IL-4 and SDF-1 Increase Adipose Tissue-Derived Stromal Cell Ability to Improve Rat Skeletal Muscle Regeneration

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
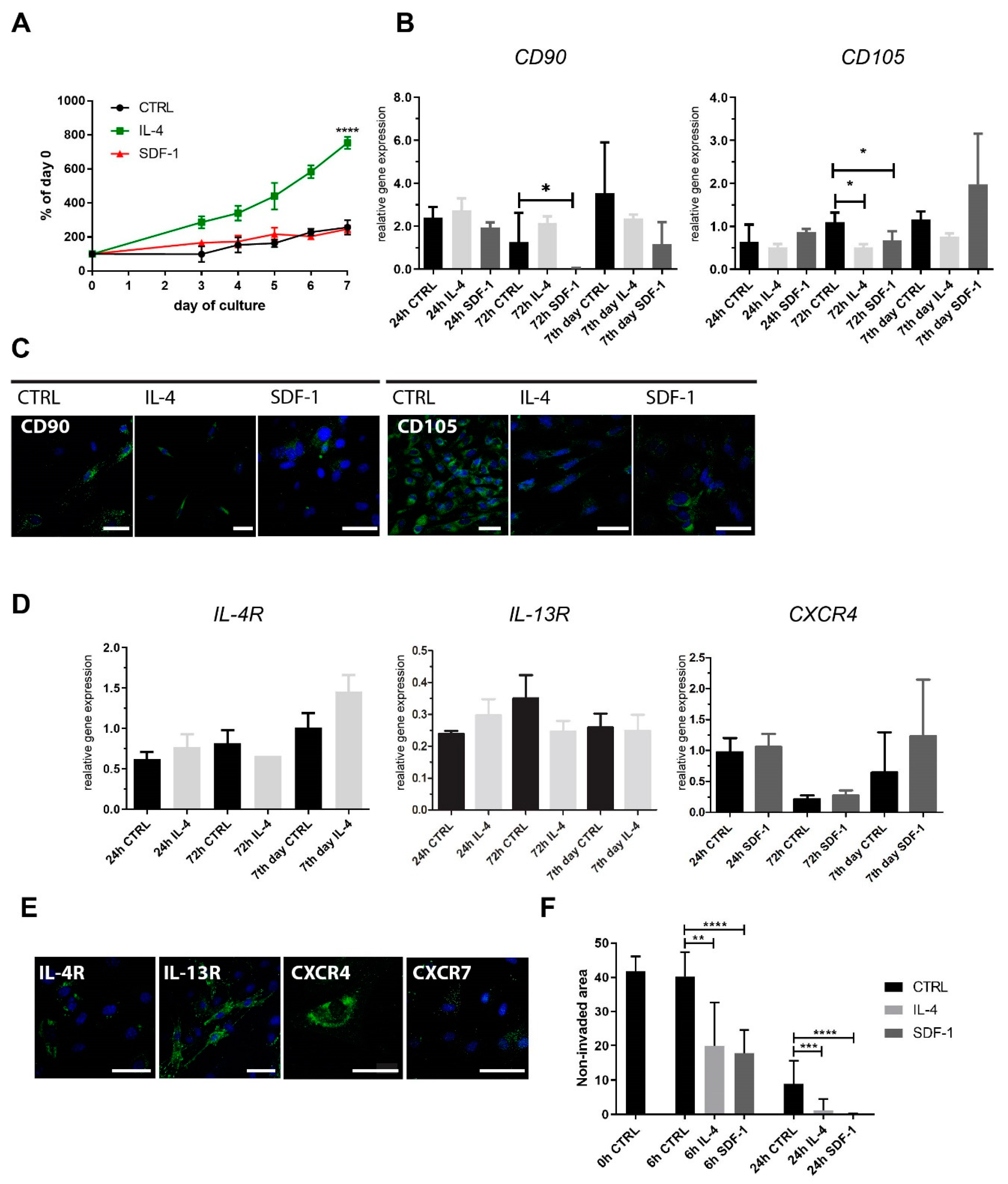
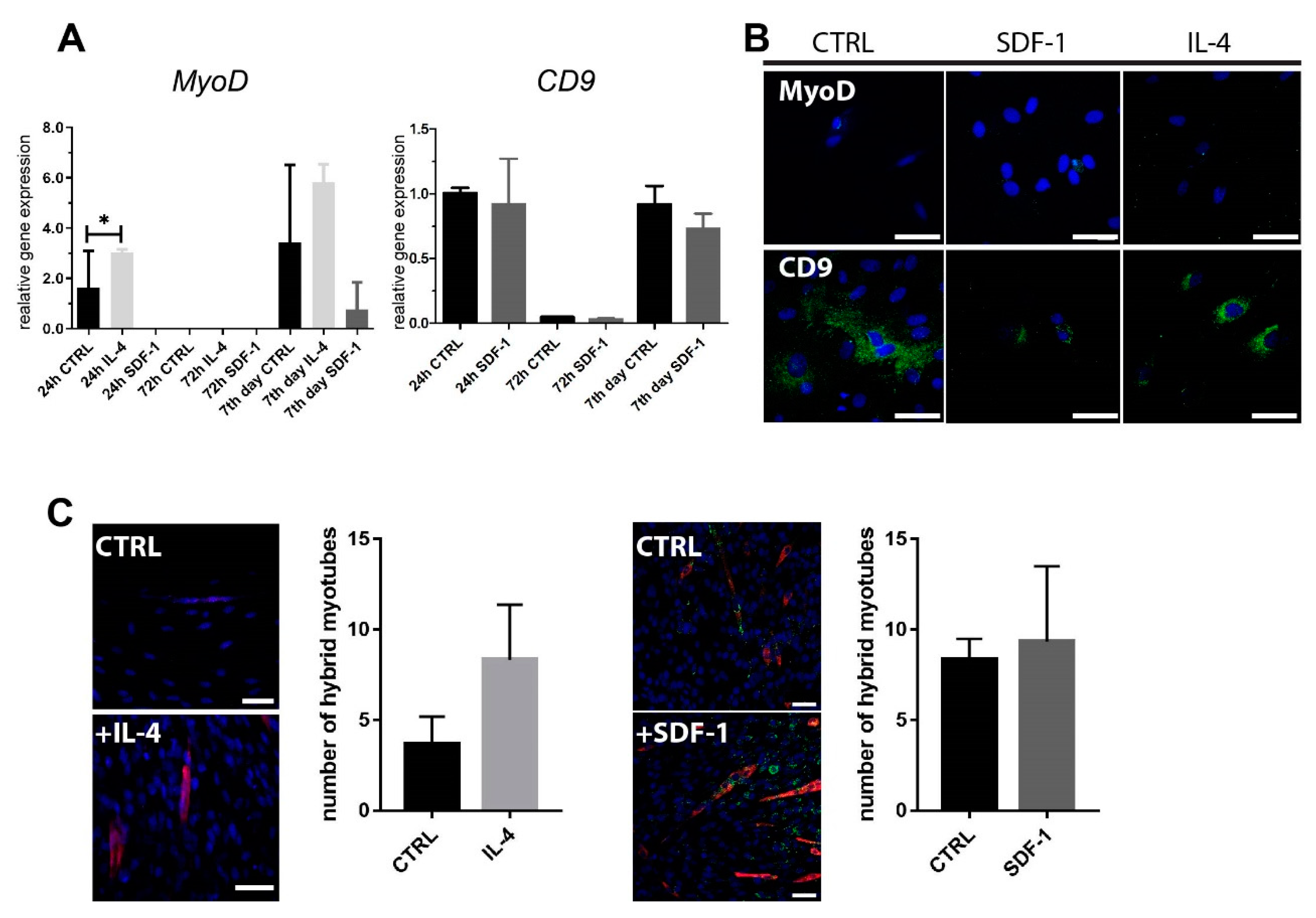
2.1. Rat ADSC Response to IL-4 or SDF-1 Treatment In Vitro
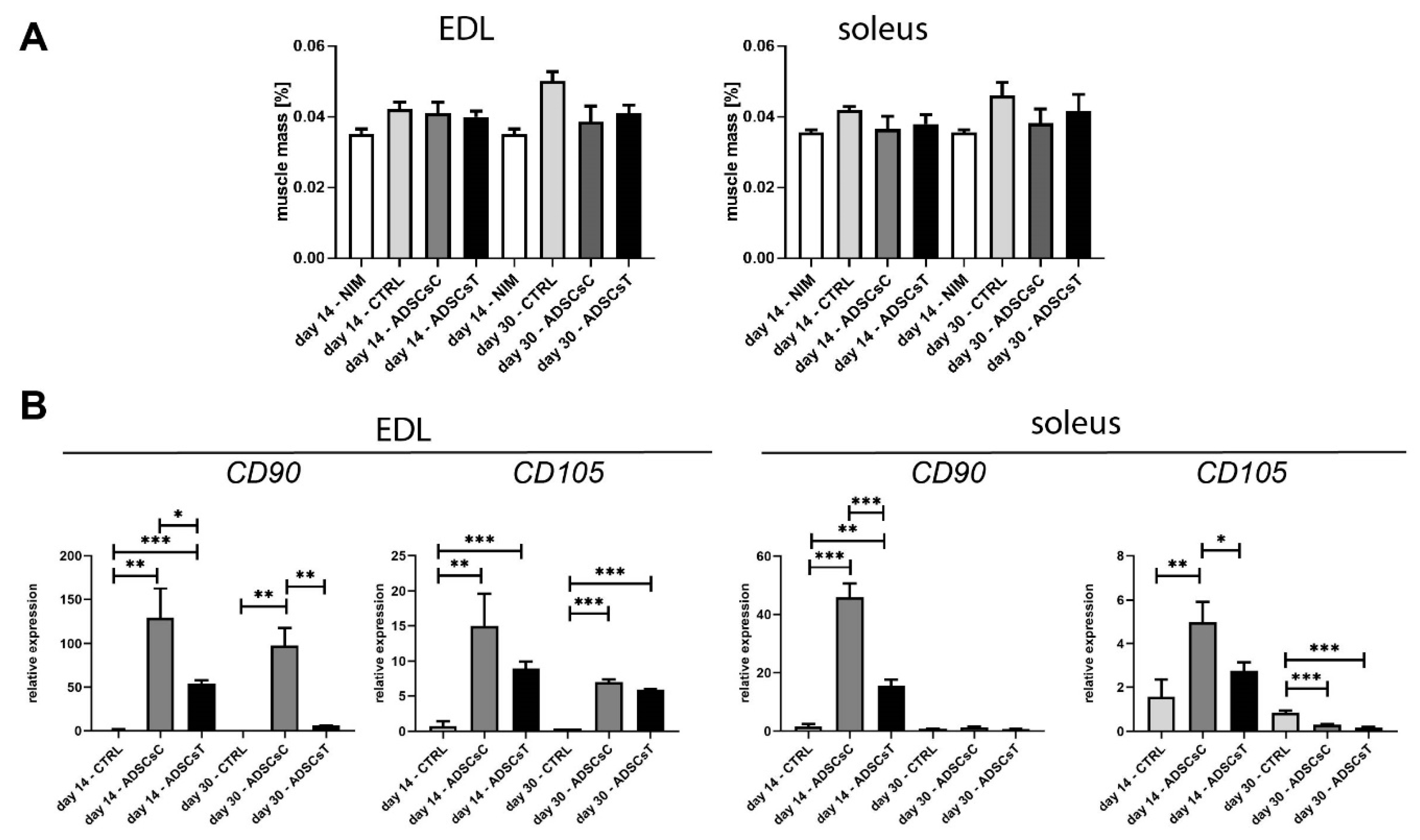
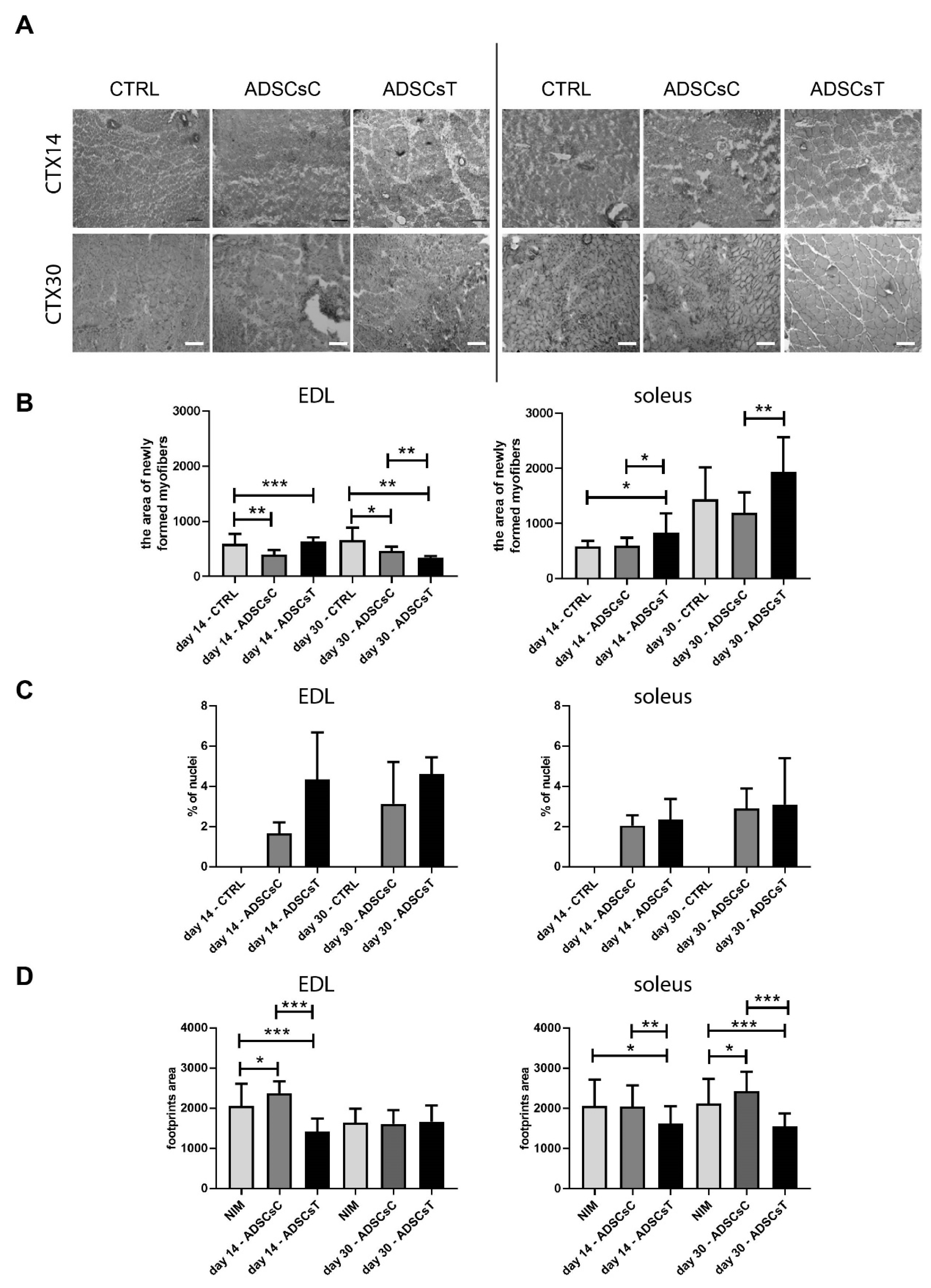
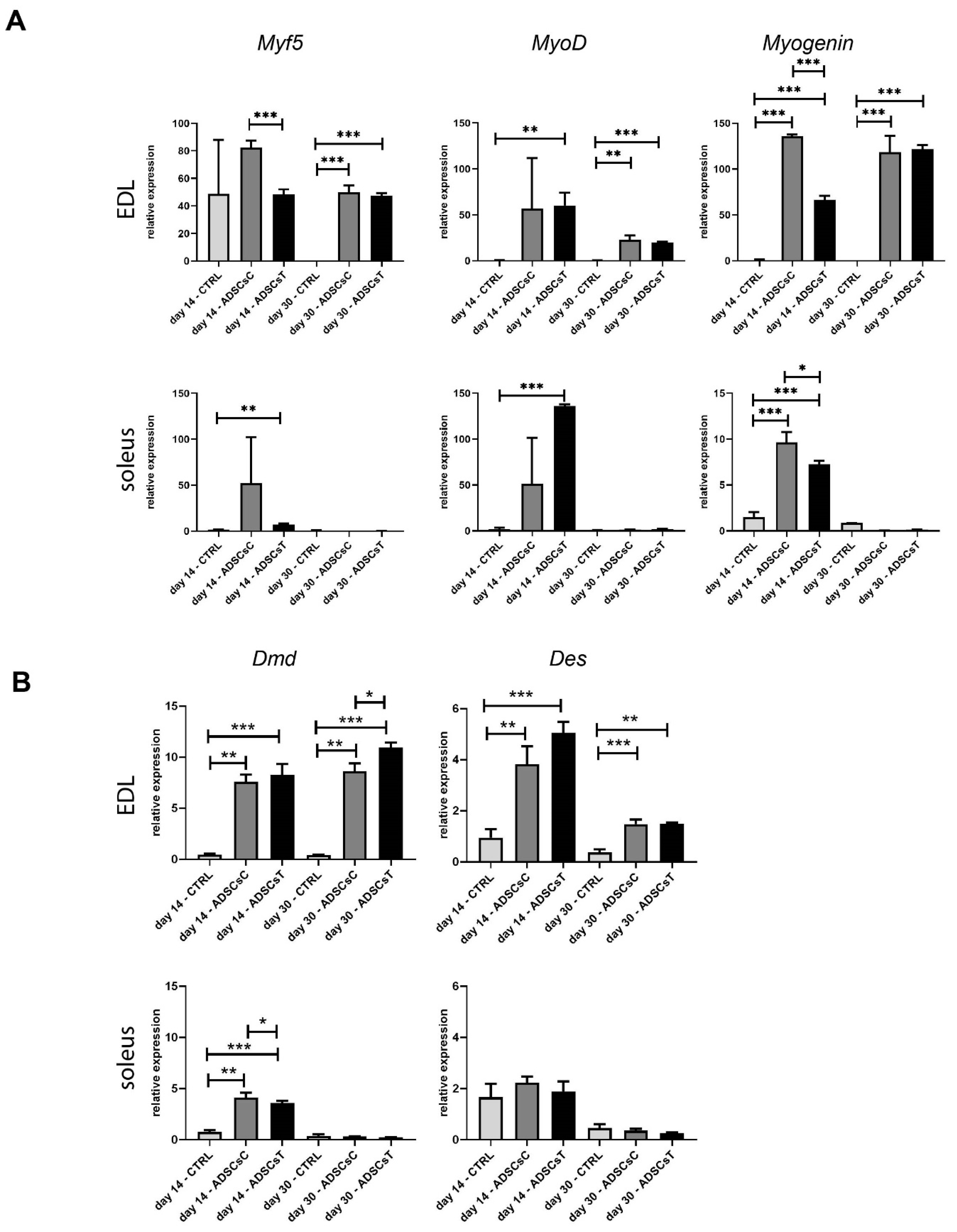
2.2. Rat ADSC Impact at the Structure and Molecular Signature of Regenerating Skeletal Muscle
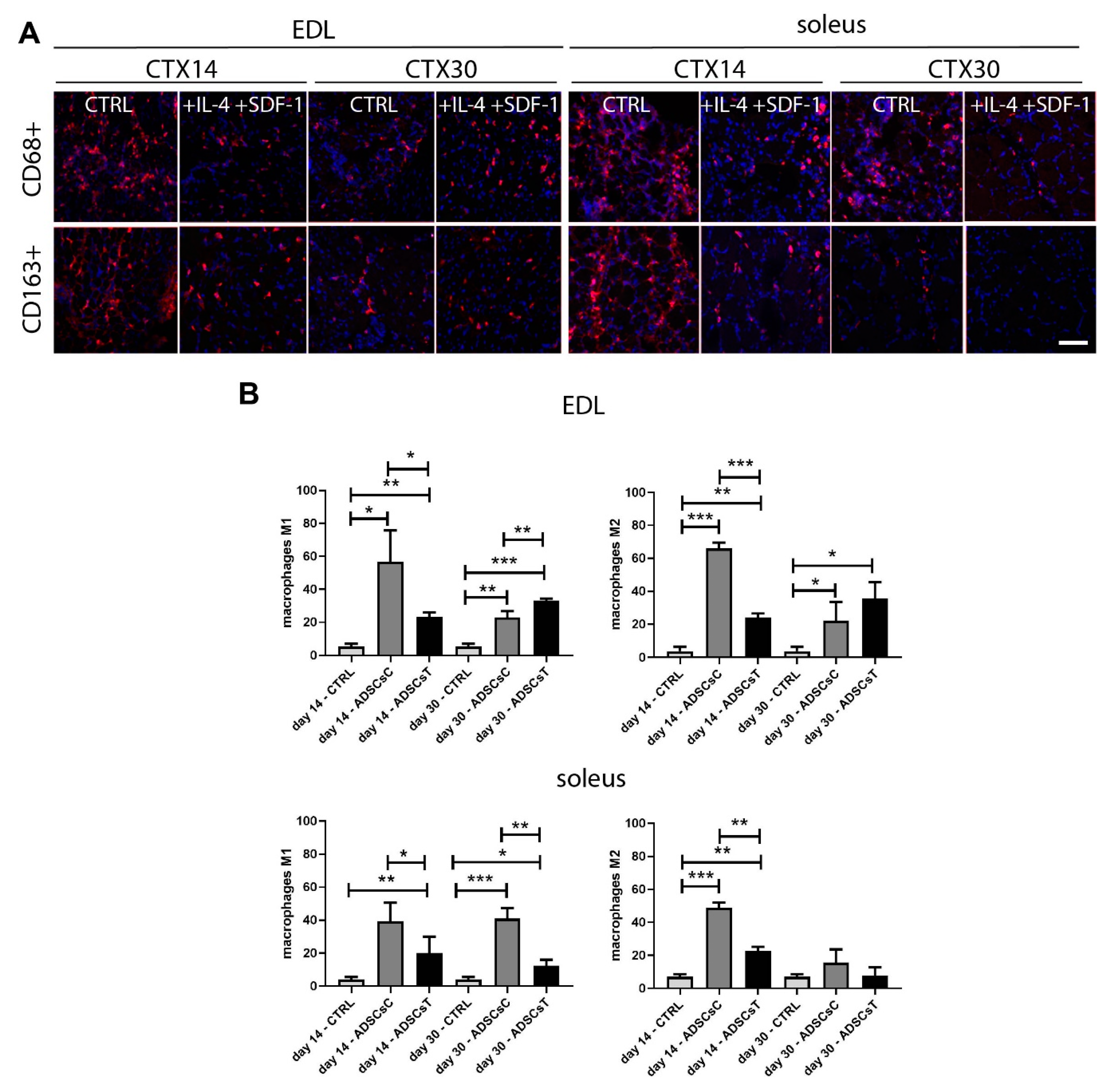
2.3. Rat ADSCs Pre-treated with IL-4 and SDF-1 Modulate Immune Response in Regenerating Skeletal Muscle
3. Discussion
4. Materials and Methods
4.1. Cell Isolation and Culture
4.2. Cell Treatment with IL-4, SDF-1, and IL-4 and SDF-1
4.3. Migration Assay
4.4. Co-culture of ADSCs with C2C12 Myoblasts
4.5. Preparation of Cells for Transplantation
4.6. Skeletal Muscle Injury
4.7. Footprint Area Measurements
4.8. Histological Analyses—Myofiber Number and Connective Tissue Area
4.9. qPCR
4.10. Immunolocalization
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADSC | adipose tissue-derived stromal cell |
| ECM | extracellular matrix |
| IL-4 | interleukin-4 |
| TGFβ | transforming growth factor beta |
| SDF-1 | stromal cell-derived factor-1 |
References
- Grabowska, I.; Archacka, K.; Czerwinska, A.M.; Krupa, M.; Ciemerych, M.A. Mouse and human pluripotent stem cells and the means of their myogenic differentiation. Results Probl. Cell Differ. 2012, 55, 321–356. [Google Scholar] [PubMed]
- Zammit, P.S. Function of the myogenic regulatory factors Myf5, MyoD, Myogenin and MRF4 in skeletal muscle, satellite cells and regenerative myogenesis. Semin. Cell Dev. Biol. 2017, 72, 19–32. [Google Scholar] [CrossRef]
- Li, H.; Capetanaki, Y. Regulation of the mouse desmin gene: Transactivated by MyoD, myogenin, MRF4 and Myf5. Nucleic Acids Res. 1993, 21, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Chazaud, B. Inflammation during skeletal muscle regeneration and tissue remodeling: Application to exercise-induced muscle damage management. Immunol. Cell Biol. 2016, 94, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Tidball, J.G. Regulation of muscle growth and regeneration by the immune system. Nat. Rev. Immunol. 2017, 17, 165–178. [Google Scholar] [CrossRef]
- Bassaglia, Y.; Gautron, J. Fast and slow rat muscles degenerate and regenerate differently after whole crush injury. J. Muscle Res. Cell Motil. 1995, 16, 420–429. [Google Scholar] [CrossRef]
- Noirez, P.; Agbulut, O.; Ferry, A. Differential modification of myosin heavy chain expression by tenotomy in regenerating fast and slow muscles of the rat. Exp. Physiol. 2000, 85, 187–191. [Google Scholar] [CrossRef]
