Peroxisome Proliferator-Activated Receptor Gamma Modulator Promotes Neonatal Mouse Primordial Follicle Activation In Vitro

 and
and
Abstract
:1. Introduction
2. Results
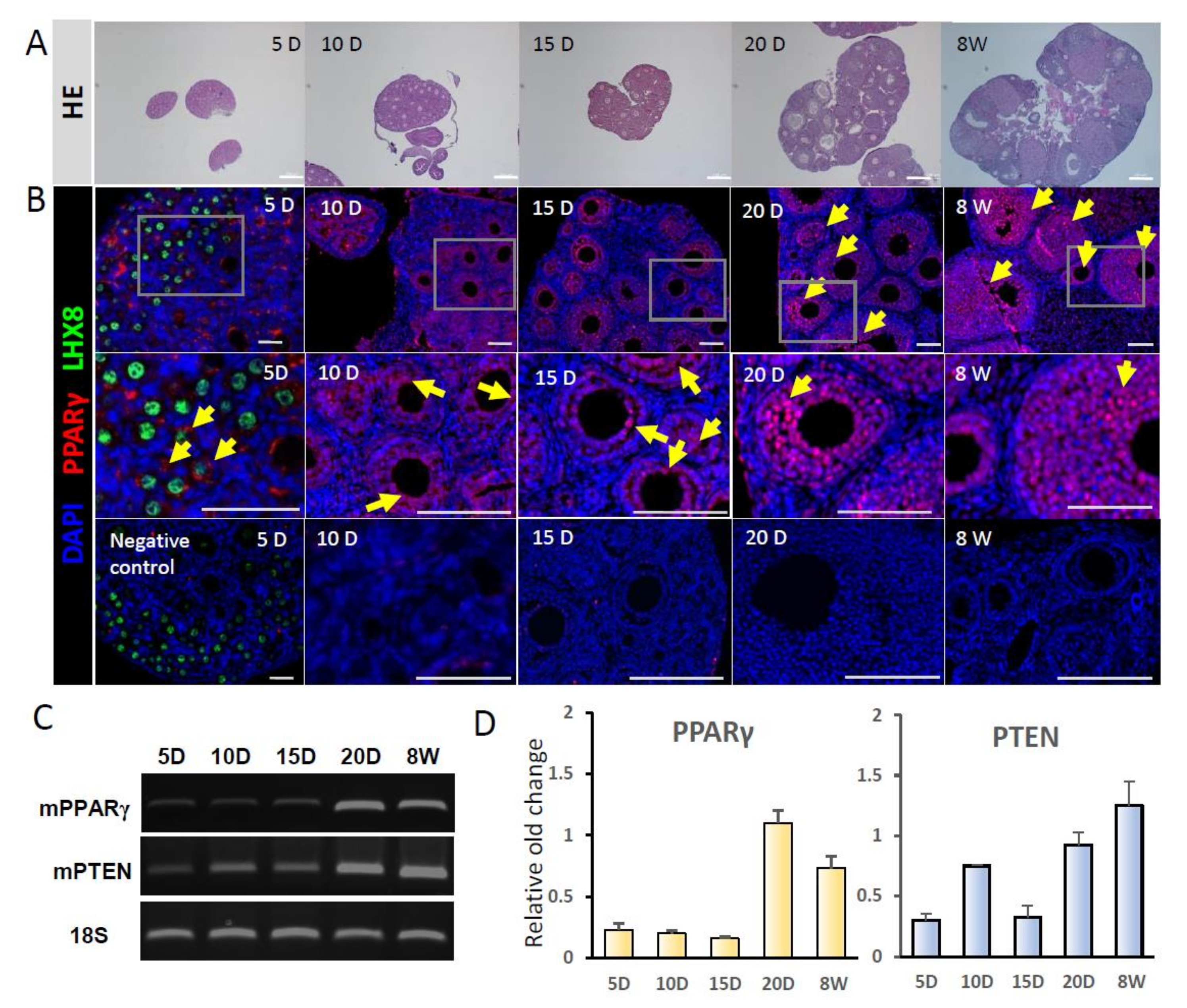
2.1. Gene Expression of PPARγ and PTEN in Mouse Ovaries
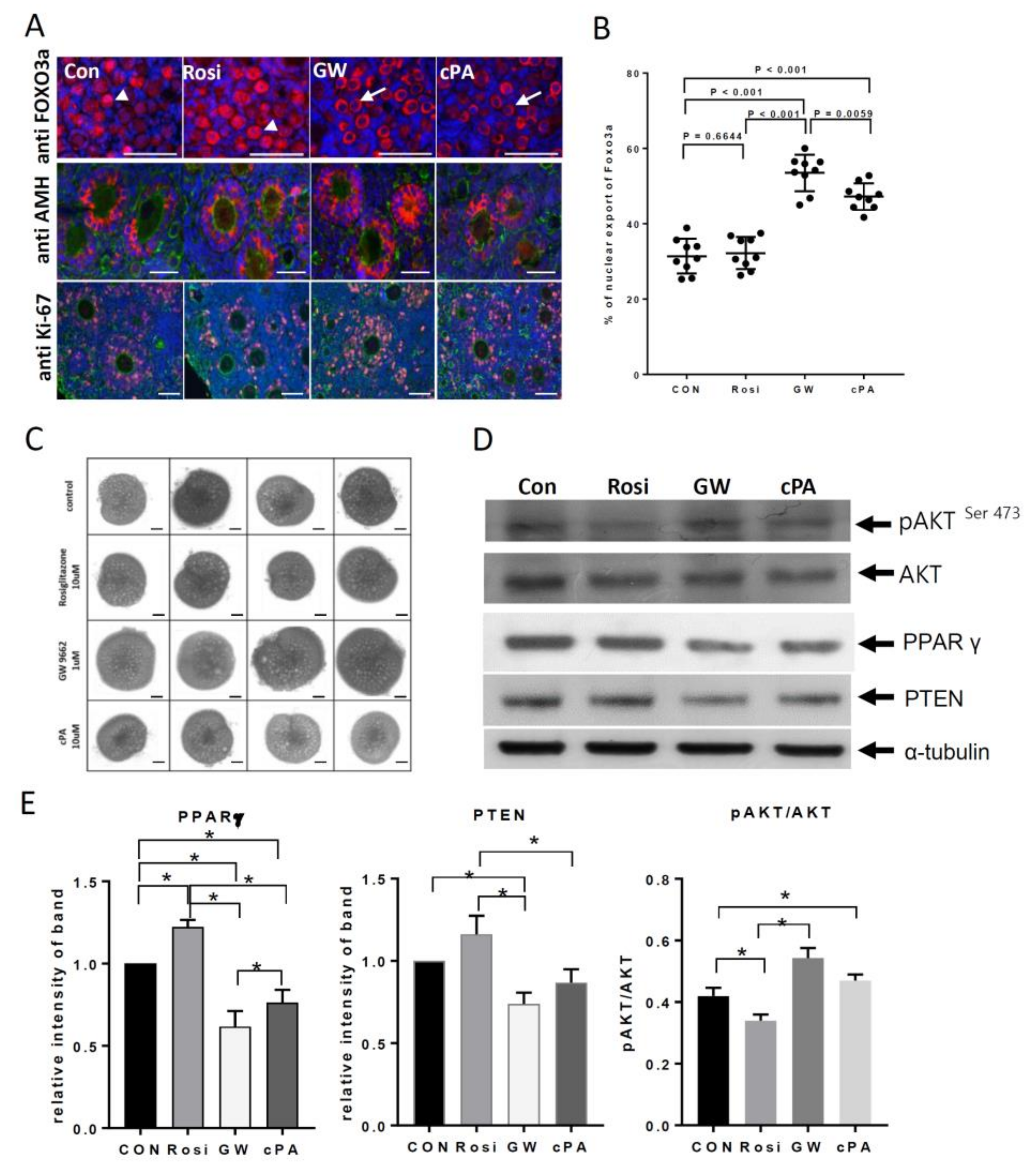
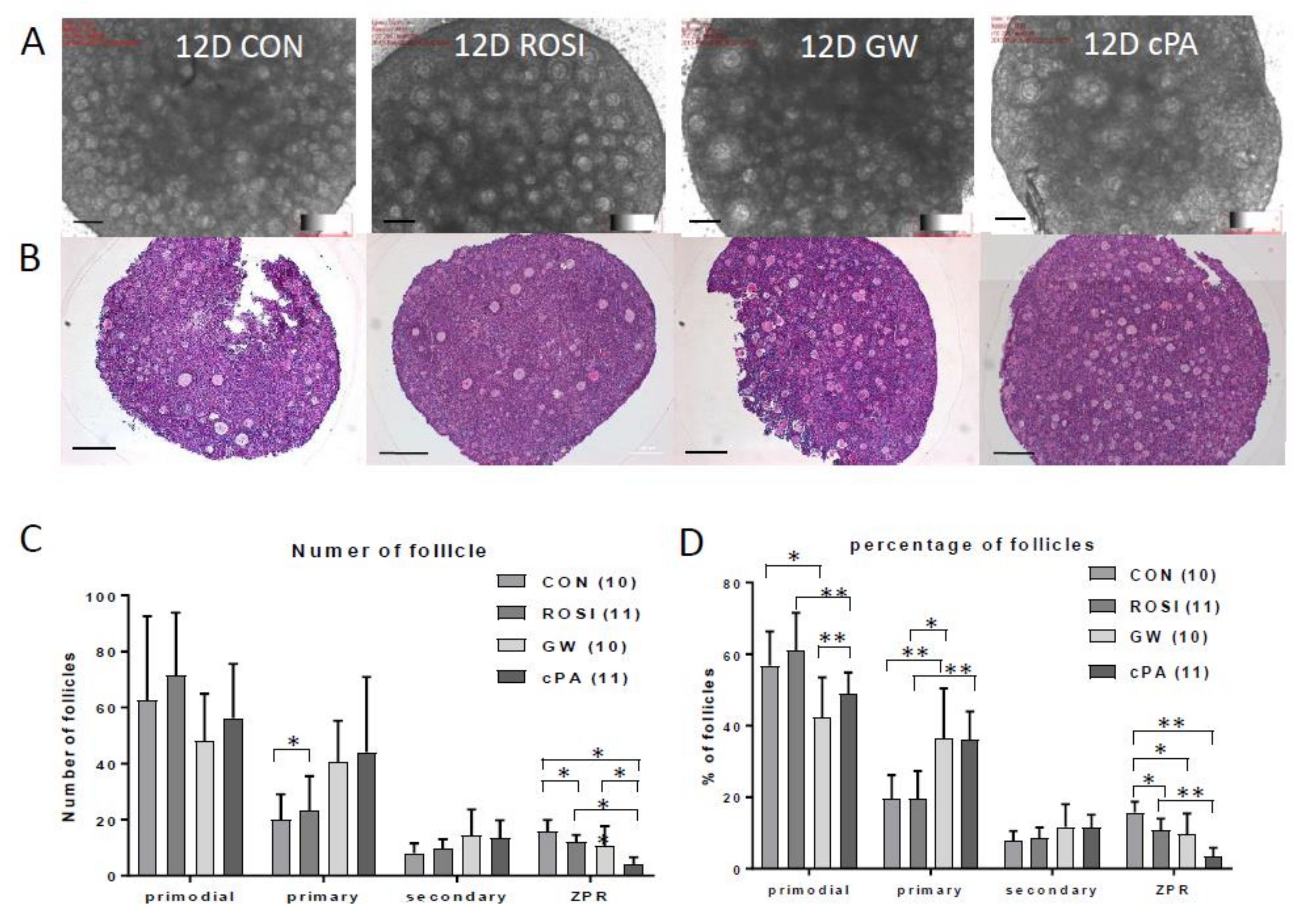
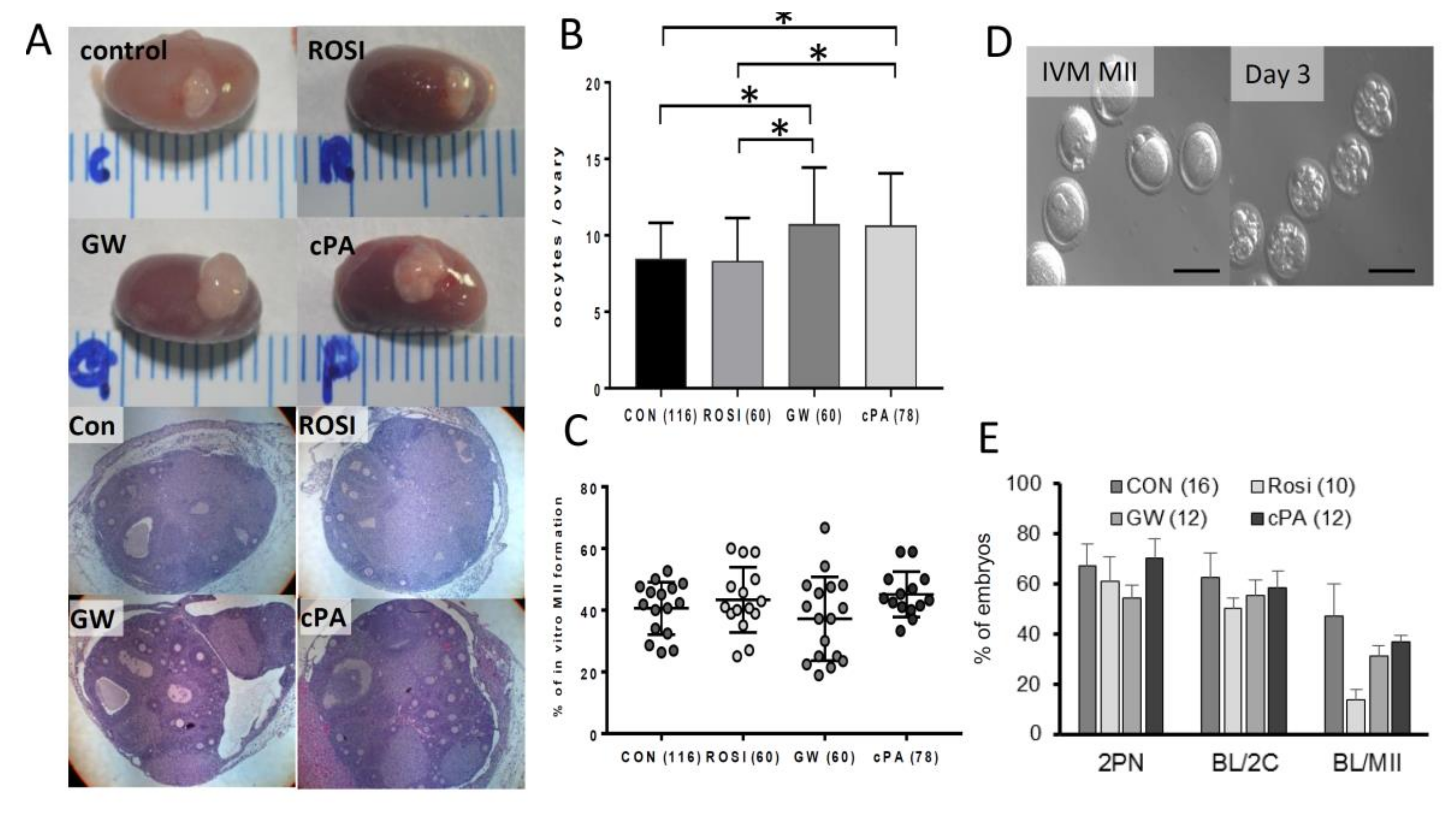
2.2. Effects on Primordial Follicle Activation and Development by PPARγ Modulators In Vitro
2.3. Embryonic Development of Oocytes from Transplanted Ovaries
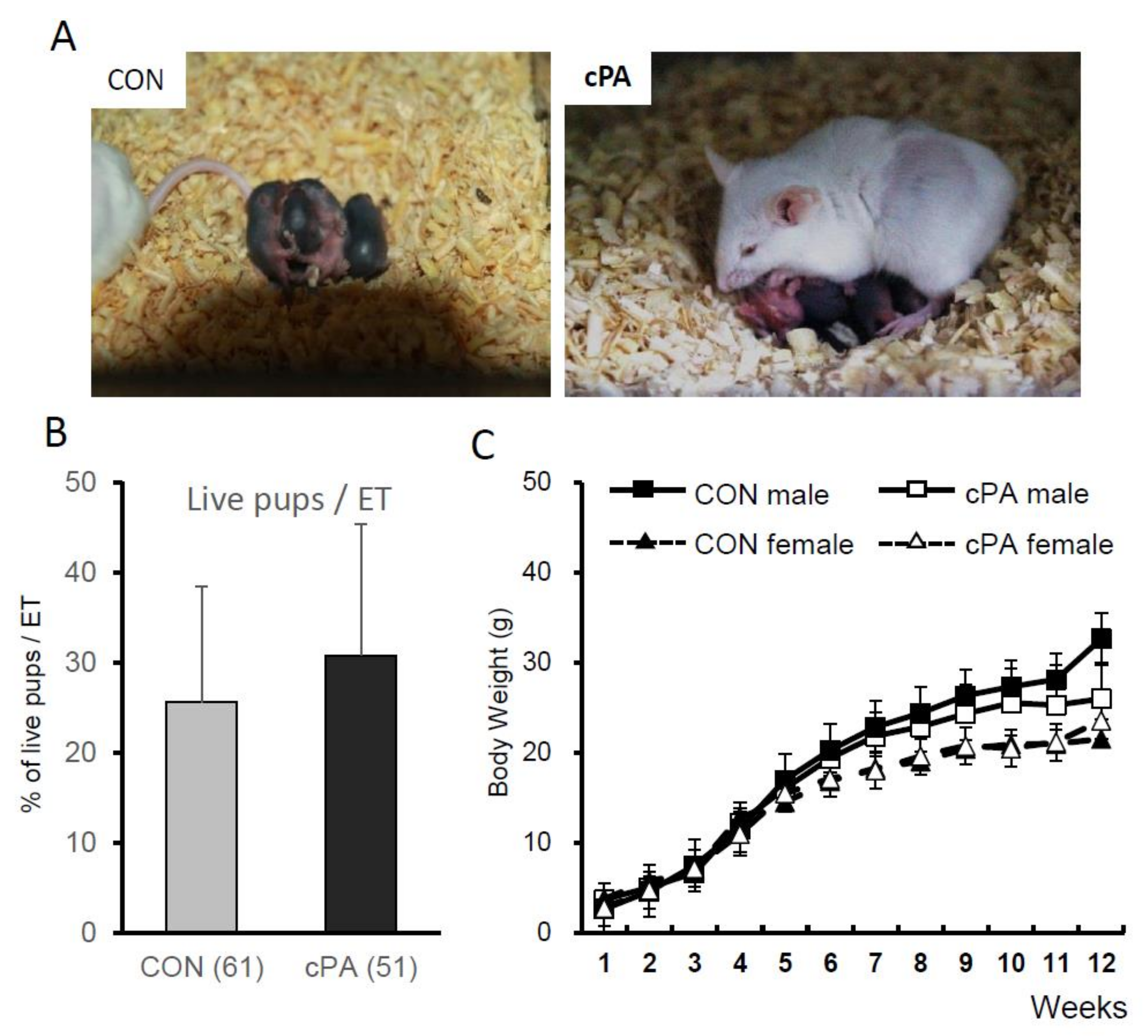
2.4. Birth of Live Pups after Embryo Transfer into Recipient Mice
3. Discussion
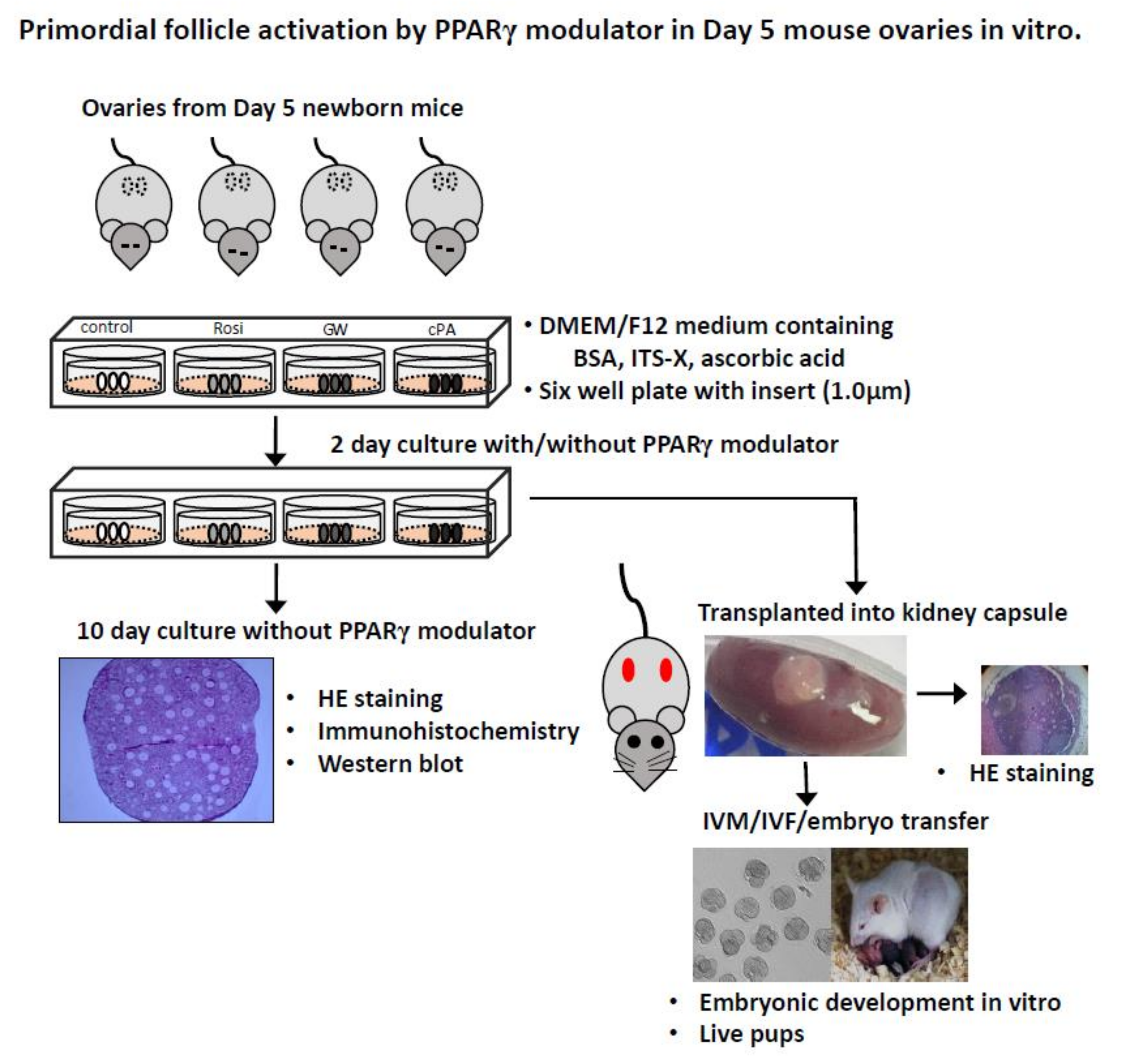
4. Materials and Methods
4.1. Animals
4.2. Gene Expression of PPARγ and PTEN in Mouse Ovaries
4.3. In Vitro Culture of Neonatal Mouse Ovaries with PPARγ Modulators
4.4. Ovarian Transplantation and In Vivo Culture
4.5. In Vitro Maturation (IVM) and Embryonic Development of Oocytes from Transplanted Ovaries
4.6. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- McGee, E.A.; Hsueh, A.J. Initial and cyclic recruitment of ovarian follicles. Endocr. Rev. 2000, 21, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, D.; Flohr, G.; Gorre, N.; Shen, Y.; Yang, H.; Lundin, E.; Lan, Z.; Gambello, M.J.; Liu, K. Disruption of Tsc2 in oocytes leads to overactivation of the entire pool of primordial follicles. Mol. Hum. Reprod. 