Bisphenol A Exposure in utero Disrupts Hypothalamic Gene Expression Particularly Genes Suspected in Autism Spectrum Disorders and Neuron and Hormone Signaling


Abstract
:1. Introduction
2. Results
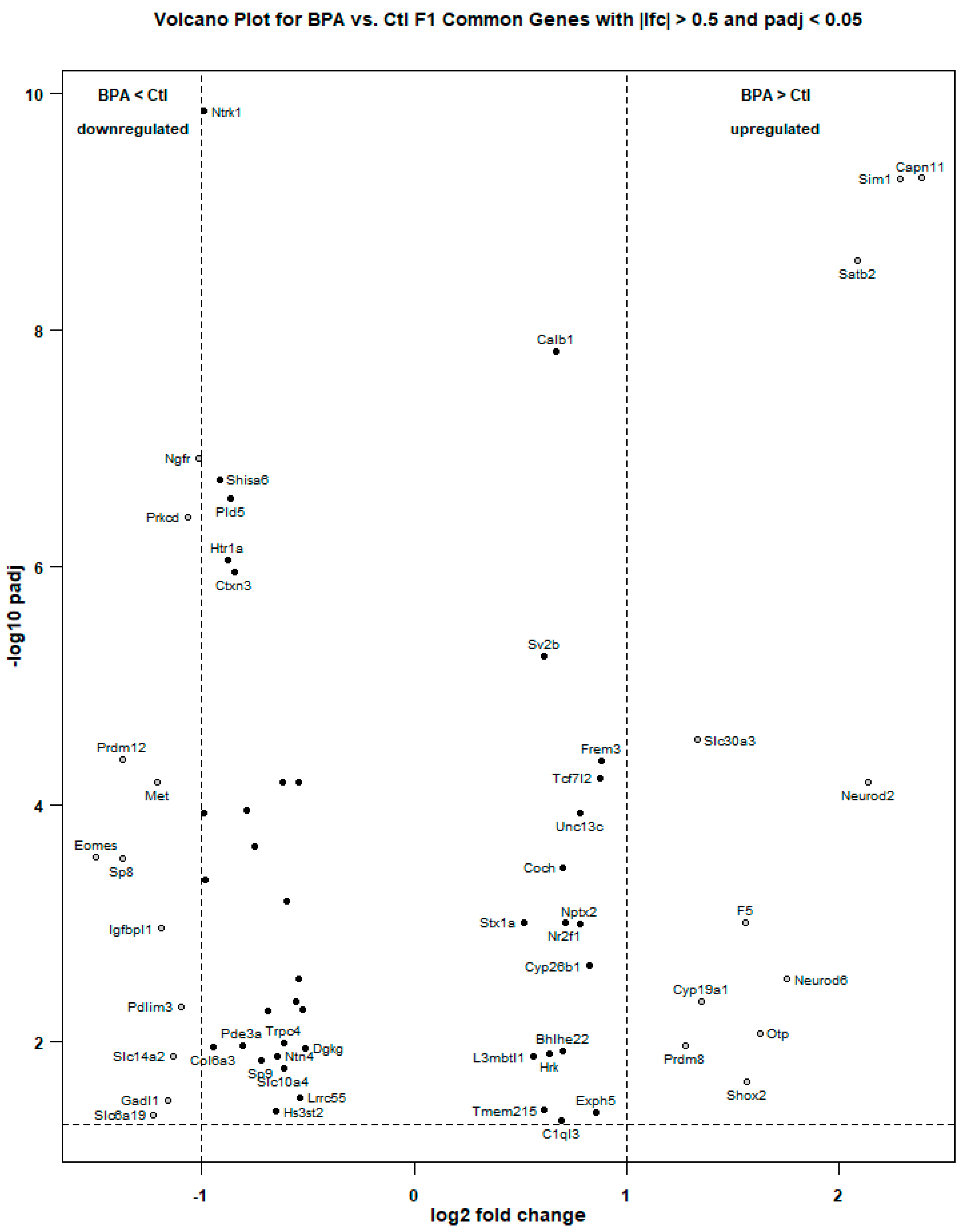
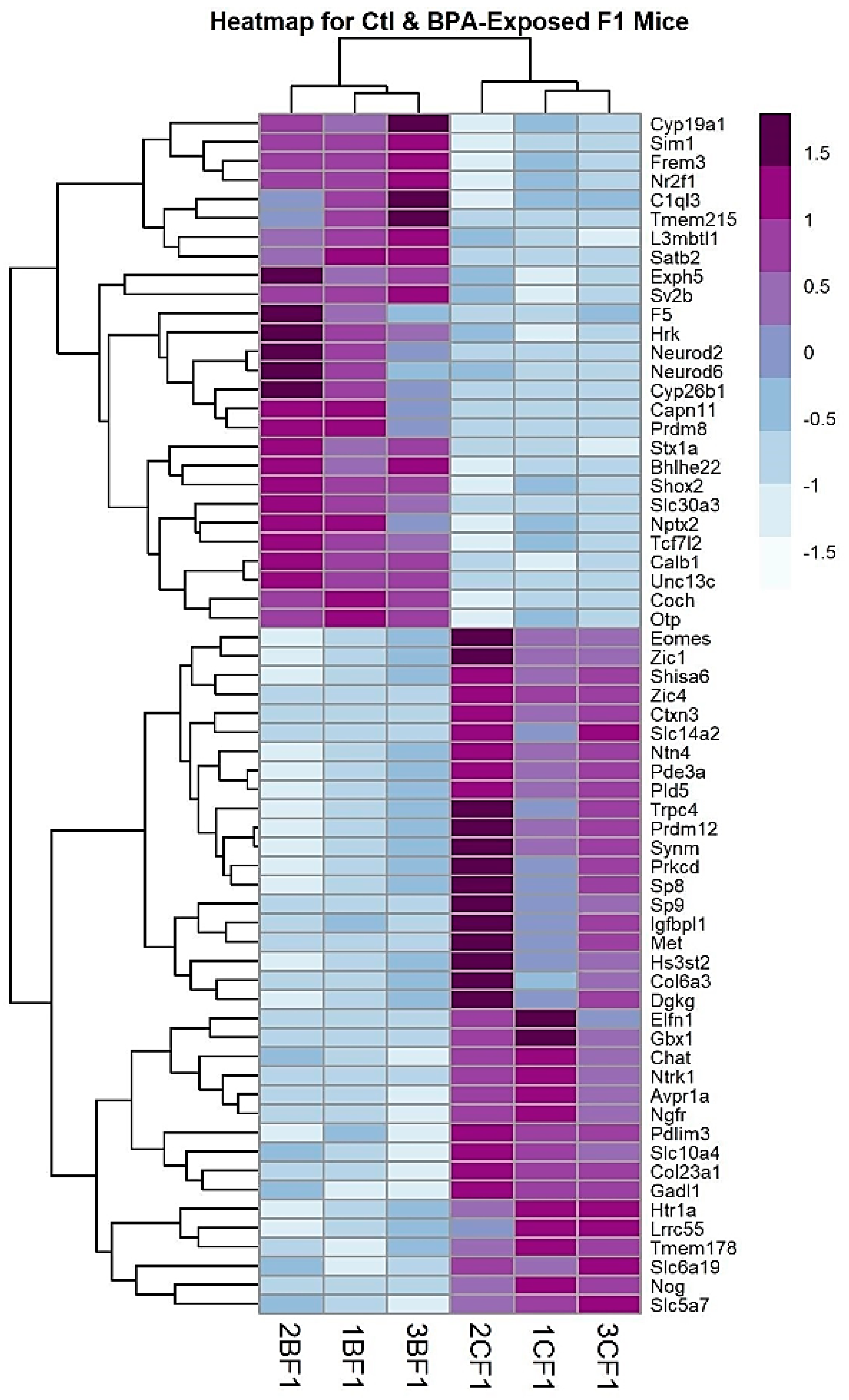
2.1. RNA-Sequencing and Analysis
2.2. Genes Involved in Cell Signaling, Hormone Activity, and Learning Behavior are Overrepresented in BPA Males
2.3. Comparison with Other Published Genomic Datasets
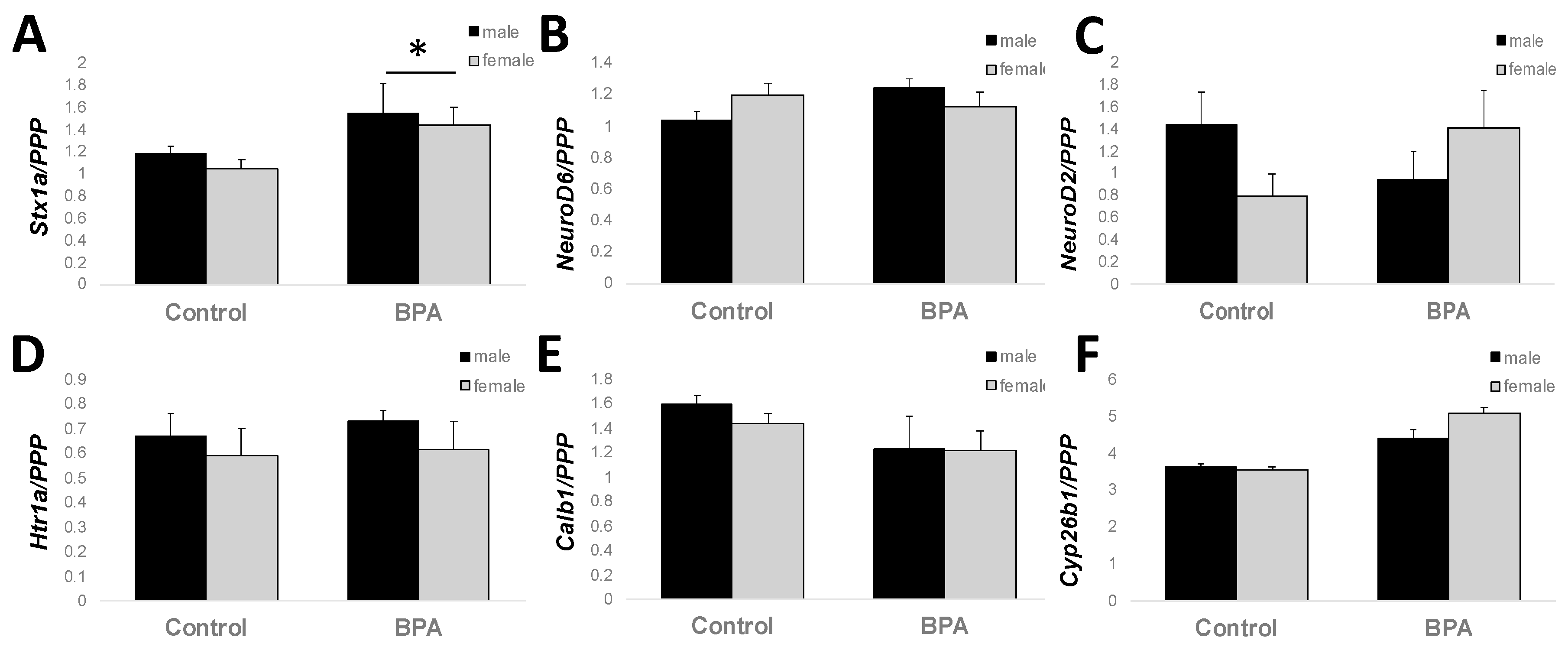
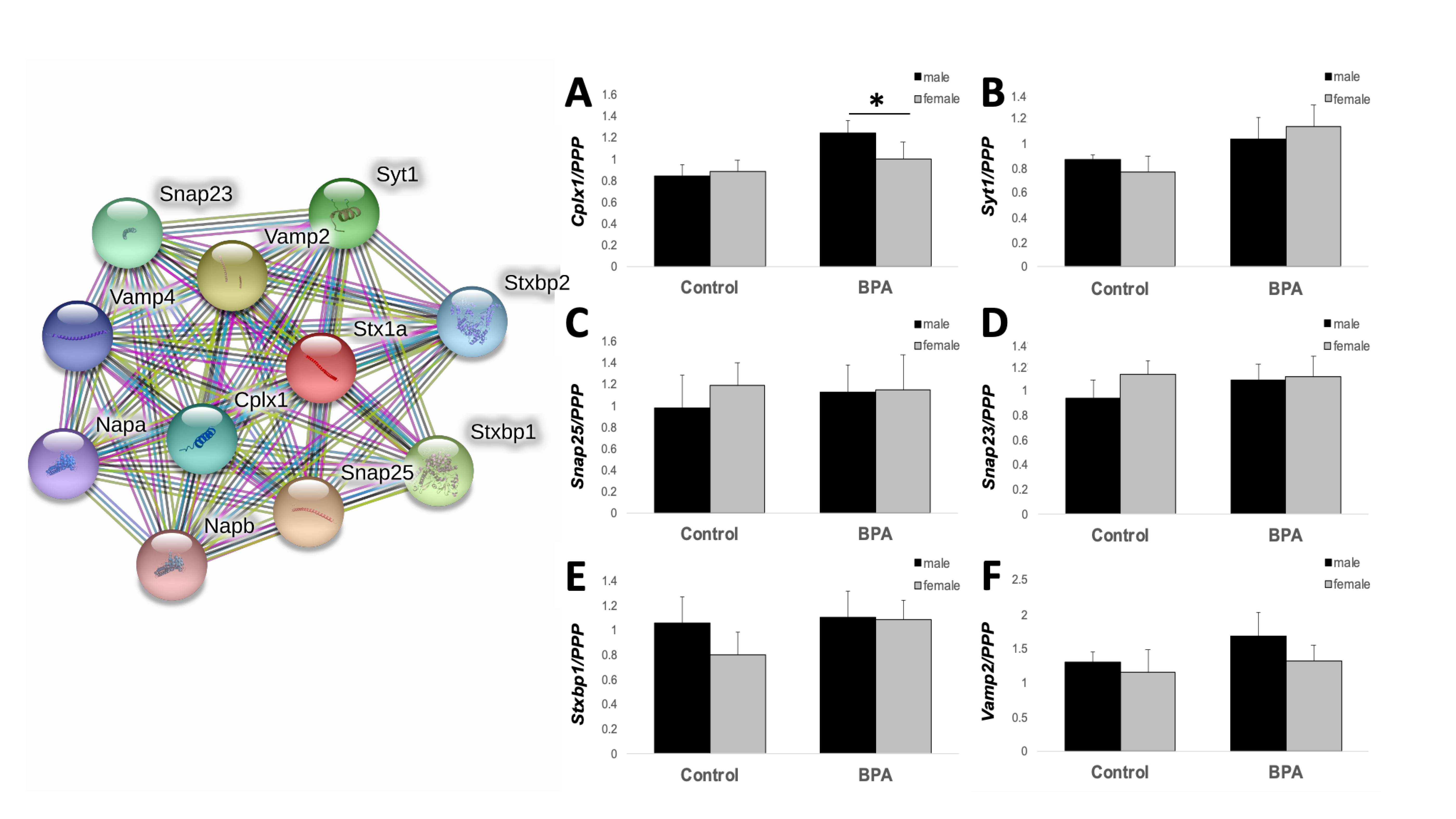
2.4. Adult Hippocampal Expression of Genes Involved in Learning and Memory
3. Discussion
4. Materials and Methods
4.1. Animals and Tissue Collection
4.2. RNA-Sequencing Analysis and Bioinformatics
4.3. RNA Isolation and qPCR Analysis
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| EDC | endocrine-disrupting compound |
| BPA | bisphenol A |
| ER | estrogen receptor |
| Esr1 | estrogen receptor alpha |
| GD | gestational day |
| PND | post-natal day |
| qPCR | quantitative polymerase chain reaction |
| SEM | standard error of the mean |
| Stx1a | syntaxin 1a |
References
- Vandenberg, L.N.; Chahoud, I.; Heindel, J.J.; Padmanabhan, V.; Paumgartten, F.J.R.; Schönfelder, G. Urinary, Circulating, and Tissue Biomonitoring Studies Indicate Widespread Exposure to Bisphenol A. Environ. Health Perspect. 2010, 118, 1055–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolstenholme, J.T.; Taylor, J.A.; Shetty, S.R.J.; Edwards, M.; Connelly, J.J.; Rissman, E.F. Gestational Exposure to Low Dose Bisphenol A Alters Social Behavior in Juvenile Mice. PLoS ONE 2011, 6, e25448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nesan, D.; Kurrasch, D.M. Gestational Exposure to Common Endocrine Disrupting Chemicals and Their Impact on Neurodevelopment and Behavior. Annu. Rev. Physiol. 2020, 82, 177–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gore, A.C.; Krishnan, K.; Reilly, M.P. Endocrine-disrupting chemicals: Effects on neuroendocrine systems and the neurobiology of social behavior. Horm. Behav. 2019, 111, 7–22. [Google Scholar] [CrossRef]
- Rosenfeld, C.S. Bisphenol A and phthalate endocrine disruption of parental and social behaviors. Front. Mol. Neurosci. 2015, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Braun, J.M.; Yolton, K.; Dietrich, K.N.; Hornung, R.; Ye, X.; Calafat, A.M.; Lanphear, B.P. Prenatal Bisphenol A Exposure and Early Childhood Behavior. Environ. Health Perspect. 2009, 117, 1945–1952. [Google Scholar] [CrossRef]
- Braun, J.M.; Kalkbrenner, A.E.; Calafat, A.M.; Yolton, K.; Ye, X.; Dietrich, K.N.; Lanphear, B.P. Impact of early-life bisphenol A exposure on behavior and executive function in children. Pediatrics 2011, 128, 873–882. [Google Scholar] [CrossRef] [Green Version]
