Human NANOS1 Represses Apoptosis by Downregulating Pro-Apoptotic Genes in the Male Germ Cell Line

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results
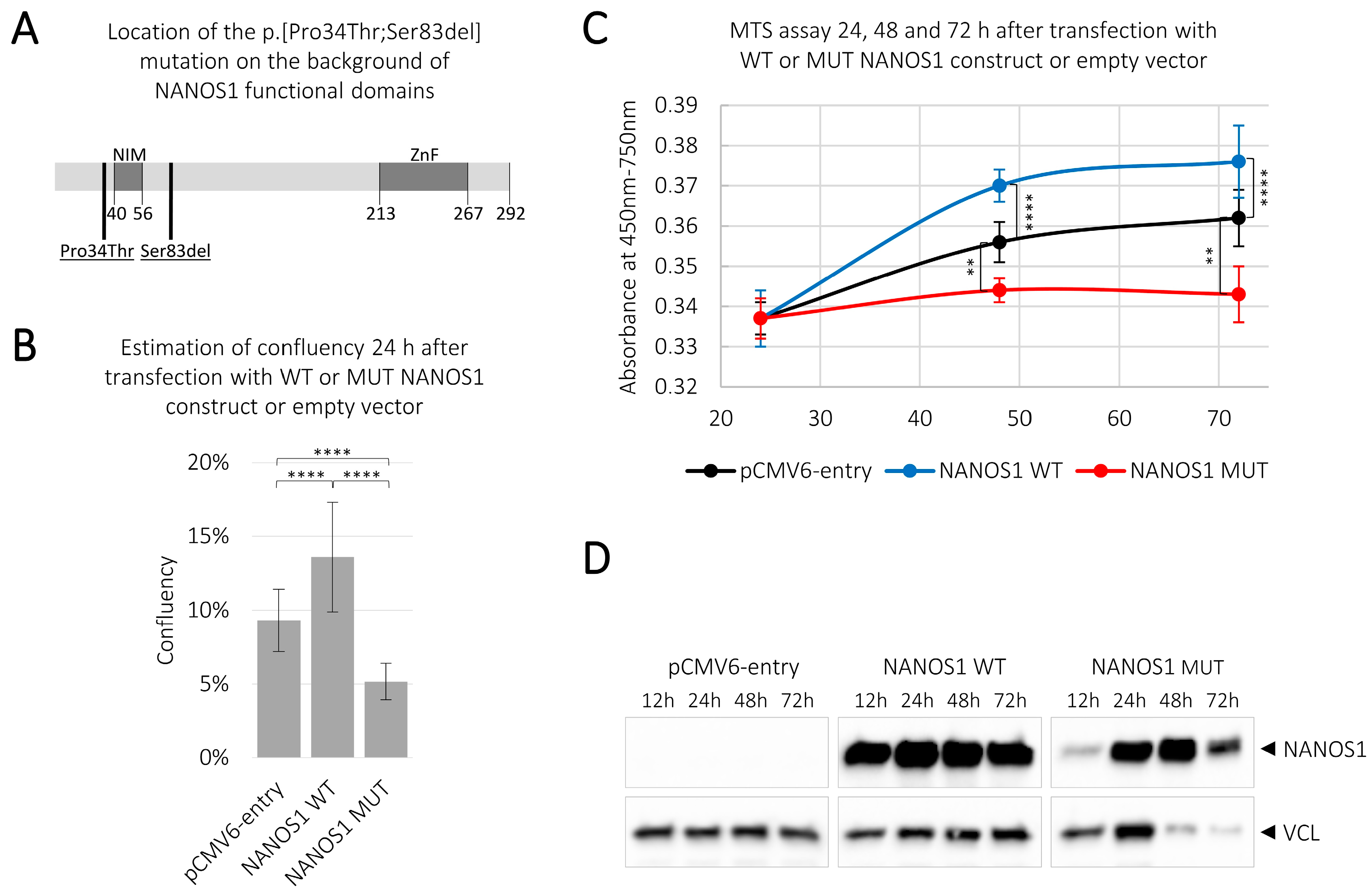
2.1. Overexpression of the Wild-Type NANOS1 Increases, While the Mutated NANOS1 p.[(Pro34Thr);(Ser83del)] Decreases, the Proliferation of TCam-2 cells
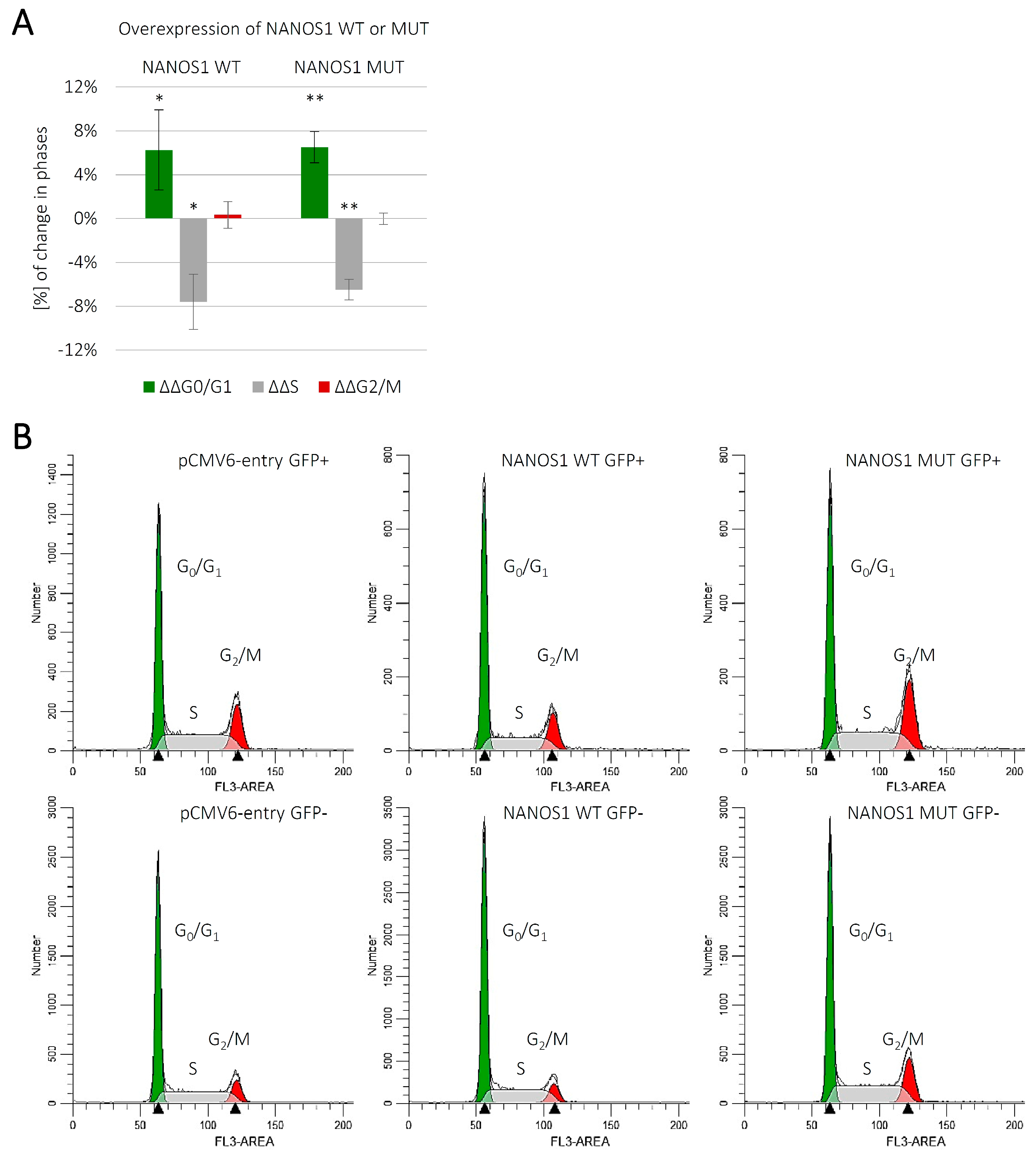
2.2. Overexpression of Both the Wild-Type and the Mutated NANOS1 Inhibits the Cell Cycle of TCam-2 cells
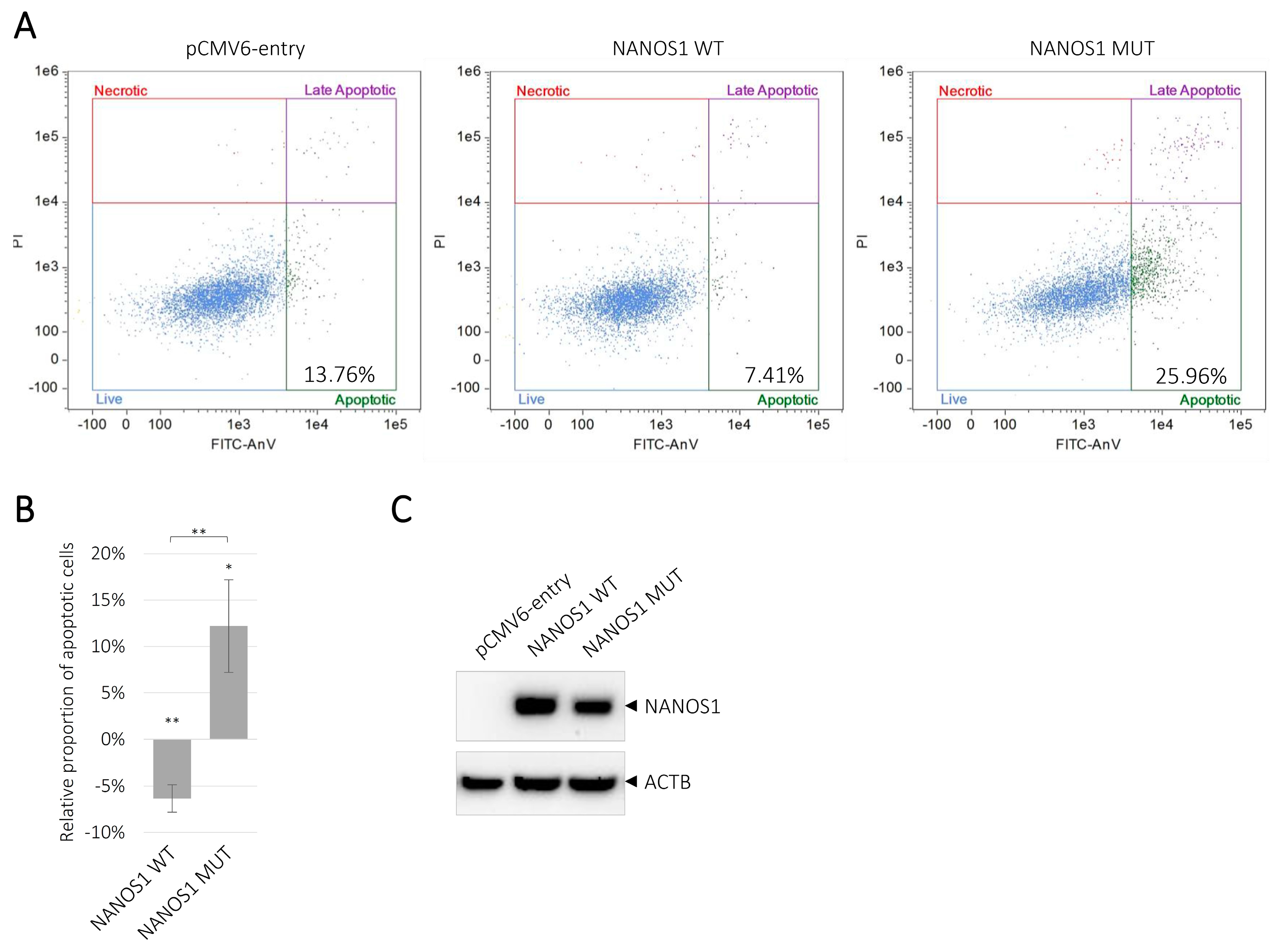
2.3. Overexpression of the Wild-Type NANOS1 Decreases, Whereas the Mutated NANOS1 p.[(Pro34Thr);(Ser83del)] Increases Apoptosis of TCam-2 cells
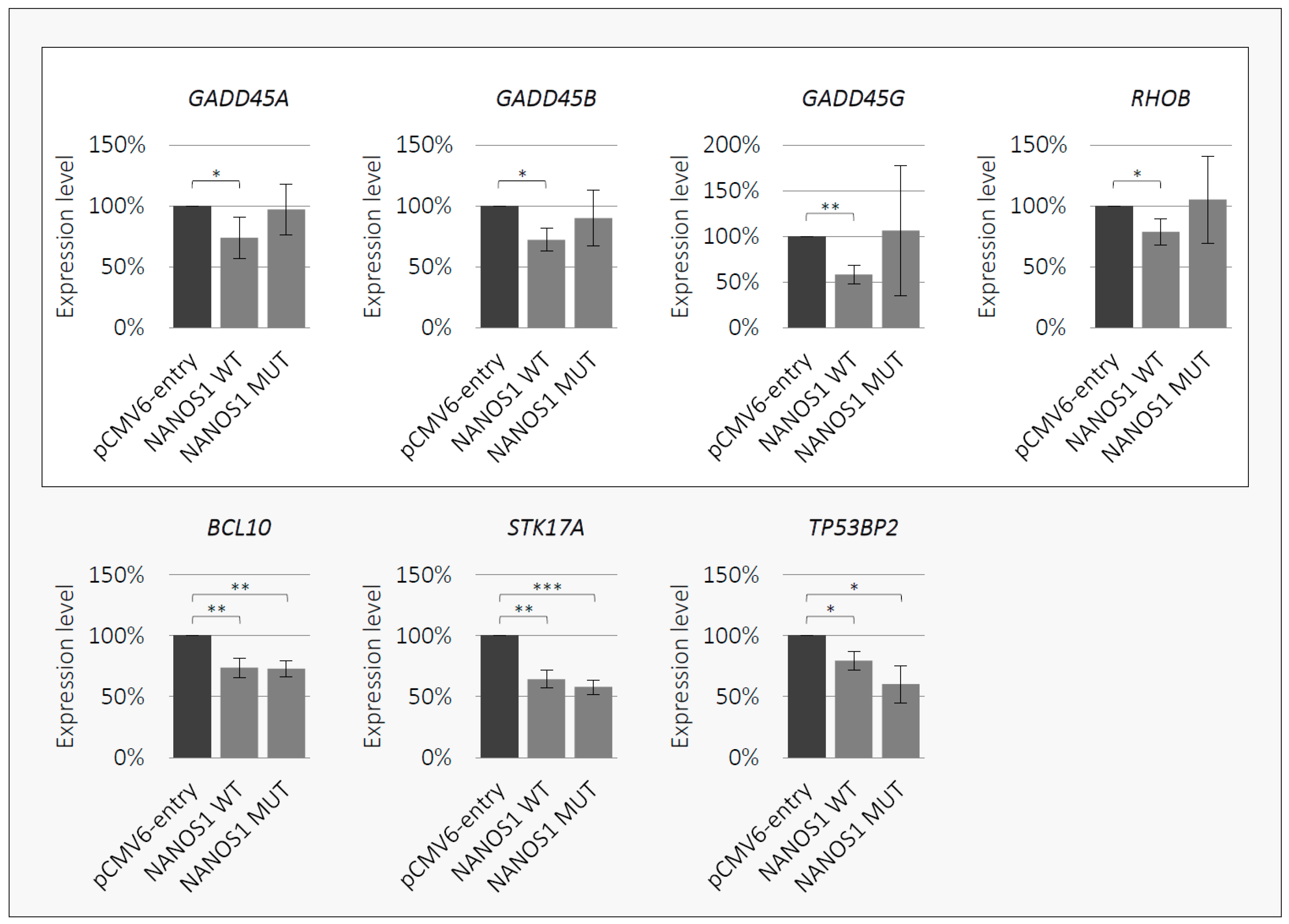
2.4. NANOS1 Negatively Regulates Pro-Apoptotic Genes and This Regulation is Disrupted by the p.[(Pro34Thr);(Ser83del)] Mutation
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Transfection
4.2. Estimation of Cell Confluency
4.3. MTS Assay
4.4. Western Blotting
4.5. Antibodies
4.6. Flow Cytometry
4.7. cDNA Library Preparation and RNA-Seq
4.8. Bioinformatic Analysis
4.9. Quantitative RT-PCR
4.10. Constructs
4.11. Accession Numbers
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACTB | actin beta |
| cDNA | complementary DNA |
| FPKM | fragments per kilobase of transcript per million mapped reads |
| GADD45 | growth arrest and DNA damage inducible 45 |
| hid | head involution defective |
| HRP | horseradish peroxidase |
| mRNA | messenger RNA |
| MTS | (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium) |
| MUT | mutated |
| NIM | NOT1 interacting motif |
| PGCs | primordial germ cells |
| POI | premature ovarian insufficiency |