- Brzoska, E.; Ciemerych, M.A.; Przewozniak, M.; Zimowska, M. Regulation of muscle stem cells activation: The role of growth factors and extracellular matrix. Vitam. Horm. 2011, 87, 239–276. [Google Scholar]
- Karalaki, M.; Fili, S.; Philippou, A.; Koutsilieris, M. Muscle regeneration: Cellular and molecular events. In Vivo 2009, 23, 779–796. [Google Scholar]
- Mahdy, M.A.A. Skeletal muscle fibrosis: An overview. Cell Tissue Res. 2019, 375, 575–588. [Google Scholar] [CrossRef]
- Moyer, A.L.; Wagner, K.R. Regeneration versus fibrosis in skeletal muscle. Curr. Opin. Rheumatol. 2011, 23, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Grabowska, I.; Zimowska, M.; Maciejewska, K.; Jablonska, Z.; Bazga, A.; Ozieblo, M.; Streminska, W.; Bem, J.; Brzoska, E.; Ciemerych, M.A. Adipose Tissue-Derived Stromal Cells in Matrigel Impacts the Regeneration of Severely Damaged Skeletal Muscles. Int. J. Mol. Sci. 2019, 20, 3313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimowska, M.; Duchesnay, A.; Dragun, P.; Oberbek, A.; Moraczewski, J.; Martelly, I. Immunoneutralization of TGFbeta1 Improves Skeletal Muscle Regeneration: Effects on Myoblast Differentiation and Glycosaminoglycan Content. Int. J. Cell Biol. 2009, 2009, 659372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.M.; Kim, A.Y.; Lee, E.J.; Park, J.K.; Lee, M.M.; Hwang, M.; Kim, C.Y.; Kim, S.Y.; Jeong, K.S. Therapeutic effects of mouse adipose-derived stem cells and losartan in the skeletal muscle of injured mdx mice. Cell Transplant. 2015, 24, 939–953. [Google Scholar] [CrossRef] [Green Version]
- Zimowska, M.; Olszynski, K.H.; Swierczynska, M.; Streminska, W.; Ciemerych, M.A. Decrease of MMP-9 Activity Improves Soleus Muscle Regeneration. Tissue Eng. 2012, 18, 1183–1192. [Google Scholar] [CrossRef] [Green Version]
- Zimowska, M.; Kasprzycka, P.; Bocian, K.; Delaney, K.; Jung, P.; Kuchcinska, K.; Kaczmarska, K.; Gladysz, D.; Streminska, W.; Ciemerych, M.A. Inflammatory response during slow- and fast-twitch muscle regeneration. Muscle Nerve. 2017, 55, 400–409. [Google Scholar] [CrossRef]
- Almada, A.E.; Wagers, A.J. Molecular circuitry of stem cell fate in skeletal muscle regeneration, ageing and disease. Nat. Rev. Mol. Cell. Biol. 2016, 17, 267–279. [Google Scholar] [CrossRef] [Green Version]
- Rando, T.A.; Blau, H.M. Primary mouse myoblast purification, characterization, and transplantation for cell-mediated gene therapy. J. Cell. Biol. 1994, 125, 1275–1287. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Maley, M.; Beilharz, M.; Grounds, M. Rapid death of injected myoblasts in myoblast transfer therapy. Muscle Nerve. 1996, 19, 853–860. [Google Scholar] [CrossRef]
- Skuk, D.; Caron, N.J.; Goulet, M.; Roy, B.; Tremblay, J.P. Resetting the problem of cell death following muscle-derived cell transplantation: Detection, dynamics and mechanisms. J. Neuropathol. Exp. Neurol. 2003, 62, 951–967. [Google Scholar] [CrossRef] [Green Version]
