Zinc and Copper Enhance Cucumber Tolerance to Fusaric Acid by Mediating Its Distribution and Toxicity and Modifying the Antioxidant System


 ,
,
Abstract
:1. Introduction
2. Results
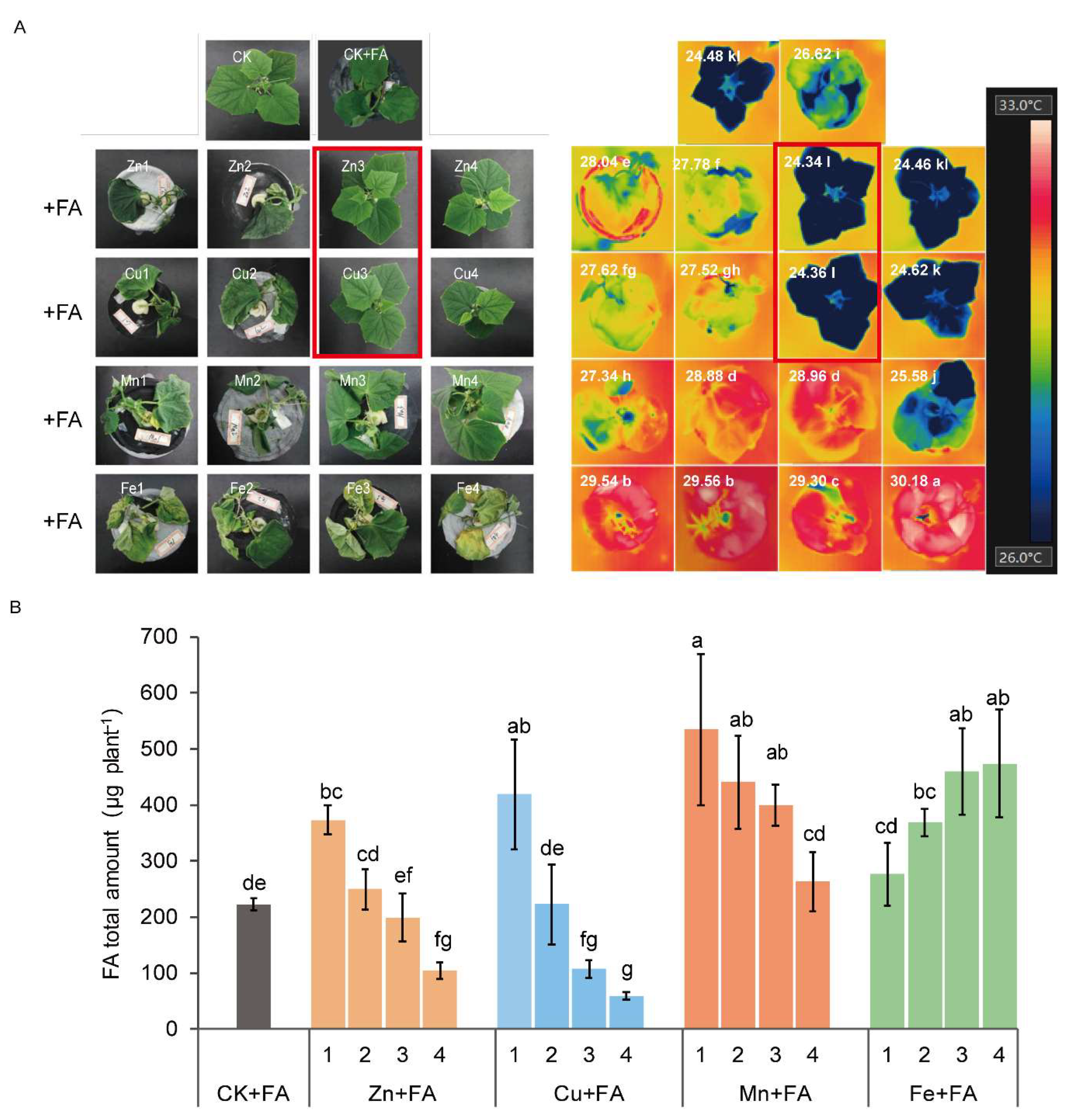
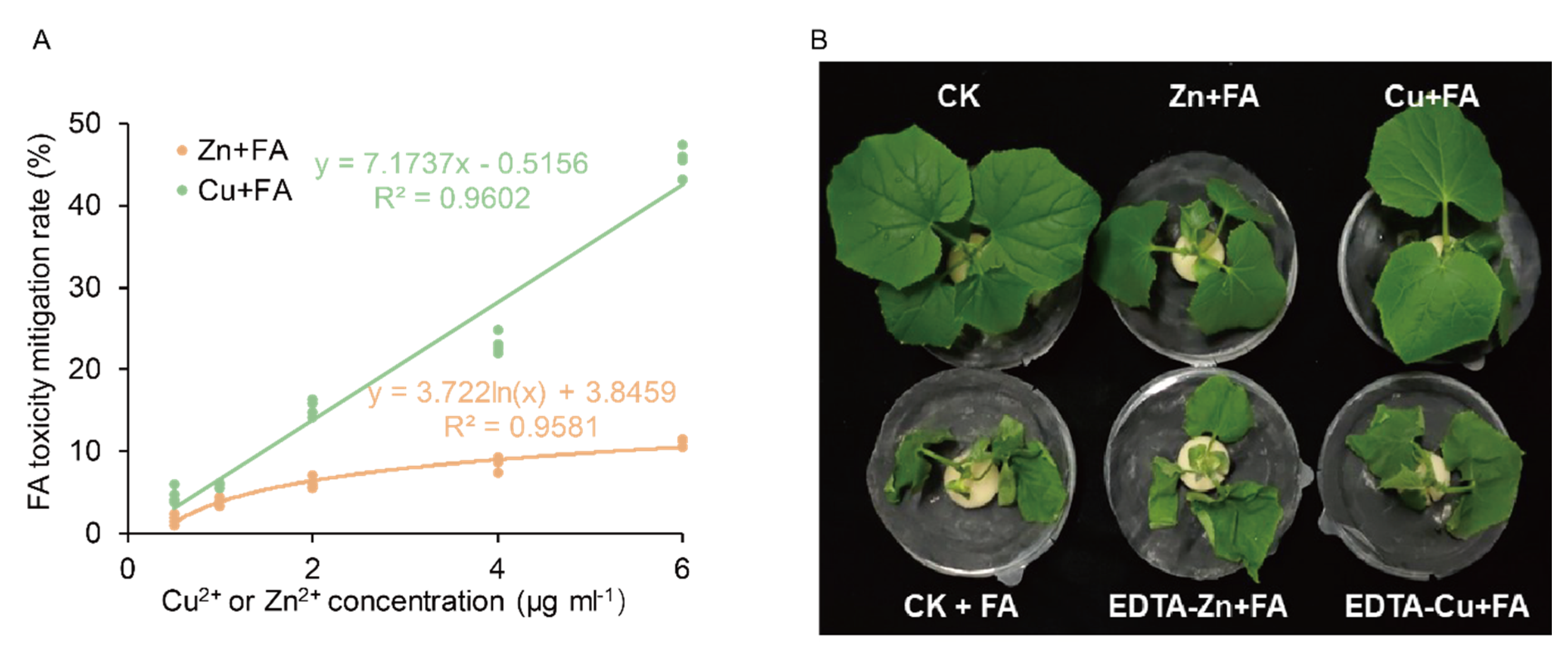
2.1. FA Toxicity in Cucumber Plants is Reversed by the Exogenous Addition of Zn and Cu
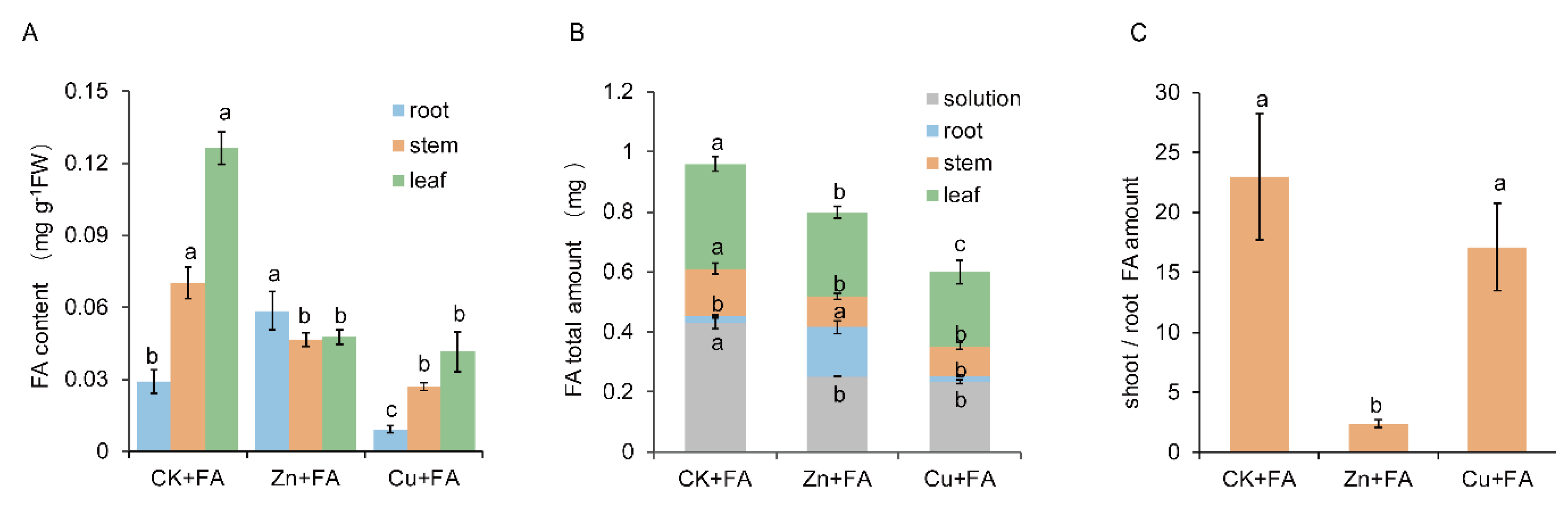
2.2. Additional Zn or Cu Improves the Tolerance of Cucumber to FA by Mediating Its Distribution
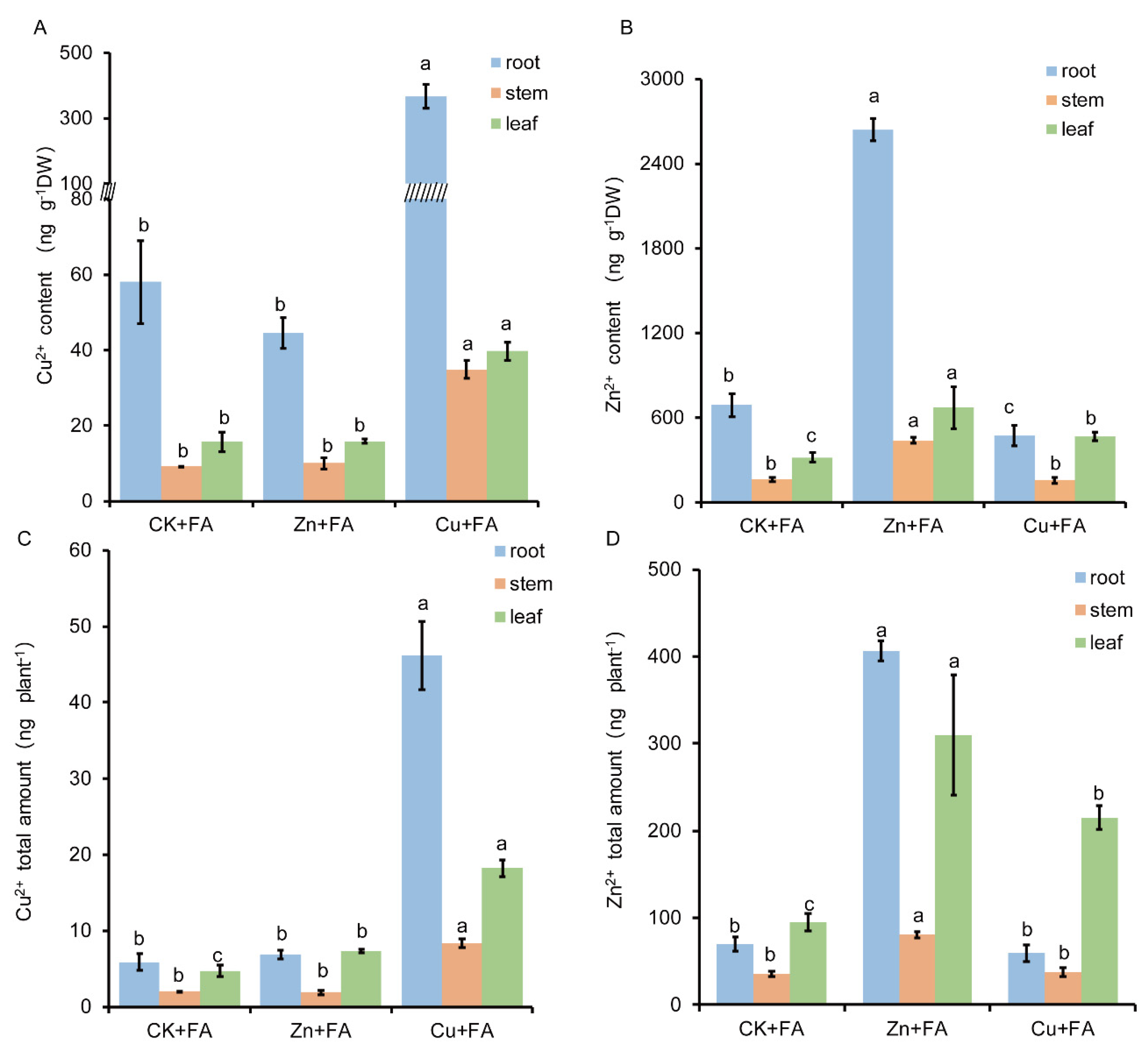
2.3. FA is Detoxified through Direct Reaction with Zn or Cu
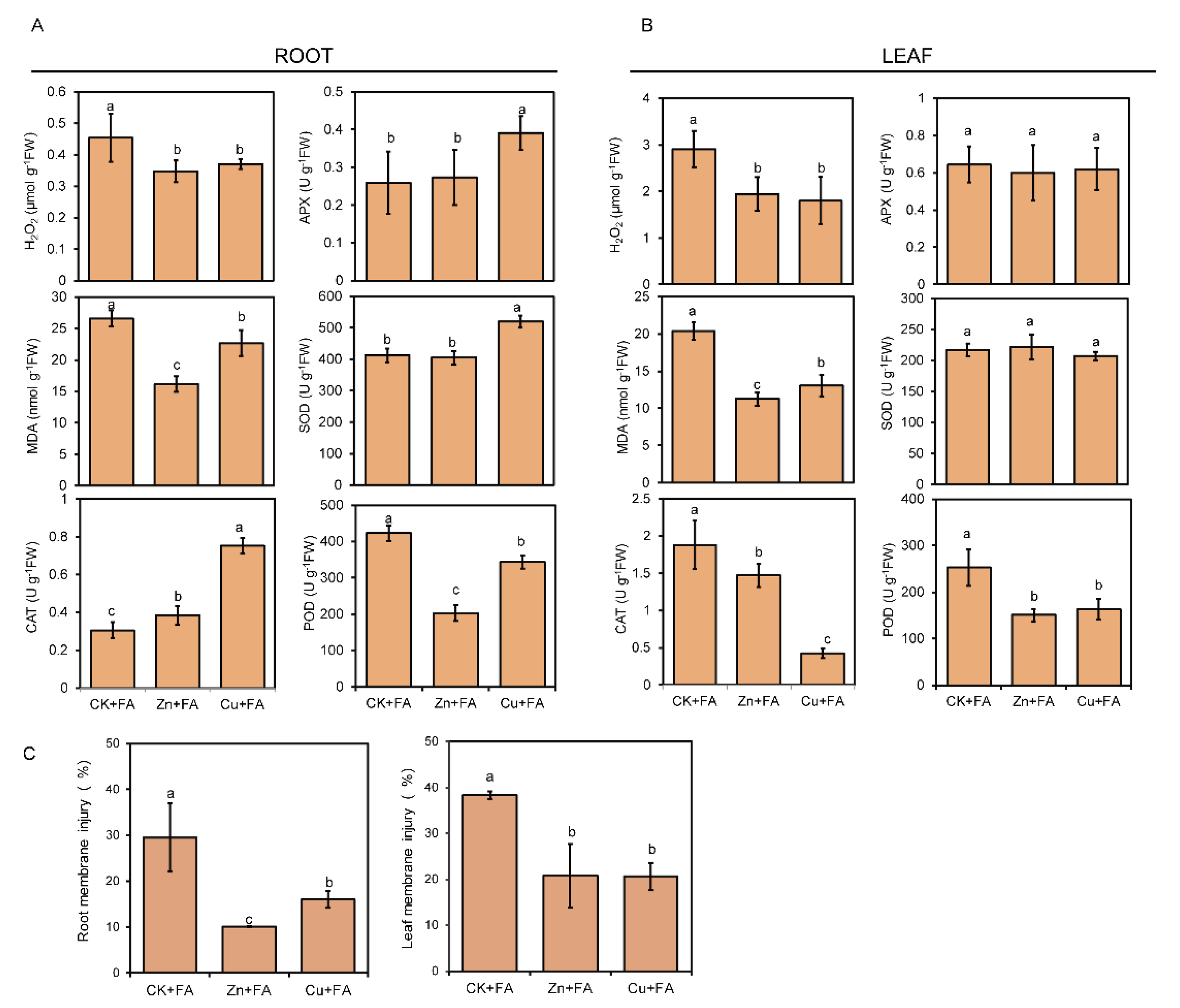
2.4. Additional Zn or Cu Alleviates the Oxidative Stress Caused by FA
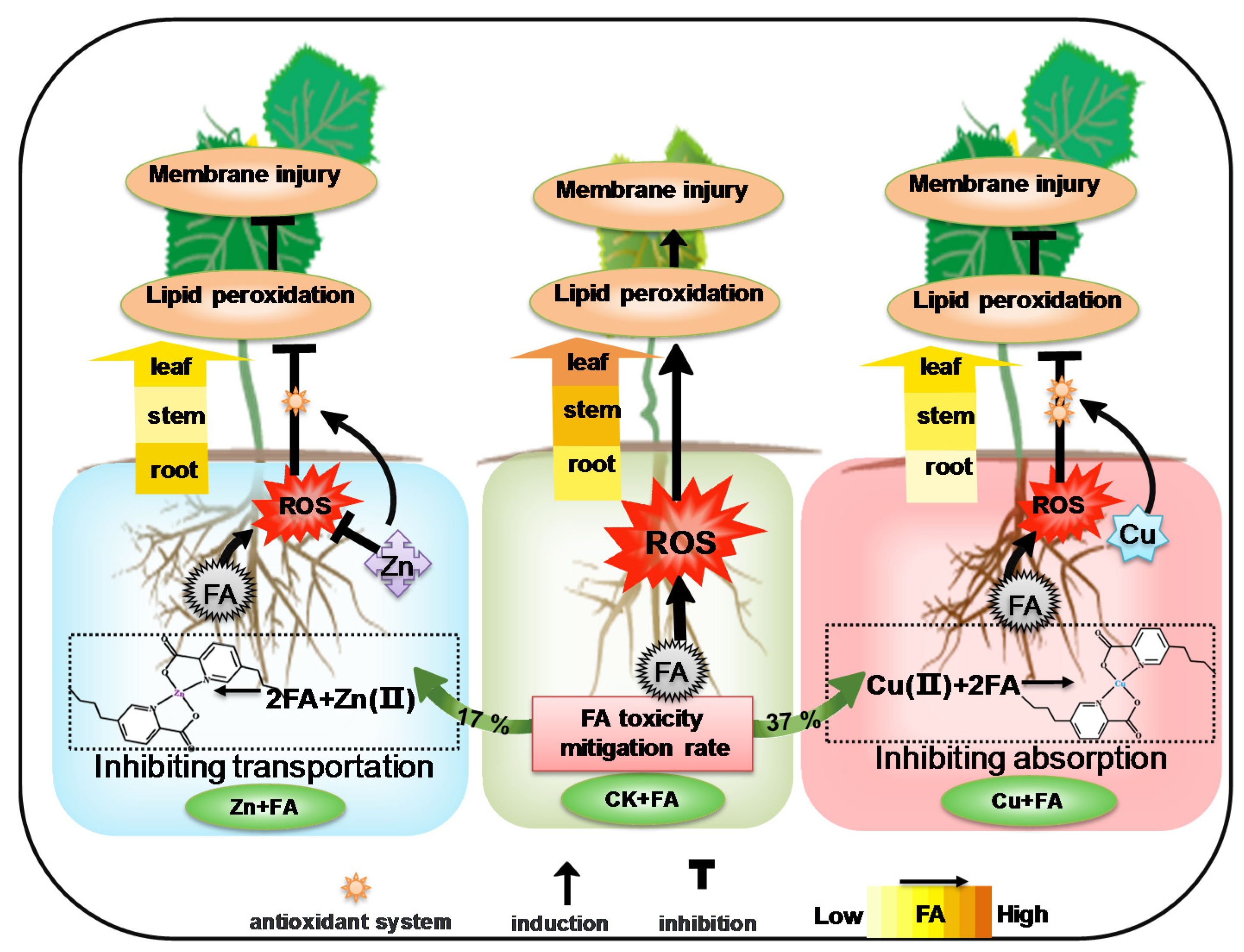
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Thermal Imaging
4.3. Wilting Index
4.4. The Leaf and Root Membrane Injury
4.5. Gas Exchange Measurements
4.6. Determination of Zn and Cu Concentration
4.7. FA Detection in Plants and Nutrient Solution
4.8. Toxicity Mitigation Rate
4.9. In Vitro Toxicity Mitigation
4.10. Quantitative Determination of Hydrogen Peroxide, Lipid Peroxidation, Antioxidant Enzyme Activity, Lignin and Cellulose
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di, P.A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 804. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Xiong, Y.F.; Ling, N.; Shen, Q.R.; Guo, S.W. Fusaric acid accelerates the senescence of leaf in banana when infected by Fusarium. World J. Microb. Biot. 2014, 30, 1399. [Google Scholar] [CrossRef] [PubMed]
- Niehaus, E.M.; Díaz-Sánchez, V.; Bargen, K.W.V.; Kleigrewe, K.; Humpf, H.U.; Limón, M.C.; Tudzynski, B. Fusarins and fusaric acid in Fusaria. In Biosynthesis and Molecular Genetics of Fungal Secondary Metabolites; Martín, J.F., García-Estrada, C., Eds.; Springer: New York, NY, USA, 2014; pp. 239–262. [Google Scholar]
- Stipanovic, R.D.; Puckhaber, L.S.; Liu, J.; Bell, A.A. Phytotoxicity of fusaric acid and analogs to cotton. Toxicon 2011, 57, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Venter, S.L.; Steyn, P.J. Correlation between fusaric acid production and virulence of isolates of Fusarium oxysporum that causes potato dry rot in South Africa. Potato Res. 1998, 41, 289–294. [Google Scholar] [CrossRef]