| RHOB | ras homologue family member B |
| RNA-Seq | RNA sequencing |
| RT-qPCR | reverse transcription and quantitative polymerase chain reaction |
| SCOS | Sertoli cell-only syndrome |
| skl | sickle |
| TGCT | testicular germ cell tumors |
| VCL | vinculin |
| WT | wild-type |
| ZnF | zinc-finger domain |
References
- Shaha, C.; Tripathi, R.; Mishra, D.P. Male germ cell apoptosis: Regulation and biology. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 1501–1515. [Google Scholar] [CrossRef] [Green Version]
- De Keuckelaere, E.; Hulpiau, P.; Saeys, Y.; Berx, G.; Van Roy, F. Nanos genes and their role in development and beyond. Cell. Mol. Life Sci. 2018, 75, 1929–1946. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Hayashi, Y.; Ninomiya, Y.; Shigenobu, S.; Arita, K.; Mukai, M.; Kobayashi, S. Maternal Nanos represses hid/skl-dependent apoptosis to maintain the germ line in Drosophila embryos. Proc. Natl. Acad. Sci. USA 2007, 104, 7455–7460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, K.M. The posterior determinant gene nanos is required for the maintenance of the adult germline stem cells during Drosophila oogenesis. Genetics 1999, 151, 1479–1492. [Google Scholar] [PubMed]
- Suzuki, A.; Saga, Y. Nanos2 suppresses meiosis and promotes male germ cell differentiation. Genes Dev. 2008, 22, 430–435. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, M.; Sasaoka, Y.; Kiso, M.; Abe, K.; Haraguchi, S.; Kobayashi, S.; Saga, Y. Conserved role of nanos proteins in germ cell development. Science 2003, 301, 1239–1241. [Google Scholar] [CrossRef]