- Morosetti, R.; Gidaro, T.; Broccolini, A.; Gliubizzi, C.; Sancricca, C.; Tonali, P.A.; Ricci, E.; Mirabella, M. Mesoangioblasts from facioscapulohumeral muscular dystrophy display in vivo a variable myogenic ability predictable by their in vitro behavior. Cell Transplant. 2011, 20, 1299–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotini, A.; Martinez-Sarra, E.; Duelen, R.; Costamagna, D.; Di Filippo, E.S.; Giacomazzi, G.; Grosemans, H.; Fulle, S.; Sampaolesi, M. Aging affects the in vivo regenerative potential of human mesoangioblasts. Aging Cell 2018, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quattrocelli, M.; Costamagna, D.; Giacomazzi, G.; Camps, J.; Sampaolesi, M. Notch signaling regulates myogenic regenerative capacity of murine and human mesoangioblasts. Cell Death Dis. 2014, 5, e1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellavalle, A.; Maroli, G.; Covarello, D.; Azzoni, E.; Innocenzi, A.; Perani, L.; Antonini, S.; Sambasivan, R.; Brunelli, S.; Tajbakhsh, S.; et al. Pericytes resident in postnatal skeletal muscle differentiate into muscle fibres and generate satellite cells. Nat. Commun. 2011, 2, 499. [Google Scholar] [CrossRef] [Green Version]
- Dellavalle, A.; Sampaolesi, M.; Tonlorenzi, R.; Tagliafico, E.; Sacchetti, B.; Perani, L.; Innocenzi, A.; Galvez, B.G.; Messina, G.; Morosetti, R.; et al. Pericytes of human skeletal muscle are myogenic precursors distinct from satellite cells. Nat. Cell. Biol. 2007, 9, 255–267. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, K.J.; Pannerec, A.; Cadot, B.; Parlakian, A.; Besson, V.; Gomes, E.R.; Marazzi, G.; Sassoon, D.A. Identification and characterization of a non-satellite cell muscle resident progenitor during postnatal development. Nat. Cell. Biol. 2010, 12, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.K.; Hall, J.K.; Troy, A.A.; Cornelison, D.D.; Majka, S.M.; Olwin, B.B. Syndecan-4-expressing muscle progenitor cells in the SP engraft as satellite cells during muscle regeneration. Cell Stem Cell 2009, 4, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Qu-Petersen, Z.; Cao, B.; Kimura, S.; Jankowski, R.; Cummins, J.; Usas, A.; Gates, C.; Robbins, P.; Wernig, A.; et al. Clonal isolation of muscle-derived cells capable of enhancing muscle regeneration and bone healing. J. Cell. Biol. 2000, 150, 1085–1100. [Google Scholar] [CrossRef] [Green Version]
- Mierzejewski, B.; Archacka, K.; Grabowska, I.; Florkowska, A.; Ciemerych, M.A.; Brzoska, E. Human and mouse skeletal muscle stem and progenitor cells in health and disease. Semin. Cell Dev. Biol. 2020. [Google Scholar] [CrossRef]
