Role of Non-Coding RNAs in Lung Circadian Clock Related Diseases

 , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Mechanism of Circadian Clock
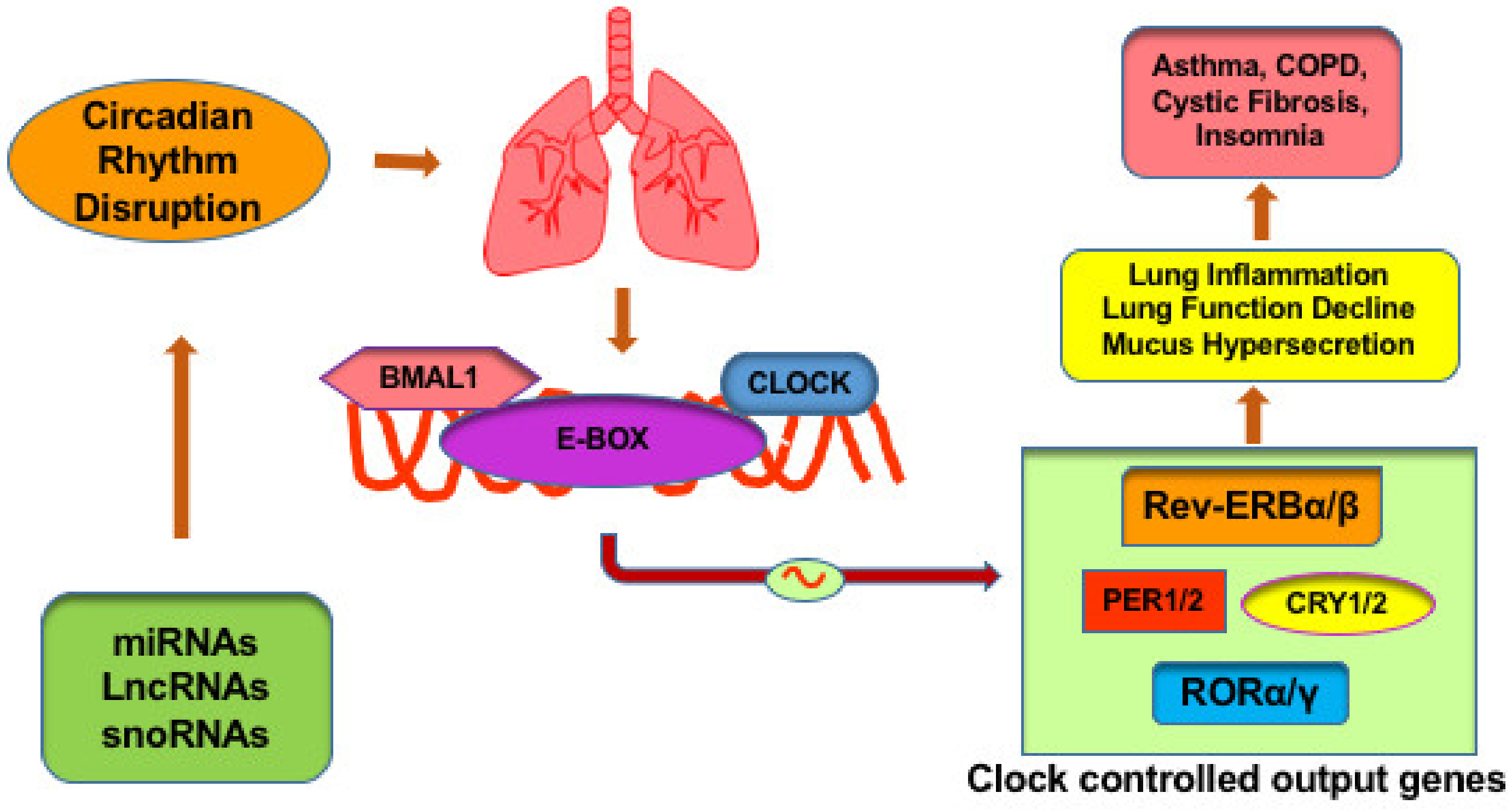
2.1. Circadian Molecular Clock in Lung Related Diseases
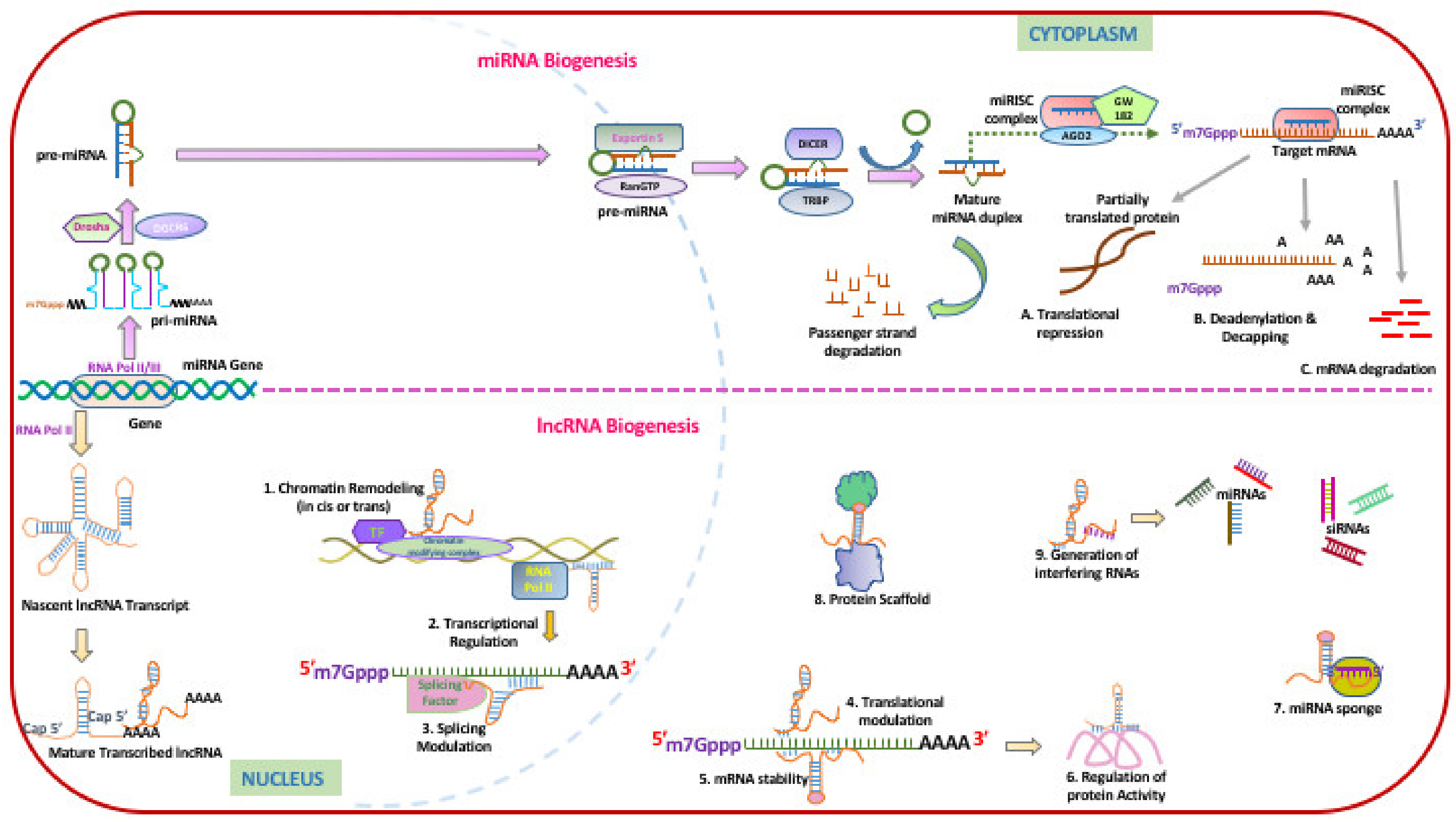
2.2. Role of Non-Coding RNAs in Molecular Clock Regulation: Implications to the Lung
2.3. Role of MicroRNAs in Molecular Clock Regulation: Implications for the Lung
2.4. TGF-β in Altered microRNAome and Its Impact on Clock Regulation: Implications to the Lung
2.5. Role of Non-Coding RNAs in Molecular Clock Regulation: Implication for the Lung
2.6. Role of Small Nucleolar RNAs (snoRNAs) in Molecular Clock Regulation: Implications to the Lung
3. Conclusions
Funding
Conflicts of Interest
References
- Skloot, G.S. Nocturnal Asthma: Mechanisms and Management; The Mount Sinai journal of medicine: New York, NY, USA, 2002; Volume 69, pp. 140–147. [Google Scholar]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef] [Green Version]
- Aschoff, J. Exogenous and Endogenous Components in Circadian Rhythms. Cold Spring Harb. Symp. Quant. Boil. 1960, 25, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Circadian variation in airway function. Am. J. Med. 1985, 79, 5–9. [Google Scholar] [CrossRef]
- Bechtold, D.A.; Gibbs, J.E.; Loudon, A.S. Circadian dysfunction in disease. Trends Pharmacol. Sci. 2010, 31, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Karbasforooshan, H.; Roohbakhsh, A.; Karimi, G. SIRT1 and microRNAs: The role in breast, lung and prostate cancers. Exp. Cell Res. 2018, 367, 1–6. [Google Scholar] [CrossRef]
- Dutta, R.K.; Chinnapaiyan, S.; Rasmussen, L.; Raju, S.V.; Unwalla, H. A Neutralizing Aptamer to TGFBR2 and miR-145 Antagonism Rescue Cigarette Smoke- and TGF-β-Mediated CFTR Expression. Mol. Ther. 2019, 27, 442–455. [Google Scholar] [CrossRef] [Green Version]
- Hoegh, S.V.; Sorensen, G.L.; Tornoe, I.; Lottenburger, T.; Ytting, H.; Nielsen, H.J.; Junker, P.; Holmskov, U. Long-term stability and circadian variation in circulating levels of surfactant protein D. Immunobiology 2010, 215, 314–320. [Google Scholar] [CrossRef]
- Hwang, J.-W.; Sundar, I.K.; Yao, H.; Sellix, M.T.; Rahman, I. Circadian clock function is disrupted by environmental tobacco/cigarette smoke, leading to lung inflammation and injury via a SIRT1-BMAL1 pathway. FASEB J. 2013, 28, 176–194. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Sundar, I.K.; Huang, Y.; Gerloff, J.; Sellix, M.T.; Sime, P.J.; Rahman, I. Disruption of Sirtuin 1–Mediated Control of Circadian Molecular Clock and Inflammation in Chronic Obstructive Pulmonary Disease. Am. J. Respir. Cell Mol. Boil. 2015, 53, 782–792. [Google Scholar] [CrossRef]
- Buhr, E.D.; Takahashi, J.S. Molecular components of the Mammalian circadian clock. Handb. Exp. Pharmacol. 2013, 217, 3–27. [Google Scholar] [CrossRef] [Green Version]
- Lowrey, P.L.; Takahashi, J.S. Genetics of Circadian Rhythms in Mammalian Model Organisms. Adv. Genet. 2011, 74, 175–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gekakis, N. Role of the CLOCK Protein in the Mammalian Circadian Mechanism. Science 1998, 280, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.-H.; Ko, C.H.; Lowrey, P.L.; Buhr, E.D.; Song, E.-J.; Chang, S.; Yoo, O.J.; Yamazaki, S.; Lee, C.; Takahashi, J.S. A noncanonical E-box enhancer drives mouse Period2 circadian oscillations in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 2608–2613. [Google Scholar] [CrossRef] [Green Version]