2009, 15, 765–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, M.K. Regulation of primordial follicle assembly and development. Hum. Reprod. Update 2005, 11, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.; Zhang, X.; Mu, Y.; Li, Y.; Hao, J. Effects of BMP4/SMAD signaling pathway on mouse primordial follicle growth and survival via up-regulation of Sohlh2 and c-kit. Mol. Reprod. Dev. 2013, 80, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Kedem, A.; Fisch, B.; Garor, R.; Ben-Zaken, A.; Gizunterman, T.; Felz, C.; Ben-Haroush, A.; Kravarusic, D.; Abir, R. Growth differentiating factor 9 (GDF9) and bone morphogenetic protein 15 both activate development of human primordial follicles in vitro, with seemingly more beneficial effects of GDF9. J. Clin. Endocrinol. Metab. 2011, 96, E1246–E1254. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, E.E.; Kezele, P.; Skinner, M.K. Leukemia inhibitory factor (LIF) promotes the primordial to primary follicle transition in rat ovaries. Mol. Cell Endocrinol. 2002, 188, 65–73. [Google Scholar] [CrossRef]
- Makker, A.; Goel, M.M.; Mahdi, A.A. PI3K/PTEN/Akt and TSC/mTOR signaling pathways, ovarian dysfunction, and infertility: An update. J. Mol. Endocrinol. 2014, 53, R103–R118. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, D.; Liu, K. Molecular mechanisms underlying the activation of mammalian primordial follicles. Endocr. Rev. 2009, 30, 438–464. [Google Scholar]
- John, G.B.; Gallardo, T.D.; Shirley, L.J.; Castrillon, D.H. Foxo3 is a PI3K-dependent molecular switch controlling the initiation of oocyte growth. Dev. Biol. 2008, 321, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.; Liu, L.; Adhikari, D.; Jagarlamudi, K.; Rajareddy, S.; Shen, Y.; Du, C.; Tang, W.; Hamalainen, T.; Peng, S.L.; et al. Oocyte-specific deletion of Pten causes premature activation of the primordial follicle pool. Science 2008, 319, 611–613. [Google Scholar] [CrossRef] [Green Version]