- Casas, M.; Forns, J.; Martinez, D.; Avella-Garcia, C.; Valvi, D.; Ballesteros-Gómez, A.; Luque, N.; Rubio, S.; Julvez, J.; Sunyer, J.; et al. Exposure to bisphenol A during pregnancy and child neuropsychological development in the INMA-Sabadell cohort. Environ. Res. 2015, 142, 671–679. [Google Scholar] [CrossRef]
- Evans, S.F.; Kobrosly, R.W.; Barrett, E.S.; Thurston, S.W.; Calafat, A.M.; Weiss, B.; Stahlhut, R.; Yolton, K.; Swan, S.H. Prenatal bisphenol A exposure and maternally reported behavior in boys and girls. Neurotoxicology 2014, 45, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Perera, F.; Vishnevetsky, J.; Herbstman, J.B.; Calafat, A.M.; Xiong, W.; Rauh, V.; Wang, S. Prenatal Bisphenol A Exposure and Child Behavior in an Inner-City Cohort. Environ. Health Perspect. 2012, 120, 1190–1194. [Google Scholar] [CrossRef] [Green Version]
- Anderson, O.S.; Peterson, K.E.; Sanchez, B.N.; Zhang, Z.; Mancuso, P.; Dolinoy, D.C. Perinatal bisphenol A exposure promotes hyperactivity, lean body composition, and hormonal responses across the murine life course. FASEB J. 2013, 27, 1784–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, K.H.; Gatewood, J.D.; Howeth, C.; Rissman, E. Gestational exposure to bisphenol A and cross-fostering affect behaviors in juvenile mice. Horm. Behav. 2010, 58, 754–761. [Google Scholar] [CrossRef] [Green Version]
- Gioiosa, L.; Palanza, P.; Parmigiani, S.; Saal, F.S.V. Risk Evaluation of Endocrine-Disrupting Chemicals: Effects of Developmental Exposure to Low Doses of Bisphenol A on Behavior and Physiology in Mice (Mus musculus). Dose Response 2015, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, Y.J.; Park, H.R.; Kim, T.H.; Yang, W.-J.; Lee, J.-J.; Choi, S.Y.; Oh, S.B.; Lee, E.; Park, J.-H.; Kim, H.-P.; et al. High dose bisphenol A impairs hippocampal neurogenesis in female mice across generations. Toxicology 2012, 296, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Kundakovic, M.; Gudsnuk, K.; Herbstman, J.B.; Tang, D.; Perera, F.P.; Champagne, F.A. DNA methylation of BDNF as a biomarker of early-life adversity. Proc. Natl. Acad. Sci. USA 2015, 112, 6807–6813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolstenholme, J.T.; Drobná, Z.; Henriksen, A.D.; Goldsby, J.A.; Stevenson, R.; Irvin, J.W.; Flaws, J.A.; Rissman, E.F. Transgenerational Bisphenol A Causes Deficits in Social Recognition and Alters Postsynaptic Density Genes in Mice. Endocrinology 2019, 160, 1854–1867. [Google Scholar] [CrossRef]
