Synthesis and Therapeutic Applications of Iminosugars in Cystic Fibrosis

 , , , ,
, , , ,  , and
, and
Abstract
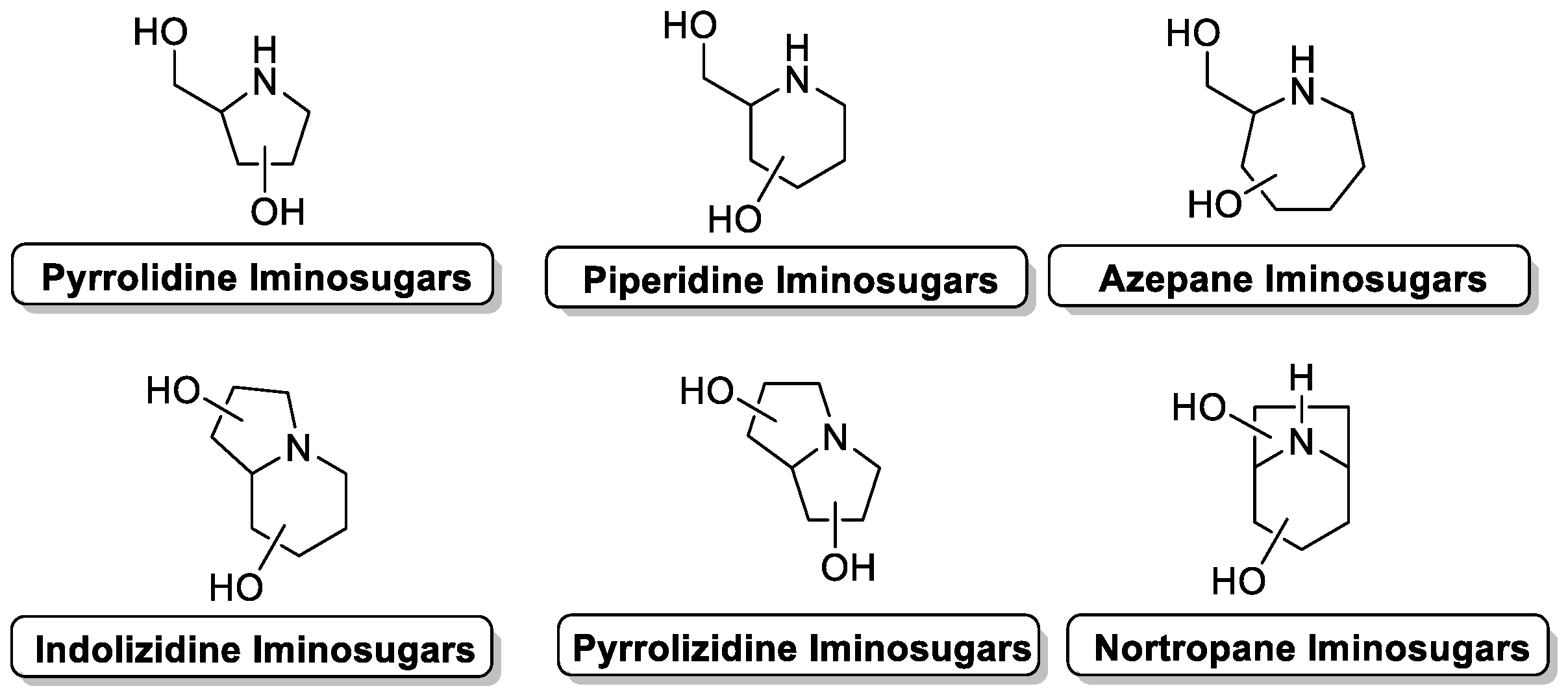
:1. Iminosugars: Powerful Glycomimetics
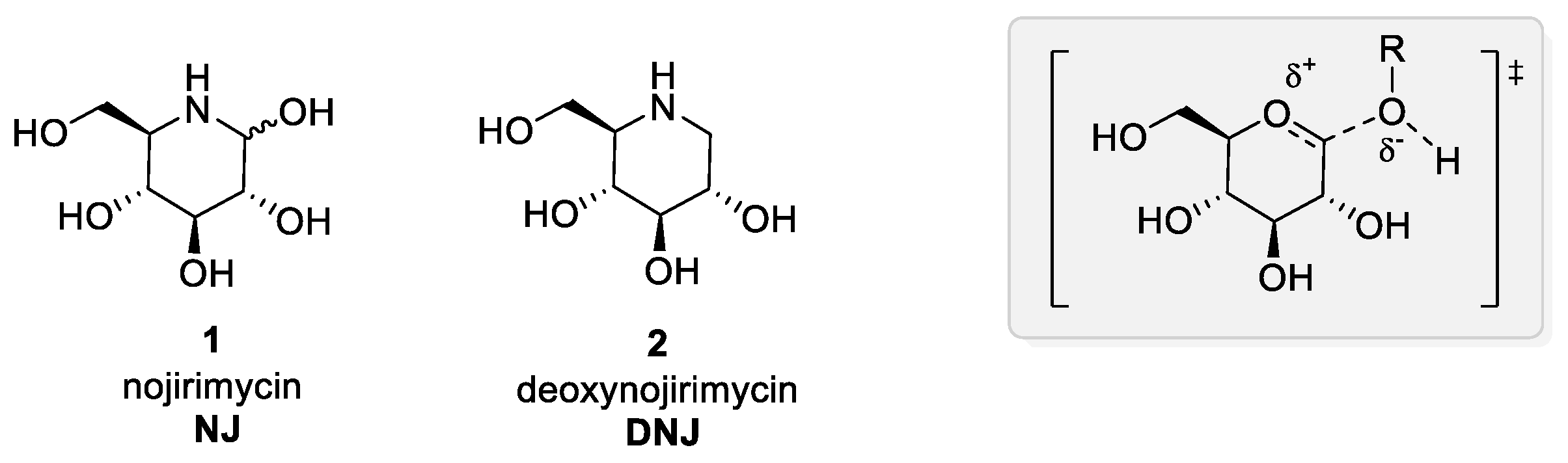
2. Iminosugars in Cystic Fibrosis
3. Rescuing the Activity of Defective CFTR: Iminosugars as Correctors
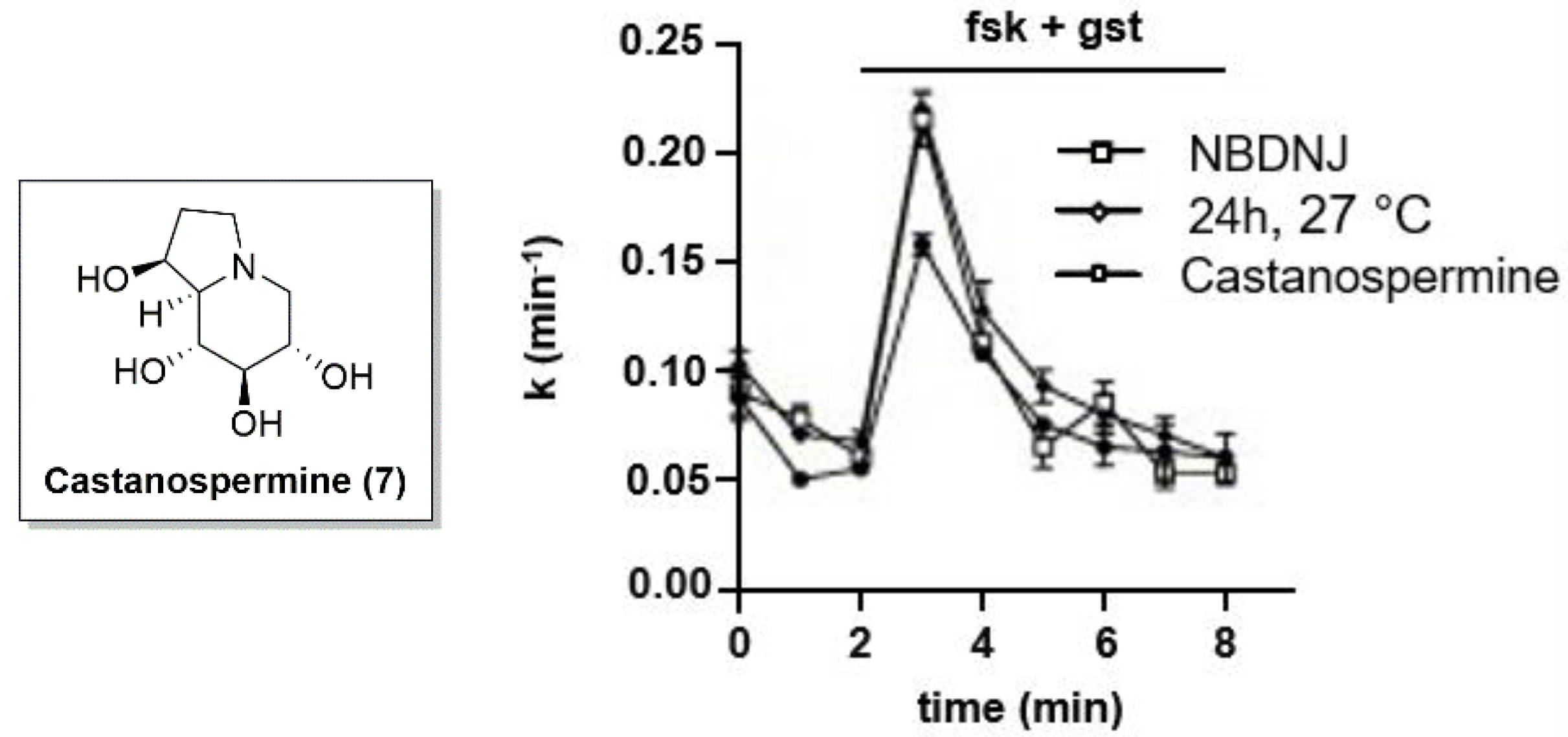
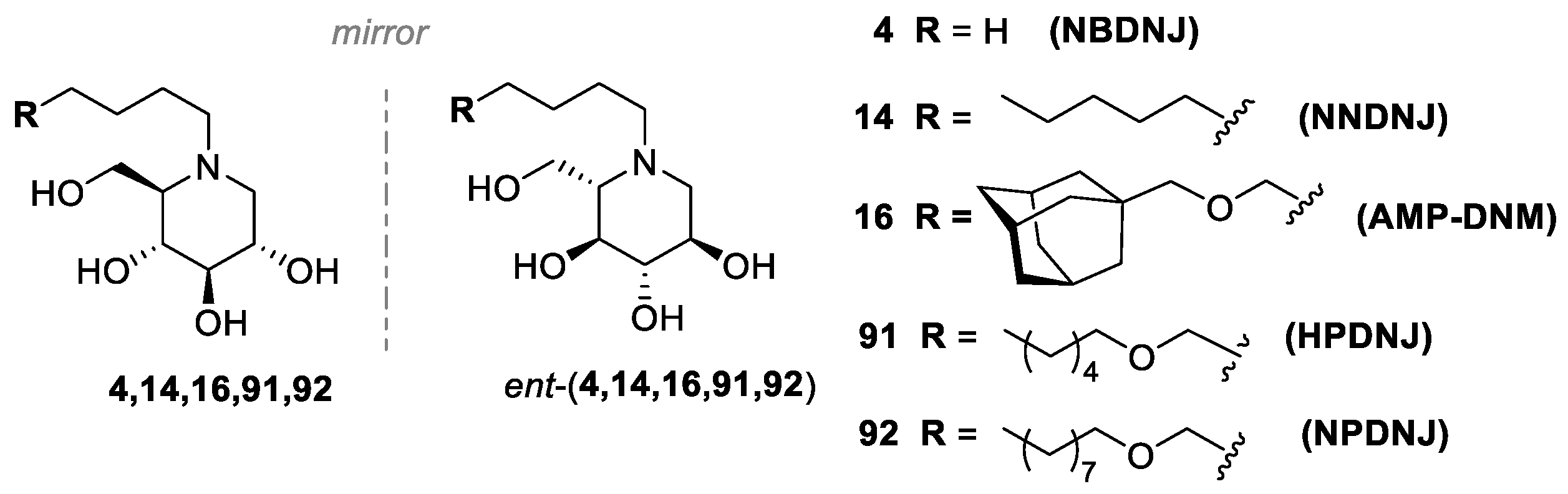
3.1. Iminosugars as CFTR Correctors: NBDNJ and beyond
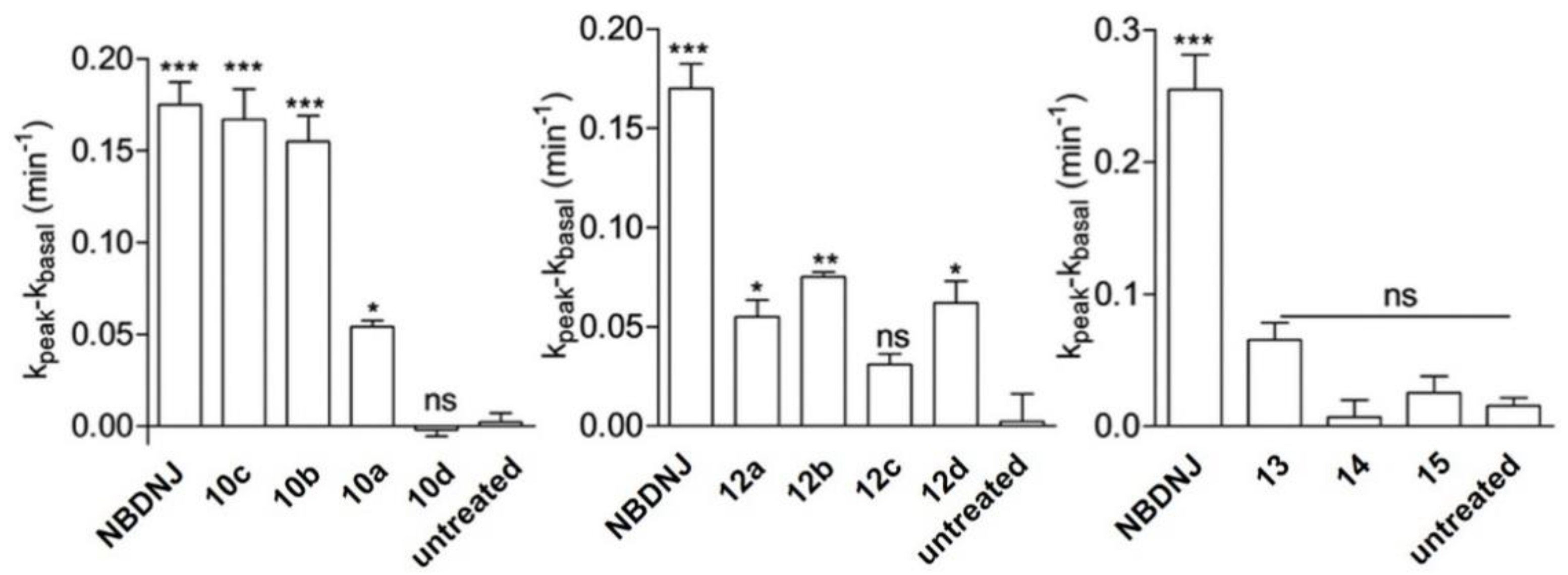
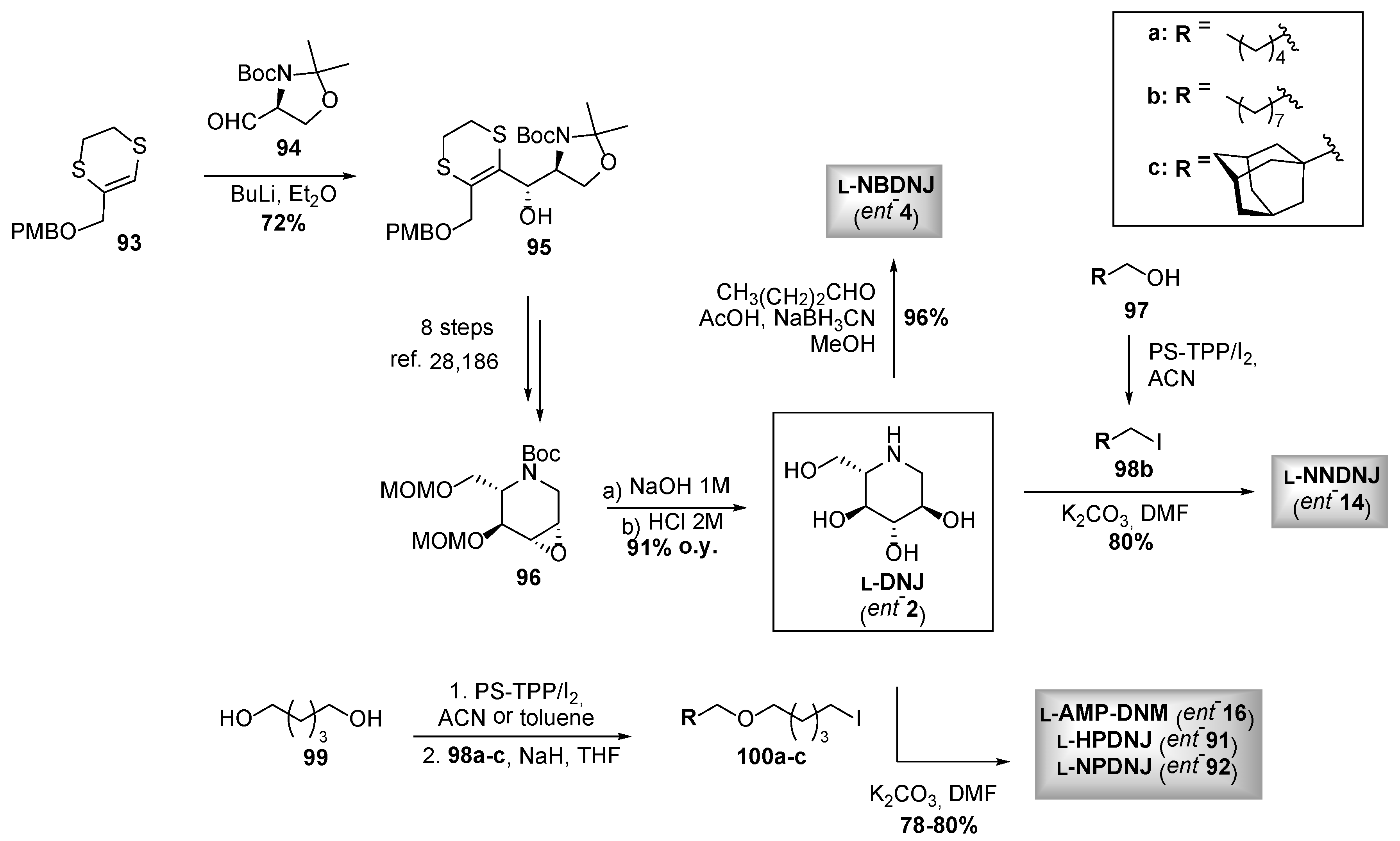
3.2. N-alkyl DNJ Derivatives: Studying the Role of Lipophilicity on the Rescue of F508del-CFTR Activity
3.3. Iminosugar Click Clusters: Multivalent Effect on the Rescue of CFTR Activity.
3.4. Pyrrolidine Iminosugars and Pyrrolidine-Containing Bicyclic Iminosugars as Highly Active and Selective CFTR Correctors
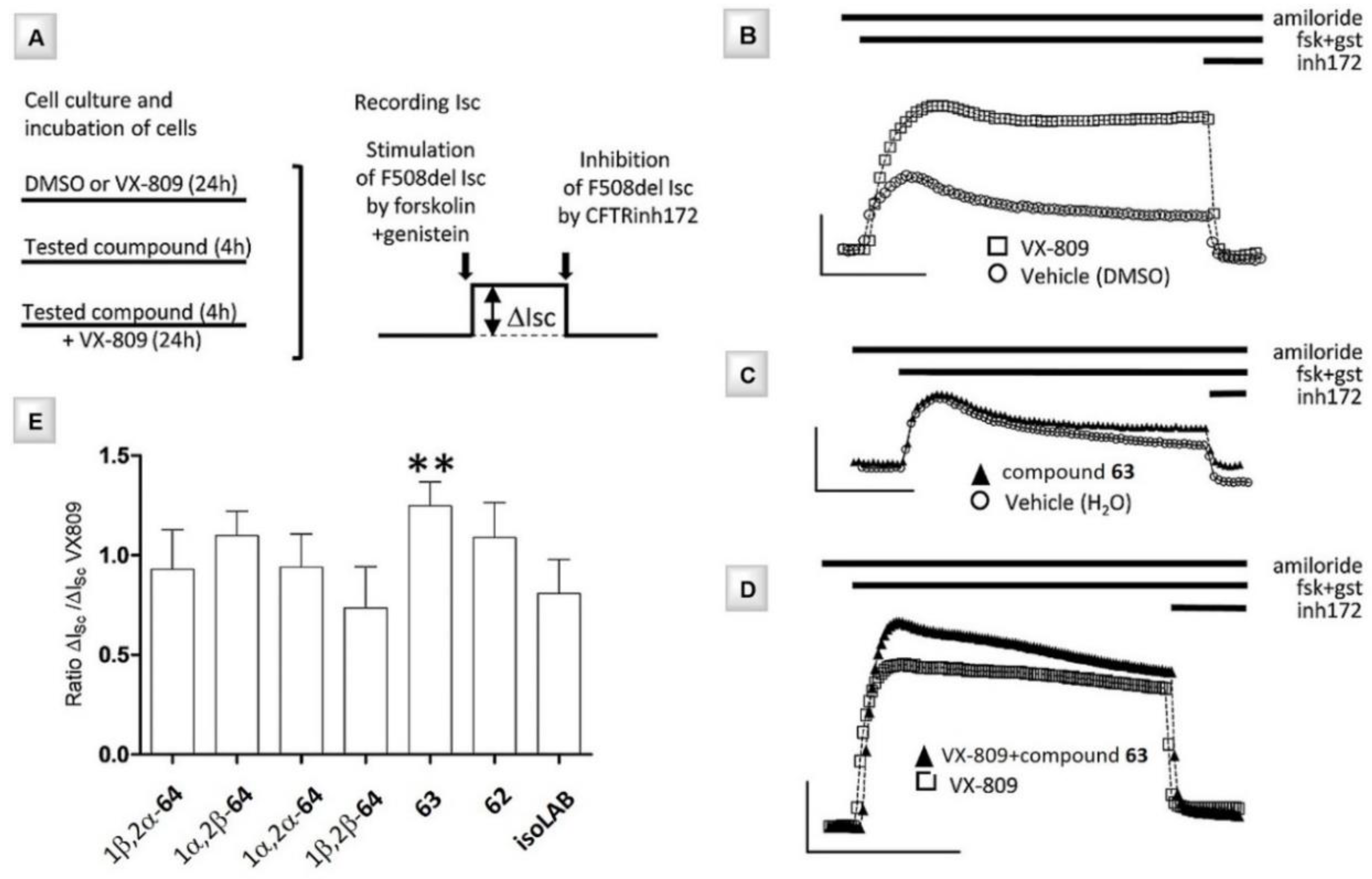
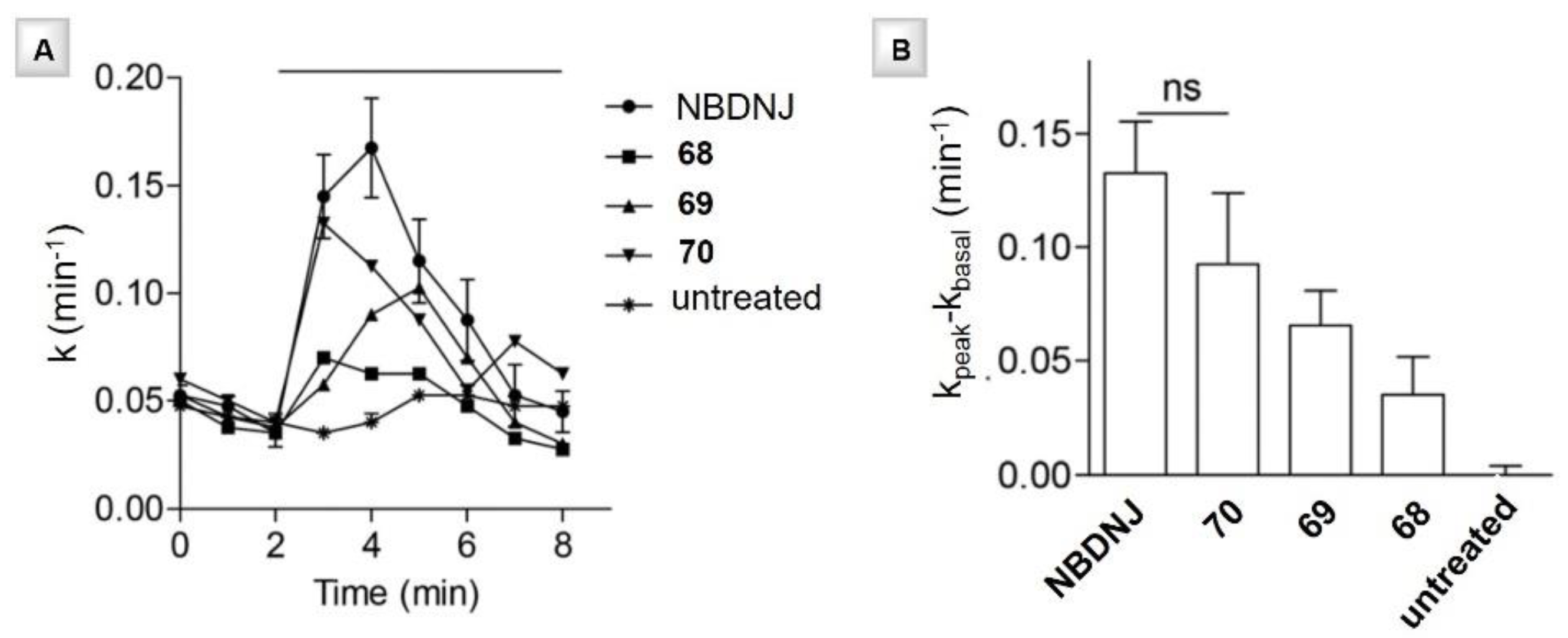
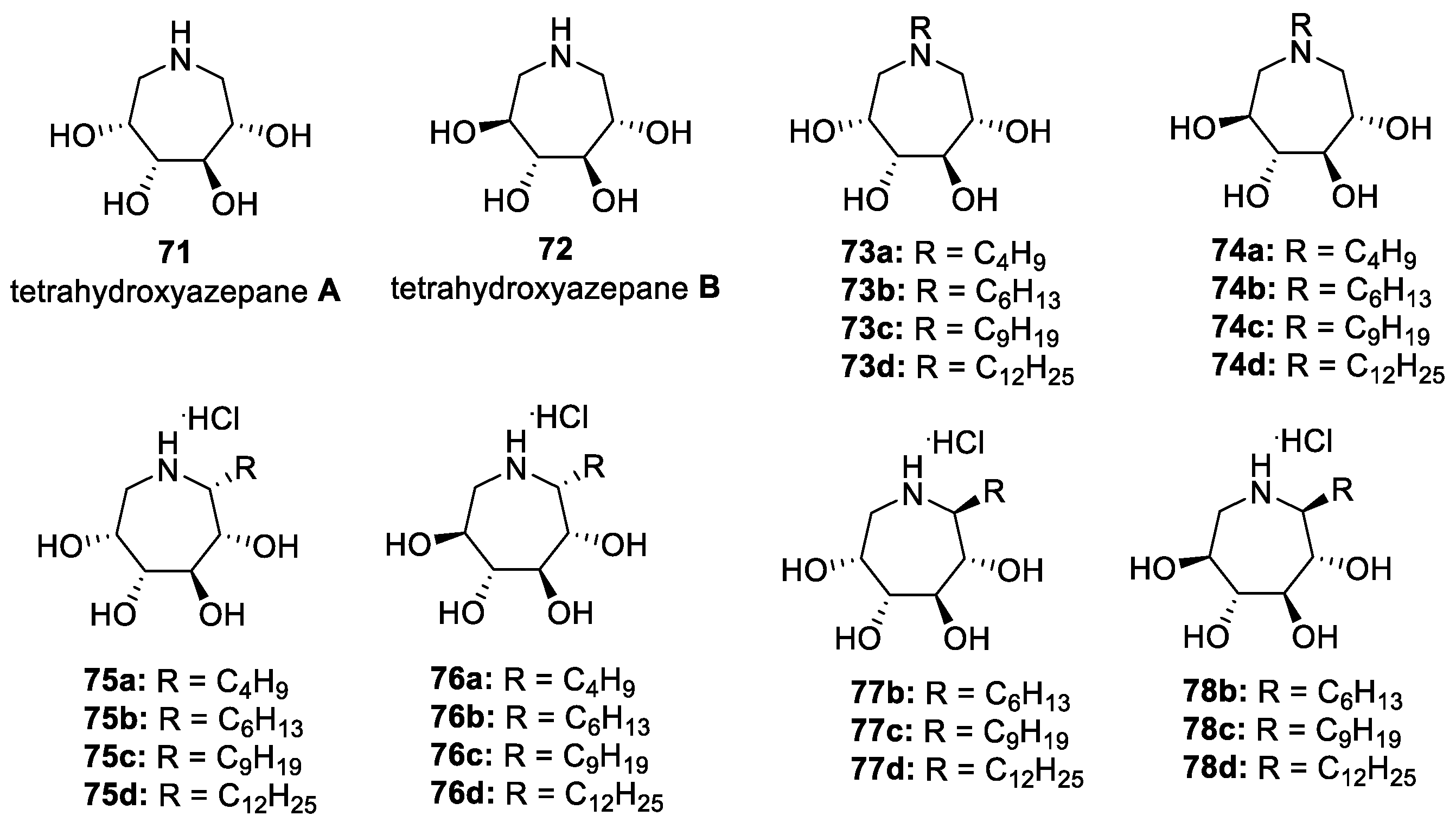
3.5. N- and C-Alkyl Azepane Iminosugars as CFTR Correctors
3.6. Combination Studies of CFTR Correctors
4. Iminosugars as Therapeutic Agents for the Treatment of CF Lung Inflammation
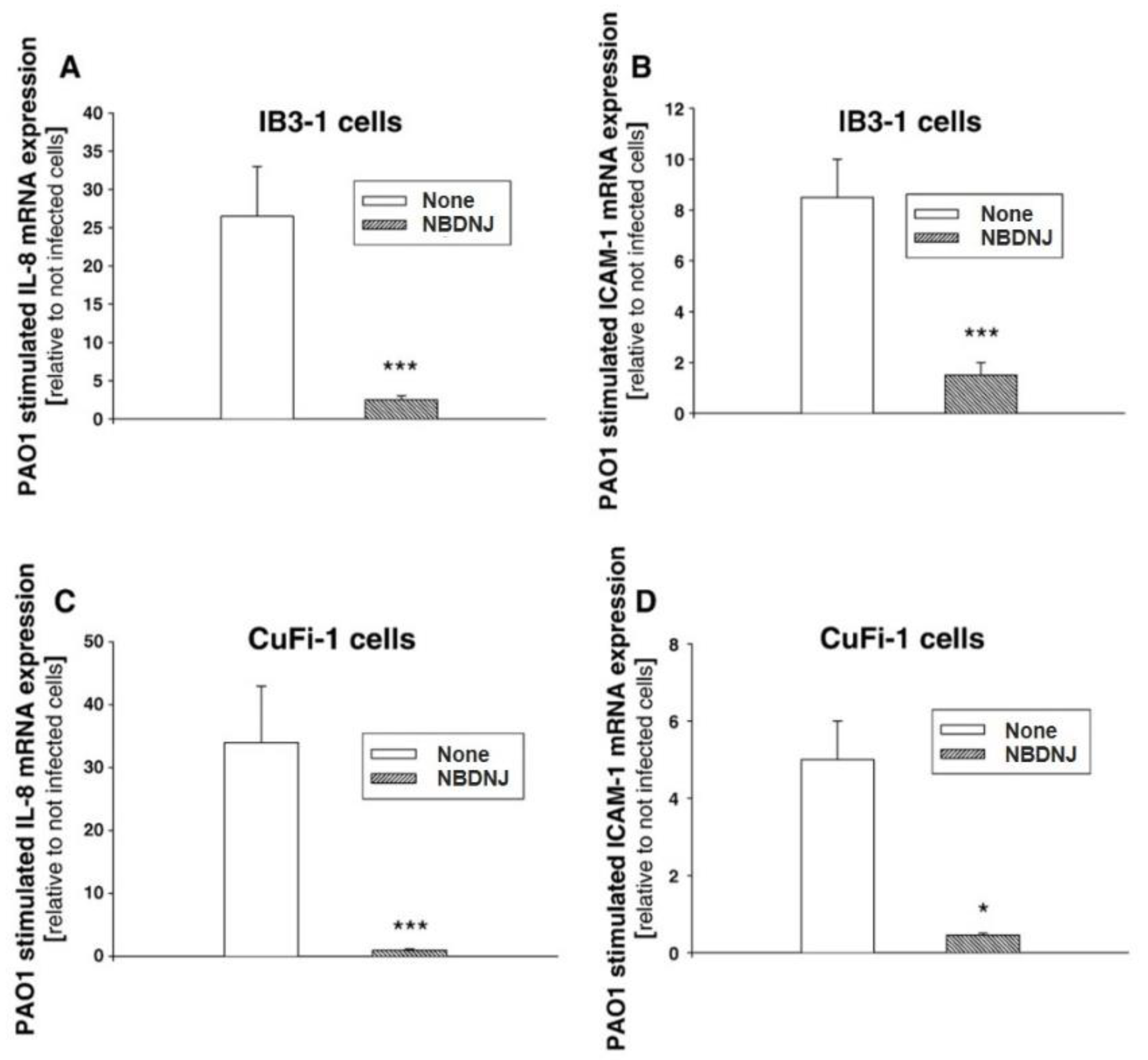
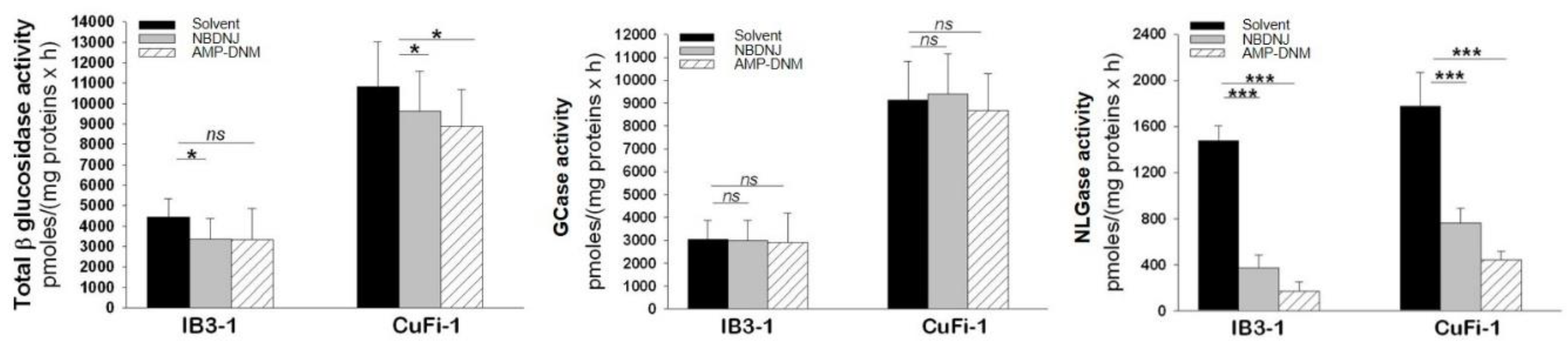
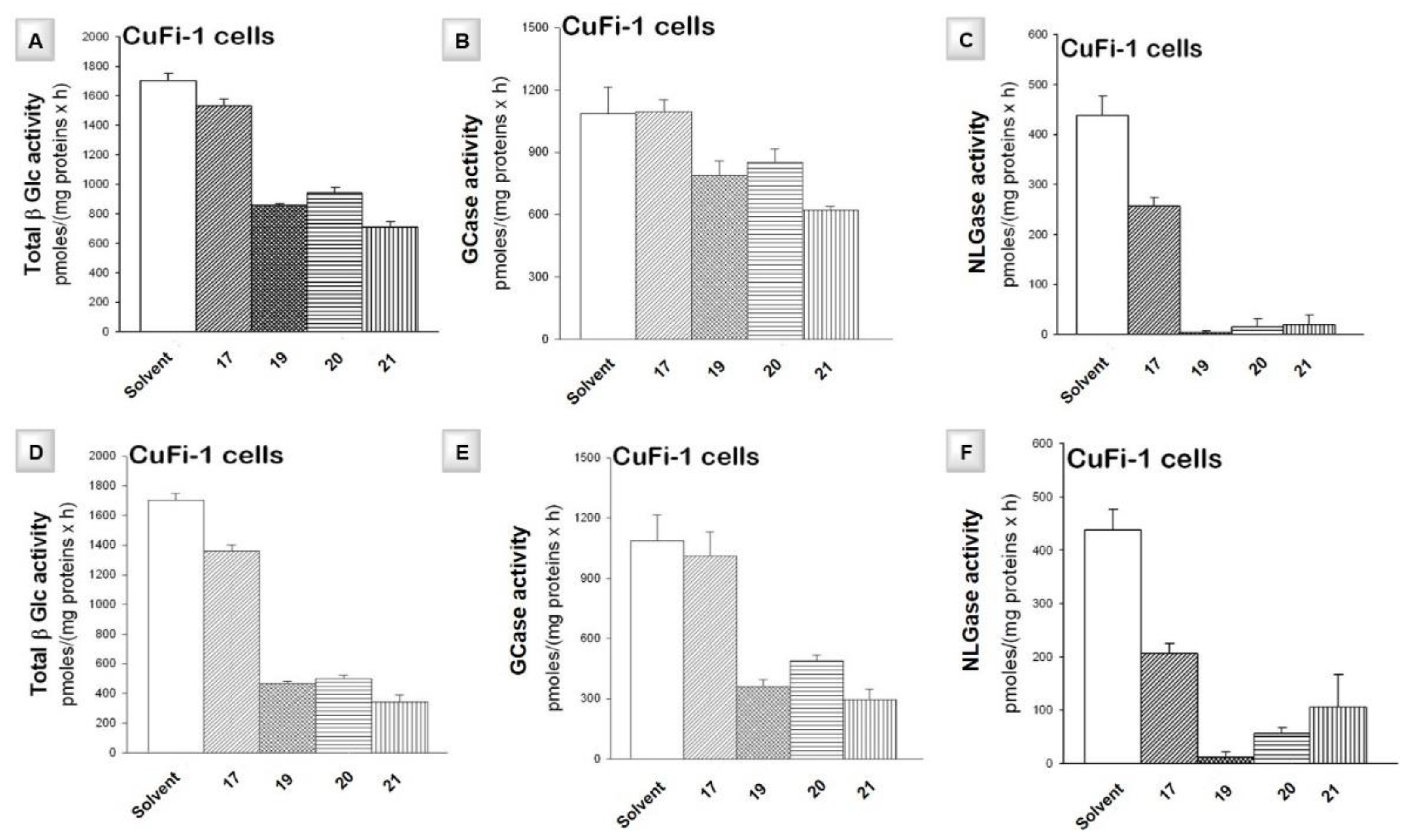
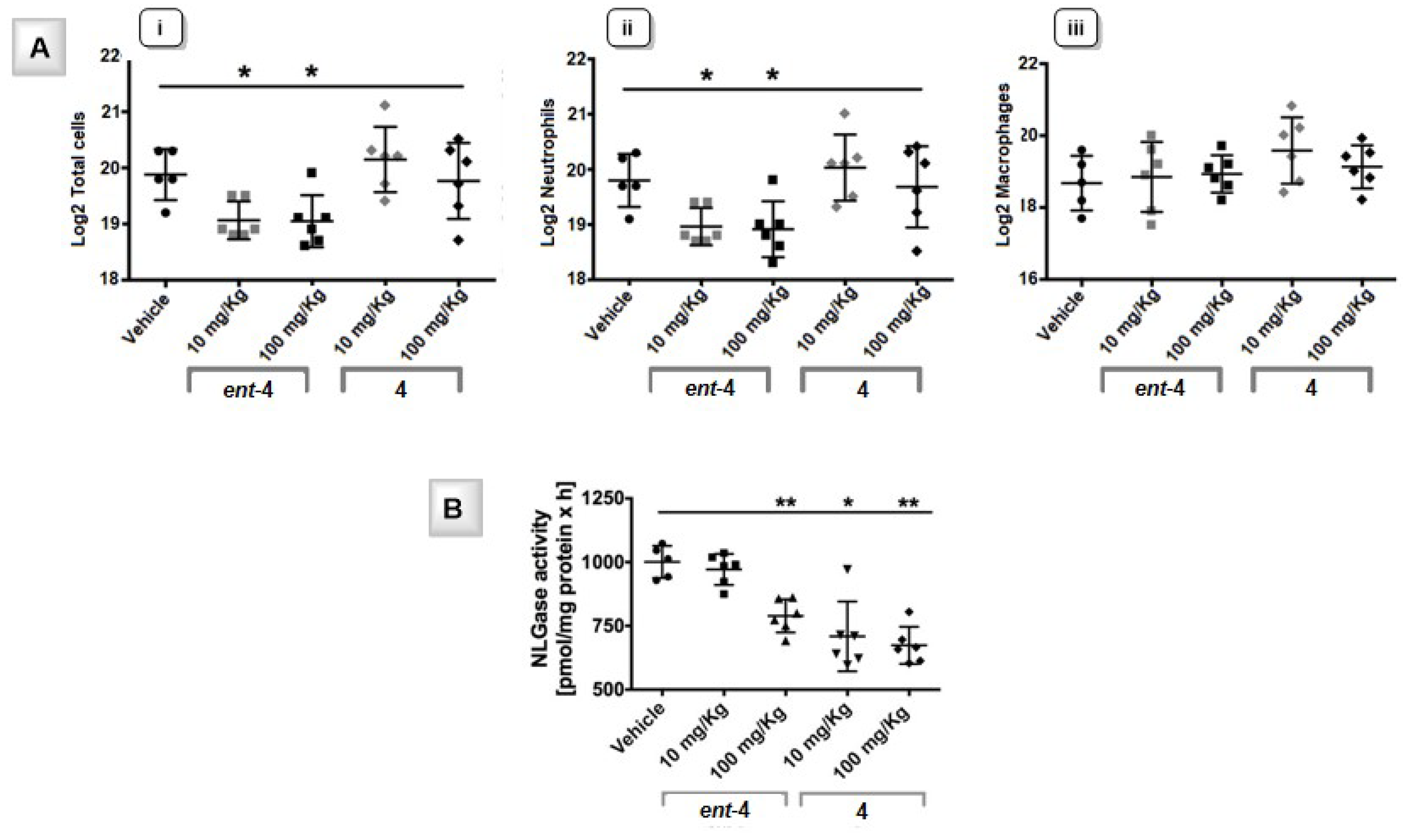
4.1. Effect of Iminosugars on the Inflammatory Response to P. aeruginosa: from NBDNJ to Its N-Alkyl Derivatives
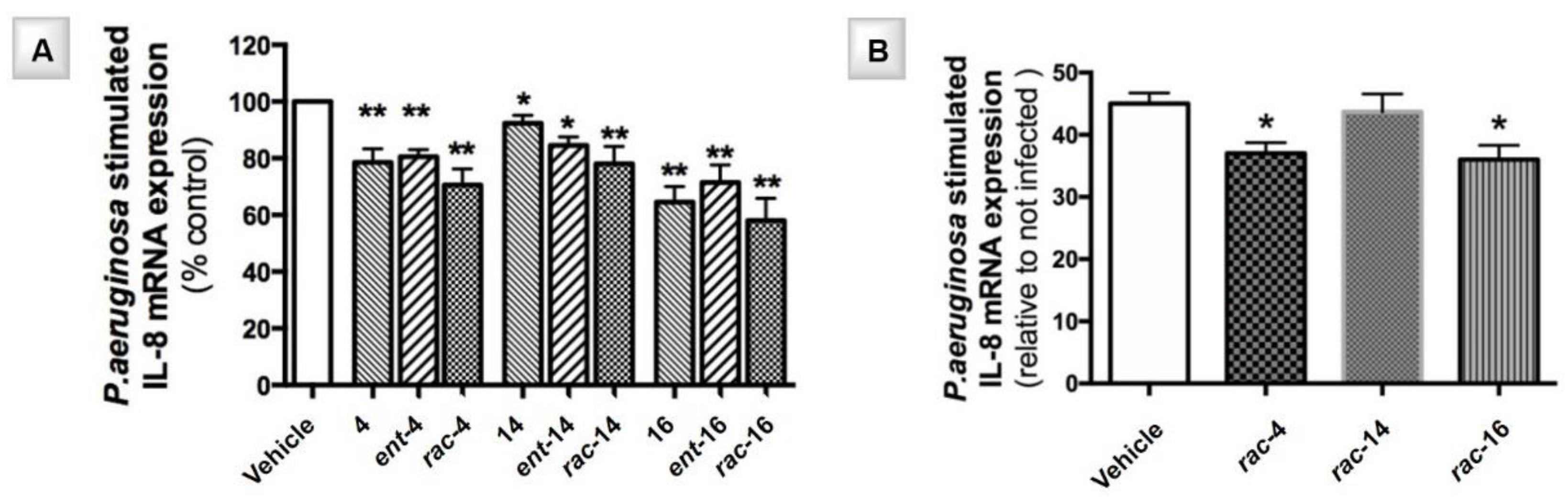
4.2. Exploring the Role of the Chirality in the Anti-Inflamatory Effect of N-alkyl DNJ Derivatives
5. Concluding Remarks
Funding
Conflicts of Interest
Abbreviations
| ACN | Acetonitrile |
| AcOH | Acetic Acid |
| AMP-DNJ | Adamantanemethoxypentyl-DNJ |
| BAL | Bronchoalveolar lavage |
| Bn | Benzyl |
| Bu | Butyl |
| BVDV | Bovine viral diarrhea virus |
| Bz | Benzoyl |
| CF | Cystic Fibrosis |
| CFTR | Cystic Fibrosis Transmembrane Conductance Regulator |
| CuAAC | CuI-catalyzed Azide-Alkyne Cycloaddition |
| βCD | β cyclodextrin |
| DAB | 1,4-dideoxy-1,4-imino-d-arabinitol |
| DCM | Dichloromethane |
| DMF | N,N-dimethylformamide |
| DMSO | Dimethyl sulfoxide |
| DNJ | Deoxynojirimycin |
| EC50 | Half maximal effective concentration |
| ENaC | Epithelial Na+ channel |
| ER | Endoplasmatic Reticulum |
| Et | Ethyl |
| EtOAc | Ethyl Acetate |
| EtOH | Ethanol |
| FOS | Free Oligosaccharides |
| fsk | Forskoline |
| GCase | Lysosomal β-glucocerebrosidase |
| gst | Genistein |
| HCV | HepatitisC virus |
| HIV | Human immunodeficiency virus |
| HPDNJ | Hexyloxypentyl-DNJ |
| HSV | Herpes Simplex virus |
| IC50 | Half maximal inhibitory concentration |
| ICAM-1 | Intracellular adhesion molecule 1 |
| IL-8 | Interleukin-8 |
| Isc | Transepithelial short-circuit currents |
| LAB | 1,4-dideoxy-1,4-imino-l-arabinitol |
| Me | Methyl |
| MeOH | Methanol |
| NBDGJ | N-butyl-deoxygalactonojirimycin |
| NBDNJ | N-butyl-deoxynojirimycin |
| NBS | N-bromosuccinimide |
| NJ | Nojirimycin |
| NLGase | non-lysosomal β-glucosidase 2 |
| NMO | N-methylmorpholine N-oxide |
| NNDNJ | N-nonyl-DNJ |
| NPDNJ | N-nonyloxypentyl-DNJ |
| PD | Potential difference |
| PS-TPP | Polymer Supported Triphenylphosphine |
| Py | Pyridine |
| SLs | Sphingolipid |
| THF | Tetrahydrofuran |
| TFA | Trifluoroacetic acid |
| TPP | Triphenylphosphine |
| UPP | Ubiquitin-Proteasome Pathway |
References
- Compain, P.; Martin, O.R. (Eds.) Iminosugars: From Synthesis to Therapeutic Applications; John Wiley & Sons, Ltd.: Chichester, UK, 2007; ISBN 9780470517437. [Google Scholar]
- Inouye, S.; Tsuruoka, T.; Nida, T. The structure of nojirimycin, a piperidinose sugar antibiotic. J. Antibiot. (Tokyo) 1966, 19, 288–292. [Google Scholar] [PubMed]
- Gao, K.; Zheng, C.; Wang, T.; Zhao, H.; Wang, J.; Wang, Z.; Zhai, X.; Jia, Z.; Chen, J.; Zhou, Y.; et al. 1-Deoxynojirimycin: Occurrence, Extraction, Chemistry, Oral Pharmacokinetics, Biological Activities and In Silico Target Fishing. Molecules 2016, 21, 1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiitz, A.E. (Ed.) Iminosugars as Glycosidase Inhibitors; Wiley: Hoboken, NJ, USA, 1998; ISBN 9783527295449. [Google Scholar]
- Asano, N.; Nash, R.J.; Molyneux, R.J.; Fleet, G.W.J. Sugar-mimic glycosidase inhibitors: Natural occurrence, biological activity and prospects for therapeutic application. Tetrahedron Asymmetry 2000, 11, 1645–1680. [Google Scholar] [CrossRef]
- Gloster, T.M.; Davies, G.J. Glycosidase inhibition: Assessing mimicry of the transition state. Org. Biomol. Chem. 2010, 8, 305–320. [Google Scholar] [CrossRef] [Green Version]
- Lillelund, V.H.; Jensen, H.H.; Liang, X.; Bols, M. Recent developments of transition-state analogue glycosidase inhibitors of non-natural product origin. Chem. Rev. 2002, 102, 515–553. [Google Scholar] [CrossRef]
- Mellor, H.R.; Neville, D.C.A.; Harvey, D.J.; Platt, F.M.; Dwek, R.A.; Butters, T.D. Cellular effects of deoxynojirimycin analogues: Inhibition of N-linked oligosaccharide processing and generation of free glucosylated oligosaccharides. Biochem. J. 2004, 381, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Compain, P.; Martin, O. Design, Synthesis and Biological Evaluation of Iminosugar-Based Glycosyltransferase Inhibitors. Curr. Top. Med. Chem. 2003, 3, 541–560. [Google Scholar] [CrossRef]
- Compain, P. Glycomimetics: Design, Synthesis, and Therapeutic Applications. Molecules 2018, 23, 1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonzi, D.S.; Scott, K.A.; Dwek, R.A.; Zitzmann, N. Iminosugar antivirals: The therapeutic sweet spot. Biochem. Soc. Trans. 2017, 45, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Block, T.M.; Guo, J.-T. Antiviral therapies targeting host ER alpha-glucosidases: Current status and future directions. Antivir. Res. 2013, 99, 251–260. [Google Scholar] [CrossRef]
- Somsak, L.; Nagy, V.; Hadady, Z.; Docsa, T.; Gergely, P. Glucose Analog Inhibitors of Glycogen Phosphorylases as Potential Antidiabetic Agents: Recent Developments. Curr. Pharm. Des. 2003, 9, 1177–1189. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Kizu, H.; Yamashita, T.; Kameda, Y.; Kato, A.; Nash, R.J.; Fleet, G.W.J.; Molyneux, R.J.; Asano, N. New Sugar-Mimic Alkaloids from the Pods of Angylocalyx pynaertii. J. Nat. Prod. 2002, 65, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Aerts, J.M.; Ottenhoff, R.; Powlson, A.S.; Grefhorst, A.; van Eijk, M.; Dubbelhuis, P.F.; Aten, J.; Kuipers, F.; Serlie, M.J.; Wennekes, T.; et al. Pharmacological Inhibition of Glucosylceramide Synthase Enhances Insulin Sensitivity. Diabetes 2007, 56, 1341–1349. [Google Scholar] [CrossRef] [Green Version]