- Tatullo, M.; Codispoti, B.; Pacifici, A.; Palmieri, F.; Marrelli, M.; Pacifici, L.; Paduano, F. Potential Use of Human Periapical Cyst-Mesenchymal Stem Cells (hPCy-MSCs) as a Novel Stem Cell Source for Regenerative Medicine Applications. Front. Cell Dev. Biol. 2017, 5, 103. [Google Scholar] [CrossRef] [Green Version]
- Tatullo, M.; Falisi, G.; Amantea, M.; Rastelli, C.; Paduano, F.; Marrelli, M. Dental Pulp Stem Cells and Human Periapical Cyst Mesenchymal Stem Cells in Bone Tissue Regeneration: Comparison of Basal and Osteogenic Differentiated Gene Expression of a Newly Discovered Mesenchymal Stem Cell Lineage. J. Biol. Regul. Homeost. Agents 2015, 29, 713–718. [Google Scholar] [PubMed]
- Bobis, S.; Jarocha, D.; Majka, M. Mesenchymal stem cells: Characteristics and clinical applications. Folia. Histochem. Cytobiol. 2006, 44, 215–230. [Google Scholar] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Horwitz, E.M.; Le Blanc, K.; Dominici, M.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Deans, R.J.; Krause, D.S.; Keating, A. Clarification of the nomenclature for MSC: The International Society for Cellular Therapy position statement. Cytotherapy 2005, 7, 393–395. [Google Scholar] [CrossRef]
- Sacchetti, B.; Funari, A.; Remoli, C.; Giannicola, G.; Kogler, G.; Liedtke, S.; Cossu, G.; Serafini, M.; Sampaolesi, M.; Tagliafico, E.; et al. No Identical “Mesenchymal Stem Cells” at Different Times and Sites: Human Committed Progenitors of Distinct Origin and Differentiation Potential Are Incorporated as Adventitial Cells in Microvessels. Stem Cell Rep. 2016, 6, 897–913. [Google Scholar] [CrossRef] [Green Version]
- Halvorsen, Y.C.; Wilkison, W.O.; Gimble, J.M. Adipose-derived stromal cells--their utility and potential in bone formation. Int. J. Obes. Relat. Metab. Disord. 2000, 24, S41–S44. [Google Scholar] [CrossRef] [Green Version]
- Gronthos, S.; Franklin, D.M.; Leddy, H.A.; Robey, P.G.; Storms, R.W.; Gimble, J.M. Surface protein characterization of human adipose tissue-derived stromal cells. J. Cell Physiol. 2001, 189, 54–63. [Google Scholar] [CrossRef]
- Halvorsen, Y.D.; Bond, A.; Sen, A.; Franklin, D.M.; Lea-Currie, Y.R.; Sujkowski, D.; Ellis, P.N.; Wilkison, W.O.; Gimble, J.M. Thiazolidinediones and glucocorticoids synergistically induce differentiation of human adipose tissue stromal cells: Biochemical, cellular, and molecular analysis. Metabolism 2001, 50, 407–413. [Google Scholar] [CrossRef]
- Sen, A.; Lea-Currie, Y.R.; Sujkowska, D.; Franklin, D.M.; Wilkison, W.O.; Halvorsen, Y.D.; Gimble, J.M. Adipogenic potential of human adipose derived stromal cells from multiple donors is heterogeneous. J. Cell. Biochem. 2001, 81, 312–319. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuk, P.A. The Adipose-derived Stem Cell: Looking Back and Looking Ahead. Mol. Biol. Cell. 2010, 21, 1783–1787. [Google Scholar] [CrossRef] [Green Version]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell. 