- Suzuki, A.; Tsuda, M.; Saga, Y. Functional redundancy among Nanos proteins and a distinct role of Nanos2 during male germ cell development. Development 2007, 134, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Haraguchi, S.; Tsuda, M.; Kitajima, S.; Sasaoka, Y.; Nomura-Kitabayashid, A.; Kurokawa, K.; Saga, Y. nanos1: A mouse nanos gene expressed in the central nervous system is dispensable for normal development. Mech. Dev. 2003, 120, 721–731. [Google Scholar] [CrossRef]
- Jaruzelska, J.; Kotecki, M.; Kusz, K.; Spik, A.; Firpo, M.; Reijo Pera, R.A. Conservation of a Pumilio-Nanos complex from Drosophila germ plasm to human germ cells. Dev. Genes Evol. 2003, 213, 120–126. [Google Scholar] [CrossRef]
- Julaton, V.T.; Reijo Pera, R.A. NANOS3 function in human germ cell development. Hum. Mol. Genet. 2011, 20, 2238–2250. [Google Scholar] [CrossRef] [Green Version]
- Kusz, K.; Tomczyk, L.; Spik, A.; Latos-Bielenska, A.; Jedrzejczak, P.; Pawelczyk, L.; Jaruzelska, J. NANOS3 gene mutations in men with isolated sterility phenotype. Mol. Reprod. Dev. 2009, 76, 804. [Google Scholar] [CrossRef] [PubMed]
- Kusz, K.M.; Tomczyk, L.; Sajek, M.; Spik, A.; Latos-Bielenska, A.; Jedrzejczak, P.; Pawelczyk, L.; Jaruzelska, J. The highly conserved NANOS2 protein: Testis-specific expression and significance for the human male reproduction. Mol. Hum. Reprod. 2009, 15, 165–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Y.; Zhao, H.; Kovanci, E.; Simpson, J.L.; Chen, Z.J.; Rajkovic, A. Mutation analysis of NANOS3 in 80 Chinese and 88 Caucasian women with premature ovarian failure. Fertil. Steril. 