- Ferhati, X.; Matassini, C.; Fabbrini, M.G.; Goti, A.; Morrone, A.; Cardona, F.; Moreno-Vargas, A.J.; Paoli, P. Dual targeting of PTP1B and glucosidases with new bifunctional iminosugar inhibitors to address type 2 diabetes. Bioorg. Chem. 2019, 87, 534–549. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, H.; Tsukida, T.; Inoue, Y.; Yokota, K.; Yoshino, K.; Kondo, H.; Miura, N.; Nishimura, S.I. Azasugar-Based MMP/ADAM Inhibitors as Antipsoriatic Agents. J. Med. Chem. 2004, 47, 1930–1938. [Google Scholar] [CrossRef] [PubMed]
- Schramm, V.; Tyler, P. Imino-Sugar-Based Nucleosides. Curr. Top. Med. Chem. 2003, 3, 525–540. [Google Scholar] [CrossRef]
- Lee, R.E.; Smith, M.D.; Nash, R.J.; Griffiths, R.C.; McNeil, M.; Grewal, R.K.; Yan, W.; Besra, G.S.; Brennan, P.J.; Fleet, G.W.J. Inhibition of UDP-Gal mutase and mycobacterial galactan biosynthesis by pyrrolidine analogues of galactofuranose. Tetrahedron Lett. 1997, 38, 6733–6736. [Google Scholar] [CrossRef]
- Lee, R.E.; Smith, M.D.; Pickering, L.; Fleet, G.W.J. An approach to combinatorial library generation of galactofuranose mimics as potential inhibitors of mycobacterial cell wall biosynthesis: Synthesis of a peptidomimetic of uridine 5′-diphosphogalactofuranose (UDP-Galf). Tetrahedron Lett. 1999, 40, 8689–8692. [Google Scholar] [CrossRef]
- Ye, X.; Sun, F.; Liu, M.; Li, Q. Synthetic Iminosugar Derivatives as New Potential Immunosuppressive Agents. J. Med. Chem. 2005, 48, 3688–3691. [Google Scholar] [CrossRef]
- Li, Q.; Ye, X. Iminosugars as Immunomodulating Agents: Synthesis and Biological Activities of 1-Deoxynojirimycin and Related Compounds. Isr. J. Chem. 2015, 55, 336–346. [Google Scholar] [CrossRef]
- Wang, G.; Xiong, Y.; Ye, J.; Zhang, L.; Ye, X. Synthetic N-Alkylated Iminosugars as New Potential Immunosuppressive Agents. ACS Med. Chem. Lett. 2011, 2, 682–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrodnigg, T.M.; Steiner, A.J.; Ueberbacher, B.J. Natural and Synthetic Iminosugars as Carbohydrate Processing Enzyme Inhibitors for Cancer Therapy. Curr. Med. Chem. Anti Cancer Agents 2008, 8, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Boyd, R.E.; Lee, G.; Rybczynski, P.; Benjamin, E.R.; Khanna, R.; Wustman, B.A.; Valenzano, K.J. Pharmacological chaperones as therapeutics for lysosomal storage diseases. J. Med. Chem. 2013, 56, 2705–2725. [Google Scholar] [CrossRef] [PubMed]
- Valenzano, K.J.; Khanna, R.; Powe, A.C.; Boyd, R.; Lee, G.; Flanagan, J.J.; Benjamin, E.R. Identification and Characterization of Pharmacological Chaperones to Correct Enzyme Deficiencies in Lysosomal Storage Disorders. Assay Drug Dev. Technol. 2011, 9, 213–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parenti, G.; Andria, G.; Valenzano, K.J. Pharmacological chaperone therapy: Preclinical development, clinical translation, and prospects for the treatment of lysosomal storage disorders. Mol. Ther. 2015, 23, 1138–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Alonzo, D.; De Fenza, M.; Porto, C.; Iacono, R.; Huebecker, M.; Cobucci-Ponzano, B.; Priestman, D.A.; Platt, F.; Parenti, G.; Moracci, M.; et al. N-Butyl-l-deoxynojirimycin (l-NBDNJ): Synthesis of an Allosteric Enhancer of α-Glucosidase Activity for the Treatment of Pompe Disease. J. Med. Chem. 2017, 60, 9462–9469. [Google Scholar] [CrossRef]
- Butters, T.D.; Dwek, R.A.; Platt, F.M. Therapeutic Applications of Imino Sugars in Lysosomal Storage Disorders. Curr. Top. Med. Chem. 2003, 3, 561–574. [Google Scholar] [CrossRef]
- Scott, L.J.; Spencer, C.M. Miglitol: A review of its therapeutic potential in type 2 diabetes mellitus. Drugs 2000, 59, 521–549. [Google Scholar] [CrossRef]
- Sels, J.P.J.E.; Huijberts, M.S.P.; Wolffenbuttel, B.H.R. Miglitol, a new α-glucosidase inhibitor. Expert Opin. Pharmacother. 1999, 1, 149–156. [Google Scholar] [CrossRef]
- Hollak, C.E.M.; Hughes, D.; van Schaik, I.N.; Schwierin, B.; Bembi, B. Miglustat (Zavesca®) in type 1 Gaucher disease: 5-year results of a post-authorisation safety surveillance programme. Pharmacoepidemiol. Drug Saf. 2009, 18, 770–777. [Google Scholar] [CrossRef]
- Platt, F.M.; Jeyakumar, M.; Andersson, U.; Priestman, D.A.; Dwek, R.A.; Butters, T.D.; Cox, T.M.; Lachmann, R.H.; Hollak, C.; Aerts, J.M.F.G.; et al. Inhibition of substrate synthesis as a strategy for glycolipid lysosomal storage disease therapy. J. Inherit. Metab. Dis. 2001, 24, 275–290. [Google Scholar] [CrossRef] [PubMed]
- Pineda, M.; Walterfang, M.; Patterson, M.C. Miglustat in Niemann-Pick disease type C patients: A review. Orphanet J. Rare Dis. 2018, 13, 140. [Google Scholar] [CrossRef] [PubMed]
- Platt, F.M.; Neises, G.R.; Dwek, R.A.; Butters, T.D. N-butyldeoxynojirimycin is a novel inhibitor of glycolipid biosynthesis. J. Biol. Chem. 1994, 269, 8362–8365. [Google Scholar] [PubMed]
- Jespersen, T.M.; Dong, W.; Sierks, M.R.; Skrydstrup, T.; Lundt, I.; Bols, M. Isofagomine, a Potent, New Glycosidase Inhibitor. Angew. Chem. Int. Ed. Engl. 1994, 33, 1778–1779. [Google Scholar] [CrossRef]
- Kornhaber, G.J.; Tropak, M.B.; Maegawa, G.H.; Tuske, S.J.; Coales, S.J.; Mahuran, D.J.; Hamuro, Y. Isofagomine Induced Stabilization of Glucocerebrosidase. ChemBioChem 2008, 9, 2643–2649. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ran, H.; Liou, B.; Quinn, B.; Zamzow, M.; Zhang, W.; Bielawski, J.; Kitatani, K.; Setchell, K.D.R.; Hannun, Y.A.; et al. Isofagomine In Vivo Effects in a Neuronopathic Gaucher Disease Mouse. PLoS ONE 2011, 6, e19037. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.Q.; Ishii, S.; Asano, N.; Suzuki, Y. Accelerated transport and maturation of lysosomal α-galactosidase A in fabry lymphoblasts by an enzyme inhibitor. Nat. Med. 1999, 5, 112–115. [Google Scholar] [CrossRef]
- Khanna, R.; Soska, R.; Lun, Y.; Feng, J.; Frascella, M.; Young, B.; Brignol, N.; Pellegrino, L.; Sitaraman, S.A.; Desnick, R.J.; et al. The pharmacological chaperone 1-deoxygalactonojirimycin reduces tissue globotriaosylceramide levels in a mouse model of fabry disease. Mol. Ther. 2010, 18, 23–33. [Google Scholar] [CrossRef]
- Benjamin, E.R.; Flanagan, J.J.; Schilling, A.; Chang, H.H.; Agarwal, L.; Katz, E.; Wu, X.; Pine, C.; Wustman, B.; Desnick, R.J.; et al. The pharmacological chaperone 1-deoxygalactonojirimycin increases α-galactosidase A levels in Fabry patient cell lines. J. Inherit. Metab. Dis. 2009, 32, 424–440. [Google Scholar] [CrossRef]
- Benjamin, E.R.; Della Valle, M.C.; Wu, X.; Katz, E.; Pruthi, F.; Bond, S.; Bronfin, B.; Williams, H.; Yu, J.; Bichet, D.G.; et al. The validation of pharmacogenetics for the identification of Fabry patients to be treated with migalastat. Genet. Med. 2017, 19, 430–438. [Google Scholar] [CrossRef] [Green Version]
- Uzuki, B.Y.S. Emerging novel concept of chaperone therapies for protein misfolding diseases. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2014, 90, 145–162. [Google Scholar]