- Jiao, J.; Sun, L.; Zhou, B.; Gao, Z.; Yu, H.; Zhu, X.; Liang, Y. Hydrogen peroxide production and mitochondrial dysfunction contribute to the fusaric acid-induced programmed cell death in tobacco cells. J. Plant Physiol. 2014, 171, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.S.; Yin, X.M.; Liu, D.Y.; Ling, N.; Bao, W.; Ying, R.R.; Zhu, Y.Y.; Guo, S.W.; Shen, Q.R. Effect of fungal fusaric acid on the root and leaf physiology of watermelon (Citrullus lanatus) seedlings. Plant. Soil 2008, 308, 255–266. [Google Scholar] [CrossRef]
- Samadi, L.; Shahsavan, B.B. Fusaric acid induces apoptosis in saffron root-tip cells: Roles of caspase-like activity, cytochrome C, and H2O2. Planta 2006, 225, 223–234. [Google Scholar] [CrossRef]
- Dong, X.; Ling, N.; Wang, M.; Shen, Q.R.; Guo, S.W. Fusaric acid is a crucial factor in the disturbance of leaf water imbalance in Fusarium-infected banana plants. Plant Physiol. Bioch. 2012, 60, 171–179. [Google Scholar] [CrossRef]
- Ruiz, J.A.; Bernar, E.M.; Jung, K. Production of siderophores increases resistance to fusaric acid in Pseudomonas protegens Pf-5. PLoS ONE 2015, 10, e0117040. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Ling, N.; Dong, X.; Zhu, J.Y.; Shen, Q.R.; Guo, S.W. Thermographic visualization of leaf response in cucumber plants infected with the soil-borne pathogen Fusarium oxysporum f. sp. Cucumerinum. Plant Physiol. Bioch. 2012, 61, 153. [Google Scholar] [CrossRef]
- Wang, M.; Sun, Y.M.; Sun, G.M.; Liu, X.; Zhai, L.; Shen, Q.R.; Guo, S.W. Water balance altered in cucumber plants infected with Fusarium oxysporum f. sp. Cucumerinum. Sci. Rep. UK 2015, 5, 7722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czymmek, K.J.; Fogg, M.; Powell, D.H.; Sweigard, J.; Park, S.Y.; Kang, S. In vivo time-lapse documentation using confocal and multi-photon microscopy reveals the mechanisms of invasion into the Arabidopsis root vascular system by Fusarium oxysporum. Fungal Genet. Biol. 2007, 44, 1011–1023. [Google Scholar] [CrossRef]
- Oubraim, S.; Sedra, M.H.; Lazrek, H.B. A relationship between Bayoud disease severity and toxin susceptibility of date palm cultivars. Emir. J. Food Agric. 2016, 28, 45. [Google Scholar] [CrossRef]
- D’Alton, A.; Etherton, B. Effects of fusaric acid on tomato root hair membrane potentials and ATP levels. Plant Physiol. 1984, 74, 39–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Ling, N.; Dong, X.; Liu, X.; Shen, Q.; Guo, S. Effect of fusaric acid on the leaf physiology of cucumber seedlings. Eur. J. Plant Pathol. 2014, 138, 103–112. [Google Scholar] [CrossRef]