- Castrillon, D.H.; Miao, L.; Kollipara, R.; Horner, J.W.; DePinho, R.A. Suppression of ovarian follicle activation in mice by the transcription factor Foxo3a. Science 2003, 301, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, D.; Gorre, N.; Risal, S.; Zhao, Z.; Zhang, H.; Shen, Y.; Liu, K. The safe use of a PTEN inhibitor for the activation of dormant mouse primordial follicles and generation of fertilizable eggs. PLoS ONE 2012, 7, e39034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.; Nagaraju, G.; Liu, Z.; Liu, K. Functional roles of the phosphatidylinositol 3-kinases (PI3Ks) signaling in the mammalian ovary. Mol. Cell Endocrinol. 2012, 356, 24–30. [Google Scholar] [CrossRef]
- Novella-Maestre, E.; Herraiz, S.; Rodriguez-Iglesias, B.; Diaz-Garcia, C.; Pellicer, A. Short-Term PTEN Inhibition Improves In Vitro Activation of Primordial Follicles, Preserves Follicular Viability, and Restores AMH Levels in Cryopreserved Ovarian Tissue From Cancer Patients. PLoS ONE 2015, 10, e0127786. [Google Scholar] [CrossRef]
- Li, J.; Kawamura, K.; Cheng, Y.; Liu, S.; Klein, C.; Liu, S.; Duan, E.K.; Hsueh, A.J. Activation of dormant ovarian follicles to generate mature eggs. Proc. Natl. Acad. Sci. USA 2010, 107, 10280–10284. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, M.; Kinnell, H.L.; Anderson, R.A.; Telfer, E.E. Inhibition of phosphatase and tensin homologue (PTEN) in human ovary in vitro results in increased activation of primordial follicles but compromises development of growing follicles. Mol. Hum. Reprod. 2014, 20, 736–744. [Google Scholar] [CrossRef] [Green Version]