- Porto, C.; Ferrara, M.C.; Meli, M.; Acampora, E.; Avolio, V.; Rosa, M.; Cobucci-Ponzano, B.; Colombo, G.; Moracci, M.; Andria, G.; et al. Pharmacological enhancement of α-glucosidase by the allosteric chaperone n-acetylcysteine. Mol. Ther. 2012, 20, 2201–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkinson, S.F.; Fleet, G.W.J.; Nash, R.J.; Koike, Y.; Adachi, I.; Yoshihara, A.; Morimoto, K.; Izumori, K.; Kato, A. Looking-glass synergistic pharmacological chaperones: DGJ and L-DGJ from the enantiomers of tagatose. Org. Lett. 2011, 13, 4064–4067. [Google Scholar] [CrossRef]
- Dechecchi, M.C.; Tamanini, A.; Cabrini, G. Molecular basis of cystic fibrosis: From bench to bedside. Ann. Transl. Med. 2018, 6, 334. [Google Scholar] [CrossRef] [PubMed]
- Fraser-Pitt, D.; O’Neil, D. Cystic fibrosis—A multiorgan protein misfolding disease. Futur. Sci. OA 2015, 1. [Google Scholar] [CrossRef]
- Brás, N.F.; Cerqueira, N.M.; Ramos, M.J.; Fernandes, P.A. Glycosidase inhibitors: A patent review (2008–2013). Expert Opin. Ther. Pat. 2014, 24, 857–874. [Google Scholar] [CrossRef]
- Lubamba, B.; Dhooghe, B.; Noel, S.; Leal, T. Cystic fibrosis: Insight into CFTR pathophysiology and pharmacotherapy. Clin. Biochem. 2012, 45, 1132–1144. [Google Scholar] [CrossRef]
- Bell, S.C.; Mall, M.A.; Gutierrez, H.; Macek, M.; Madge, S.; Davies, J.C.; Burgel, P.R.; Tullis, E.; Castaños, C.; Castellani, C.; et al. The future of cystic fibrosis care: A global perspective. Lancet Respir. Med. 2020, 8, 65–124. [Google Scholar] [CrossRef] [Green Version]
- Pedemonte, N.; Galietta, L.J.V. Pharmacological correctors of mutant CFTR mistrafficking. Front. Pharmacol. 2012, 3, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Ghelani, D.P.; Schneider-Futschik, E.K. Emerging Cystic Fibrosis Transmembrane Conductance Regulator Modulators as New Drugs for Cystic Fibrosis: A Portrait of in Vitro Pharmacology and Clinical Translation. ACS Pharmacol. Transl. Sci. 2020, 3, 4–10. [Google Scholar] [CrossRef]
- Zegarra-Moran, O.; Galietta, L.J.V. CFTR pharmacology. Cell. Mol. Life Sci. 2017, 74, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Spanò, V.; Montalbano, A.; Carbone, A.; Scudieri, P.; Galietta, L.J.V.; Barraja, P. An overview on chemical structures as ΔF508-CFTR correctors. Eur. J. Med. Chem. 2019, 180, 430–448. [Google Scholar] [CrossRef] [PubMed]
- Cantin, A.M.; Hartl, D.; Konstan, M.W.; Chmiel, J.F. Inflammation in cystic fibrosis lung disease: Pathogenesis and therapy. J. Cyst. Fibros. 2015, 14, 419–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dechecchi, M.C.; Nicolis, E.; Mazzi, P.; Cioffi, F.; Bezzerri, V.; Lampronti, I.; Huang, S.; Wiszniewski, L.; Gambari, R.; Scupoli, M.T.; et al. Modulators of sphingolipid metabolism reduce lung inflammation. Am. J. Respir. Cell Mol. Biol. 2011, 45, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Roesch, E.A.; Nichols, D.P.; Chmiel, J.F. Inflammation in cystic fibrosis: An update. Pediatr. Pulmonol. 2018, 53, S30–S50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flume, P.A.; Robinson, K.A.; O’Sullivan, B.P.; Finder, J.D.; Vender, R.L.; Wiley-Courand, D.B.; White, T.B.; Marshall, B.C.; Bujan, J.; Downs, A.; et al. Cystic fibrosis pulmonary guidelines: Airway clearance therapies. Respir. Care 2009, 54, 522–537. [Google Scholar]
- Martinucci, M.; Roscetto, E.; Iula, V.D.; Votsi, A.; Catania, M.R.; De Gregorio, E. Accurate identification of members of the Burkholderia cepacia complex in cystic fibrosis sputum. Lett. Appl. Microbiol. 2016, 62, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, B.Z.; Haaf, J.B.; Leal, T.; Noel, S. Cystic fibrosis transmembrane conductance regulator modulators in cystic fibrosis: Current perspectives. Clin. Pharmacol. Adv. Appl. 2016, 8, 127–140. [Google Scholar]
- Ratjen, F.; Bell, S.C.; Rowe, S.M.; Goss, C.H.; Quittner, A.L.; Bush, A. Cystic fibrosis. Nat. Rev. Dis. Prim. 2015, 1, 15010. [Google Scholar] [CrossRef]
- Ramsey, B.W.; Davies, J.; McElvaney, N.G.; Tullis, E.; Bell, S.C.; Dřevínek, P.; Griese, M.; McKone, E.F.; Wainwright, C.E.; Konstan, M.W.; et al. A CFTR Potentiator in Patients with Cystic Fibrosis and the G551D Mutation. N. Engl. J. Med. 2011, 365, 1663–1672. [Google Scholar] [CrossRef] [Green Version]
- Wainwright, C.E.; Elborn, J.S.; Ramsey, B.W.; Marigowda, G.; Huang, X.; Cipolli, M.; Colombo, C.; Davies, J.C.; De Boeck, K.; Flume, P.A.; et al. Lumacaftor–Ivacaftor in Patients with Cystic Fibrosis Homozygous for Phe508del CFTR. N. Engl. J. Med. 2015, 373, 220–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor-Cousar, J.L.; Munck, A.; McKone, E.F.; van der Ent, C.K.; Moeller, A.; Simard, C.; Wang, L.T.; Ingenito, E.P.; McKee, C.; Lu, Y.; et al. Tezacaftor–Ivacaftor in Patients with Cystic Fibrosis Homozygous for Phe508del. N. Engl. J. Med. 2017, 377, 2013–2023. [Google Scholar] [CrossRef] [PubMed]
- Heijerman, H.G.M.; McKone, E.F.; Downey, D.G.; Van Braeckel, E.; Rowe, S.M.; Tullis, E.; Mall, M.A.; Welter, J.J.; Ramsey, B.W.; McKee, C.M.; et al. Efficacy and safety of the elexacaftor plus tezacaftor plus ivacaftor combination regimen in people with cystic fibrosis homozygous for the F508del mutation: A double-blind, randomised, phase 3 trial. Lancet 2019, 394, 1940–1948. [Google Scholar] [CrossRef]
- Galietta, L.J.V.; Springsteel, M.F.; Eda, M.; Niedzinski, E.J.; By, K.; Haddadin, M.J.; Kurth, M.J.; Nantz, M.H.; Verkman, A.S. Novel CFTR Chloride Channel Activators Identified by Screening of Combinatorial Libraries Based on Flavone and Benzoquinolizinium Lead Compounds. J. Biol. Chem. 2001, 276, 19723–19728. [Google Scholar] [CrossRef] [Green Version]
- Elborn, J.S. Cystic fibrosis. Lancet 2016, 388, 2519–2531. [Google Scholar] [CrossRef]
- Wennekes, T.; Meijer, A.J.; Groen, A.K.; Boot, R.G.; Groener, J.E.; Van Eijk, M.; Ottenhoff, R.; Bijl, N.; Ghauharali, K.; Song, H.; et al. Dual-action lipophilic iminosugar improves glycemic control in obese rodents by reduction of visceral glycosphingolipids and buffering of carbohydrate assimilation. J. Med. Chem. 2010, 53, 689–698. [Google Scholar] [CrossRef]
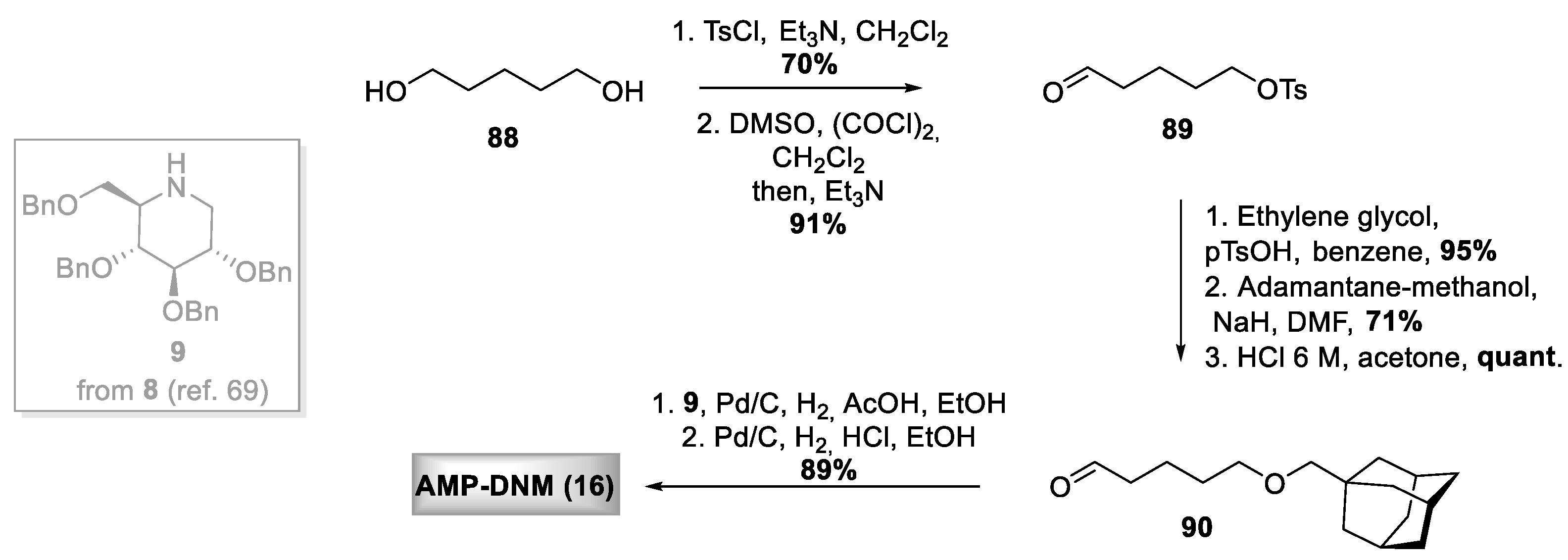
- Wennekes, T.; van den Berg, R.J.B.H.N.; Donker, W.; van der Marel, G.A.; Strijland, A.; Aerts, J.M.F.G.; Overkleeft, H.S. Development of Adamantan-1-yl-methoxy-Functionalized 1-Deoxynojirimycin Derivatives as Selective Inhibitors of Glucosylceramide Metabolism in Man. J. Org. Chem. 2007, 72, 1088–1097. [Google Scholar] [CrossRef]
- Wennekes, T.; Lang, B.; Leeman, M.; Van Der Marel, G.A.; Smits, E.; Weber, M.; Van Wiltenburg, J.; Wolberg, M.; Aerts, J.M.F.G.; Overkleeft, H.S. Large-scale synthesis of the glucosylceramide synthase inhibitor N-[5-(adamantan-1-yl-methoxy)-pentyl]-1-deoxynojirimycin. Org. Process Res. Dev. 2008, 12, 414–423. [Google Scholar] [CrossRef]