- Yang, S.S. Studies on cross protection of fusarium wilt of cucumber-(4)-protective effect by a nonpathogenic isolate of Fusarium oxysporum in a greenhouse and fields. Korean J. Plant Pathol. 1996, 10, 29–33. [Google Scholar]
- Boyd, R. Elemental defenses of plants by metals. Nat. Educ. Knowl. 2010, 3, 57. [Google Scholar]
- Boyd, R.S. Plant defense using toxic inorganic ions: Conceptual models of the defensive enhancement and joint effects hypotheses. Plant Sci. 2012, 195, 88. [Google Scholar] [CrossRef]
- Cabot, C.; Martos, S.; Llugany, M.; Gallego, B.; Tolrà, R.; Poschenrieder, C. A role for zinc in plant defense against pathogens and herbivores. Front. Plant Sci. 2019, 10, 1171. [Google Scholar] [CrossRef]
- Cobbett, C. Heavy metals and plants-model systems and hyperaccumulators. New Phytol. 2003, 159, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Poschenrieder, C.; Roser, T.; Juan, B. Can metals defend plants against biotic stress? Trends Plant Sci. 2006, 11, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Rolke, Y.; Liu, S.; Quidde, T.; Williamson, B.; Schouten, A.; Weltring, K.M.; Siewers, V.; Tenberge, K.B.; Tudzynski, B.; Tudzynski, P. Functional analysis of H2O2-generating systems in B. cinerea: The major Cu-Zn SOD (BCSOD1) has impact on virulence on bean, whereas a glucose oxidase (BCGOD1) is dispensable. Mol. Plant Pathol. 2004, 5, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Stolpe, C.; Giehren, F.; Krämer, U.; Müller, C. Both heavy metal-amendment of soil and aphid-infestation increase Cd and Zn concentrations in phloem exudates of a metal-hyperaccumulating plant. Phytochemistry 2017, 139, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Franza, T.; Mahé, B.; Expert, D. Erwinia chrysanthemi requires a second iron transport route dependent of the siderophore achromobactin for extracellular growth and plant infection. Mol. Microbiol. 2005, 55, 261. [Google Scholar] [CrossRef] [PubMed]
- Babitha, M.P.; Bhat, S.G.; Prakash, H.S.; Shetty, H.S. Differential induction of superoxide dismutase in downy mildew-resistant and -susceptible genotypes of pearl millet. Plant Pathol. 2002, 51, 480–486. [Google Scholar] [CrossRef] [Green Version]
- Parisot, D.; Dufresne, M.; Veneault, C.; Laugé, R.; Langin, T. Clap1, a gene encoding a copper-transporting ATPase involved in the process of infection by the phytopathogenic fungus Colletotrichum lindemuthianum. Mol. Genet. Genom. 2002, 268, 139–151. [Google Scholar] [CrossRef]
- Fones, H.; Davis, C.A.R.; Rico, A.; Fang, F.; Smith, J.A.C.; Preston, G.M. Metal hyperaccumulation armors plants against disease. PLoS Pathog. 2010, 6, e1001093. [Google Scholar] [CrossRef]