- Ohno, T.; Onishi, Y.; Ishida, N. A novel E4BP4 element drives circadian expression of mPeriod2. Nucleic Acids Res. 2006, 35, 648–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- HogenEsch, J.B.; Gu, Y.-Z.; Jain, S.; Bradfield, C.A. The basic-helix-loop-helix-PAS orphan MOP3 forms transcriptionally active complexes with circadian and hypoxia factors. Proc. Natl. Acad. Sci. USA 1998, 95, 5474–5479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kume, K.; Zylka, M.J.; Sriram, S.; Shearman, L.P.; Weaver, D.; Jin, X.; Maywood, E.S.; Hastings, M.H.; Reppert, S.M. mCRY1 and mCRY2 are essential components of the negative limb of the circadian clock feedback loop. Cell 1999, 98, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Griffin, E.A.; Staknis, D.; Weitz, C.J. Light-Independent Role of CRY1 and CRY2 in the Mammalian Circadian Clock. Science 1999, 286, 768–771. [Google Scholar] [CrossRef]
- Sangoram, A.M.; Sáez, L.; Young, M.W.; Weitz, C.J.; Takahashi, J.S.; Antoch, M.P.; Gekakis, N.; Staknis, D.; Whiteley, A.; Fruechte, E.M.; et al. Mammalian Circadian Autoregulatory Loop: A timeless ortholog and mPer1 interact and negatively regulate CLOCK-BMAL1-induced transcription. Neuron 1998, 21, 1101–1113. [Google Scholar] [CrossRef] [Green Version]
- Field, M.D.; Maywood, E.S.; A O’Brien, J.; Weaver, D.; Reppert, S.M.; Hastings, M.H. Analysis of clock proteins in mouse SCN demonstrates phylogenetic divergence of the circadian clockwork and resetting mechanisms. Neuron 2000, 25, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.K.; Yamada, R.G.; Ukai, H.; E Baggs, J.; Miraglia, L.J.; Kobayashi, T.J.; Welsh, D.K.; A Kay, S.; Ueda, H.R.; HogenEsch, J.B. Feedback repression is required for mammalian circadian clock function. Nat. Genet. 2006, 38, 312–319. [Google Scholar] [CrossRef]
- Li, N.; Ren, A.; Wang, X.; Fan, X.; Zhao, Y.; Gao, G.F.; Cleary, P.; Wang, B. Influenza viral neuraminidase primes bacterial coinfection through TGF-β–mediated expression of host cell receptors. Proc. Natl. Acad. Sci. USA 2014, 112, 238–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakahata, Y.; Kaluzova, M.; Grimaldi, B.; Sahar, S.; Hirayama, J.; Chen, D.; Guarente, L.P.; Sassone-Corsi, P. The NAD+-Dependent Deacetylase SIRT1 Modulates CLOCK-Mediated Chromatin Remodeling and Circadian Control. Cell 2008, 134, 329–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asher, G.; Gatfield, D.; Stratmann, M.; Reinke, H.; Dibner, C.; Kreppel, F.; Mostoslavsky, R.; Alt, F.W.; Schibler, U. SIRT1 Regulates Circadian Clock Gene Expression through PER2 Deacetylation. Cell 2008, 134, 317–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutter, J.; Reick, M.; Wu, L.C.; McKnight, S.L. Regulation of Clock and NPAS2 DNA Binding by the Redox State of NAD Cofactors. Science 2001, 293, 510–514. [Google Scholar] [CrossRef] [Green Version]
- Sundar, I.K.; Sellix, M.T.; Rahman, I. Redox regulation of circadian molecular clock in chronic airway diseases. Free Radic. Boil. Med. 2018, 119, 121–128. [Google Scholar] [CrossRef]
- Hirayama, J.; Sahar, S.; Grimaldi, B.; Tamaru, T.; Takamatsu, K.; Nakahata, Y.; Sassone-Corsi, P. CLOCK-mediated acetylation of BMAL1 controls circadian function. Nature 2007, 450, 1086–1090. [Google Scholar] [CrossRef]
- Trump, R.P.; Bresciani, S.; Cooper, A.W.J.; Tellam, J.P.; Wojno, J.; Blaikley, J.F.; Orband-Miller, L.A.; Kashatus, J.A.; Boudjelal, M.; Dawson, H.C.; et al. Optimized Chemical Probes for REV-ERBα. J. Med. Chem. 2013, 56, 4729–4737. [Google Scholar] [CrossRef]