- Semple, R.K.; Chatterjee, V.K.; O’Rahilly, S. PPAR gamma and human metabolic disease. J. Clin. Investig. 2006, 116, 581–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minge, C.E.; Robker, R.L.; Norman, R.J. PPAR Gamma: Coordinating Metabolic and Immune Contributions to Female Fertility. PPAR Res. 2008, 2008, 243791. [Google Scholar] [CrossRef] [PubMed]
- Froment, P.; Gizard, F.; Staels, B.; Dupont, J.; Monget, P. A role of PPARgamma in reproduction? Med. Sci. (Paris) 2005, 21, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Patel, L.; Pass, I.; Coxon, P.; Downes, C.P.; Smith, S.A.; Macphee, C.H. Tumor suppressor and anti-inflammatory actions of PPARgamma agonists are mediated via upregulation of PTEN. Curr. Biol. 2001, 11, 764–768. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.S.; Park, S.J.; Hwang, P.H.; Yi, H.K.; Song, C.H.; Chai, O.H.; Kim, J.S.; Lee, M.K.; Lee, Y.C. PPAR-gamma modulates allergic inflammation through up-regulation of PTEN. FASEB J. 2005, 19, 1033–1035. [Google Scholar] [CrossRef] [PubMed]
- Tsukahara, T. PPAR gamma Networks in Cell Signaling: Update and Impact of Cyclic Phosphatidic Acid. J. Lipids 2013, 2013, 246597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, Y. Cyclic phosphatidic acid—A unique bioactive phospholipid. Biochim. Biophys. Acta 2008, 1781, 519–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukahara, T.; Tsukahara, R.; Fujiwara, Y.; Yue, J.; Cheng, Y.; Guo, H.; Bolen, A.; Zhang, C.; Balazs, L.; Re, F.; et