- Fones, H.; Preston, G.M. The impact of transition metals on bacterial plant disease. FEMS Microbiol. Rev. 2013, 37, 495–519. [Google Scholar] [CrossRef] [Green Version]
- Gallego, B.; Martos, S.; Cabot, C.; Barceló, J.; Poschenrieder, C. Zinc hyperaccumulation substitutes for defense failures beyond salicylate and jasmonate signaling pathways of Alternaria brassicicola attack in Noccaea caerulescens. Physiol. Plant. 2017, 159, 401–415. [Google Scholar] [CrossRef]
- Heckman, J.R.; Clarke, B.B.; Murphy, J.A. Optimizing manganese fertilization for the suppression of take-all patch disease on creeping bentgrass. Crop Sci. 2003, 43, 1395–1398. [Google Scholar] [CrossRef]
- Simoglou, K.B.; Dordas, C. Effect of foliar applied boron, manganese and zinc on tan spot in winter durum wheat. Crop Prot. 2006, 25, 657–663. [Google Scholar] [CrossRef]
- Mithöfer, A.; Schulze, B.; Boland, W. Biotic and heavy metal stress response in plants: Evidence for common signals. FEBS Lett. 2004, 566, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Martos, S.; Gallego, B.; Cabot, C.; Llugany, M.; Barceló, J.; Poschenrieder, C. Zinc triggers signaling mechanisms and defense responses promoting resistance to Alternaria brassicicola in Arabidopsis thaliana. Plant Sci. 2016, 249, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.L.; Garcia, D.; Kim, D.; Hopf, A.; Salt, D.E. Constitutively elevated salicylic acid signals glutathione-mediated nickel tolerance in Thlaspi nickel hyperaccumulators. Plant Physiol. 2005, 137, 1082–1091. [Google Scholar] [CrossRef] [Green Version]
- Jonak, C.; Nakagami, H.; Hirt, H. Heavy metal stress. Activation of distinct mitogen-activated protein kinase pathways by copper and cadmium. Plant Physiol. 2004, 136, 3276–3283. [Google Scholar] [CrossRef] [Green Version]
- Mittra, B.; Ghosh, P.; Henry, S.L.; Mishra, J.; Das, T.K.; Ghosh, S.; Babu, C.R.; Mohanty, P. Novel mode of resistance to Fusarium infection by a mild dose pre-exposure of cadmium in wheat. Plant Physiol. Biochem. 2004, 42, 781–787. [Google Scholar] [CrossRef]
- Malini, S. Heavy metal chelates of fusaric acid: In vitro spectrophotometry. J. Phytopathol. 2010, 57, 221–231. [Google Scholar] [CrossRef]
- Pan, J.H.; Lin, Y.C.; Ni, T.; Gu, Y.C. Cu (II): A “signaling molecule” of the mangrove endophyte Fusarium oxysporum ZZF51? Biometals 2010, 23, 1053–1060. [Google Scholar] [CrossRef]