- Gibbs, J.E.; Beesley, S.; Plumb, J.; Singh, D.; Farrow, S.; Ray, D.W.; Loudon, A. Circadian timing in the lung; a specific role for bronchiolar epithelial cells. Endocrinology 2008, 150, 268–276. [Google Scholar] [CrossRef]
- Korenčič, A.; Kosir, R.; Bordyugov, G.; Lehmann, R.; Rozman, D.; Herzel, H. Timing of circadian genes in mammalian tissues. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.A.; Davidson, A.J. Health Consequences of Circadian Disruption in Humans and Animal Models. Prog. Mol. Biol. Transl. Sci. 2013, 119, 283–323. [Google Scholar]
- Marcheva, B.; Ramsey, K.M.; Peek, C.B.; Affinati, A.; Maury, E.; Bass, J. Circadian clocks and metabolism. Handb. Exp. Pharmacol. 2013, 217, 127–155. [Google Scholar] [CrossRef] [Green Version]
- Sundar, I.K.; Yao, H.; Sellix, M.T.; Rahman, I. Circadian molecular clock in lung pathophysiology. Am. J. Physiol. Cell. Mol. Physiol. 2015, 309, L1056–L1075. [Google Scholar] [CrossRef] [PubMed]
- Mortola, J.P.; Seifert, E.L. Circadian patterns of breathing. Respir. Physiol. Neurobiol. 2002, 131, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Hadden, H.; Soldin, S.J.; Massaro, D. Circadian disruption alters mouse lung clock gene expression and lung mechanics. J. Appl. Physiol. 2012, 113, 385–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boysen, P.G.; Block, A.J.; Wynne, J.W.; Hunt, L.A.; Flick, M.R. Nocturnal Pulmonary Hypertension in Patients with Chronic Obstructive Pulmonary Disease. Chest 1979, 76, 536–542. [Google Scholar] [CrossRef]
- Spengler, C.; Shea, S. Endogenous Circadian Rhythm of Pulmonary Function in Healthy Humans. Am. J. Respir. Crit. Care Med. 2000, 162, 1038–1046. [Google Scholar] [CrossRef]
- Sukumaran, S.; Jusko, W.J.; Dubois, D.C.; Almon, R.R. Light-dark oscillations in the lung transcriptome: Implications for lung homeostasis, repair, metabolism, disease, and drug action. J. Appl. Physiol. 2011, 110, 1732–1747. [Google Scholar] [CrossRef] [Green Version]
- Petty, T.L. Circadian variations in chronic asthma and chronic obstructive pulmonary disease. Am. J. Med. 1988, 85, 21–23. [Google Scholar] [CrossRef]
- Tsai, C.-L.; Brenner, B.E.; Camargo, C.A., Jr. Circadian-Rhythm Differences among Emergency Department Patients with Chronic Obstructive Pulmonary Disease Exacerbation. Chrono. Int. 2007, 24, 699–713. [Google Scholar] [CrossRef]
- Agusti, A.; Hedner, J.; Marín, J.M.; Barbé, F.; Cazzola, M.; Rennard, S.; Garcia-Navarro, A.A. Night-time symptoms: A forgotten dimension of COPD. Eur. Respir. Rev. 2011, 20, 183–194. [Google Scholar] [CrossRef]
- Yao, H.; Rahman, I. Current concepts on oxidative/carbonyl stress, inflammation and epigenetics in pathogenesis of chronic obstructive pulmonary disease. Toxicol. Appl. Pharmacol. 2011, 254, 72–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, A.; Petro, W.; Konietzko, N. The circadian rhythm of ciliary beat frequency of human nasal cilia in probands with healthy lungs and in patients with chronic obstructive lung disease. Includes adrenergic stimulation by terbutaline. Pneumologie 1993, 47, 526–530. [Google Scholar]
- Gebel, S.; Gerstmayer, B.; Kuhl, P.; Borlak, J.; Meurrens, K.; Müller, T. The Kinetics of Transcriptomic Changes Induced by Cigarette Smoke in Rat Lungs Reveals a Specific Program of Defense, Inflammation, and Circadian Clock Gene Expression. Toxicol. Sci. 2006, 93, 422–431. [Google Scholar] [CrossRef]