- Wolstenholme, J.T.; Edwards, M.; Shetty, S.R.J.; Gatewood, J.D.; Taylor, J.A.; Rissman, E.; Connelly, J.J. Gestational exposure to bisphenol a produces transgenerational changes in behaviors and gene expression. Endocrinology 2012, 153, 3828–3838. [Google Scholar] [CrossRef] [Green Version]
- Wolstenholme, J.T.; Goldsby, J.A.; Rissman, E.F. Transgenerational effects of prenatal bisphenol A on social recognition. Horm. Behav. 2013, 64, 833–839. [Google Scholar] [CrossRef] [Green Version]
- Kundakovic, M.; Gudsnuk, K.; Franks, B.; Madrid, J.; Miller, R.L.; Perera, F.P.; Champagne, F.A. Sex-specific epigenetic disruption and behavioral changes following low-dose in utero bisphenol A exposure. Proc. Natl. Acad. Sci. USA 2013, 110, 9956–9961. [Google Scholar] [CrossRef] [Green Version]
- Shu, L.; Meng, Q.; Diamante, G.; Tsai, B.; Chen, Y.-W.; Mikhail, A.; Luk, H.; Ritz, B.; Allard, P.; Yang, X. Prenatal Bisphenol A Exposure in Mice Induces Multitissue Multiomics Disruptions Linking to Cardiometabolic Disorders. Endocrinology 2018, 160, 409–429. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.A.; Spollen, W.G.; Manshack, L.K.; Bivens, N.; Givan, S.; Rosenfeld, C.S. Hypothalamic transcriptomic alterations in male and female California mice (Peromyscus californicus) developmentally exposed to bisphenol A or ethinyl estradiol. Physiol. Rep. 2017, 5, e13133. [Google Scholar] [CrossRef] [PubMed]
- Arambula, S.E.; Belcher, S.M.; Planchart, A.; Turner, S.D.; Patisaul, H.B. Impact of Low Dose Oral Exposure to Bisphenol A (BPA) on the Neonatal Rat Hypothalamic and Hippocampal Transcriptome: A CLARITY-BPA Consortium Study. Endocrinology 2016, 157, 3856–3872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kardas, F.; Bayram, A.K.; Demirci, E.; Akin, L.; Ozmen, S.; Kendirci, M.; Canpolat, M.; Oztop, D.B.; Narin, F.; Gumus, H.; et al. Increased Serum Phthalates (MEHP, DEHP) and Bisphenol A Concentrations in Children with Autism Spectrum Disorder: The Role of Endocrine Disruptors in Autism Etiopathogenesis. J. Child. Neurol. 2016, 31, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Kondolot, M.; Özmert, E.N.; Ascı, A.; Erkekoğlu, P.; Oztop, D.B.; Gümüş, H.; Kocer-Gumusel, B.; Yurdakök, K. Plasma phthalate and bisphenol a levels and oxidant-antioxidant status in autistic children. Environ. Toxicol. Pharmacol. 2016, 43, 149–158. [Google Scholar] [CrossRef]
- Stein, T.P.; Schluter, M.D.; Steer, R.A.; Guo, L.; Ming, X. Bisphenol A Exposure in Children with Autism Spectrum Disorders. Autism Res. 2015, 8, 272–283. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Diamond, D.M. The stressed hippocampus, synaptic plasticity and lost memories. Nat. Rev. Neurosci. 2002, 3, 453–462. [Google Scholar] [CrossRef]