- Lakshminarayanan, K.; Subramanian, D. Is fusaric acid a vivotoxin? Nature 1955, 176, 697–698. [Google Scholar] [CrossRef]
- Tamari, K.; Kaji, J. Studies on the mechanism of injurious action of fusarinic acid on plant growth. J. Agric. Chem. Soc. Jpn. 1952, 26, 345–349. [Google Scholar]
- Yin, E.S.; Rakhmankulova, M.; Kucera, K.; Filho, J.G.D.S.; Portero, C.E.; Narváez-Trujillo, A.; Holley, S.A.; Strobel, S.A. Fusaric acid induces a notochord malformation in zebrafish via copper chelation. Biometals 2015, 28, 783–789. [Google Scholar] [CrossRef] [PubMed]
- López-Díaz, C.; Rahjoo, V.; Sulyok, M.; Ghionna, V.; Martín-Vicente, A.; Capilla, J.; Di, P.A.; López-Berges, M.S. Fusaric acid contributes to virulence of Fusarium oxysporum on plant and mammalian hosts. Mol. Plant Pathol. 2017, 19, 440–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.K.; Singh, H.B.; Upadhyay, R.S. Role of fusaric acid in the development of ’Fusarium wilt’ symptoms in tomato: Physiological, biochemical and proteomic perspectives. Plant Physiol. Biochem. 2017, 118, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Chawla, H.S.; Wenzel, G. In vitroselection for fusaric acid resistant barley plants. Plant Breed. 1987, 99, 159–163. [Google Scholar] [CrossRef]
- Jiang, R.F.; Ma, D.Y.; Zhao, F.J.; Mcgrath, S.P. Cadmium hyperaccumulation protects Thlaspi caerulescens from leaf feeding damage by thrips (Frankliniella occidentalis). New Phytol. 2010, 167, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Behmer, S.T.; Lloyd, C.M.; Raubenheimer, D.; Stewart-Clark, J.; Knight, J.; Leighton, R.S.; Harper, F.A.; Smith, J.A.C. Metal hyperaccumulation in plants: Mechanisms of defence against insect herbivores. Funct. Ecol. 2010, 19, 55–66. [Google Scholar] [CrossRef]
- Ent, A.V.D.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of metal and metalloid trace elements: Facts and fiction. Plant Soil 2013, 362, 319–334. [Google Scholar]
- Reeves, R.D. Tropical hyperaccumulators of metals and their potential for phytoextraction. Plant Soil 2003, 249, 57–65. [Google Scholar] [CrossRef]
- Saikia, R.; Varghese, S.; Singh, B.P.; Arora, D.K. Influence of mineral amendment on disease suppressive activity of Pseudomonas fluorescens to Fusarium wilt of chickpea. Microbiol. Res. 2009, 164, 365–373. [Google Scholar] [CrossRef]
- Marrè, M.T.; Vergani, P.; Albergoni, F.G. Relationship between fusaric acid uptake and its binding to cell structures by leaves of Egeria densa and its toxic effects on membrane permeability and respiration. Physiol. Mol. Plant Pathol. 1993, 42, 141–157. [Google Scholar] [CrossRef]