- Casale, R.; Pasqualetti, P. Cosinor Analysis of Circadian Peak Expiratory Flow Variability in Normal Subjects, Passive Smokers, Heavy Smokers, Patients with Chronic Obstructive Pulmonary Disease and Patients with Interstitial Lung Disease. Respiration 1997, 64, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, A.; Xie, W.; Agapov, E.; Brown, S.; Steinberg, D.; Tidwell, R.; Sajol, G.; Schutz, R.; Weaver, R.; Yu, H.; et al. BMAL1 links the circadian clock to viral airway pathology and asthma phenotypes. Mucosal Immunol. 2017, 11, 97–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lechasseur, A.; Jubinville, É.; Routhier, J.; Bérubé, J.; Hamel-Auger, M.; Talbot, M.; Lamothe, J.; Aubin, S.; Paré, M.; Beaulieu, M.; et al. Exposure to electronic cigarette vapors affects pulmonary and systemic expression of circadian molecular clock genes. Physiol. Rep. 2017, 5, e13440. [Google Scholar] [CrossRef] [PubMed]
- Belden, W.J.; Dunlap, J.C. SIRT1 Is a Circadian Deacetylase for Core Clock Components. Cell 2008, 134, 212–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, S.-I.; Armstrong, C.M.; Kaeberlein, M.; Guarente, L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature 2000, 403, 795–800. [Google Scholar] [CrossRef]
- Grimaldi, B.; Nakahata, Y.; Kaluzova, M.; Masubuchi, S.; Sassone-Corsi, P. Chromatin remodeling, metabolism and circadian clocks: The interplay of CLOCK and SIRT1. Int. J. Biochem. Cell Boil. 2009, 41, 81–86. [Google Scholar] [CrossRef]
- Rajendrasozhan, S.; Yang, S.-R.; Kinnula, V.L.; Rahman, I. SIRT1, an Antiinflammatory and Antiaging Protein, Is Decreased in Lungs of Patients with Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2008, 177, 861–870. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Chung, S.; Hwang, J.-W.; Rajendrasozhan, S.; Sundar, I.K.; Dean, D.A.; McBurney, M.W.; Guarente, L.; Gu, W.; Rönty, M.; et al. SIRT1 protects against emphysema via FOXO3-mediated reduction of premature senescence in mice. J. Clin. Investig. 2012, 122, 2032–2045. [Google Scholar] [CrossRef]
- Yang, S.-R.; Wright, J.; Bauter, M.; Seweryniak, K.; Kode, A.; Rahman, I. Sirtuin regulates cigarette smoke-induced proinflammatory mediator release via RelA/p65 NF-κB in macrophages in vitro and in rat lungs in vivo: Implications for chronic inflammation and aging. Am. J. Physiol. Cell. Mol. Physiol. 2007, 292, L567–L576. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Dutta, R.K.; Chinnapaiyan, S.; Unwalla, H. Aberrant MicroRNAomics in Pulmonary Complications: Implications in Lung Health and Diseases. Mol. Ther. Nucleic Acids 2019, 18, 413–431. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Boil. 2018, 20, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Haase, A.D.; Jaskiewicz, L.; Zhang, H.; Lainé, S.; Sack, R.; Gatignol, A.; Filipowicz, W. TRBP, a regulator of cellular PKR and HIV-1 virus expression, interacts with Dicer and functions in RNA silencing. EMBO Rep. 2005, 6, 961–967. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Wahid, F.; Shehzad, A.; Khan, T.; Kim, Y.Y. MicroRNAs: Synthesis, mechanism, function, and recent clinical trials. Biochim. et Biophys. Acta BBA Bioenerg. 2010, 1803, 1231–1243. [Google Scholar] [CrossRef] [Green Version]
- Macfarlane, L.-A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [Green Version]
- Kim, V.N.; Han, J.; Siomi, M.C. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Boil. 2009, 10, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Kawamata, T.; Tomari, Y. Making RISC. Trends Biochem. Sci. 2010, 35, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Khraiwesh, B.; Arif, M.A.; Seumel, G.I.; Ossowski, S.; Weigel, D.; Reski, R.; Frank, W. Transcriptional Control of Gene Expression by MicroRNAs. Cell 2010, 140, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Shende, V.R.; Neuendorff, N.; Earnest, D.J. Role of miR-142-3p in the Post-Transcriptional Regulation of the Clock Gene Bmal1 in the Mouse SCN. PLoS ONE 2013, 8, e65300. [Google Scholar] [CrossRef] [PubMed]