- MacLusky, N.J.; Hajszan, T.; Leranth, C. The Environmental Estrogen Bisphenol A Inhibits Estradiol-Induced Hippocampal Synaptogenesis. Environ. Health Perspect. 2005, 113, 675–679. [Google Scholar] [CrossRef]
- Hajszan, T.; Leranth, C. Bisphenol A interferes with synaptic remodeling. Front. Neuroendocrinol. 2010, 31, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Elsworth, J.D.; Jentsch, J.D.; Vandevoort, C.A.; Roth, R.H.; Redmond, D.E., Jr.; Leranth, C. Prenatal exposure to bisphenol A impacts midbrain dopamine neurons and hippocampal spine synapses in non-human primates. Neurotoxicology 2013, 35, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Kimura, E.; Matsuyoshi, C.; Miyazaki, W.; Benner, S.; Hosokawa, M.; Yokoyama, K.; Kakeyama, M.; Tohyama, C. Prenatal exposure to bisphenol A impacts neuronal morphology in the hippocampal CA1 region in developing and aged mice. Arch. Toxicol. 2015, 90, 691–700. [Google Scholar] [CrossRef] [Green Version]
- Goldsby, J.A.; Wolstenholme, J.T.; Rissman, E.F. Multi- and Transgenerational Consequences of Bisphenol A on Sexually Dimorphic Cell Populations in Mouse Brain. Endocrinology 2017, 158, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thongkorn, S.; Kanlayaprasit, S.; Jindatip, D.; Tencomnao, T.; Hu, V.W.; Sarachana, T. Sex Differences in the Effects of Prenatal Bisphenol A Exposure on Genes Associated with Autism Spectrum Disorder in the Hippocampus. Sci. Rep. 2019, 9, 3038. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Li, T.; Gong, H.; Chen, Z.; Jin, Y.; Xu, G.; Wang, M. Bisphenol A Impairs Synaptic Plasticity by Both Pre- and Postsynaptic Mechanisms. Adv. Sci. 2017, 4, 1600493. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Xie, L.; Hong, X.; Ruan, Q.; Lu, H.; Zhang, Q.; Zhang, G.; Liu, X. Perinatal exposure to bisphenol-A inhibits synaptogenesis and affects the synaptic morphological development in offspring male mice. Chemosphere 2013, 91, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Thakur, M.K. Anxiety like behavior due to perinatal exposure to Bisphenol-A is associated with decrease in excitatory to inhibitory synaptic density of male mouse brain. Toxicology 2017, 378, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Komada, M.; Asai, Y.; Morii, M.; Matsuki, M.; Sato, M.; Nagao, T. Maternal bisphenol A oral dosing relates to the acceleration of neurogenesis in the developing neocortex of mouse fetuses. Toxicology 2012, 295, 31–38. [Google Scholar] [CrossRef]
- Nakamura, K.; Itoh, K.; Sugimoto, T.; Fushiki, S. Prenatal exposure to bisphenol A affects adult murine neocortical structure. Neurosci. Lett. 2007, 420, 100–105. [Google Scholar] [CrossRef]