- Pavlovkin, J.; Mistrík, I.; Prokop, M. Some aspects of the phytotoxic action of fusaric acid on primary Ricinus roots. Plant Soil Environ. 2004, 50, 397–401. [Google Scholar] [CrossRef]
- Vázquez, M.D.; Poschenrieder, C.; Barceló, J.; Baker, A.J.M.; Hatton, P.; Cope, G.H. Compartmentation of zinc in roots and leaves of the zinc hyperaccumulator Thlaspi caerulescens J. C. Presl. Bot. Acta 1994, 107, 243–250. [Google Scholar] [CrossRef]
- Schneider, T.; Persson, D.P.; Husted, S.; Schellenberg, M.; Gehrig, P.; Lee, Y.; Martinoia, E.; Schjoerring, J.K.; Meyer, S. A proteomics approach to investigate the process of Zn hyperaccumulation in Noccaea caerulescens (J & C. Presl) F.K. Meyer. Plant J. 2013, 73, 131–142. [Google Scholar] [PubMed]
- Peng, J.; Gong, J. Vacuolar sequestration capacity and long-distance metal transport in plants. Front. Plant Sci. 2014, 5, 19. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Tian, S.; Zhang, J.; Yang, X.; Labavitch, J.M.; Webb, S.M.; Latimer, M.; Brown, P.H. Efficient xylem transport and phloem remobilization of Zn in the hyperaccumulator plant species S edum alfredii. New Phytol. 2013, 198, 721–731. [Google Scholar] [CrossRef]
- Álvarez-Fernández, A.; Díaz-Benito, P.; Abadía, A.; López-Millán, A.-F.; Abadía, J. Metal species involved in long distance metal transport in plants. Front. Plant Sci. 2014, 5, 105. [Google Scholar] [CrossRef] [Green Version]
- Pilon, M.; Abdelghany, S.E.; Cohu, C.M.; Gogolin, K.A.; Ye, H. Copper cofactor delivery in plant cells. Curr. Opin. Plant Biol. 2006, 9, 256–263. [Google Scholar] [CrossRef]
- Claudia, C.; Enrico, M.; Catherine, K. Hyperaccumulation of cadmium and zinc in Thlaspi caerulescens and Arabidopsis halleri at the leaf cellular level. Plant Physiol. 2004, 134, 716–725. [Google Scholar]
- Tan, N.; Pan, J.H.; Peng, G.T.; Mou, C.B.; Tao, Y.W.; She, Z.G.; Yang, Z.L.; Zhou, S.N.; Lin, Y.C. A copper coordination compound produced by a marine fungus Fusarium sp. ZZF51 with biosorption of Cu (II) ions. Chin. J. Chem. 2008, 26, 516–521. [Google Scholar] [CrossRef]
- Jian, Y.; Meredith, M.; Stack, B.C. Effects of fusaric acid treatment on HEp2 and docetaxel-resistant HEp2 laryngeal squamous cell carcinoma. Chemotherapy 2013, 59, 121–128. [Google Scholar]
