Disease-Specific Derangement of Circulating Endocannabinoids and N-Acylethanolamines in Myeloproliferative Neoplasms

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Study Cohort
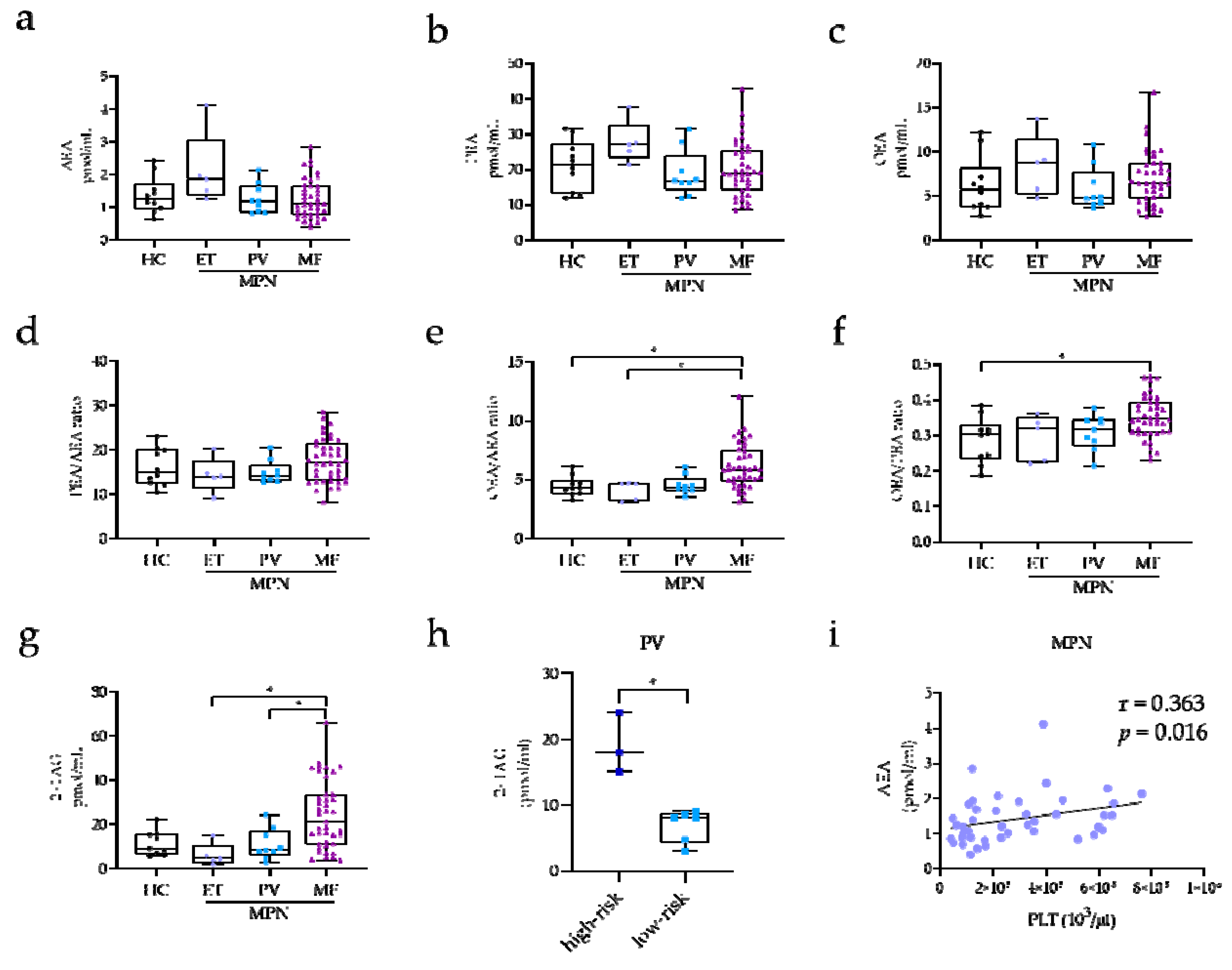
2.2. EC and NAE Plasma Profile of MPN Subclasses
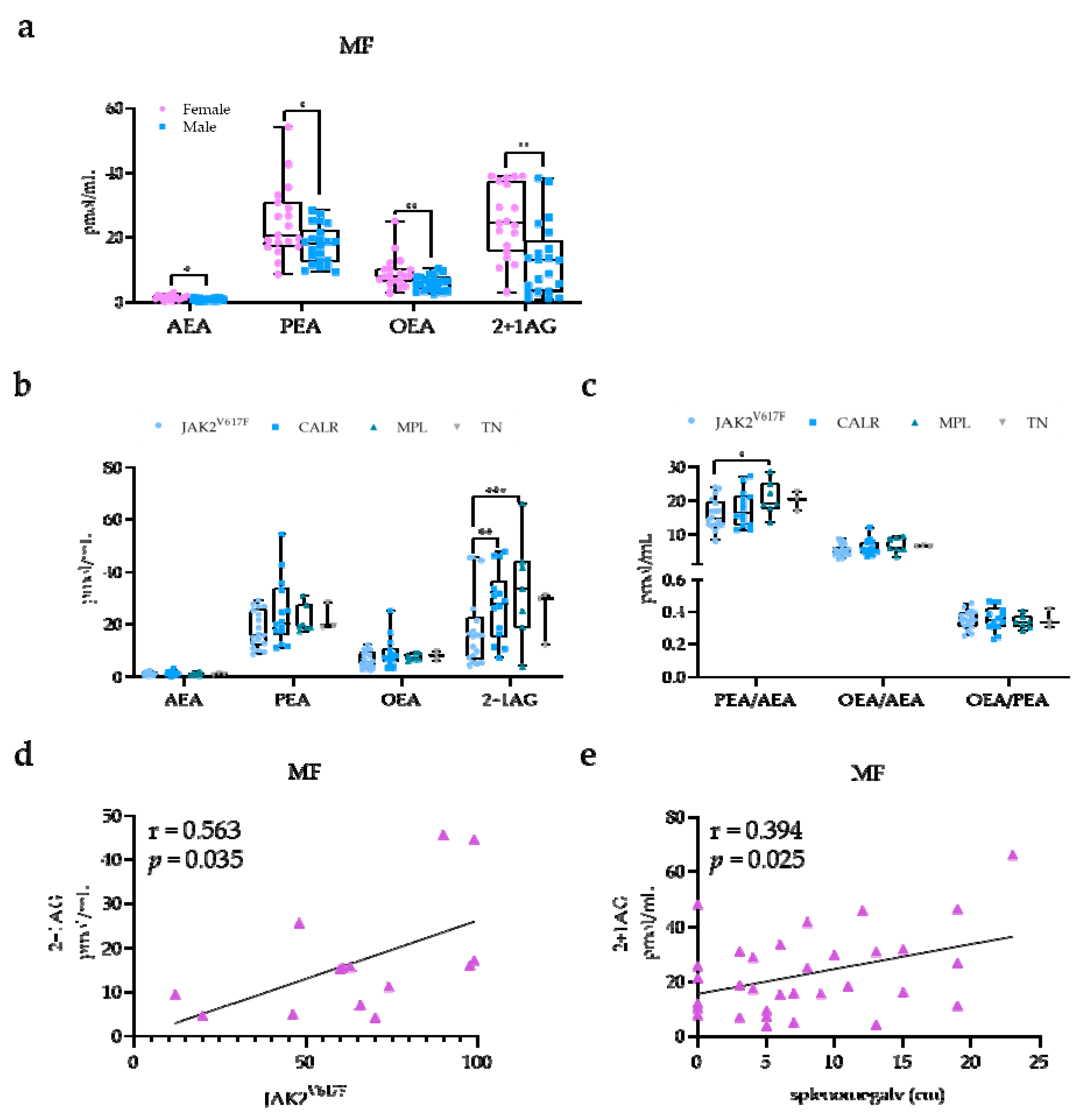
2.3. EC and NAE Plasma Profile According to Risk Classification, Mutational Status, and Clinical Manifestations in MF Patients
3. Discussion
4. Materials and Methods
4.1. Study Cohort
4.2. Blood Sampling
4.3. Mutation Analysis
4.4. EC and NAE Measurements
4.5. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MPN | myeloproliferative neoplasms |
| MF | myelofibrosis |
| ET | essential thrombocythemia |
| PV | polycythemia vera |
| HC | healthy control |
| EC | endocannabinoid |
| NAE | N-acylethanolamide |
| AEA | N-arachidonoyl-ethanolamine (anandamide) |
| 1AG | 1-arachidonoyl-glycerol |
| 2AG | 2-arachidonoyl-glycerol |
| 2+1AG | 1/2-arachidonoyl-glycerol |
| PEA | palmitoyl-ethanolamide |
| OEA | oleoyl-ethanolamide |
| LC–MS/MS | liquid chromatography–tandem mass spectrometry |
| CB1/2 | cannabinoid receptors ½ |
| WBC | white blood cell |
| PLT | platelet count |
| Hct | hematocrit |
| RBC | red blood cell |
| Hgb | hemoglobin |
| TSS | total symptoms score |
| DIPSS | dynamic international prognostic scoring system |
| JAK2 | janus kinase 2 |
| CALR | calreticulin |