- Komada, M.; Itoh, S.; Kawachi, K.; Kagawa, N.; Ikeda, Y.; Nagao, T. Newborn mice exposed prenatally to bisphenol A show hyperactivity and defective neocortical development. Toxicology 2014, 323, 51–60. [Google Scholar] [CrossRef]
- Lin, C.-H.; Hansen, S.; Wang, Z.; Storm, D.R.; Tapscott, S.J.; Olson, J.M. The dosage of the neuroD2 transcription factor regulates amygdala development and emotional learning. Proc. Natl. Acad. Sci. USA 2005, 102, 14877–14882. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, T.; Mishima, T.; Kofuji, T.; Chiba, T.; Tanaka, K.; Yamamoto, A.; Akagawa, K. Analysis of knock-out mice to determine the role of HPC-1/syntaxin 1A in expressing synaptic plasticity. J. Neurosci. 2006, 26, 5767–5776. [Google Scholar] [CrossRef]
- Fujiwara, T.; Snada, M.; Kofuji, T.; Yoshikawa, T.; Akagawa, K. HPC-1/syntaxin 1A gene knockout mice show abnormal behavior possibly related to a disruption in 5-HTergic systems. Eur. J. Neurosci. 2010, 32, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T.; Sanada, M.; Kofuji, T.; Akagawa, K. Unusual social behavior in HPC-1/syntaxin1A knockout mice is caused by disruption of the oxytocinergic neural system. J. Neurochem. 2016, 138, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kofuji, T.; Hayashi, Y.; Fujiwara, T.; Sanada, M.; Tamaru, M.; Akagawa, K. A part of patients with autism spectrum disorder has haploidy of HPC-1/syntaxin1A gene that possibly causes behavioral disturbance as in experimentally gene ablated mice. Neurosci. Lett. 2017, 644, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Drobná, Z.; Henriksen, A.D.; Wolstenholme, J.T.; Montiel, C.; Lambeth, P.S.; Shang, S.; Harris, E.; Zhou, C.; Flaws, J.A.; Adli, M.; et al. Transgenerational Effects of Bisphenol A on Gene Expression and DNA Methylation of Imprinted Genes in Brain. Endocrinology 2018, 159, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2012, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Blankenberg, D.; Von Kuster, G.; Coraor, N.; Ananda, G.; Lazarus, R.; Mangan, M.; Nekrutenko, A.; Taylor, J. Galaxy: A Web-Based Genome Analysis Tool for Experimentalists. Curr. Protoc. Mol. Biol. 2010, 19, 1–21. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.A.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Gordon, A.; Hannon, G. Fastx-toolkit. FASTQ/A Short-reads Preprocessing Tools. Available online: http://hannonlab.cshl.edu/fastx_toolkit/ (accessed on 31 August 2019).
- Kroll, K.W.; Mokaram, N.E.; Pelletier, A.R.; Frankhouser, D.E.; Westphal, M.S.; Stump, P.A.; Stump, C.L.; Bundschuh, R.; Blachly, J.S.; Yan, P. Quality Control for RNA-Seq (QuaCRS): An Integrated Quality Control Pipeline. Cancer Inform. 2014, 13 (Suppl. S3), 7–14. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolstenholme, J.; Mahmood, T.; Harris, G.M.; Abbas, S.; Miles, M.F. Intermittent Ethanol during Adolescence Leads to Lasting Behavioral Changes in Adulthood and Alters Gene Expression and Histone Methylation in the PFC. Front. Mol. Neurosci. 2017, 10, 307. [Google Scholar] [CrossRef] [PubMed]