- Shende, V.R.; Goldrick, M.M.; Ramani, S.; Earnest, D.J. Expression and Rhythmic Modulation of Circulating MicroRNAs Targeting the Clock Gene Bmal1 in Mice. PLoS ONE 2011, 6, e22586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olena, A.F.; Patton, J.G. Genomic organization of microRNAs. J. Cell. Physiol. 2010, 222, 540–545. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.K.; Kim, V.N. Processing of intronic microRNAs. EMBO J. 2007, 26, 775–783. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.-L.; Miller, J.D.; Ying, S.-Y. Intronic MicroRNA (miRNA). J. Biomed. Biotechnol. 2006, 2006, 1–13. [Google Scholar] [CrossRef]
- Ramalingam, P.; Palanichamy, J.K.; Singh, A.; Das, P.; Bhagat, M.; Kassab, M.A.; Sinha, S.; Chattopadhyay, P. Biogenesis of intronic miRNAs located in clusters by independent transcription and alternative splicing. RNA 2013, 20, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Nagel, R.; Clijsters, L.; Agami, R. The miRNA-192/194 cluster regulates the Period gene family and the circadian clock. FEBS J. 2009, 276, 5447–5455. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.-H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Hagedorn, C.H.; Cullen, B.R. Human microRNAs are processed from capped, polyadenylated transcripts that can also function as mRNAs. RNA 2004, 10, 1957–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzoccoli, G.; Colangelo, T.; Panza, A.; Rubino, R.; Tiberio, C.; Palumbo, O.; Carella, M.; Trombetta, D.; Gentile, A.; Tavano, F.; et al. Analysis of clock gene-miRNA correlation networks reveals candidate drivers in colorectal cancer. Oncotarget 2016, 7, 45444–45461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasakova, K.; Reis, R.; Vician, M.; Zeman, M.; Herichova, I. Expression of miR-34a-5p is up-regulated in human colorectal cancer and correlates with survival and clock gene PER2 expression. PLoS ONE 2019, 14, e0224396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borchert, G.M.; Lanier, W.; Davidson, B.L. RNA polymerase III transcribes human microRNAs. Nat. Struct. Mol. Boil. 2006, 13, 1097–1101. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; D’Alessandro, M.; Lee, C. miRNAs are required for generating a time delay critical for the circadian oscillator. Curr. Boil. 2013, 23, 1959–1968. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, J.S.; Hong, H.-K.; Ko, C.H.; McDearmon, E.L. The genetics of mammalian circadian order and disorder: Implications for physiology and disease. Nat. Rev. Genet. 2008, 9, 764–775. [Google Scholar] [CrossRef]
- Ju, J.; Wu, S.; Fesler, A. Implications of Circadian Rhythm Regulation by microRNAs in Colorectal Cancer. Cancer Transl. Med. 2016, 2, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Esau, C.; Davis, S.; Murray, S.F.; Yu, X.X.; Pandey, S.K.; Pear, M.; Watts, L.; Booten, S.L.; Graham, M.; McKay, R.; et al. miR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab. 2006, 3, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Gatfield, D.; Le Martelot, G.; Vejnar, C.E.; Gerlach, D.; Schaad, O.; Fleury-Olela, F.; Ruskeepää, A.-L.; Orešič, M.; Esau, C.C.; Zdobnov, E.M.; et al. Integration of microRNA miR-122 in hepatic circadian gene expression. Genome Res. 2009, 23, 1313–1326. [Google Scholar] [CrossRef] [Green Version]