| MPL | myeloproliferative leukemia protein |
| TN | triple-negative |
| VAF | variant allele frequency |
| BMI | body mass index |
References
- Hillard, C.J. The Endocannabinoid Signaling System in the CNS: A Primer. Int. Rev. Neurobiol. 2015, 125, 1–47. [Google Scholar] [PubMed]
- Knight, J.M.; Szabo, A.; Zhao, S. Circulating endocannabinoids during hematopoietic stem cell transplantation: A pilot study. Neurobiol. Stress. 2015, 2, 44–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuboi, K.; Uyama, T.; Okamoto, Y.; Ueda, N. Endocannabinoids and related N-acylethanolamines: Biological activities and metabolism. Inflamm. Regen. 2018, 38, 28. [Google Scholar] [CrossRef] [PubMed]
- Howlett, A.C.; Blume, L.C.; Dalton, G.D. CB(1) cannabinoid receptors and their associated proteins. Curr. Med. Chem. 2010, 17, 1382–1393. [Google Scholar] [CrossRef] [Green Version]
- Lakiotaki, E.; Giaginis, C.; Tolia, M. Clinical Significance of Cannabinoid Receptors CB1 and CB2 Expression in Human Malignant and Benign Thyroid Lesions. Biomed. Res. Int. 2015, 2015, 839403. [Google Scholar] [CrossRef] [Green Version]
- Kose, S.; Aerts-Kaya, F.; Kopru, C.Z. Human bone marrow mesenchymal stem cells secrete endocannabinoids that stimulate in vitro hematopoietic stem cell migration effectively comparable to beta-adrenergic stimulation. Exp. Hematol. 2018, 57, 30–41. [Google Scholar] [CrossRef]