- Wolstenholme, J.T.; Rissman, E.F.; Bekiranov, S. Sexual differentiation in the developing mouse brain: Contributions of sex chromosome genes. Genes, Brain Behav. 2013, 12, 166–180. [Google Scholar] [CrossRef] [Green Version]
- Wolstenholme, J.T.; Warner, J.A.; Capparuccini, M.I.; Archer, K.J.; Shelton, K.L.; Miles, M.F. Genomic Analysis of Individual Differences in Ethanol Drinking: Evidence for Non-Genetic Factors in C57BL/6 Mice. PLoS ONE 2011, 6, e21100. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DESeq2/Galaxy Union Genes | DESeq2 DE Results | Galaxy CuffDiff DE Results | ||||||
|---|---|---|---|---|---|---|---|---|
| Gene | RefSeqID | Chr | lfc | p value | padj | log2fc | p_value | q_value |
| Eomes | NM_010136 | 9 | −1.4979 | 0.0000 | 0.0003 | −1.9697 | 0.0001 | 0.0055 |
| Prdm12 | NM_001123362 | 2 | −1.3741 | 0.0000 | 0.0000 | −1.4659 | 0.0001 | 0.0055 |
| Sp8 | NM_177082 | 12 | −1.3726 | 0.0000 | 0.0003 | −1.5349 | 0.0001 | 0.0055 |
| Slc6a19 | NM_028878 | 13 | −1.2254 | 0.0003 | 0.0419 | −1.9559 | 0.0001 | 0.0055 |
| Met | NM_008591 | 6 | −1.2116 | 0.0000 | 0.0001 | −1.3163 | 0.0001 | 0.0055 |
| Igfbpl1 | NM_018741 | 4 | −1.1905 | 0.0000 | 0.0011 | −1.2246 | 0.0001 | 0.0055 |
| Gadl1 | NM_028638 | 9 | −1.1615 | 0.0002 | 0.0313 | −1.2434 | 0.0001 | 0.0055 |
| Slc14a2 | NM_001110274 | 18 | −1.1338 | 0.0001 | 0.0133 | −1.4175 | 0.0001 | 0.0055 |
| Pdlim3 | NM_016798 | 8 | −1.0986 | 0.0000 | 0.0051 | −1.1447 | 0.0001 | 0.0055 |
| Prkcd | NM_011103 | 14 | −1.0667 | 0.0000 | 0.0000 | −1.1190 | 0.0001 | 0.0055 |
| Ngfr | NM_033217 | 11 | −1.0178 | 0.0000 | 0.0000 | −1.0344 | 0.0001 | 0.0055 |
| Ntrk1 | NM_001033124 | 3 | −0.9894 | 0.0000 | 0.0000 | −0.9987 | 0.0001 | 0.0055 |
| Avpr1a | NM_016847 | 10 | −0.9880 | 0.0000 | 0.0001 | −1.0424 | 0.0001 | 0.0055 |
| Gbx1 | NM_015739 | 5 | −0.9859 | 0.0000 | 0.0004 | −1.1339 | 0.0001 | 0.0055 |
| Col6a3 | NM_001243009 | 1 | −0.9442 | 0.0001 | 0.0112 | −1.1950 | 0.0001 | 0.0055 |
| Shisa6 | NM_001034874 | 11 | −0.9164 | 0.0000 | 0.0000 | −0.9462 | 0.0001 | 0.0055 |
| Htr1a | NM_008308 | 13 | −0.8765 | 0.0000 | 0.0000 | −0.8989 | 0.0001 | 0.0055 |
| Pld5 | NM_176916 | 1 | −0.8610 | 0.0000 | 0.0000 | −0.9883 | 0.0004 | 0.0307 |
| Ctxn3 | NM_001134697 | 18 | −0.8473 | 0.0000 | 0.0000 | −0.9263 | 0.0001 | 0.0055 |
| Pde3a | NM_018779 | 6 | −0.8090 | 0.0001 | 0.0108 | −0.8426 | 0.0001 | 0.0055 |