- Shilts, J.; Chen, G.; Hughey, J.J. Evidence for widespread dysregulation of circadian clock progression in human cancer. PeerJ 2018, 6, e4327. [Google Scholar] [CrossRef] [PubMed]
- Na, Y.-J.; Sung, J.H.; Lee, S.C.; Lee, Y.-J.; Choi, Y.J.; Park, W.-Y.; Shin, H.S.; Kim, J.H. Comprehensive analysis of microRNA-mRNA co-expression in circadian rhythm. Exp. Mol. Med. 2009, 41, 638–647. [Google Scholar] [CrossRef]
- Guo, X.; Zhu, Y.; Hong, X.; Zhang, M.; Qiu, X.; Wang, Z.; Qi, Z.; Hong, X. miR-181d and c-myc-mediated inhibition of CRY2 and FBXL3 reprograms metabolism in colorectal cancer. Cell Death Dis. 2017, 8, e2958. [Google Scholar] [CrossRef]
- Wood, P.A.; Yang, X.; Hrushesky, W. Clock Genes and Cancer. Integr. Cancer Ther. 2009, 8, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Aubert, J.-D.; Dalal, B.I.; Bai, T.R.; Roberts, C.R.; Hayashi, S.; Hogg, J.C. Transforming growth factor beta 1 gene expression in human airways. Thorax 1994, 49, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Vignola, A.M.; Chanez, P.; Chiappara, G.; Merendino, A.; Zinnanti, E.; Bousquet, J.; Bellia, V.; Bonsignore, G. Release of transforming growth factor-beta (TGF-β) and fibronectin by alveolar macrophages in airway diseases. Clin. Exp. Immunol. 1996, 106, 114–119. [Google Scholar] [CrossRef]
- Vignola, A.M.; Chanez, P.; Chiappara, G.; Merendino, A.; Pace, E.; Rizzo, A.; La Rocca, A.M.; Bellia, V.; Bonsignore, G.; Bousquet, J. Transforming Growth Factor- β Expression in Mucosal Biopsies in Asthma and Chronic Bronchitis. Am. J. Respir. Crit. Care Med. 1997, 156, 591–599. [Google Scholar] [CrossRef]
- Takizawa, H.; Tanaka, M.; Takami, K.; Ohtoshi, T.; Ito, K.; Satoh, M.; Okada, Y.; Yamasawa, F.; Nakahara, K.; Umeda, A. Increased Expression of Transforming Growth Factor- β 1 in Small Airway Epithelium from Tobacco Smokers and Patients with Chronic Obstructive Pulmonary Disease (COPD). Am. J. Respir. Crit. Care Med. 2001, 163, 1476–1483. [Google Scholar] [CrossRef] [PubMed]
- Gast, H.; Gordic, S.; Petrzilka, S.; Lopez, M.; Müller, A.; Gietl, A.; Hock, C.; Birchler, T.; Fontana, A. Transforming growth factor-beta inhibits the expression of clock genes. Ann. New York Acad. Sci. 2012, 1261, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Gongora, R.; Sosulski, M.L.; Luo, F.; Sanchez, C.G. Regulation of transforming growth factor-beta1 (TGF-β1)-induced pro-fibrotic activities by circadian clock gene BMAL1. Respir. Res. 2016, 17, 4. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.-D.; Yeh, J.-K.; Peng, M.-T.; Shie, S.-S.; Lin, S.-L.; Yang, C.-H.; Chen, T.-H.; Hung, K.-C.; Wang, C.-C.; Hsieh, I.-C.; et al. Circadian CLOCK Mediates Activation of Transforming Growth Factor-β Signaling and Renal Fibrosis through Cyclooxygenase 2. Am. J. Pathol. 2015, 185, 3152–3163. [Google Scholar] [CrossRef]
- Kheir, T.B.; Futoma-Kazmierczak, E.; Skanderup, A.J.; Krogh, A.; Bardram, L.; Hother, C.; Grønbæk, K.; Federspiel, B.; Lund, A.H.; Friis-Hansen, L. miR-449 inhibits cell proliferation and is down-regulated in gastric cancer. Mol. Cancer 2011, 10, 29. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Zhao, Y.; Sun, J.; Mu, P.; Deng, Y. miR449a/SIRT1/PGC-1α Is Necessary for Mitochondrial Biogenesis Induced by T-2 Toxin. Front. Pharmacol. 2018, 8. [Google Scholar] [CrossRef]
- Xu, J.-Q.; Liu, P.; Si, M.-J.; Ding, X.-Y. MicroRNA-126 inhibits osteosarcoma cells proliferation by targeting Sirt1. Tumor Boil. 2013, 34, 3871–3877. [Google Scholar] [CrossRef]