2007, 88, 1465–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, M.G.; Machado, A.Z.; Martins, C.N.; Domenice, S.; Costa, E.M.; Nishi, M.Y.; Ferraz-de-Souza, B.; Jorge, S.A.; Pereira, C.A.; Soardi, F.C.; et al. Homozygous inactivating mutation in NANOS3 in two sisters with primary ovarian insufficiency. BioMed Res. Int. 2014, 2014, 787465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, B.L.; Nishi, M.Y.; Santos, M.G.; Brito, V.N.; Domenice, S.; Mendonca, B.B. Mutation analysis of NANOS3 in Brazilian women with primary ovarian failure. Clinics 2016, 71, 695–698. [Google Scholar] [CrossRef]
- Wu, X.; Wang, B.; Dong, Z.; Zhou, S.; Liu, Z.; Shi, G.; Cao, Y.; Xu, Y. A NANOS3 mutation linked to protein degradation causes premature ovarian insufficiency. Cell Death Dis. 2013, 4, e825. [Google Scholar] [CrossRef] [Green Version]
- Kusz-Zamelczyk, K.; Sajek, M.; Spik, A.; Glazar, R.; Jedrzejczak, P.; Latos-Bielenska, A.; Kotecki, M.; Pawelczyk, L.; Jaruzelska, J. Mutations of NANOS1, a human homologue of the Drosophila morphogen, are associated with a lack of germ cells in testes or severe oligo-astheno-teratozoospermia. J. Med. Genet. 2013, 50, 187–193. [Google Scholar] [CrossRef]
- Bhandari, D.; Raisch, T.; Weichenrieder, O.; Jonas, S.; Izaurralde, E. Structural basis for the Nanos-mediated recruitment of the CCR4-NOT complex and translational repression. Genes Dev. 2014, 28, 888–901. [Google Scholar] [CrossRef]
- De Jong, J.; Stoop, H.; Gillis, A.J.; Hersmus, R.; Van Gurp, R.J.; Van de Geijn, G.J.; Van Drunen, E.; Beverloo, H.B.; Schneider, D.T.; Sherlock, J.K.; et al. Further characterization of the first seminoma cell line TCam-2. Genes Chromosomes Cancer 2008, 47, 185–196. [Google Scholar] [CrossRef]