- Devaraja, S.; Girish, K.S.; Santhosh, M.S.; Hemshekhar, M.; Nayaka, S.C.; Kemparaju, K. Fusaric acid, a mycotoxin, and its influence on blood coagulation and platelet function. Blood Coagul. Fibrinolysis 2013, 24, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Krämer, U. Metal hyperaccumulation in plants. Annu. Rev. Plant Biol. 2010, 61, 517–534. [Google Scholar] [CrossRef]
- Noman, A.; Aqeel, M.; Khalid, N.; Islam, W.; Sanaullah, T.; Anwar, M.; Khan, S.; Ye, W.; Lou, Y. Zinc finger protein transcription factors: Integrated line of action for plant antimicrobial activity. Microb. Pathog. 2019, 132, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Yruela, I. Copper in plants: Acquisition, transport and interactions. Funct. Plant Biol. 2009, 36, 409–430. [Google Scholar] [CrossRef] [Green Version]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B., Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef]
- Cakmak, I. Tansley review no. 111: Possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytol. 2010, 146, 185–205. [Google Scholar] [CrossRef]
- Singh, V.K.; Upadhyay, R. Induction of defence responses by fusaric acid (Fusarium toxin) in tomato plant. In Proceedings of the 6th International Conference on Agriculture, Environment and Biological Sciences, Kuala Lumpur, Malaysia, 21–22 December 2016; pp. 1–22. [Google Scholar]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F.J. Function of nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 2012; pp. 191–248. [Google Scholar]
- Li, J.; Essemine, J.; Shang, C.; Zhang, H.; Zhu, X.; Yu, J.; Chen, G.; Qu, M.; Sun, D. Combined proteomics and metabolism analysis unravels prominent roles of antioxidant system in the prevention of alfalfa (Medicago sativa L.) against salt stress. Int. J. Mol. Sci. 2020, 21, 909. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, R.M.; Cheng, C.; Sheng, J.; Wang, W.; Ren, H.; Aslam, M.; Yan, Y. Interruption of jasmonic acid biosynthesis causes differential responses in the roots and shoots of maize seedlings against salt stress. Int. J. Mol. Sci. 2019, 20, 6202. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.K.; Sousadias, M.G. Fusaric acid content of swine feedstuffs. J. Agric. Food Chem. 1994, 41, 2296–2298. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, M.; Sun, Y.; Gu, Z.; Wang, R.; Saydin, A.; Shen, Q.; Guo, S. Nitrate increased cucumber tolerance to Fusarium wilt by regulating fungal toxin production and distribution. Toxins 2017, 9, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Gu, Z.C.; Wang, R.R.; Guo, J.J.; Ling, N.; Firbank, L.G.; Guo, S.W. Plant primary metabolism regulated by nitrogen contributes to plant-pathogen interactions. Plant Cell Physiol. 2019, 60, 329–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Stewart, R.R.C.; Bewley, J.D. Lipid peroxidation associated with accelerated aging of soybean axes. Plant Physiol. 1980, 65, 245–248. [Google Scholar] [CrossRef] [Green Version]