- Asano, N.; Kizu, H.; Oseki, K.; Tomioka, E.; Matsui, K.; Okamoto, M.; Baba, M. N-Alkylated Nitrogen-in-the-Ring Sugars: Conformational Basis of Inhibition of Glycosidases and HIV-1 Replication. J. Med. Chem. 1995, 38, 2349–2356. [Google Scholar] [CrossRef]
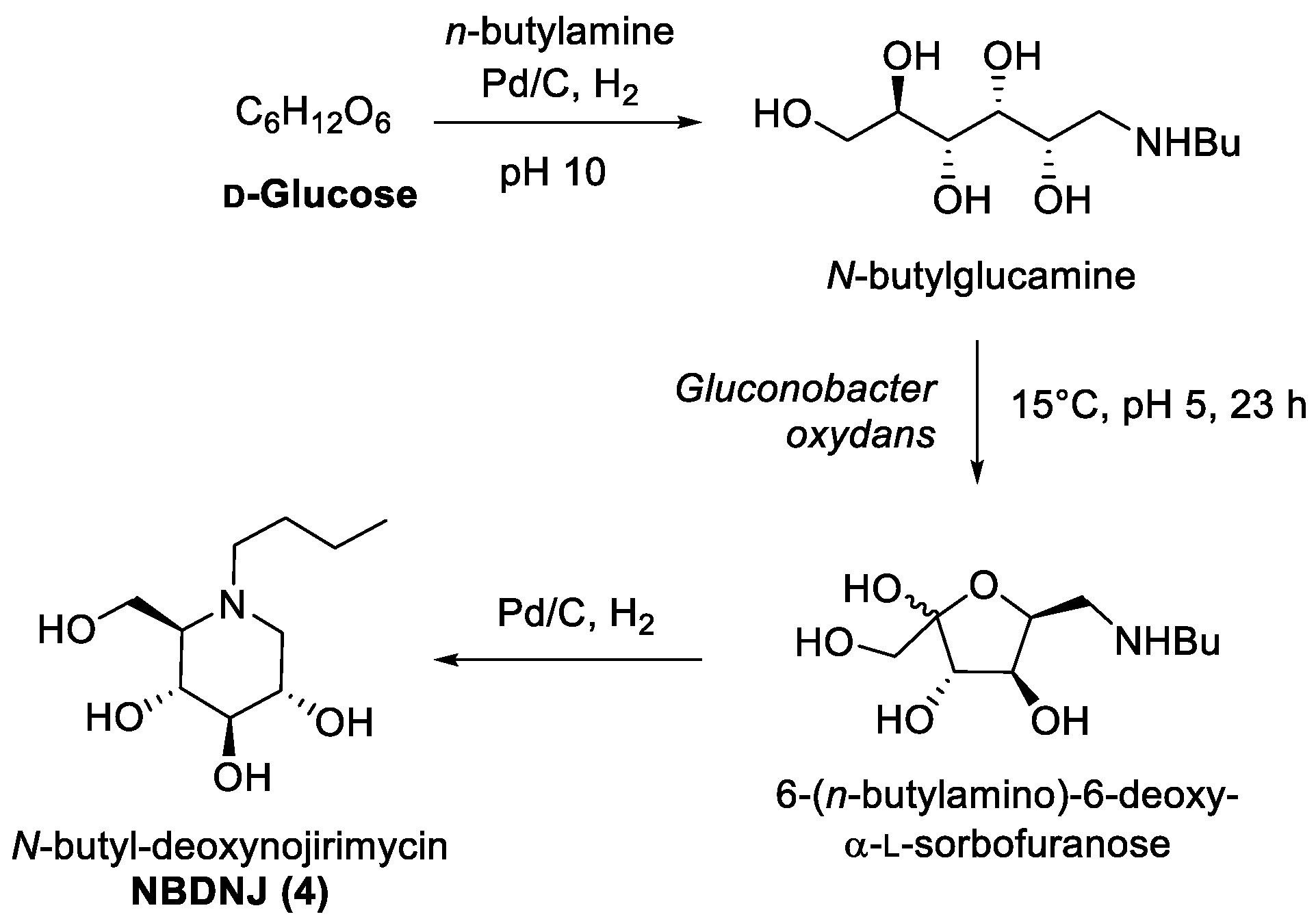
- Landis, B.H.; McLaughlin, J.K.; Heeren, R.; Grabner, R.W.; Wang, P.T. Bioconversion of N -Butylglucamine to 6-Deoxy-6-butylamino Sorbose by Gluconobacter o xydans. Org. Process Res. Dev. 2002, 6, 547–552. [Google Scholar] [CrossRef]
- Grabner, R.W.; Landis, B.H.; Wang, P.T.; Prunier, M.L.; Scaros, M.G. Process for Producing N-Substituted-1-Deoxynojirimycin. US Patent 5610039, 11 March 1997. [Google Scholar]
- Tierney, M.; Pottage, J.; Kessler, H.; Fischl, M.; Richman, D.; Merigan, T.; Powderly, W.; Smith, S.; Karim, A.; Sherman, J.; et al. The tolerability and pharmacokinetics of N-butyl-deoxynojirimycin in patients with advanced HIV disease (ACTG 100). J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 1995, 10, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Norez, C.; Noel, S.; Wilke, M.; Bijvelds, M.; Jorna, H.; Melin, P.; Dejonge, H.; Becq, F. Rescue of functional delF508-CFTR channels in cystic fibrosis epithelial cells by the α-glucosidase inhibitor miglustat. FEBS Lett. 2006, 580, 2081–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norez, C.; Antigny, F.; Becq, F.; Vandebrouck, C. Maintaining low Ca2+ level in the endoplasmic reticulum restores abnormal endogenous F508del-CFTR trafficking in airway epithelial cells. Traffic 2006, 7, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Denning, G.M.; Anderson, M.P.; Amara, J.F.; Marshall, J.; Smith, A.E.; Welsh, M.J. Processing of mutant cystic fibrosis transmembrane conductance regulator is temperature-sensitive. Nature 1992, 358, 761–764. [Google Scholar] [CrossRef] [PubMed]
- Dechecchi, M.C.; Nicolis, E.; Norez, C.; Bezzerri, V.; Borgatti, M.; Mancini, I.; Rizzotti, P.; Ribeiro, C.M.P.; Gambari, R.; Becq, F.; et al. Anti-inflammatory effect of miglustat in bronchial epithelial cells. J. Cyst. Fibros. 2008, 7, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Noël, S.; Wilke, M.; Bot, A.G.M.; De Jonge, H.R.; Becq, F. Parallel improvement of sodium and chloride transport defects by miglustat (n-butyldeoxynojyrimicin) in cystic fibrosis epithelial cells. J. Pharmacol. Exp. Ther. 2008, 325, 1016–1023. [Google Scholar] [CrossRef] [Green Version]
- Antigny, F.; Norez, C.; Becq, F.; Vandebrouck, C. Calcium homeostasis is abnormal in cystic fibrosis airway epithelial cells but is normalized after rescue of F508del-CFTR. Cell Calcium 2008, 43, 175–183. [Google Scholar] [CrossRef]
- Norez, C.; Antigny, F.; Noel, S.; Vandebrouck, C.; Becq, F. A cystic fibrosis respiratory epithelial cell chronically treated by miglustat acquires a non-cystic fibrosis-like phenotype. Am. J. Respir. Cell Mol. Biol. 2009, 41, 217–225. [Google Scholar] [CrossRef]
- Lubamba, B.; Lebacq, J.; Lebecque, P.; Vanbever, R.; Leonard, A.; Wallemacq, P.; Leal, T. Airway delivery of low-dose miglustat normalizes nasal potential difference in F508del cystic fibrosis mice. Am. J. Respir. Crit. Care Med. 2009, 179, 1022–1028. [Google Scholar] [CrossRef] [Green Version]
- Le Henaff, C.; Haÿ, E.; Velard, F.; Marty, C.; Tabary, O.; Marie, P.J.; Jacquot, J.P. Enhanced F508del-CFTR channel activity ameliorates bone pathology in murine cystic fibrosis. Am. J. Pathol. 2014, 184, 1132–1141. [Google Scholar] [CrossRef]
- Leonard, A.; Lebecque, P.; Dingemanse, J.; Leal, T. A randomized placebo-controlled trial of miglustat in cystic fibrosis based on nasal potential difference. J. Cyst. Fibros. 2012, 11, 231–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, A.; Lebecque, P.; Dingemanse, J.; Leal, T. Miglustat effects on the basal nasal potential differences in cystic fibrosis. J. Cyst. Fibros. 2013, 12, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horne, G.; Wilson, F.X.; Tinsley, J.; Williams, D.H.; Storer, R. Iminosugars past, present and future: Medicines for tomorrow. Drug Discov. Today 2011, 16, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Nash, R.J.; Kato, A.; Yu, C.Y.; Fleet, G.W. Iminosugars as therapeutic agents: Recent advances and promising trends. Future Med. Chem. 2011, 3, 1513–1521. [Google Scholar] [CrossRef]
- Cendret, V.; Legigan, T.; Mingot, A.; Thibaudeau, S.; Adachi, I.; Forcella, M.; Parenti, P.; Bertrand, J.; Becq, F.; Norez, C.; et al. Synthetic deoxynojirimycin derivatives bearing a thiolated, fluorinated or unsaturated N-alkyl chain: Identification of potent α-glucosidase and trehalase inhibitors as well as F508del-CFTR correctors. Org. Biomol. Chem. 2015, 13, 10734–10744. [Google Scholar] [CrossRef]
- Overkleeft, H.S.; van Wiltenburg, J.; Pandit, U.K. A facile transformation of sugar lactones to azasugars. Tetrahedron 1994, 50, 4215–4224. [Google Scholar] [CrossRef]
- Compain, G.; Jouvin, K.; Martin-Mingot, A.; Evano, G.; Marrot, J.; Thibaudeau, S. Stereoselective hydrofluorination of ynamides: A straightforward synthesis of novel α-fluoroenamides. Chem. Commun. 2012, 48, 5196–5198. [Google Scholar] [CrossRef]
- Cantet, A.-C.; Carreyre, H.; Gesson, J.-P.; Jouannetaud, M.-P.; Renoux, B. gem -Difluorination of Aminoalkynes via Highly Reactive Dicationic Species in Superacid HF−SbF 5: Application to the Efficient Synthesis of Difluorinated Cinchona Alkaloid Derivatives. J. Org. Chem. 2008, 73, 2875–2878. [Google Scholar] [CrossRef]
- Bilska-Markowska, M.; Szwajca, A.; Marciniak, B. Design, properties and applications of fluorinated and fluoroalkylated N-containing monosaccharides and their analogues. J. Fluor. Chem. 2019, 227, 109364. [Google Scholar] [CrossRef]
- Kammouni, W.; Moreau, B.; Becq, F.; Saleh, A.; Pavirani, A.; Figarella, C.; Merten, M.D. A Cystic Fibrosis Tracheal Gland Cell Line, CF-KM4. Am. J. Respir. Cell Mol. Biol. 1999, 20, 684–691. [Google Scholar] [CrossRef]
- Norez, C.; Heda, G.D.; Jensen, T.; Kogan, I.; Hughes, L.K.; Auzanneau, C.; Dérand, R.; Bulteau-Pignoux, L.; Li, C.; Ramjeesingh, M.; et al. Determination of CFTR chloride channel activity and pharmacology using radiotracer flux methods. J. Cyst. Fibros. 2004, 3, 119–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brumshtein, B.; Wormald, M.R.; Silman, I.; Futerman, A.H.; Sussman, J.L. Structural comparison of differently glycosylated forms of acid-β-glucosidase, the defective enzyme in Gaucher disease. Acta Crystallogr. Sect. D Biol. Crystallogr. 2006, 62, 1458–1465. [Google Scholar] [CrossRef] [PubMed]