- Smialek, M.J. Human PUM1 and PUM2 exhibit regulation of divergent mRNA targets in male germ cells. Biorxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Takekawa, M.; Saito, H. A family of stress-inducible GADD45-like proteins mediate activation of the stress-responsive MTK1/MEKK4 MAPKKK. Cell 1998, 95, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Fritz, G.; Kaina, B.; Aktories, K. The ras-related small GTP-binding protein RhoB is immediate-early inducible by DNA damaging treatments. J. Biol. Chem. 1995, 270, 25172–25177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapp, T.; Olah, L.; Holker, I.; Besselmann, M.; Tiesler, C.; Maeda, K.; Hossmann, K.A. GTPase RhoB: An early predictor of neuronal death after transient focal ischemia in mice. Mol. Cell. Neurosci. 2001, 17, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Harkin, D.P.; Bean, J.M.; Miklos, D.; Song, Y.H.; Truong, V.B.; Englert, C.; Christians, F.C.; Ellisen, L.W.; Maheswaran, S.; Oliner, J.D.; et al. Induction of GADD45 and JNK/SAPK-dependent apoptosis following inducible expression of BRCA1. Cell 1999, 97, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Marlow, L.A.; Bok, I.; Smallridge, R.C.; Copland, J.A. RhoB upregulation leads to either apoptosis or cytostasis through differential target selection. Endocr. Relat. Cancer 2015, 22, 777–792. [Google Scholar] [CrossRef] [Green Version]
- Srougi, M.C.; Burridge, K. The nuclear guanine nucleotide exchange factors Ect2 and Net1 regulate RhoB-mediated cell death after DNA damage. PLoS ONE 2011, 6, e17108. [Google Scholar] [CrossRef]
- Adnane, J.; Muro-Cacho, C.; Mathews, L.; Sebti, S.M.; Munoz-Antonia, T. Suppression of rho B expression in invasive carcinoma from head and neck cancer patients. Clin. Cancer Res. 2002, 8, 2225–2232. [Google Scholar]