al. Phospholipase D2-dependent inhibition of the nuclear hormone receptor PPARgamma by cyclic phosphatidic acid. Mol. Cell 2010, 39, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Tsukahara, T.; Hanazawa, S.; Kobayashi, T.; Iwamoto, Y.; Murakami-Murofushi, K. Cyclic phosphatidic acid decreases proliferation and survival of colon cancer cells by inhibiting peroxisome proliferator-activated receptor gamma. Prostaglandins Other Lipid Mediat. 2010, 93, 126–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukahara, T.; Murakami-Murofushi, K. Release of cyclic phosphatidic acid from gelatin-based hydrogels inhibit colon cancer cell growth and migration. Sci. Rep. 2012, 2, 687. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Hur, G.Y.; Jung, K.H.; Jung, H.C.; Lee, S.Y.; Kim, J.H.; Shin, C.; Shim, J.J.; In, K.H.; Kang, K.H.; et al. PPAR-gamma agonist increase gefitinib’s antitumor activity through PTEN expression. Lung Cancer 2006, 51, 297–301. [Google Scholar] [CrossRef]
- Goetze, S.; Eilers, F.; Bungenstock, A.; Kintscher, U.; Stawowy, P.; Blaschke, F.; Graf, K.; Law, R.E.; Fleck, E.; Grafe, M. PPAR activators inhibit endothelial cell migration by targeting Akt. Biochem. Biophys. Res. Commun. 2002, 293, 1431–1437. [Google Scholar] [CrossRef]
- Komar, C.M. Peroxisome proliferator-activated receptors (PPARs) and ovarian function—implications for regulating steroidogenesis, differentiation, and tissue remodeling. Reprod. Biol. Endocrinol. 