- Wang, X. Plant Physiology and Biochemistry Experiment Principle and Technology, 2nd ed.; Wang, X., Ed.; Higher Education Press: Beijing, China, 2006; pp. 169–173. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | CK | CK + FA | Zn + FA | Cu + FA |
|---|---|---|---|---|
| Plant height (cm) | 18.3 ± 1.46 a | 15.2 ± 0.28 b | 18.7 ± 2.08 a | 17.5 ± 0.50 ab |
| Root length (cm) | 15.5 ± 1.15 a | 14.7 ± 0.60 a | 15.0 ± 1.70 a | 14.3 ± 0.57 a |
| SPAD | 48.9 ± 1.30 a | 43.3 ± 0.60 b | 49.6 ± 2.00 a | 49.9 ± 1.90 a |
| Plant dry weight (g) | 0.71 ± 0.02 b | 0.62 ± 0.03 c | 0.83 ± 0.02 a | 0.80 ± 0.04 a |
| Pn(μmolCO2m−2s−1) | 22.0 ± 0.67 a | 13.8 ± 1.51 c | 17.0 ± 0.16 b | 18.0 ± 0.28 b |
| gs(molH2Om−2s−1) | 0.74 ± 0.01 a | 0.12 ± 0.00 c | 0.48 ± 0.01ab | 0.62 ± 0.02 a |
| Ci(μmolCO2mol−1) | 357 ± 7.63 a | 258 ± 5.50 c | 328 ± 0.95 b | 365 ± 6.35 a |
| E(mmolH2Om−2s−1) | 9.06 ± 0.49 a | 3.08 ± 0.29 b | 9.43 ± 0.02 a | 9.16 ± 0.02 a |
| E/gs(10−3) | 7.42 ± 0.79 b | 18.7 ± 1.62 a | 8.42 ± 0.03 b | 7.39 ± 0.11 b |
| Wilting index (%) | - | 100 ± 5.77 a | 7.3 ± 0.76 b | 8.63 ± 0.30 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Huang, J.; Liang, A.; Wang, Y.; Mur, L.A.J.; Wang, M.; Guo, S. Zinc and Copper Enhance Cucumber Tolerance to Fusaric Acid by Mediating Its Distribution and Toxicity and Modifying the Antioxidant System. Int. J. Mol. Sci. 2020, 21, 3370. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093370
Wang R, Huang J, Liang A, Wang Y, Mur LAJ, Wang M, Guo S. Zinc and Copper Enhance Cucumber Tolerance to Fusaric Acid by Mediating Its Distribution and Toxicity and Modifying the Antioxidant System. International Journal of Molecular Sciences. 2020; 21(9):3370. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093370
Chicago/Turabian StyleWang, Ruirui, Jian Huang, Aichen Liang, Ying Wang, Luis Alejandro Jose Mur, Min Wang, and Shiwei Guo. 2020. "Zinc and Copper Enhance Cucumber Tolerance to Fusaric Acid by Mediating Its Distribution and Toxicity and Modifying the Antioxidant System" International Journal of Molecular Sciences 21, no. 9: 3370. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093370