| Zic1 | NM_009573 | 9 | −0.7906 | 0.0000 | 0.0001 | −0.8006 | 0.0001 | 0.0055 |
| Chat | NM_009891 | 14 | −0.7488 | 0.0000 | 0.0002 | −0.7671 | 0.0001 | 0.0055 |
| Sp9 | NM_001005343 | 2 | −0.7181 | 0.0001 | 0.0142 | −0.7211 | 0.0001 | 0.0055 |
| Nog | NM_008711 | 11 | −0.6856 | 0.0000 | 0.0055 | −0.6826 | 0.0001 | 0.0097 |
| Hs3st2 | NM_001081327 | 7 | −0.6508 | 0.0003 | 0.0391 | −0.6381 | 0.0001 | 0.0055 |
| Ntn4 | NM_021320 | 10 | −0.6440 | 0.0001 | 0.0134 | −0.6611 | 0.0001 | 0.0097 |
| Synm | NM_207663 | 7 | −0.6170 | 0.0000 | 0.0001 | −0.7814 | 0.0002 | 0.0138 |
| Trpc4 | NM_001253682 | 3 | −0.6157 | 0.0000 | 0.0103 | −1.9274 | 0.0003 | 0.0243 |
| Slc10a4 | NM_173403 | 5 | −0.6138 | 0.0001 | 0.0167 | −0.6536 | 0.0001 | 0.0055 |
| Zic4 | NM_009576 | 9 | −0.6017 | 0.0000 | 0.0007 | −0.6062 | 0.0001 | 0.0055 |
| Elfn1 | NM_175522 | 5 | −0.5569 | 0.0000 | 0.0045 | −0.5598 | 0.0001 | 0.0055 |
| Slc5a7 | NM_022025 | 17 | −0.5469 | 0.0000 | 0.0030 | −0.5656 | 0.0001 | 0.0055 |
| Col23a1 | NM_153393 | 11 | −0.5445 | 0.0000 | 0.0001 | −0.5308 | 0.0001 | 0.0055 |
| Lrrc55 | NM_001033346 | 2 | −0.5366 | 0.0002 | 0.0299 | −0.5715 | 0.0001 | 0.0055 |
| Tmem178 | NM_026516 | 17 | −0.5249 | 0.0000 | 0.0054 | −0.5279 | 0.0001 | 0.0055 |
| Dgkg | NM_138650 | 16 | −0.5125 | 0.0001 | 0.0113 | −0.5270 | 0.0001 | 0.0055 |
| Stx1a | NM_016801 | 5 | 0.5159 | 0.0000 | 0.0010 | 0.5597 | 0.0001 | 0.0055 |
| L3mbtl1 | NM_001081338 | 2 | 0.5599 | 0.0001 | 0.0134 | 0.5238 | 0.0002 | 0.0175 |
| Sv2b | NM_001109753 | 7 | 0.6092 | 0.0000 | 0.0000 | 0.7609 | 0.0006 | 0.0389 |
| Tmem215 | NM_001166009 | 4 | 0.6126 | 0.0003 | 0.0379 | 0.6488 | 0.0001 | 0.0055 |
| Hrk | NM_007545 | 5 | 0.6365 | 0.0001 | 0.0127 | 0.6613 | 0.0001 | 0.0055 |
| Calb1 | NM_009788 | 4 | 0.6679 | 0.0000 | 0.0000 | 0.6607 | 0.0001 | 0.0055 |
| C1ql3 | NM_153155 | 2 | 0.6905 | 0.0003 | 0.0467 | 0.7652 | 0.0001 | 0.0055 |
| Bhlhe22 | NM_021560 | 3 | 0.6993 | 0.0001 | 0.0119 | 0.7375 | 0.0001 | 0.0055 |
| Coch | NM_001198835 | 12 | 0.7013 | 0.0000 | 0.0003 | 0.7497 | 0.0001 | 0.0055 |
| Nr2f1 | NM_010151 | 13 | 0.7079 | 0.0000 | 0.0010 | 0.7103 | 0.0001 | 0.0055 |
| Nptx2 | NM_016789 | 5 | 0.7789 | 0.0000 | 0.0010 | 0.8747 | 0.0001 | 0.0055 |
| Unc13c | NM_001081153 | 9 | 0.7824 | 0.0000 | 0.0001 | 0.7985 | 0.0001 | 0.0055 |
| Cyp26b1 | NM_001177713 | 6 | 0.8214 | 0.0000 | 0.0023 | 0.8481 | 0.0001 | 0.0055 |
| Exph5 | NM_176846 | 9 | 0.8528 | 0.0003 | 0.0393 | 0.8928 | 0.0001 | 0.0055 |
| Tcf7l2 | NM_001142924 | 19 | 0.8720 | 0.0000 | 0.0001 | 1.3745 | 0.0001 | 0.0055 |
| Frem3 | NM_001167898 | 8 | 0.8789 | 0.0000 | 0.0000 | 0.9520 | 0.0001 | 0.0055 |
| Prdm8 | NM_029947 | 5 | 1.2730 | 0.0001 | 0.0109 | 1.6396 | 0.0001 | 0.0055 |