- Yamakuchi, M.; Lowenstein, C.J. MiR-34, SIRT1, and p53: The feedback loop. Cell Cycle 2009, 8, 712–715. [Google Scholar] [CrossRef]
- Zhang, H.-S.; Chen, X.-Y.; Wu, T.-C.; Sang, W.-W.; Ruan, Z. MiR-34a is involved in Tat-induced HIV-1 long terminal repeat (LTR) transactivation through the SIRT1/NFκB pathway. FEBS Lett. 2012, 586, 4203–4207. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zhu, X.; Cheng, S.; Xie, Y.; Wang, Z.; Liu, Y.; Jiang, Z.; Xiao, J.; Guo, H. MiR-29a/b/c regulate human circadian gene hPER1 expression by targeting its 3’UTR. Acta Biochim. et Biophys. Sin. 2014, 46, 313–317. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, C.; Goedeke, L.; Rotllan, N.; Yoon, J.-H.; Cirera-Salinas, D.; Mattison, J.A.; Suárez, Y.; De Cabo, R.; Gorospe, M.; Fernández-Hernando, C. MicroRNA 33 Regulates Glucose Metabolism. Mol. Cell. Boil. 2013, 33, 2891–2902. [Google Scholar] [CrossRef] [Green Version]
- Devanna, P.; Vernes, S.C. A direct molecular link between the autism candidate gene RORa and the schizophrenia candidate MIR137. Sci. Rep. 2014, 4, 3994. [Google Scholar] [CrossRef] [Green Version]
- Chinnapaiyan, S.; Parira, T.; Dutta, R.K.; Agudelo, M.; Morris, A.; Nair, M.; Unwalla, H. HIV Infects Bronchial Epithelium and Suppresses Components of the Mucociliary Clearance Apparatus. PLoS ONE 2017, 12, e0169161. [Google Scholar] [CrossRef] [Green Version]
- Unwalla, H.; Ivonnet, P.; Dennis, J.S.; Conner, G.E.; Salathe, M. Transforming Growth Factor-β1 and Cigarette Smoke Inhibit the Ability of β2-Agonists to Enhance Epithelial Permeability. Am. J. Respir. Cell Mol. Boil. 2015, 52, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, J.; Acuña, S.M.; Aoki, J.I.; Floeter-Winter, L.M.; Muxel, S.M. Long Non-Coding RNAs in the Regulation of Gene Expression: Physiology and Disease. Noncoding RNA 2019, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Coon, S.L.; Munson, P.J.; Cherukuri, P.F.; Sugden, D.; Rath, M.F.; Møller, M.; Clokie, S.; Fu, C.; Olanich, M.E.; Rangel, Z.; et al. Circadian changes in long noncoding RNAs in the pineal gland. Proc. Natl. Acad. Sci. USA 2012, 109, 13319–13324. [Google Scholar] [CrossRef] [Green Version]
- Vollmers, C.; Schmitz, R.J.; Nathanson, J.; Yeo, G.; Ecker, J.R.; Panda, S. Circadian oscillations of protein-coding and regulatory RNAs in a highly dynamic mammalian liver epigenome. Cell Metab. 2012, 16, 833–845. [Google Scholar] [CrossRef] [Green Version]
- Palczewski, K.; Kevany, B.M.; Genoud, C.; Bai, X.; Palczewski, K. Photoreceptor phagocytosis is mediated by phosphoinositide signaling. FASEB J. 2013, 27, 4585–4595. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Belden, W.J. Long non-coding RNAs have age-dependent diurnal expression that coincides with age-related changes in genome-wide facultative heterochromatin. BMC Genom. 2018, 19, 777. [Google Scholar] [CrossRef]
- Park, J.; Zhu, Q.; Mirek, E.; Na, L.; Raduwan, H.; Anthony, T.G.; Belden, W.J. BMAL1 associates with chromosome ends to control rhythms in TERRA and telomeric heterochromatin. PLoS ONE 2019, 14, e0223803. [Google Scholar] [CrossRef] [Green Version]