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.B.; McIntosh, K.; Zvonic, S.; Garrett, S.; Floyd, Z.E.; Kloster, A.; Di Halvorsen, Y.; Storms, R.W.; Goh, B.; Kilroy, G.; et al. Immunophenotype of human adipose-derived cells: Temporal changes in stromal-associated and stem cell-associated markers. Stem Cells 2006, 24, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Bourin, P.; Bunnell, B.A.; Casteilla, L.; Dominici, M.; Katz, A.J.; March, K.L.; Redl, H.; Rubin, J.P.; Yoshimura, K.; Gimble, J.M. Stromal cells from the adipose tissue-derived stromal vascular fraction and culture expanded adipose tissue-derived stromal/stem cells: A joint statement of the International Federation for Adipose Therapeutics and Science (IFATS) and the International Society for Cellular Therapy (ISCT). Cytotherapy 2013, 15, 641–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilak, F.; Lott, K.E.; Awad, H.A.; Cao, Q.; Hicok, K.C.; Fermor, B.; Gimble, J.M. Clonal analysis of the differentiation potential of human adipose-derived adult stem cells. J. Cell. Physiol. 2006, 206, 229–237. [Google Scholar] [CrossRef]
- Mitchell, R.; Mellows, B.; Sheard, J.; Antonioli, M.; Kretz, O.; Chambers, D.; Zeuner, M.T.; Tomkins, J.E.; Denecke, B.; Musante, L.; et al. Secretome of adipose-derived mesenchymal stem cells promotes skeletal muscle regeneration through synergistic action of extracellular vesicle cargo and soluble proteins. Stem Cell Res. Ther. 2019, 10, 116. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, Y.; Li, Y.; Cao, J.; Zhang, H.; Chen, M.; Wang, L.; Zhang, C. Long-term engraftment of myogenic progenitors from adipose-derived stem cells and muscle regeneration in dystrophic mice. Hum. Mol. Genet. 2015, 24, 6029–6040. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, K.; Archacki, R.; Archacka, K.; Streminska, W.; Paciorek, A.; Golabek, M.; Ciemerych, M.A.; Brzoska, E. Stromal derived factor-1 and granulocyte-colony stimulating factor treatment improves regeneration of Pax7-/- mice skeletal muscles. J. Cachexia. Sarcopenia. Muscle 2016, 7, 483–496. [Google Scholar] [CrossRef]
- Brzoska, E.; Kowalski, K.; Markowska-Zagrajek, A.; Kowalewska, M.; Archacki, R.; Plaskota, I.; Streminska, W.; Janczyk-Ilach, K.; Ciemerych, M.A. Sdf-1 (CXCL12) induces CD9 expression in stem cells engaged in muscle regeneration. Stem Cell Res. Ther. 2015, 6, 46. [Google Scholar] [CrossRef] [Green Version]
- Grabowska, I.; Brzoska, E.; Gawrysiak, A.; Streminska, W.; Moraczewski, J.; Polanski, Z.; Hoser, G.; Kawiak, J.; Machaj, E.K.; Pojda, Z.; et al. Restricted Myogenic Potential of Mesenchymal Stromal Cells Isolated From Umbilical Cord. Cell Transplant. 2012, 21, 1711–1726. [Google Scholar] [CrossRef] [PubMed]