- Liu, B.; Yang, L.; Li, X.J.; Li, R.; Sun, W.; Chen, X.Y.; Liu, J.C. Expression and significance of ASPP2 in squamous carcinoma of esophagus. Kaohsiung J. Med. Sci. 2018, 34, 321–329. [Google Scholar] [CrossRef]
- Mazieres, J.; Antonia, T.; Daste, G.; Muro-Cacho, C.; Berchery, D.; Tillement, V.; Pradines, A.; Sebti, S.; Favre, G. Loss of RhoB expression in human lung cancer progression. Clin. Cancer Res. 2004, 10, 2742–2750. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, K.; Sha, X.; Magimaidas, A.; Maifrede, S.; Skorski, T.; Bhatia, R.; Hoffman, B.; Liebermann, D.A. Gadd45a deficiency accelerates BCR-ABL driven chronic myelogenous leukemia. Oncotarget 2017, 8, 10809–10821. [Google Scholar] [CrossRef]
- Prendergast, G.C. Farnesyltransferase inhibitors define a role for RhoB in controlling neoplastic pathophysiology. Histol. Histopathol. 2001, 16, 269–275. [Google Scholar] [PubMed]
- Sha, X.; Hoffman, B.; Liebermann, D.A. Loss of Gadd45b accelerates BCR-ABL-driven CML. Oncotarget 2018, 9, 33360–33367. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Bian, Q.; Zhang, Y.J.; Shao, C.H.; Li, G.; Liu, A.A.; Jing, W.; Liu, R.; Zhou, Y.Q.; Jin, G.; et al. Downregulation of ASPP2 in pancreatic cancer cells contributes to increased resistance to gemcitabine through autophagy activation. Mol. Cancer 2015, 14, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, L.; Deng, Z.; Xu, L.; Yang, T.; Yao, W.; Ji, L.; Lu, Y.; Zhang, J.; Liu, Y.; Wang, J. Downregulation of ASPP2 promotes gallbladder cancer metastasis and macrophage recruitment via aPKC-iota/GLI1 pathway. Cell Death Dis. 2018, 9, 1115. [Google Scholar] [CrossRef]