- Cui, M.; Zheng, M.; Sun, B.; Wang, Y.; Ye, L.; Zhang, X. A long noncoding RNA perturbs the circadian rhythm of hepatoma cells to facilitate hepatocarcinogenesis. Neoplasia 2015, 17, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Zhao, X.; Wu, X.; Xiang, L.; Yuan, Y.; Zhou, S.; Yu, W. LncRNA UCA1 facilitated cell growth and invasion through the miR-206/CLOCK axis in glioma. Cancer Cell Int. 2019, 19, 316–319. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, H.; Xu, C.; Yu, C.; Li, Y. Long Non-Coding RNA Profiling in a Non-Alcoholic Fatty Liver Disease Rodent Model: New Insight into Pathogenesis. Int. J. Mol. Sci. 2017, 18, 21. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Chen, L.; Zeb, F.; Huang, Y.; An, J.; Ren, J.; Yang, F.; Feng, Q. Regulation of circadian rhythms by NEAT1 mediated TMAO-induced endothelial proliferation: A protective role of asparagus extract. Exp. Cell Res. 2019, 382, 111451. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wu, X.; Zeb, F.; Huang, Y.; An, J.; Jiang, P.; Chen, A.; Xu, C.; Feng, Q. Acrolein-induced apoptosis of smooth muscle cells through NEAT1-Bmal1/Clock pathway and a protection from asparagus extract. Environ. Pollut. 2020, 258, 113735. [Google Scholar] [CrossRef] [PubMed]
- Bratkovič, T.; Rogelj, B. The many faces of small nucleolar RNAs. Biochim. et Biophys. Acta BBA Bioenerg. 2014, 1839, 438–443. [Google Scholar] [CrossRef]
- Dupuis-Sandoval, F.; Poirier, M.; Scott, M.S. The emerging landscape of small nucleolar RNAs in cell biology. Wiley Interdiscip. Rev. RNA 2015, 6, 381–397. [Google Scholar] [CrossRef]
- Jorjani, H.; Kehr, S.; Jedlinski, D.J.; Gumienny, R.; Hertel, J.; Stadler, P.F.; Zavolan, M.; Gruber, A. An updated human snoRNAome. Nucleic Acids Res. 2016, 44, 5068–5082. [Google Scholar] [CrossRef]
- Massenet, S.; Bertrand, E.; Verheggen, C. Assembly and trafficking of box C/D and H/ACA snoRNPs. RNA Boil. 2016, 14, 680–692. [Google Scholar] [CrossRef] [Green Version]
- Aitken, S.; Semple, C.A. The circadian dynamics of small nucleolar RNA in the mouse liver. J. R. Soc. Interface 2017, 14, 20170034. [Google Scholar] [CrossRef] [Green Version]
- Powell, W.; Coulson, R.; Crary, F.K.; Wong, S.S.; Ach, R.A.; Tsang, P.; Yamada, N.A.; Yasui, D.H.; LaSalle, J. A Prader-Willi locus lncRNA cloud modulates diurnal genes and energy expenditure. Hum. Mol. Genet. 2013, 22, 4318–4328. [Google Scholar] [CrossRef] [Green Version]
- Coulson, R.; Yasui, D.H.; Dunaway, K.; Laufer, B.; Ciernia, A.V.; Zhu, Y.; Mordaunt, C.; Totah, T.S.; LaSalle, J. Snord116-dependent diurnal rhythm of DNA methylation in mouse cortex. Nat. Commun. 2018, 9, 1616. [Google Scholar] [CrossRef] [Green Version]
- Mei, Y.-P.; Liao, J.-P.; Shen, J.; Yu, L.; Liu, B.; Liu, L.; Li, R.-Y.; Ji, L.; Dorsey, S.G.; Jiang, Z.-R.; et al. Small nucleolar RNA 42 acts as an oncogene in lung tumorigenesis. Oncogene 2011, 31, 2794–2804. [Google Scholar] [CrossRef]
- Mannoor, K.; Shen, J.; Liao, J.; Liu, Z.; Jiang, F. Small nucleolar RNA signatures of lung tumor-initiating cells. Mol. Cancer 2014, 13, 104. [Google Scholar] [CrossRef] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chinnapaiyan, S.; Dutta, R.K.; Devadoss, D.; Chand, H.S.; Rahman, I.; Unwalla, H.J. Role of Non-Coding RNAs in Lung Circadian Clock Related Diseases. Int. J. Mol. Sci. 2020, 21, 3013. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21083013
Chinnapaiyan S, Dutta RK, Devadoss D, Chand HS, Rahman I, Unwalla HJ. Role of Non-Coding RNAs in Lung Circadian Clock Related Diseases. International Journal of Molecular Sciences. 2020; 21(8):3013. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21083013
Chicago/Turabian StyleChinnapaiyan, Srinivasan, Rajib Kumar Dutta, Dinesh Devadoss, Hitendra S Chand, Irfan Rahman, and Hoshang Jehangir Unwalla. 2020. "Role of Non-Coding RNAs in Lung Circadian Clock Related Diseases" International Journal of Molecular Sciences 21, no. 8: 3013. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21083013