- Ghisaidoobe, A.; Bikker, P.; De Bruijn, A.C.J.; Godschalk, F.D.; Rogaar, E.; Guijt, M.C.; Hagens, P.; Halma, J.M.; Van’t Hart, S.M.; Luitjens, S.B.; et al. Identification of potent and selective glucosylceramide synthase inhibitors from a library of N-alkylated iminosugars. ACS Med. Chem. Lett. 2011, 2, 119–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellor, H.R.; Nolan, J.; Pickering, L.; Wormald, M.R.; Platt, F.M.; Dwek, R.A.; Fleet, G.W.J.; Butters, T.D. Preparation, biochemical characterization and biological properties of radiolabelled N-alkylated deoxynojirimycins. Biochem. J. 2002, 233, 225–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
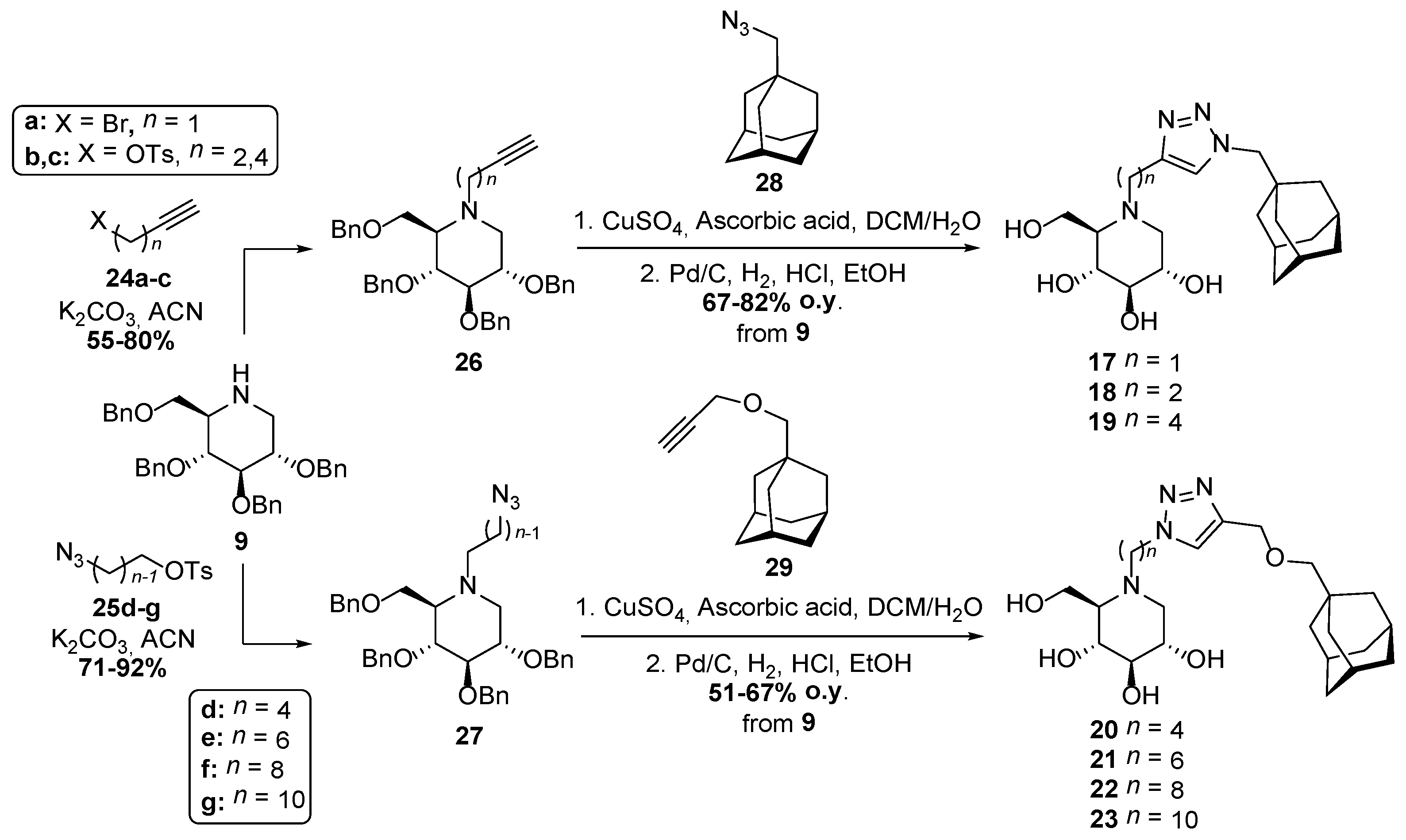
- Ardes-Guisot, N.; Alonzi, D.S.; Reinkensmeier, G.; Butters, T.D.; Norez, C.; Becq, F.; Shimada, Y.; Nakagawa, S.; Kato, A.; Blériot, Y.; et al. Selection of the biological activity of DNJ neoglycoconjugates through click length variation of the side chain. Org. Biomol. Chem. 2011, 9, 5373–5388. [Google Scholar] [CrossRef] [PubMed]
- Overkleeft, H.S.; Renkema, G.H.; Neele, J.; Vianello, P.; Hung, I.O.; Strijland, A.; Van Der Burg, A.M.; Koomen, G.J.; Pandit, U.K.; Aerts, J.M.F.G. Generation of specific deoxynojirimycin-type inhibitors of the non- lysosomal glucosylceramidase. J. Biol. Chem. 1998, 273, 26522–26527. [Google Scholar] [CrossRef] [Green Version]
- Butters, T.D.; Dwek, R.A.; Platt, F.M. Inhibition of glycosphingolipid biosynthesis: Application to lysosomal storage disorders. Chem. Rev. 2000, 100, 4683–4696. [Google Scholar] [CrossRef]
- Wennekes, T.; van den Berg, R.J.B.H.N.; Bonger, K.M.; Donker-Koopman, W.E.; Ghisaidoobe, A.; van der Marel, G.A.; Strijland, A.; Aerts, J.M.F.G.; Overkleeft, H.S. Synthesis and evaluation of dimeric lipophilic iminosugars as inhibitors of glucosylceramide metabolism. Tetrahedron Asymmetry 2009, 20, 836–846. [Google Scholar] [CrossRef]
- Van Den Berg, R.J.B.H.N.; Wennekes, T.; Ghisaidoobe, A.; Donker-Koopman, W.E.; Strijland, A.; Boot, R.G.; Van Der Marel, G.A.; Aerts, J.M.F.G.; Overkleeft, H.S. Assessment of partially deoxygenated deoxynojirimycin derivatives as glucosylceramide synthase inhibitors. ACS Med. Chem. Lett. 2011, 2, 519–522. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Eguchi, S.; Katada, T.; Hiroaki, O. Synthesis of adamantane derivatives. 37. A convenient and efficient synthesis of 1-azidoadamantane and related bridgehead azides, and some of their reactions. J. Org. Chem. 1977, 42, 3741–3743. [Google Scholar] [CrossRef]
- Tornøe, C.W.; Christensen, C.; Meldal, M. Peptidotriazoles on solid phase: [1,2,3]-Triazoles by regiospecific copper(I)-catalyzed 1,3-dipolar cycloadditions of terminal alkynes to azides. J. Org. Chem. 2002, 67, 3057–3064. [Google Scholar] [CrossRef] [PubMed]
- Baron, A.; Blériot, Y.; Sollogoub, M.; Vauzeilles, B. Phenylenediamine catalysis of “click glycosylations” in water: Practical and direct access to unprotected neoglycoconjugates. Org. Biomol. Chem. 2008, 6, 1898. [Google Scholar] [CrossRef] [Green Version]
- Compain, P.; Decroocq, C.; Joosten, A.; de Sousa, J.; Rodríguez-Lucena, D.; Butters, T.D.; Bertrand, J.; Clément, R.; Boinot, C.; Becq, F.; et al. Rescue of functional CFTR channels in cystic fibrosis: A dramatic multivalent effect using iminosugar cluster-based correctors. ChemBioChem 2013, 14, 2050–2058. [Google Scholar] [CrossRef] [PubMed]
- Compain, P.; Bodlenner, A. The multivalent effect in glycosidase inhibition: A new, rapidly emerging topic in glycoscience. ChemBioChem 2014, 15, 1239–1251. [Google Scholar] [CrossRef] [PubMed]
- Compain, P. Multivalent Effect in Glycosidase Inhibition: The End of the Beginning. Chem. Rec. 2020, 20, 10–22. [Google Scholar] [CrossRef]
- Matassini, C.; Parmeggiani, C.; Cardona, F.; Goti, A. Are enzymes sensitive to the multivalent effect? Emerging evidence with glycosidases. Tetrahedron Lett. 2016, 57, 5407–5415. [Google Scholar] [CrossRef]
- Joosten, A.; Decroocq, C.; De Sousa, J.; Schneider, J.P.; Etamé, E.; Bodlenner, A.; Butters, T.D.; Compain, P. A systematic investigation of iminosugar click clusters as pharmacological chaperones for the treatment of gaucher disease. ChemBioChem 2014, 15, 309–319. [Google Scholar] [CrossRef]
- Lohse, A.; Jensen, K.B.; Lundgren, K.; Bols, M. Synthesis and deconvolution of the first combinatorial library of glycosidase inhibitors. Bioorg. Med. Chem. 1999, 7, 1965–1971. [Google Scholar] [CrossRef]
- Gouin, S.G. Multivalent Inhibitors for Carbohydrate-Processing Enzymes: Beyond the “Lock-and-Key” Concept. Chem. Eur. J. 2014, 20, 11616–11628. [Google Scholar] [CrossRef] [PubMed]
- Zelli, R.; Longevial, J.-F.; Dumy, P.; Marra, A. Synthesis and biological properties of multivalent iminosugars. New J. Chem. 2015, 39, 5050–5074. [Google Scholar] [CrossRef]
- Diot, J.; García-Moreno, M.I.; Gouin, S.G.; Mellet, C.O.; Haupt, K.; Kovensky, J. Multivalent iminosugars to modulate affinity and selectivity for glycosidases. Org. Biomol. Chem. 2009, 7, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Rísquez-Cuadro, R.; García Fernández, J.M.; Nierengarten, J.F.; Ortiz Mellet, C. Fullerene-sp2-iminosugar balls as multimodal ligands for lectins and glycosidases: A mechanistic hypothesis for the inhibitory multivalent effect. Chem. Eur. J. 2013, 19, 16791–16803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decroocq, C.; Rodríguez-Lucena, D.; Russo, V.; Mena Barragán, T.; Ortiz Mellet, C.; Compain, P. The multivalent effect in glycosidase inhibition: Probing the influence of architectural parameters with cyclodextrin-based iminosugar click clusters. Chem. Eur. J. 2011, 17, 13825–13831. [Google Scholar] [CrossRef] [PubMed]
- Decroocq, C.; Joosten, A.; Sergent, R.; Mena Barragán, T.; Ortiz Mellet, C.; Compain, P. The multivalent effect in glycosidase inhibition: Probing the influence of valency, peripheral ligand structure, and topology with cyclodextrin-based iminosugar click clusters. ChemBioChem 2013, 14, 2038–2049. [Google Scholar] [CrossRef]
- Stauffert, F.; Serra-Vinardell, J.; Gómez-Grau, M.; Michelakakis, H.; Mavridou, I.; Grinberg, D.; Vilageliu, L.; Casas, J.; Bodlenner, A.; Delgado, A.; et al. Stereodivergent synthesis of right- and left-handed iminoxylitol heterodimers and monomers. Study of their impact on β-glucocerebrosidase activity. Org. Biomol. Chem. 2017, 15, 3681–3705. [Google Scholar] [CrossRef]
- Decroocq, C.; Rodríguez-Lucena, D.; Ikeda, K.; Asano, N.; Compain, P. Cyclodextrin-Based Iminosugar Click Clusters: The First Examples of Multivalent Pharmacological Chaperones for the Treatment of Lysosomal Storage Disorders. ChemBioChem 2012, 13, 661–664. [Google Scholar] [CrossRef]