- Wu, T.; Song, H.; Xie, D.; Zhao, B.; Xu, H.; Wu, C.; Hua, K.; Deng, Y.; Ji, C.; Hu, J.; et al. Silencing of ASPP2 promotes the proliferation, migration and invasion of triple-negative breast cancer cells via the PI3K/AKT pathway. Int. J. Oncol. 2018, 52, 2001–2010. [Google Scholar] [CrossRef] [Green Version]
- Bonnomet, A.; Polette, M.; Strumane, K.; Gilles, C.; Dalstein, V.; Kileztky, C.; Berx, G.; Van Roy, F.; Birembaut, P.; Nawrocki-Raby, B. The E-cadherin-repressed hNanos1 gene induces tumor cell invasion by upregulating MT1-MMP expression. Oncogene 2008, 27, 3692–3699. [Google Scholar] [CrossRef] [Green Version]
- Blackinton, J.G.; Keene, J.D. Post-transcriptional RNA regulons affecting cell cycle and proliferation. Semin. Cell Dev. Biol. 2014, 34, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janecki, D.M.; Sajek, M.; Smialek, M.J.; Kotecki, M.; Ginter-Matuszewska, B.; Kuczynska, B.; Spik, A.; Kolanowski, T.; Kitazawa, R.; Kurpisz, M.; et al. SPIN1 is a proto-oncogene and SPIN3 is a tumor suppressor in human seminoma. Oncotarget 2018, 9, 32466–32477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sajek, M.; Janecki, D.M.; Smialek, M.J.; Ginter-Matuszewska, B.; Spik, A.; Oczkowski, S.; Ilaslan, E.; Kusz-Zamelczyk, K.; Kotecki, M.; Blazewicz, J.; et al. PUM1 and PUM2 exhibit different modes of regulation for SIAH1 that involve cooperativity with NANOS paralogues. Cell. Mol. Life Sci. 2019, 76, 147–161. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HGNC | Base Mean | log2 Fold Change | Adjusted p Value | p Value | Standard Error |
|---|---|---|---|---|---|
| SIAH1 | 26.98 | −0.78 | 9.31 × 10−3 | 1.11 × 10−3 | 0.24 |
| GADD45B | 311.44 | −0.65 | 6.91 × 10−4 | 3.08 × 10−5 | 0.16 |