2005, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Froment, P.; Gizard, F.; Defever, D.; Staels, B.; Dupont, J.; Monget, P. Peroxisome proliferator-activated receptors in reproductive tissues: From gametogenesis to parturition. J. Endocrinol. 2006, 189, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Komar, C.M.; Braissant, O.; Wahli, W.; Curry, T.E., Jr. Expression and localization of PPARs in the rat ovary during follicular development and the periovulatory period. Endocrinology 2001, 142, 4831–4838. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Miyoshi, K.; Claudio, E.; Siebenlist, U.K.; Gonzalez, F.J.; Flaws, J.; Wagner, K.U.; Hennighausen, L. Loss of the peroxisome proliferation-activated receptor gamma (PPARgamma) does not affect mammary development and propensity for tumor formation but leads to reduced fertility. J. Biol. Chem. 2002, 277, 17830–17835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, B.H.; Baek, K.H. Pro12Ala and His447His polymorphisms of PPAR-gamma are associated with polycystic ovary syndrome. Reprod. Biomed. Online 2009, 18, 644–650. [Google Scholar] [CrossRef]
- Thangavelu, M.; Godla, U.R.; Paul Solomon, F.D.; Maddaly, R. Single-nucleotide polymorphism of INS, INSR, IRS1, IRS2, PPAR-G and CAPN10 genes in the pathogenesis of polycystic ovary syndrome. J. Genet. 2017, 96, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Savage, D.B.; Tan, G.D.; Acerini, C.L.; Jebb, S.A.; Agostini, M.; Gurnell, M.; Williams, R.L.; Umpleby, A.M.; Thomas, E.L.; Bell, J.D.; et al. Human metabolic syndrome resulting from dominant-negative mutations in the nuclear receptor peroxisome proliferator-activated receptor-gamma. Diabetes 2003, 52, 910–917. [Google Scholar] [CrossRef] [Green Version]