- Grabowska, I.; Streminska, W.; Janczyk-Ilach, K.; Machaj, E.K.; Pojda, Z.; Hoser, G.; Kawiak, J.; Moraczewski, J.; Ciemerych, M.A.; Brzoska, E. Myogenic potential of mesenchymal stem cells-the case of adhesive fraction of human umbilical cord blood cells. Curr. Stem Cell Res. Ther. 2013, 8, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Swierczek-Lasek, B.; Neska, J.; Kominek, A.; Tolak, L.; Czajkowski, T.; Janczyk-Ilach, K.; Streminska, W.; Piwocka, K.; Ciemerych, M.A.; Archacka, K. Interleukin 4 Moderately Affects Competence of Pluripotent Stem Cells for Myogenic Conversion. Int. J. Mol. Sci. 2019, 20, 3932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horsley, V.; Jansen, K.M.; Mills, S.T.; Pavlath, G.K. IL-4 acts as a myoblast recruitment factor during mammalian muscle growth. Cell 2003, 113, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Schulze, M.; Belema-Bedada, F.; Technau, A.; Braun, T. Mesenchymal stem cells are recruited to striated muscle by NFAT/IL-4-mediated cell fusion. Genes Dev. 2005, 19, 1787–1798. [Google Scholar] [CrossRef] [Green Version]
- Schleimer, R.P.; Sterbinsky, S.A.; Kaiser, J.; Bickel, C.A.; Klunk, D.A.; Tomioka, K.; Newman, W.; Luscinskas, F.W.; Gimbrone, M.A., Jr.; McIntyre, B.W.; et al. IL-4 induces adherence of human eosinophils and basophils but not neutrophils to endothelium. Association with expression of VCAM-1. J. Immunol. 1992, 148, 1086–1092. [Google Scholar]
- Lafreniere, J.F.; Mills, P.; Bouchentouf, M.; Tremblay, J.P. Interleukin-4 improves the migration of human myogenic precursor cells in vitro and in vivo. Exp. Cell Res. 2006, 312, 1127–1141. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, K.; Kolodziejczyk, A.; Sikorska, M.H.; Placzkiewicz, J.; Cichosz, P.; Kowalewska, M.; Streminska, W.; Janczyk-Ilach, K.; Koblowska, M.; Fogtman, A.; et al. Stem cells migration during skeletal muscle regeneration-the role of Sdf-1/Cxcr4 and Sdf-1/Cxcr7 axis. Cell Adh. Migr. 2016. [Google Scholar] [CrossRef] [Green Version]
- Zimowska, M.; Brzoska, E.; Swierczynska, M.; Streminska, W.; Moraczewski, J. Distinct patterns of MMP-9 and MMP-2 activity in slow and fast twitch skeletal muscle regeneration in vivo. Int. J. Dev. Biol. 2008, 52, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Zimowska, M.; Szczepankowska, D.; Streminska, W.; Papy, D.; Tournaire, M.C.; Gautron, J.; Barritault, D.; Moraczewski, J.; Martelly, I. Heparan sulfate mimetics modulate calpain activity during rat Soleus muscle regeneration. J. Cell Physiol. 2001, 188, 178–187. [Google Scholar] [CrossRef]
- Zimowska, M.; Constantin, B.; Papy-Garcia, D.; Raymond, G.; Cognard, C.; Caruelle, J.P.; Moraczewski, J.; Martelly, I. Novel glycosaminoglycan mimetic (RGTA, RGD120) contributes to enhance skeletal muscle satellite cell fusion by increasing intracellular Ca2+ and calpain activity. J. Cell Physiol. 2005, 205, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.N.; Ahn, J.S.; Lee, S.G.; Lee, H.S.; Choi, A.M.K.; Yoon, J.H. Integrins alphavbeta5 and alphavbeta6 Mediate IL-4-Induced Collective Migration in Human Airway Epithelial Cells. Am. J. Respir. Cell Mol. Biol. 2018. [Google Scholar] [CrossRef]