- Laigre, E.; Hazelard, D.; Casas, J.; Serra-Vinardell, J.; Michelakakis, H.; Mavridou, I.; Aerts, J.M.F.G.; Delgado, A.; Compain, P. Investigation of original multivalent iminosugars as pharmacological chaperones for the treatment of Gaucher disease. Carbohydr. Res. 2016, 429, 98–104. [Google Scholar] [CrossRef]
- Compain, P.; Decroocq, C.; Iehl, J.; Holler, M.; Hazelard, D.; Barragán, T.M.; Mellet, C.O.; Nierengarten, J.F. Glycosidase inhibition with fullerene iminosugar balls: A dramatic multivalent effect. Angew. Chem. Int. Ed. 2010, 49, 5753–5756. [Google Scholar] [CrossRef]
- Rostovtsev, V.V.; Green, L.G.; Fokin, V.V.; Sharpless, K.B. A Stepwise Huisgen Cycloaddition Process: Copper(I)-Catalyzed Regioselective “Ligation” of Azides and Terminal Alkynes. Angew. Chem. Int. Ed. 2002, 41, 2596–2599. [Google Scholar] [CrossRef]
- Mollard, A.; Zharov, I. Tricarboranyl Pentaerythritol-Based Building Block. Inorg. Chem. 2006, 45, 10172–10179. [Google Scholar] [CrossRef]
- Papp, I.; Dernedde, J.; Enders, S.; Haag, R. Modular synthesis of multivalent glycoarchitectures and their unique selectin binding behavior. Chem. Commun. 2008, 5851–5853. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Flores, F.G.; Isac-García, J.; Hernández-Mateo, F.; Pérez-Balderas, F.; Calvo-Asín, J.A.; Sanchéz-Vaquero, E.; Santoyo-González, F. 1,3-Dipolar Cycloadditions as a Tool for the Preparation of Multivalent Structures. Org. Lett. 2000, 2, 2499–2502. [Google Scholar] [CrossRef] [PubMed]
- Takeo, K.; Mitoh, H.; Uemura, K. Selective chemical modification of cyclomalto-oligosaccharides via tert-butyldimethylsilylation. Carbohydr. Res. 1989, 187, 203–221. [Google Scholar] [CrossRef]
- Ortega-Caballero, F.; Giménez-Martínez, J.J.; Vargas-Berenguel, A. Diverse Motifs of Mannoside Clustering on a β-Cyclodextrin Core. Org. Lett. 2003, 5, 2389–2392. [Google Scholar] [CrossRef]
- Fleet, G.W.J.; Nicholas, S.J.; Smith, P.W.; Evans, S.V.; Fellows, L.E.; Nash, R.J. Potent competitive inhibition of α-galactosidase and α-glucosidase activity by 1,4-dideoxy-1,4-iminopentitols: Syntheses of 1,4-dideoxy-1,4-imino-d-lyxitol and of both enantiomers of 1,4-dideoxy-1,4-iminoarabinitol. Tetrahedron Lett. 1985, 26, 3127–3130. [Google Scholar] [CrossRef]
- Watson, A.A.; Fleet, G.W.J.; Asano, N.; Molyneux, R.J.; Nash, R.J. Polyhydroxylated alkaloids—Natural occurrence and therapeutic applications. Phytochemistry 2001, 56, 265–295. [Google Scholar] [CrossRef]
- Davis, B.G. A silver-lined anniversary of Fleet iminosugars: 1984–2009, from DIM to DRAM to LABNAc. Tetrahedron Asymmetry 2009, 20, 652–671. [Google Scholar] [CrossRef]
- Cruz, F.P.D.; Newberry, S.; Jenkinson, S.F.; Wormald, M.R.; Butters, T.D.; Alonzi, D.S.; Nakagawa, S.; Becq, F.; Norez, C.; Nash, R.J.; et al. 4-C-Me-DAB and 4-C-Me-LAB—Enantiomeric alkyl-branched pyrrolidine iminosugars—Are specific and potent α-glucosidase inhibitors; Acetone as the sole protecting group. Tetrahedron Lett. 2011, 52, 219–223. [Google Scholar] [CrossRef] [Green Version]
- Fleet, G.W.J.; Karpas, A.; Dwek, R.A.; Fellows, L.E.; Tyms, A.S.; Petursson, S.; Namgoong, S.K.; Ramsden, N.G.; Smith, P.W.; Son, J.C.; et al. Inhibition of HIV replication by amino-sugar derivatives. FEBS Lett. 1988, 237, 128–132. [Google Scholar] [CrossRef] [Green Version]
- Behling, J.R.; Campbell, A.L.; Babiak, K.A.; Ng, J.S.; Medic, J.; Farid, P.; Fleet, G.W.J. Synthesis from D-lyxonolactone of 1,4-Dideoxy-1,4-imino-L-arabinitol, a glucosidase inhibitor with in vitro anti-viral activity. Tetrahedron 1993, 49, 3359–3366. [Google Scholar] [CrossRef]
- Scofield, A.M.; Fellows, L.E.; Nash, R.J.; Fleet, G.W.J. Inhibition of mammalian digestive disaccharidases by polyhydroxy alkaloids. Life Sci. 1986, 39, 645–650. [Google Scholar] [CrossRef]
- Karpas, A.; Fleet, G.W.J.; Dwek, R.A.; Petursson, S.; Namgoong, S.K.; Ramsden, N.G.; Jacob, G.S.; Rademacher, T.W. Aminosugar derivatives as potential anti-human immunodeficiency virus agents. Proc. Natl. Acad. Sci. USA 1988, 85, 9229–9233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
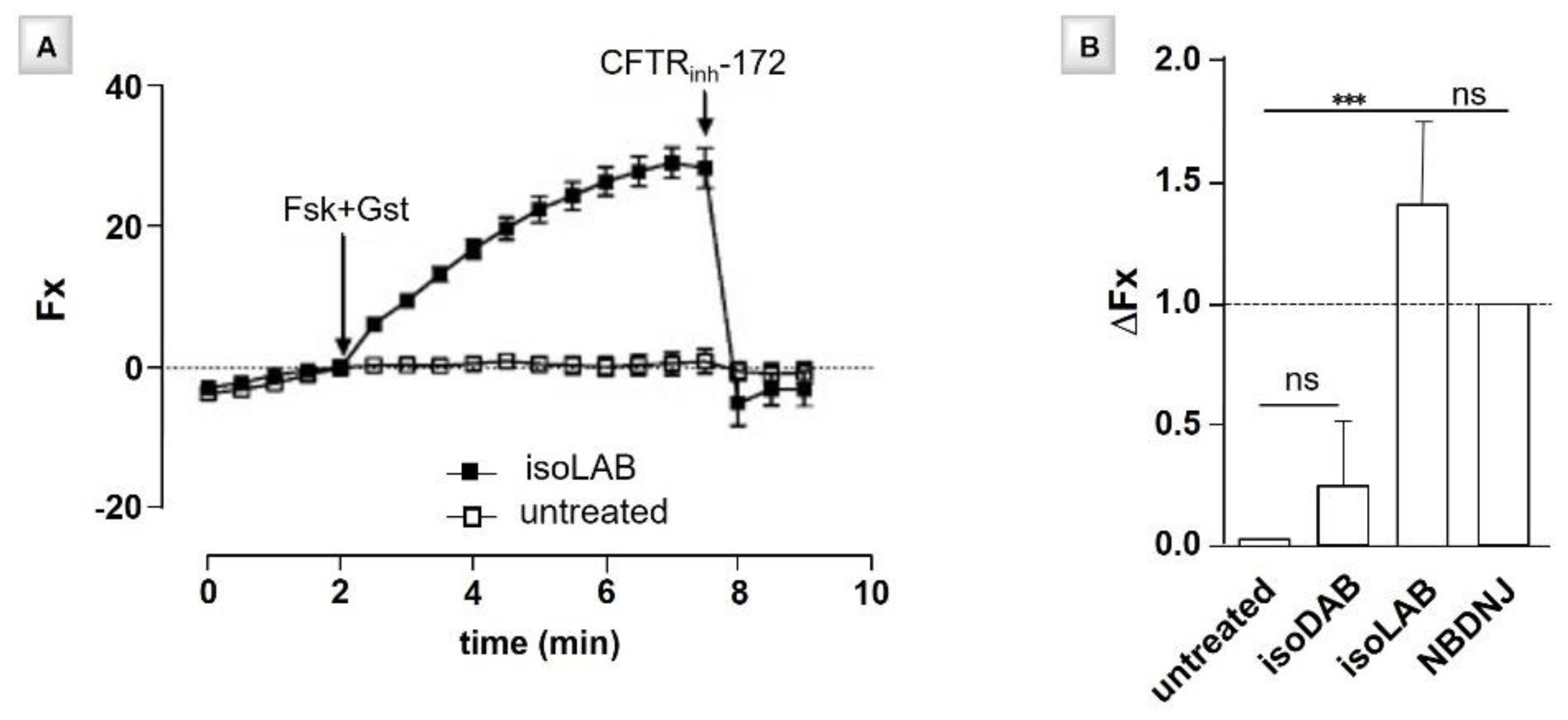
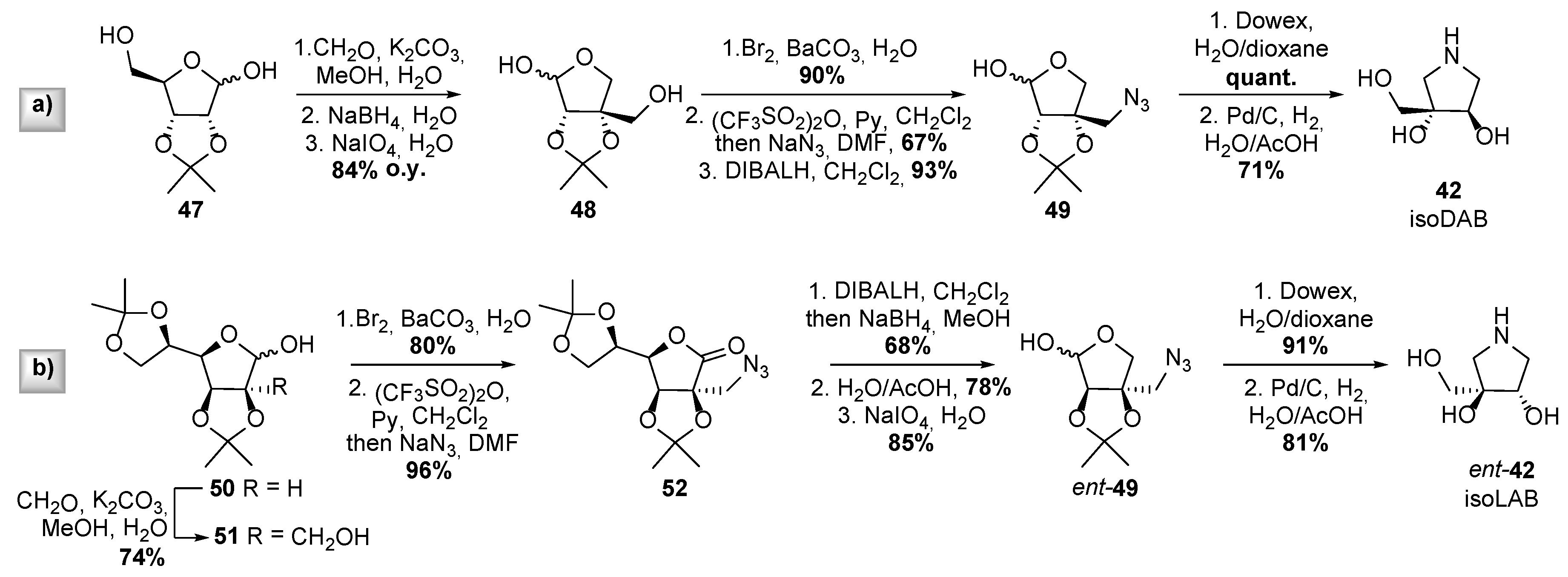
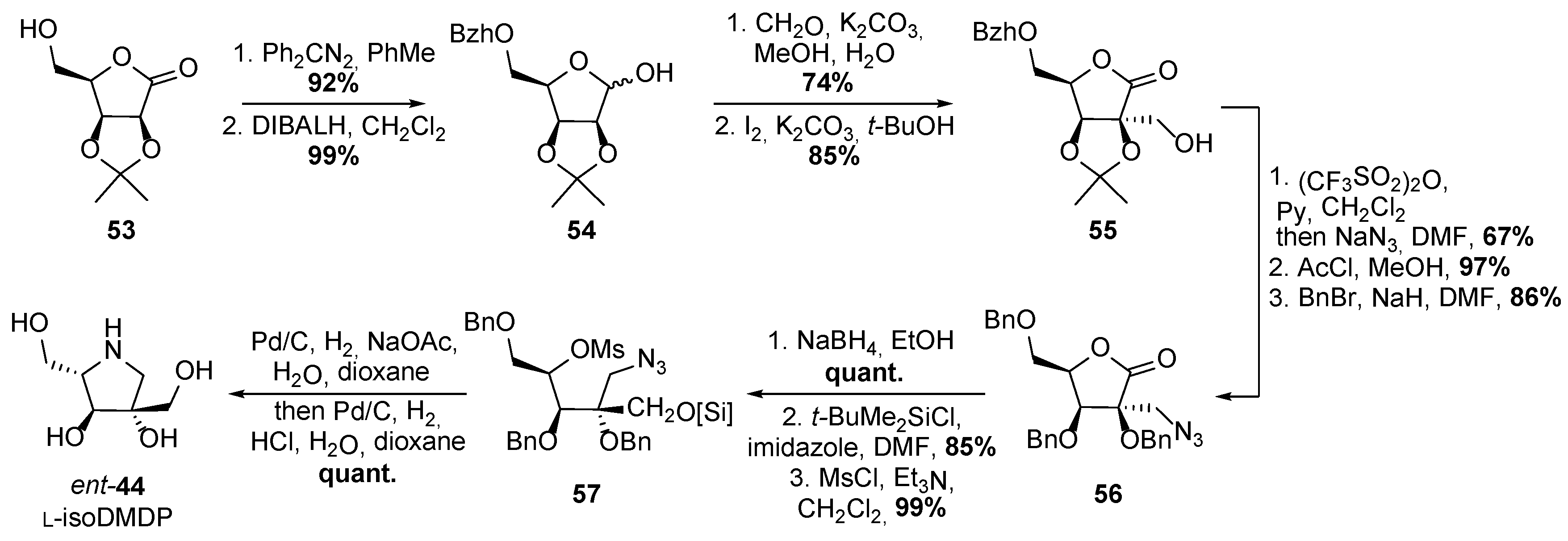
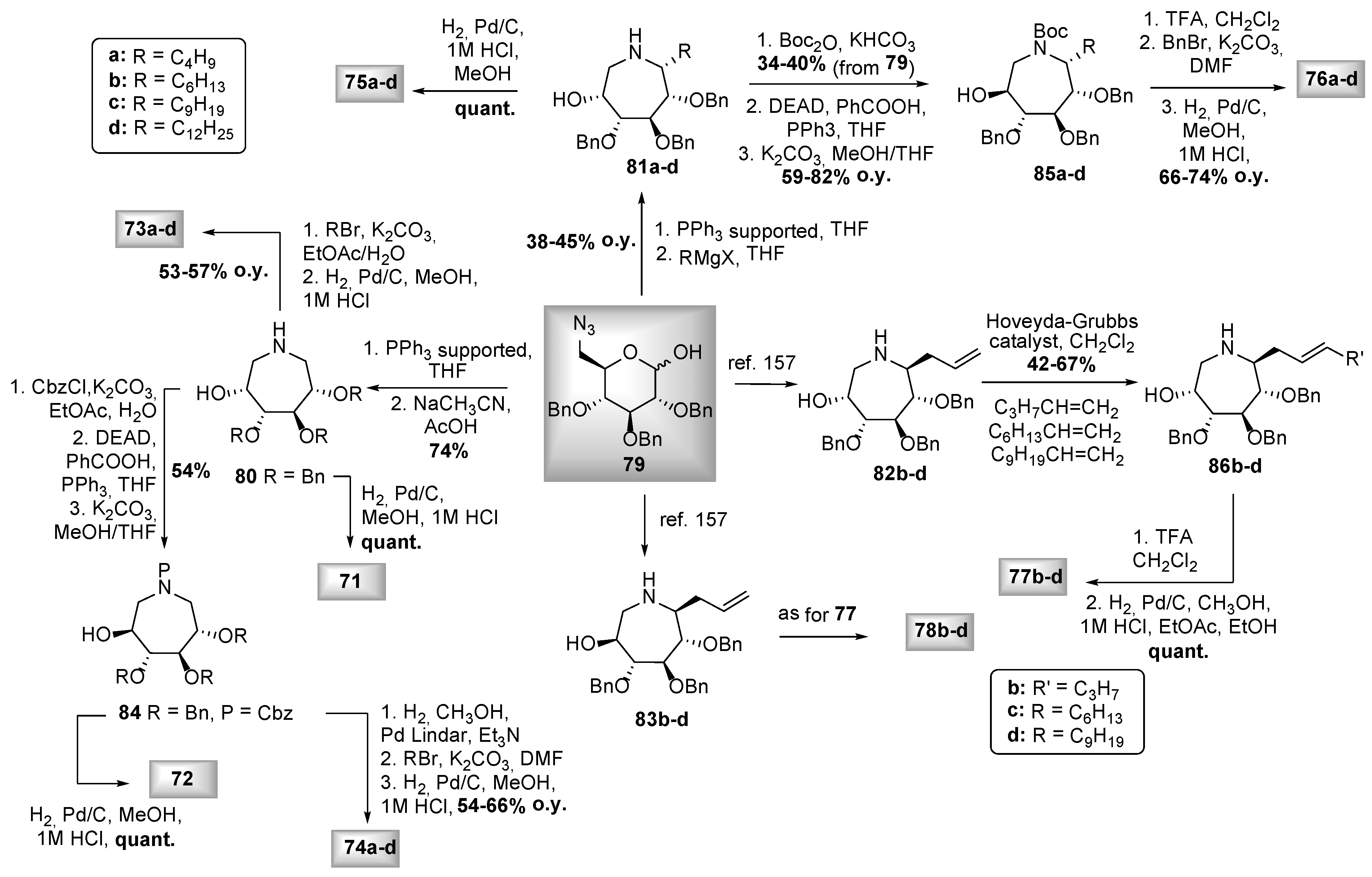
- Best, D.; Jenkinson, S.F.; Saville, A.W.; Alonzi, D.S.; Wormald, M.R.; Butters, T.D.; Norez, C.; Becq, F.; Blériot, Y.; Adachi, I.; et al. Cystic fibrosis and diabetes: IsoLAB and isoDAB, enantiomeric carbon-branched pyrrolidine iminosugars. Tetrahedron Lett. 2010, 51, 4170–4174. [Google Scholar] [CrossRef]
- Fleet, G.W.J. Pyrrolidine Iminosugars used in the treatment of Cystic Fibrosis. WIPO (PCT) Patent Application No WO2011086347-A1, 21 July 2011. [Google Scholar]
- Yu, C.-Y.; Asano, N.; Ikeda, K.; Wang, M.-X.; Butters, T.D.; Wormald, M.R.; Dwek, R.A.; Winters, A.L.; Nash, R.J.; Fleet, G.W.J. Looking glass inhibitors: L-DMDP, a more potent and specific inhibitor of α-glucosidases than the enantiomeric natural product DMDP. Chem. Commun. 2004, 1936–1937. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, S.F.; Best, D.; Saville, A.W.; Mui, J.; Martínez, R.F.; Nakagawa, S.; Kunimatsu, T.; Alonzi, D.S.; Butters, T.D.; Norez, C.; et al. C-branched iminosugars: α-glucosidase inhibition by enantiomers of isoDMDP, isoDGDP, and isoDAB- l -isoDMDP compared to miglitol and miglustat. J. Org. Chem. 2013, 78, 7380–7397. [Google Scholar] [CrossRef]
- Robertson, J.; Stevens, K. Pyrrolizidine alkaloids: Occurrence, biology, and chemical synthesis. Nat. Prod. Rep. 2017, 34, 62–89. [Google Scholar] [CrossRef]
- Dehoux-Baudoin, C.; Génisson, Y. C-Branched Imino Sugars: Synthesis and Biological Relevance. Eur. J. Org. Chem. 2019, 2019, 4765–4777. [Google Scholar] [CrossRef]
- Kurasaki, H.; Okamoto, I.; Morita, N.; Tamura, O. Total Synthesis of Grandisine D. Org. Lett. 2009, 11, 1179–1181. [Google Scholar] [CrossRef]
- Dehoux, C.; Castellan, T.; Enel, M.; André-Barrès, C.; Mirval, S.; Becq, F.; Ballereau, S.; Génisson, Y. Transalpinecine and Analogues: First Total Synthesis, Stereochemical Revision and Biological Evaluation. Eur. J. Org. Chem. 2019, 2019, 1830–1834. [Google Scholar] [CrossRef]
- Van Goor, F.; Straley, K.S.; Cao, D.; González, J.; Hadida, S.; Hazlewood, A.; Joubran, J.; Knapp, T.; Makings, L.R.; Miller, M.; et al. Rescue of ΔF508-CFTR trafficking and gating in human cystic fibrosis airway primary cultures by small molecules. Am. J. Physiol. Cell. Mol. Physiol. 2006, 290, 1117–1130. [Google Scholar] [CrossRef]
- Van Goor, F.; Hadida, S.; Grootenhuis, P.D.J.; Burton, B.; Stack, J.H.; Straley, K.S.; Decker, C.J.; Miller, M.; McCartney, J.; Olson, E.R.; et al. Correction of the F508del-CFTR protein processing defect in vitro by the investigational drug VX-809. Proc. Natl. Acad. Sci. USA 2011, 108, 18843–18848. [Google Scholar] [CrossRef] [Green Version]