- Barroso, I.; Gurnell, M.; Crowley, V.E.; Agostini, M.; Schwabe, J.W.; Soos, M.A.; Maslen, G.L.; Williams, T.D.; Lewis, H.; Schafer, A.J.; et al. Dominant negative mutations in human PPARgamma associated with severe insulin resistance, diabetes mellitus and hypertension. Nature 1999, 402, 880–883. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Zhou, L.L.; Ding, L.; Zhao, X.; Cao, H.; Fan, F.; Li, H.; Lou, R.; Du, Y.; Dong, S.; et al. PPARgamma agonist efatutazone and gefitinib synergistically inhibit the proliferation of EGFR-TKI-resistant lung adenocarcinoma cells via the PPARgamma/PTEN/Akt pathway. Exp. Cell Res. 2017, 361, 246–256. [Google Scholar] [CrossRef]
- Nakamura, N.; Ramaswamy, S.; Vazquez, F.; Signoretti, S.; Loda, M.; Sellers, W.R. Forkhead transcription factors are critical effectors of cell death and cell cycle arrest downstream of PTEN. Mol. Cell Biol. 2000, 20, 8969–8982. [Google Scholar] [CrossRef] [Green Version]
- Kawamura, K.; Cheng, Y.; Suzuki, N.; Deguchi, M.; Sato, Y.; Takae, S.; Ho, C.H.; Kawamura, N.; Tamura, M.; Hashimoto, S.; et al. Hippo signaling disruption and Akt stimulation of ovarian follicles for infertility treatment. Proc. Natl. Acad. Sci. USA 2013, 110, 17474–17479. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, M.O.; Sharma, S.; Kim, S.; Rickert, E.; Hsueh, K.; Hwang, V.; Olefsky, J.M.; Webster, N.J. Obese Neuronal PPARgamma Knockout Mice Are Leptin Sensitive but Show Impaired Glucose Tolerance and Fertility. Endocrinology 2017, 158, 121–133. [Google Scholar]