| GADD45G | 334.60 | −0.64 | 4.47 × 10−3 | 3.97 × 10−4 | 0.18 |
| BCL10 | 32.64 | −0.64 | 3.32 × 10−2 | 6.68 × 10−3 | 0.24 |
| STK17A | 392.07 | −0.62 | 7.51 × 10−4 | 3.48 × 10−5 | 0.15 |
| JUN | 234.83 | −0.62 | 1.63 × 10−3 | 1.00 × 10−4 | 0.16 |
| GADD45A | 1051.99 | −0.55 | 8.22 × 10−4 | 3.93 × 10−5 | 0.13 |
| TP53BP2 | 894.20 | −0.51 | 4.09 × 10−4 | 1.57 × 10−5 | 0.12 |
| RHOB | 516.17 | −0.42 | 3.00 × 10−2 | 5.78 × 10−3 | 0.15 |
| RIPK1 | 294.70 | −0.41 | 3.69 × 10−2 | 7.70 × 10−3 | 0.15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janecki, D.M.; Ilaslan, E.; Smialek, M.J.; Sajek, M.P.; Kotecki, M.; Ginter-Matuszewska, B.; Krainski, P.; Jaruzelska, J.; Kusz-Zamelczyk, K. Human NANOS1 Represses Apoptosis by Downregulating Pro-Apoptotic Genes in the Male Germ Cell Line. Int. J. Mol. Sci. 2020, 21, 3009. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21083009
Janecki DM, Ilaslan E, Smialek MJ, Sajek MP, Kotecki M, Ginter-Matuszewska B, Krainski P, Jaruzelska J, Kusz-Zamelczyk K. Human NANOS1 Represses Apoptosis by Downregulating Pro-Apoptotic Genes in the Male Germ Cell Line. International Journal of Molecular Sciences. 2020; 21(8):3009. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21083009
Chicago/Turabian StyleJanecki, Damian M., Erkut Ilaslan, Maciej J. Smialek, Marcin P. Sajek, Maciej Kotecki, Barbara Ginter-Matuszewska, Patryk Krainski, Jadwiga Jaruzelska, and Kamila Kusz-Zamelczyk. 2020. "Human NANOS1 Represses Apoptosis by Downregulating Pro-Apoptotic Genes in the Male Germ Cell Line" International Journal of Molecular Sciences 21, no. 8: 3009. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21083009