondrial Lon protease and identification of oxidized protein in mouse brain following exposure to MPTP: Implications for Parkinson disease. Free Radic. Biol. Med. 2017, 108, 236–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.Y.; Hsieh, H.Y.; Chen, C.M.; Wu, S.R.; Tsai, C.H.; Huang, C.Y.; Hua, M.Y.; Wei, K.C.; Yeh, C.K.; Liu, H.L. Non-invasive, neuron-specific gene therapy by focused ultrasound-induced blood-brain barrier opening in Parkinson’s disease mouse model. J. Control. Release 2016, 235, 72–81. [Google Scholar] [CrossRef]
- Alieva, A.K.; Zyrin, V.S.; Rudenok, M.M.; Kolacheva, A.A.; Shulskaya, M.V.; Ugryumov, M.V.; Slominsky, P.A.; Shadrina, M. Whole-Transcriptome Analysis of Mouse Models with MPTP-Induced Early Stages of Parkinson’s Disease Reveals Stage-Specific Response of Transcriptome and a Possible Role of Myelin-Linked Genes in Neurodegeneration. Mol. Neurobiol. 2018, 55, 7229–7241. [Google Scholar] [CrossRef]
- Middleton, F.A.; Strick, P.L. Basal ganglia and cerebellar loops: Motor and cognitive circuits. Brain Res. Rev. 2000, 31, 236–250. [Google Scholar] [CrossRef]
- Ichinohe, N.; Mori, F.; Shoumura, K. A di-synaptic projection from the lateral cerebellar nucleus to the laterodorsal part of the striatum via the central lateral nucleus of the thalamus in the rat. Brain Res. 2000, 880, 191–197. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequences |
|---|---|
| GAPDH | F: 5’-CACTGAGCAAGAGAGGCCCTAT-3’ R: 5’-GCAGCGAACTTTATTGATGGTATT-3’ |
| mmu_circRNA_0004144 | F: 5’-TTAGCAGAGGAGCAAGCGTT-3’ R: 5’-TGCTCCTGAACCTGAAAATGT-3’ |
| mmu_circRNA_0000468 | F: 5’-CGTCACCAATCACACGGAGT-3’ R: 5’-CAGATGGCAGACCGTAGTCG-3’ |
| mmu_circRNA_0013321 | F: 5’-GTCTGACTGGTGGAACCCTG-3’ R: 5’-CCCAGAGGGATGGTGTAGCA-3’ |
| mmu_circRNA_0001320 | F: 5’-ACTCTGCTCGGGCGGT-3’ R: 5’-GCCGGCTTCGTGGATAATCT-3’ |
| mmu_circRNA_0003292 | F: GGTTTCACAGGTCTGGCGT R: CAACCGTTCCGTGGCTAACT |
| mmu_circRNA_0005157 | F: GCAGCATTTGCAGGCACATAA R: AGCCTTGCCACACTCATCTC |
| mmu_circRNA_0000870 | F: CGTCTAGAAGCATTGGGGCA R: CCACCATGAATAACTTTCACAAGC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, E.; Zhou, Y.; Liu, Z.; Wang, L.; Ouyang, T.; Pan, M.; Bai, Y.; Ge, Q. Transcriptomic Profiling of Circular RNA in Different Brain Regions of Parkinson’s Disease in a Mouse Model. Int. J. Mol. Sci. 2020, 21, 3006. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21083006
Jia E, Zhou Y, Liu Z, Wang L, Ouyang T, Pan M, Bai Y, Ge Q. Transcriptomic Profiling of Circular RNA in Different Brain Regions of Parkinson’s Disease in a Mouse Model. International Journal of Molecular Sciences. 2020; 21(8):3006. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21083006
Chicago/Turabian StyleJia, Erteng, Ying Zhou, Zhiyu Liu, Liujing Wang, Tinglan Ouyang, Min Pan, Yunfei Bai, and Qinyu Ge. 2020. "Transcriptomic Profiling of Circular RNA in Different Brain Regions of Parkinson’s Disease in a Mouse Model" International Journal of Molecular Sciences 21, no. 8: 3006. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21083006