- Choi, Y.; Ballow, D.J.; Xin, Y.; Rajkovic, A. Lim homeobox gene, lhx8, is essential for mouse oocyte differentiation and survival. Biol. Reprod. 2008, 79, 442–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rak-Mardyla, A.; Drwal, E. In vitro interaction between resistin and peroxisome proliferator-activated receptor gamma in porcine ovarian follicles. Reprod. Fertil. Dev. 2016, 28, 357–368. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | cPA | |
|---|---|---|
| Total number of embryos | 61 | 57 |
| Number of pseudo mother | 9 | 7 |
| Delivery mother | 3 | 3 |
| Live pup | 20 | 18 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, S.Y.; Kim, R.; Jang, H.; Shin, D.H.; Lee, J.I.; Seol, D.; Lee, D.R.; Chang, E.M.; Lee, W.S. Peroxisome Proliferator-Activated Receptor Gamma Modulator Promotes Neonatal Mouse Primordial Follicle Activation In Vitro. Int. J. Mol. Sci. 2020, 21, 3120. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093120
Yoon SY, Kim R, Jang H, Shin DH, Lee JI, Seol D, Lee DR, Chang EM, Lee WS. Peroxisome Proliferator-Activated Receptor Gamma Modulator Promotes Neonatal Mouse Primordial Follicle Activation In Vitro. International Journal of Molecular Sciences. 2020; 21(9):3120. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093120
Chicago/Turabian StyleYoon, Sook Young, Ran Kim, Hyunmee Jang, Dong Hyuk Shin, Jin Il Lee, Dongwon Seol, Dong Ryul Lee, Eun Mi Chang, and Woo Sik Lee. 2020. "Peroxisome Proliferator-Activated Receptor Gamma Modulator Promotes Neonatal Mouse Primordial Follicle Activation In Vitro" International Journal of Molecular Sciences 21, no. 9: 3120. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093120