- Nocquet, P.A.; Hensienne, R.; Wencel-Delord, J.; Laigre, E.; Sidelarbi, K.; Becq, F.; Norez, C.; Hazelard, D.; Compain, P. Pushing the limits of catalytic C-H amination in polyoxygenated cyclobutanes. Org. Biomol. Chem. 2016, 14, 2780–2796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nocquet, P.A.; Hensienne, R.; Wencel-Delord, J.; Wimmer, E.; Hazelard, D.; Compain, P. Synthesis of a new class of iminosugars based on constrained azaspirocyclic scaffolds by way of catalytic C-H amination. Org. Biomol. Chem. 2015, 13, 9176–9180. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, M.; Hensienne, R.; Kern, N.; Tardieu, D.; Bodlenner, A.; Hazelard, D.; Compain, P. Stereocontrolled synthesis of polyhydroxylated bicyclic azetidines as a new class of iminosugars. Org. Biomol. Chem. 2018, 16, 4688–4700. [Google Scholar] [CrossRef] [PubMed]
- Heightman, T.D.; Vasella, A.T. Recent Insights into Inhibition, Structure, and Mechanism of Configuration-Retaining Glycosidases. Angew. Chem. Int. Ed. 1999, 38, 750–770. [Google Scholar] [CrossRef]
- Nocquet, P.A.; Hazelard, D.; Gruntz, G.; Compain, P. Synthesis of 4-membered carbasugars by way of stereoselective SmI 2-mediated aldehyde-alkene cyclization. J. Org. Chem. 2013, 78, 6751–6757. [Google Scholar] [CrossRef]
- Du Bois, J. Rhodium-catalyzed C-H amination. an enabling method for chemical synthesis. Org. Process Res. Dev. 2011, 15, 758–762. [Google Scholar] [CrossRef] [Green Version]
- Hazelard, D.; Nocquet, P.A.; Compain, P. Catalytic C-H amination at its limits: Challenges and solutions. Org. Chem. Front. 2017, 4, 2500–2521. [Google Scholar] [CrossRef]
- Morís-Varas, F.; Qian, X.H.; Wong, C.H. Enzymatic/chemical synthesis and biological evaluation of seven-membered iminocyclitols. J. Am. Chem. Soc. 1996, 118, 7647–7652. [Google Scholar] [CrossRef]
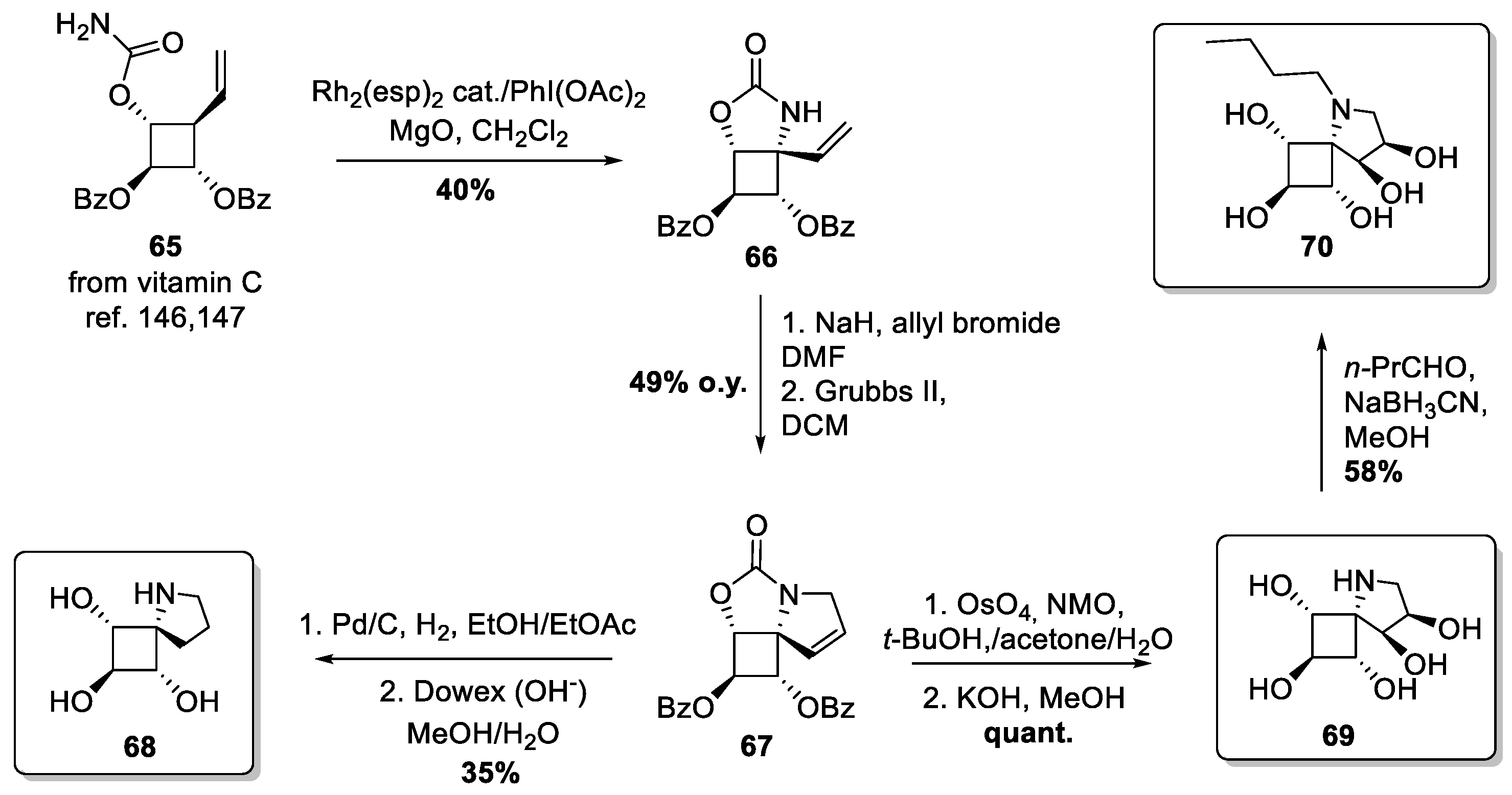
- Orwig, S.D.; Tan, Y.L.; Grimster, N.P.; Yu, Z.; Powers, E.T.; Kelly, J.W.; Lieberman, R.L. Binding of 3,4,5,6-tetrahydroxyazepanes to the acid-β-glucosidase active site: Implications for pharmacological chaperone design for Gaucher disease. Biochemistry 2011, 50, 10647–10657. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.-N.; Twigg, G.; Butters, T.D.; Zhang, S.; Zhang, L.; Zhang, L.-H.; Ye, X.-S. Synthesis of N-substituted ε-hexonolactams as pharmacological chaperones for the treatment of N370S mutant Gaucher disease. Org. Biomol. Chem. 2012, 10, 2923. [Google Scholar] [CrossRef] [PubMed]
- Désiré, J.; Mondon, M.; Fontelle, N.; Nakagawa, S.; Hirokami, Y.; Adachi, I.; Iwaki, R.; Fleet, G.W.J.; Alonzi, D.S.; Twigg, G.; et al. N- and C-alkylation of seven-membered iminosugars generates potent glucocerebrosidase inhibitors and F508del-CFTR correctors. Org. Biomol. Chem. 2014, 12, 8977–8996. [Google Scholar] [CrossRef] [PubMed]
- Mondon, M.; Fontelle, N.; Désiré, J.; Lecornué, F.; Guillard, J.; Marrot, J.; Blériot, Y. Access to L-and D-iminosugar C-glycosides from a D-gluco-derived 6-azidolactol exploiting a ring isomerization/alkylation strategy. Org. Lett. 2012, 14, 870–873. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, Y.; Vogel, P.; Sinaÿ, P.; Blériot, Y. Tandem Staudinger–azaWittig mediated ring expansion: Rapid access to new isofagomine-tetrahydroxyazepane hybrids. Chem. Commun. 2007, 183–185. [Google Scholar] [CrossRef] [PubMed]
- Boinot, C.; Souchet, M.J.; Ferru-Clément, R.; Becq, F. Searching for combinations of small-molecule correctors to restore F508del-cystic fibrosis transmembrane conductance regulator function and processing. J. Pharmacol. Exp. Ther. 2014, 350, 624–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veit, G.; Xu, H.; Dreano, E.; Avramescu, R.G.; Bagdany, M.; Beitel, L.K.; Roldan, A.; Hancock, M.A.; Lay, C.; Li, W.; et al. Structure-guided combination therapy to potently improve the function of mutant CFTRs. Nat. Med. 2018, 24, 1732–1742. [Google Scholar] [CrossRef]
- Rowe, S.M.; Verkman, A.S. Cystic Fibrosis Transmembrane Regulator Correctors and Potentiators. Cold Spring Harb. Perspect. Med. 2013, 3, a009761. [Google Scholar] [CrossRef] [Green Version]
- Hanrahan, J.W.; Matthes, E.; Carlile, G.; Thomas, D.Y. Corrector combination therapies for F508del-CFTR. Curr. Opin. Pharmacol. 2017, 34, 105–111. [Google Scholar] [CrossRef]
- Hutt, D.M.; Herman, D.; Rodrigues, A.P.C.; Noel, S.; Pilewski, J.M.; Matteson, J.; Hoch, B.; Kellner, W.; Kelly, J.W.; Schmidt, A.; et al. Reduced histone deacetylase 7 activity restores function to misfolded CFTR in cystic fibrosis. Nat. Chem. Biol. 2010, 6, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Cutting, G.R. Cystic fibrosis genetics: From molecular understanding to clinical application. Nat. Rev. Genet. 2015, 16, 45–56. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, B.P.; Freedman, S.D. Cystic fibrosis. Lancet 2009, 373, 1891–1904. [Google Scholar] [CrossRef]
- Nichols, D.P.; Chmiel, J.F. Inflammation and its genesis in cystic fibrosis. Pediatr. Pulmonol. 2015, 50, S39–S56. [Google Scholar] [CrossRef]
- Perrem, L.; Ratjen, F. Anti-inflammatories and mucociliary clearance therapies in the age of CFTR modulators. Pediatr. Pulmonol. 2019, 54, S46–S55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiumarini, D.; Loberto, N.; Mancini, G.; Bassi, R.; Giussani, P.; Chiricozzi, E.; Samarani, M.; Munari, S.; Tamanini, A.; Cabrini, G.; et al. Evidence for the Involvement of Lipid Rafts and Plasma Membrane Sphingolipid Hydrolases in Pseudomonas aeruginosa Infection of Cystic Fibrosis Bronchial Epithelial Cells. Mediat. Inflamm. 2017, 2017, 1730245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aureli, M.; Schiumarini, D.; Loberto, N.; Bassi, R.; Tamanini, A.; Mancini, G.; Tironi, M.; Munari, S.; Cabrini, G.; Dechecchi, M.C.; et al. Unravelling the role of sphingolipids in cystic fibrosis lung disease. Chem. Phys. Lipids 2016, 200, 94–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, A.; Issler, A.C.; Cotton, C.U.; Kelley, T.J.; Verkman, A.S.; Davis, P.B. CFTR inhibition mimics the cystic fibrosis inflammatory profile. Am. J. Physiol. Cell. Mol. Physiol. 2007, 292, 383–395. [Google Scholar] [CrossRef] [Green Version]
- Chiricozzi, E.; Loberto, N.; Schiumarini, D.; Samarani, M.; Mancini, G.; Tamanini, A.; Lippi, G.; Dechecchi, M.C.; Bassi, R.; Giussani, P.; et al. Sphingolipids role in the regulation of inflammatory response: From leukocyte biology to bacterial infection. J. Leukoc. Biol. 2018, 103, 445–456. [Google Scholar] [CrossRef]