- Puchert, M.; Engele, J. The peculiarities of the SDF-1/CXCL12 system: In some cells, CXCR4 and CXCR7 sing solos, in others, they sing duets. Cell Tissue Res. 2014, 355, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Guo, Y.; Chen, F.; Liu, J.; Jin, P. Stromal cell-derived factor-1 promotes human adipose tissue-derived stem cell survival and chronic wound healing. Exp. Ther. Med. 2016, 12, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Costamagna, D.; Costelli, P.; Sampaolesi, M.; Penna, F. Role of Inflammation in Muscle Homeostasis and Myogenesis. Mediat. Inflamm. 2015, 2015, 805172. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, C.H.; de Queiroz, J.C.; Guimaraes-Ferreira, L.; Vitzel, K.F.; Nachbar, R.T.; de Sousa, L.G.; de Souza, A.L., Jr.; Nunes, M.T.; Curi, R. Local injections of adipose-derived mesenchymal stem cells modulate inflammation and increase angiogenesis ameliorating the dystrophic phenotype in dystrophin-deficient skeletal muscle. Stem Cell Rev. 2012, 8, 363–374. [Google Scholar] [CrossRef]
- Olea, F.D.; Locatelli, P.; Hnatiuk, A.; De Lorenzi, A.; Valdivieso, L.; Rocha, E.; Ramirez, R.; Laguens, R.; Crottogini, A. Vascular endothelial growth factor overexpression does not enhance adipose stromal cell-induced protection on muscle damage in critical limb ischemia. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 184–188. [Google Scholar] [CrossRef] [Green Version]
- Kasprzycka, P.; Archacka, K.; Kowalski, K.; Mierzejewski, B.; Zimowska, M.; Grabowska, I.; Piotrowski, M.; Rafalko, M.; Ryzko, A.; Irhashava, A.; et al. The factors present in regenerating muscles impact bone marrow-derived mesenchymal stromal/stem cell fusion with myoblasts. Stem Cell Res. Ther. 2019, 10, 343. [Google Scholar] [CrossRef] [Green Version]
- Wakitani, S.; Saito, T.; Caplan, A.I. Myogenic cells derived from rat bone marrow mesenchymal stem cells exposed to 5-azacytidine. Muscle Nerve. 1995, 18, 1417–1426. [Google Scholar] [CrossRef]
- Gussoni, E.; Soneoka, Y.; Strickland, C.D.; Buzney, E.A.; Khan, M.K.; Flint, A.F.; Kunkel, L.M.; Mulligan, R.C. Dystrophin expression in the mdx mouse restored by stem cell transplantation. Nature 1999, 401, 390–394. [Google Scholar] [CrossRef]
- Fukada, S.; Miyagoe-Suzuki, Y.; Tsukihara, H.; Yuasa, K.; Higuchi, S.; Ono, S.; Tsujikawa, K.; Takeda, S.; Yamamoto, H. Muscle regeneration by reconstitution with bone marrow or fetal liver cells from green fluorescent protein-gene transgenic mice. J. Cell Sci. 2002, 115, 1285–1293. [Google Scholar] [PubMed]