| Slc30a3 | NM_011773 | 5 | 1.3293 | 0.0000 | 0.0000 | 1.3863 | 0.0001 | 0.0055 |
| Cyp19a1 | NM_007810 | 9 | 1.3506 | 0.0000 | 0.0045 | 1.6930 | 0.0001 | 0.0055 |
| F5 | NM_007976 | 1 | 1.5600 | 0.0000 | 0.0010 | 1.9068 | 0.0001 | 0.0055 |
| Shox2 | NM_001302358 | 3 | 1.5631 | 0.0001 | 0.0218 | 3.4934 | 0.0001 | 0.0055 |
| Otp | NM_011021 | 13 | 1.6254 | 0.0000 | 0.0086 | 2.7132 | 0.0003 | 0.0243 |
| Neurod6 | NM_009717 | 6 | 1.7555 | 0.0000 | 0.0030 | 3.1023 | 0.0008 | 0.0494 |
| Satb2 | NM_139146 | 1 | 2.0837 | 0.0000 | 0.0000 | 2.4912 | 0.0001 | 0.0055 |
| Neurod2 | NM_010895 | 11 | 2.1360 | 0.0000 | 0.0001 | 3.8885 | 0.0003 | 0.0212 |
| Sim1 | NM_011376 | 10 | 2.2845 | 0.0000 | 0.0000 | 2.8305 | 0.0001 | 0.0055 |
| Capn11 | NM_001013767 | 17 | 2.3889 | 0.0000 | 0.0000 | 2.7544 | 0.0001 | 0.0055 |
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| Stx1a | CCGAACCCCGATGAGAAGAC | TGCTCTTTAGCTTGGAGCGA |
| NeuroD6 | ATGCGACACTCAGCCTGAAA | CTGGGATTCGGGCATTACGA |
| NeuroD2 | AAGCCAGTGTCTCTTCGTGG | TTGGACAGCTTCTGCGTCTT |
| Htr1a | TACTCCACTTTCGGCGCTTT | GGAGGTAGCTCCTGATTCGC |
| Calb1 | ATTTCGACGCTGACGGAAGT | TGGGTAAGACGTGAGCCAAC |
| Cyp26b1 | CTGGTTGCTACAGGGTTCCG | CTCAAGTGCCTCATGGCTGA |
| Cplx1 | TTGGAAGGCAGAAGACCCTGA | TTATACCCTGCCGCATGACC |
| Syt1 | CAGGCCCTTAAGGATGACGA | TCCGAGTATGGCACCTTGAA |
| Snap25 | CAAGGCGAACAACTGGAACG | AGCTTGTTACAGGGACACACA |
| Snap23 | AGCGGGACAGAGTATCCGTA | CCATCTCCCCATGTTGCCTT |
| Stxbp1 | TCACGGATTCCACACTACGC | TCCATGAGGAATTTGGTGGGA |
| Vamp2 | CTGGTGGAAAAACCTCAAGATGAT | GGGTGTTAAGGACAACCGGA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henriksen, A.D.; Andrade, A.; Harris, E.P.; Rissman, E.F.; Wolstenholme, J.T. Bisphenol A Exposure in utero Disrupts Hypothalamic Gene Expression Particularly Genes Suspected in Autism Spectrum Disorders and Neuron and Hormone Signaling. Int. J. Mol. Sci. 2020, 21, 3129. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093129
Henriksen AD, Andrade A, Harris EP, Rissman EF, Wolstenholme JT. Bisphenol A Exposure in utero Disrupts Hypothalamic Gene Expression Particularly Genes Suspected in Autism Spectrum Disorders and Neuron and Hormone Signaling. International Journal of Molecular Sciences. 2020; 21(9):3129. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093129
Chicago/Turabian StyleHenriksen, Anne D., Alejandro Andrade, Erin P. Harris, Emilie F. Rissman, and Jennifer T. Wolstenholme. 2020. "Bisphenol A Exposure in utero Disrupts Hypothalamic Gene Expression Particularly Genes Suspected in Autism Spectrum Disorders and Neuron and Hormone Signaling" International Journal of Molecular Sciences 21, no. 9: 3129. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093129