- Dechecchi, M.C.; Nicolis, E.; Mazzi, P.; Paroni, M.; Cioffi, F.; Tamanini, A.; Bezzerri, V.; Tebon, M.; Lampronti, I.; Huang, S.; et al. Pharmacological Modulators of Sphingolipid Metabolism for the Treatment of Cystic Fibrosis Lung Inflammation. In Cystic Fibrosis—Renewed Hopes Through Research; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Loberto, N.; Tebon, M.; Lampronti, I.; Marchetti, N.; Aureli, M.; Bassi, R.; Giri, M.G.; Bezzerri, V.; Lovato, V.; Cantù, C.; et al. GBA2-Encoded β-Glucosidase Activity Is Involved in the Inflammatory Response to Pseudomonas aeruginosa. PLoS ONE 2014, 9, e104763. [Google Scholar] [CrossRef] [Green Version]
- Samarani, M.; Loberto, N.; Soldà, G.; Straniero, L.; Asselta, R.; Duga, S.; Lunghi, G.; Zucca, F.A.; Mauri, L.; Ciampa, M.G.; et al. A lysosome–plasma membrane–sphingolipid axis linking lysosomal storage to cell growth arrest. FASEB J. 2018, 32, 5685–5702. [Google Scholar] [CrossRef]
- Ghisaidoobe, A.T.; Van Den Berg, R.J.B.H.N.; Butt, S.S.; Strijland, A.; Donker-Koopman, W.E.; Scheij, S.; Van Den Nieuwendijk, A.M.C.H.; Koomen, G.J.; Van Loevezijn, A.; Leemhuis, M.; et al. Identification and development of biphenyl substituted iminosugars as improved dual glucosylceramide synthase/neutral glucosylceramidase inhibitors. J. Med. Chem. 2014, 57, 9096–9104. [Google Scholar] [CrossRef]
- Munari, S.; Loberto, N.; Aureli, M.; Baron, A.; Guisot, N.; Bassi, R.; Tironi, M.; Giri, M.G.; Tamanini, A.; Lippi, G.; et al. Neoglycoconjugates Derived from Deoxynojirimycin as Possible Therapeutic Agents for Cystic Fibrosis Lung Disease, by Modulation of the Sphingolipid Metabolism. JSM Genet Genom. 2016, 3, 2–7. [Google Scholar]
- Rountree, J.S.S.; Butters, T.D.; Wormald, M.R.; Boomkamp, S.D.; Dwek, R.A.; Asano, N.; Ikeda, K.; Evinson, E.L.; Nash, R.J.; Fleet, G.W.J. Design, synthesis, and biological evaluation of enantiomeric β-N-acetylhexosaminidase inhibitors LABNAc and DABNAc as potential agents against Tay-Sachs and Sandhoff disease. ChemMedChem 2009, 4, 378–392. [Google Scholar] [CrossRef] [PubMed]
- Lahav, D.; Liu, B.; Van Den Berg, R.J.B.H.N.; Van Den Nieuwendijk, A.M.C.H.; Wennekes, T.; Ghisaidoobe, A.T.; Breen, I.; Ferraz, M.J.; Kuo, C.L.; Wu, L.; et al. A Fluorescence Polarization Activity-Based Protein Profiling Assay in the Discovery of Potent, Selective Inhibitors for Human Nonlysosomal Glucosylceramidase. J. Am. Chem. Soc. 2017, 139, 14192–14197. [Google Scholar] [CrossRef] [PubMed]
- De Fenza, M.; D’Alonzo, D.; Esposito, A.; Munari, S.; Loberto, N.; Santangelo, A.; Lampronti, I.; Tamanini, A.; Rossi, A.; Ranucci, S.; et al. Exploring the effect of chirality on the therapeutic potential of N-alkyl-deoxyiminosugars: Anti-inflammatory response to Pseudomonas aeruginosa infections for application in CF lung disease. Eur. J. Med. Chem. 2019, 175, 63–71. [Google Scholar] [CrossRef] [PubMed]
- D’Alonzo, D.; Amato, J.; Schepers, G.; Froeyen, M.; Van Aerschot, A.; Herdewijn, P.; Guaragna, A. Enantiomeric selection properties of β-homoDNA: Enhanced pairing for heterochiral complexes. Angew. Chem. Int. Ed. 2013, 52, 6662–6665. [Google Scholar] [CrossRef]
- Paolella, C.; D’Alonzo, D.; Palumbo, G.; Guaragna, A. Sulfur-assisted domino access to bicyclic dihydrofurans: Case study and early synthetic applications. Org. Biomol. Chem. 2013, 11, 7825–7829. [Google Scholar] [CrossRef]
- D’Alonzo, D.; Froeyen, M.; Schepers, G.; Di Fabio, G.; Van Aerschot, A.; Herdewijn, P.; Palumbo, G.; Guaragna, A. 1′,5′-Anhydro-L-ribo-hexitol Adenine Nucleic Acids (α-L-HNA-A): Synthesis and Chiral Selection Properties in the Mirror Image World. J. Org. Chem. 2015, 80, 5014–5022. [Google Scholar] [CrossRef]
- Caputo, R.; Ciriello, U.; Festa, P.; Guaragna, A.; Palumbo, G.; Pedatella, S. Stereoselective synthesis of fully protected (S)-1,7-dioxaspiro [5,5]undec-4-ene derivatives of sugars. Eur. J. Org. Chem. 2003, 2003, 2617–2621. [Google Scholar] [CrossRef]
- Caputo, R.; De Nisco, M.; Festa, P.; Guaragna, A.; Palumbo, G.; Pedatella, S. Synthesis of 4-deoxy-L-and D-hexoses from chiral noncarbohydrate building blocks. J. Org. Chem. 2004, 69, 7033–7037. [Google Scholar] [CrossRef]
- D’Alonzo, D.; Guaragna, A.; Van Aerschot, A.; Herdewijn, P.; Palumbo, G. Toward L-Homo-DNA: Stereoselective de Novo Synthesis of β-L-erythro-Hexopyranosyl Nucleosides. J. Org. Chem. 2010, 75, 6402–6410. [Google Scholar] [CrossRef]
- Guaragna, A.; D’Errico, S.; D’Alonzo, D.; Pedatella, S.; Palumbo, G. A general approach to the synthesis of 1-deoxy-L-iminosugars. Org. Lett. 2007, 9, 3473–3476. [Google Scholar] [CrossRef] [PubMed]
- Drewry, D.H.; Coe, D.M.; Poon, S. Solid-supported reagents in organic synthesis. Med. Res. Rev. 1999, 19, 97–148. [Google Scholar] [CrossRef]
- Caputo, R.; Cecere, G.; Guaragna, A.; Palumbo, G.; Pedatella, S. β-Amino-α-hydroxy Esters by Asymmetric Hydroxylation ofhomo-β-Amino Acid Esters. Eur. J. Org. Chem. 2002, 17, 3050–3054. [Google Scholar] [CrossRef]





































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
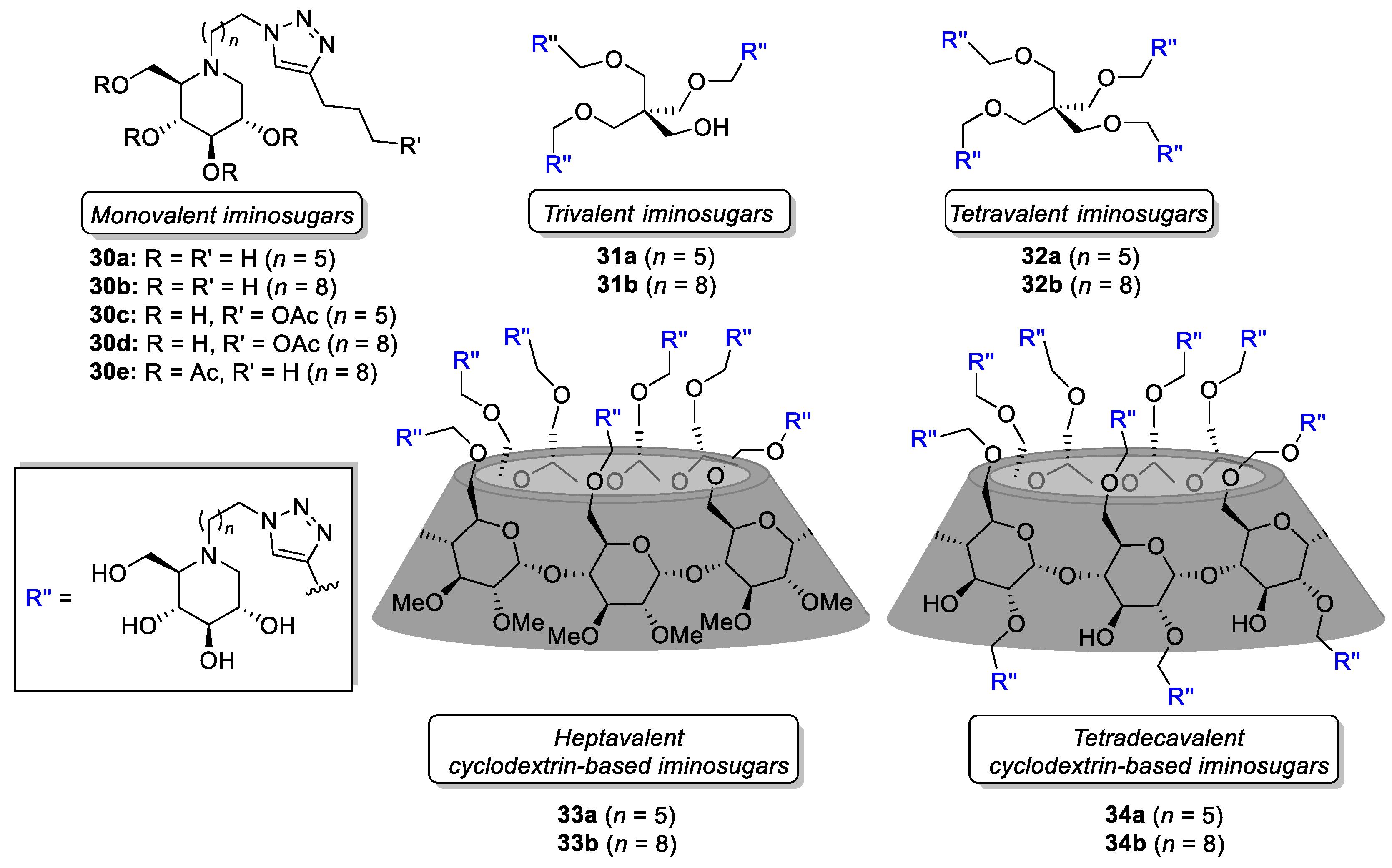
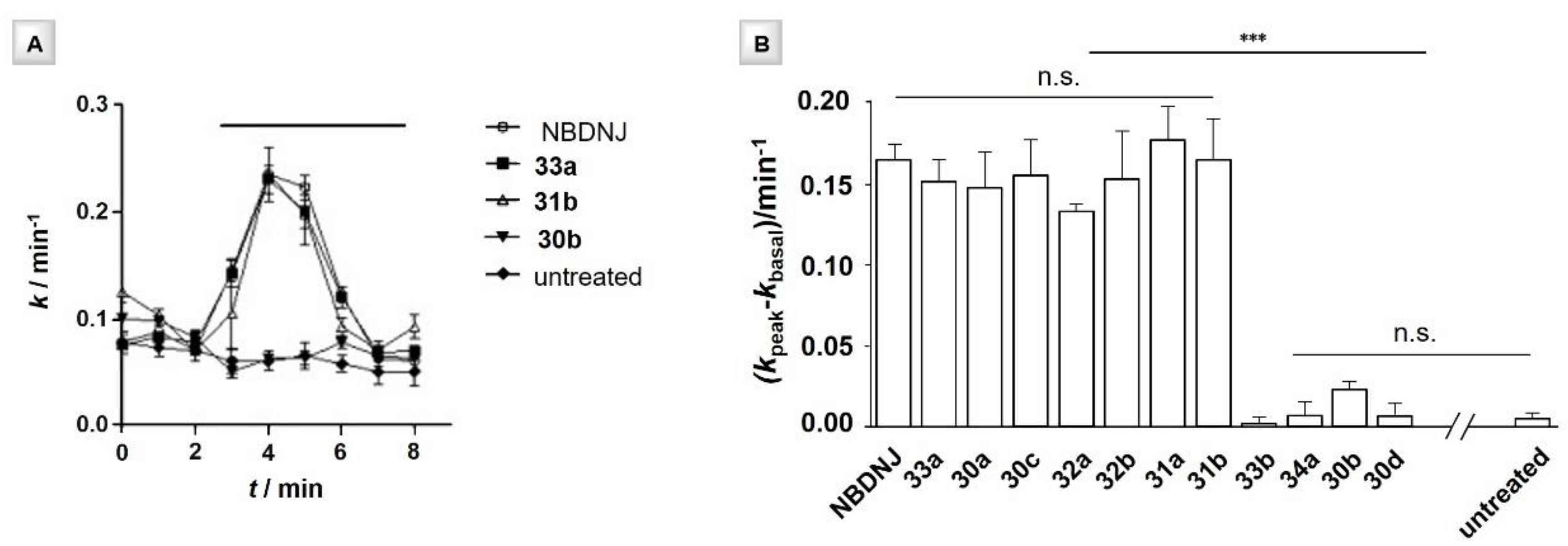
| Entry | Iminosugar | Valency | Spacer Length | EC50 (μM) | EC50 (mg·L−1) |
|---|---|---|---|---|---|
| 1 | NBDNJ | 1 | - | 112 | 24.6 |
| 2 | 30a | 1 | C6 | 12 | 4.3 |
| 3 | 30c | 1 | C6 | 20 | 8.3 |
| 4 | 31a | 3 | C6 | 1 | 1.1 |
| 5 | 32a | 4 | C6 | 110 | 158.6 |
| 6 | 33a | 7 | C6 | 71 | 256.7 |
| 7 | 30b | 1 | C9 | 458 | 182.5 |
| 8 | 31b | 3 | C9 | 0.5 | 0.6 |
| 9 | 32b | 4 | C9 | 49 | 78.9 |
| 10 | 33b | 7 | C9 | -b | -b |
 | |||
|---|---|---|---|
| Entry | Iminosugar | IC50 (nM) | M.I. (%) |
| 1 | NBDNJ (4) | 1900 | 51.5 |
| 2 | AMP-DNM (16) | 2 | 46 |
| 3 | 17 | 12 | 43 |
| 4 | 18 | 5 | 23 |
| 5 | 19 | 45 | 36 |
| 6 | 20 | 35 | 40 |
| 7 | 21 | 620 | 36 |
| 8 | 22 | 7 | 36 |
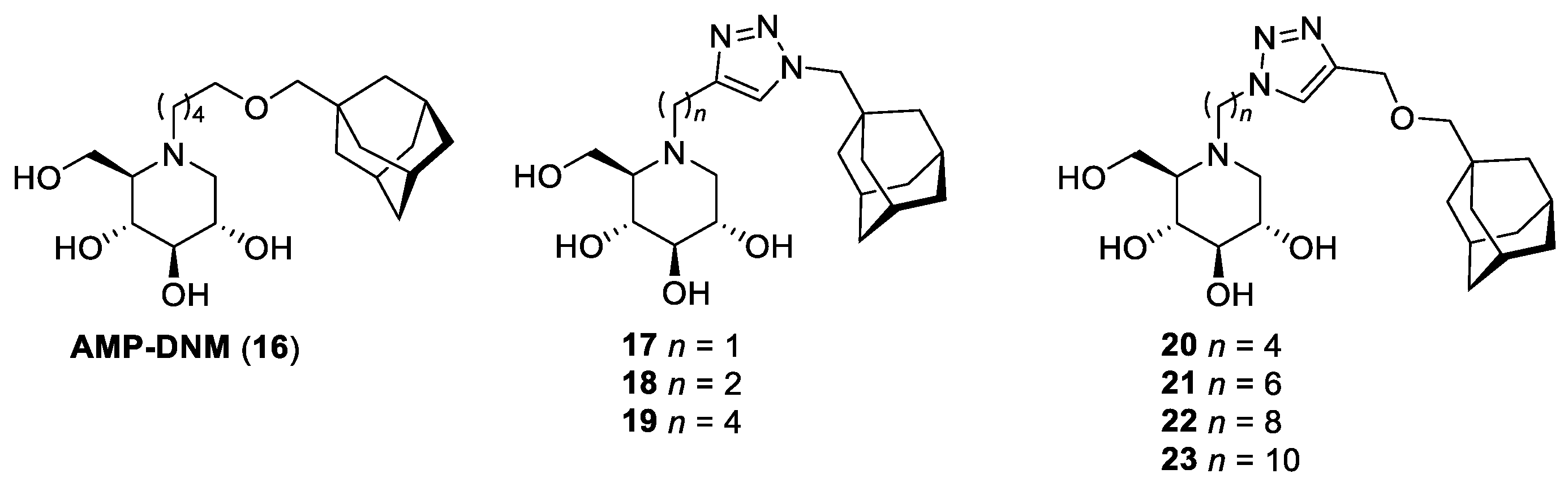
| 9 | 23 | 2 | 28 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esposito, A.; D’Alonzo, D.; De Fenza, M.; De Gregorio, E.; Tamanini, A.; Lippi, G.; Dechecchi, M.C.; Guaragna, A. Synthesis and Therapeutic Applications of Iminosugars in Cystic Fibrosis. Int. J. Mol. Sci. 2020, 21, 3353. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093353
Esposito A, D’Alonzo D, De Fenza M, De Gregorio E, Tamanini A, Lippi G, Dechecchi MC, Guaragna A. Synthesis and Therapeutic Applications of Iminosugars in Cystic Fibrosis. International Journal of Molecular Sciences. 2020; 21(9):3353. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093353
Chicago/Turabian StyleEsposito, Anna, Daniele D’Alonzo, Maria De Fenza, Eliana De Gregorio, Anna Tamanini, Giuseppe Lippi, Maria Cristina Dechecchi, and Annalisa Guaragna. 2020. "Synthesis and Therapeutic Applications of Iminosugars in Cystic Fibrosis" International Journal of Molecular Sciences 21, no. 9: 3353. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093353