- Brazelton, T.R.; Nystrom, M.; Blau, H.M. Significant differences among skeletal muscles in the incorporation of bone marrow-derived cells. Dev. Biol. 2003, 262, 64–74. [Google Scholar] [CrossRef] [Green Version]
- Abarbanell, A.M.; Coffey, A.C.; Fehrenbacher, J.W.; Beckman, D.J.; Herrmann, J.L.; Weil, B.; Meldrum, D.R. Proinflammatory cytokine effects on mesenchymal stem cell therapy for the ischemic heart. Ann. Thorac. Surg. 2009, 88, 1036–1043. [Google Scholar] [CrossRef] [PubMed]
- Shohara, R.; Yamamoto, A.; Takikawa, S.; Iwase, A.; Hibi, H.; Kikkawa, F.; Ueda, M. Mesenchymal stromal cells of human umbilical cord Wharton’s jelly accelerate wound healing by paracrine mechanisms. Cytotherapy 2012, 14, 1171–1181. [Google Scholar] [CrossRef]
- Heo, S.C.; Jeon, E.S.; Lee, I.H.; Kim, H.S.; Kim, M.B.; Kim, J.H. Tumor necrosis factor-alpha-activated human adipose tissue-derived mesenchymal stem cells accelerate cutaneous wound healing through paracrine mechanisms. J. Invest. Dermatol. 2011, 131, 1559–1567. [Google Scholar] [CrossRef] [Green Version]
- Case, J.; Horvath, T.L.; Howell, J.C.; Yoder, M.C.; March, K.L.; Srour, E.F. Clonal multilineage differentiation of murine common pluripotent stem cells isolated from skeletal muscle and adipose stromal cells. Ann. N. Y. Acad. Sci. 2005, 1044, 183–200. [Google Scholar] [CrossRef]
- Case, J.; Horvath, T.L.; Ballas, C.B.; March, K.L.; Srour, E.F. In vitro clonal analysis of murine pluripotent stem cells isolated from skeletal muscle and adipose stromal cells. Exp. Hematol. 2008, 36, 224–234. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zimowska, M.; Archacka, K.; Brzoska, E.; Bem, J.; Czerwinska, A.M.; Grabowska, I.; Kasprzycka, P.; Michalczewska, E.; Stepaniec, I.; Soszynska, M.; et al. IL-4 and SDF-1 Increase Adipose Tissue-Derived Stromal Cell Ability to Improve Rat Skeletal Muscle Regeneration. Int. J. Mol. Sci. 2020, 21, 3302. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093302
Zimowska M, Archacka K, Brzoska E, Bem J, Czerwinska AM, Grabowska I, Kasprzycka P, Michalczewska E, Stepaniec I, Soszynska M, et al. IL-4 and SDF-1 Increase Adipose Tissue-Derived Stromal Cell Ability to Improve Rat Skeletal Muscle Regeneration. International Journal of Molecular Sciences. 2020; 21(9):3302. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093302
Chicago/Turabian StyleZimowska, Małgorzata, Karolina Archacka, Edyta Brzoska, Joanna Bem, Areta M. Czerwinska, Iwona Grabowska, Paulina Kasprzycka, Emilia Michalczewska, Igor Stepaniec, Marta Soszynska, and et al. 2020. "IL-4 and SDF-1 Increase Adipose Tissue-Derived Stromal Cell Ability to Improve Rat Skeletal Muscle Regeneration" International Journal of Molecular Sciences 21, no. 9: 3302. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093302