- Fanelli, F.; Di Lallo, V.D.; Belluomo, I. Estimation of reference intervals of five endocannabinoids and endocannabinoid related compounds in human plasma by two dimensional- LC/MS/MS. J. Lipid Res. 2012, 53, 481–493. [Google Scholar] [CrossRef] [Green Version]
- Fanelli, F.; Mezzullo, M.; Belluomo, I. Plasma 2-arachidonoylglycerol is a biomarker of age and menopause related insulin resistance and dyslipidemia in lean but not in obese men and women. Mol. Metab. 2017, 6, 406–415. [Google Scholar] [CrossRef]
- Docs, K.; Meszar, Z.; Gonda, S. The Ratio of 2-AG to Its Isomer 1-AG as an Intrinsic Fine Tuning Mechanism of CB1 Receptor Activation. Front Cell Neurosci. 2017, 11, 39. [Google Scholar] [CrossRef] [Green Version]
- Balvers, M.G.; Wortelboer, H.M.; Witkamp, R.F.; Verhoeckx, K.C. Liquid chromatography-tandem mass spectrometry analysis of free and esterified fatty acid N-acyl ethanolamines in plasma and blood cells. Anal. Biochem. 2013, 434, 275–283. [Google Scholar] [CrossRef]
- Borrelli, F.; Romano, B.; Petrosino, S. Palmitoylethanolamide, a naturally occurring lipid, is an orally effective intestinal anti-inflammatory agent. Br. J. Pharmacol. 2015, 172, 142–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jumpertz, R.; Guijarro, A.; Pratley, R.E.; Piomelli, D.; Krakoff, J. Central and peripheral endocannabinoids and cognate acylethanolamides in humans: Association with race, adiposity, and energy expenditure. J. Clin. Endocrinol. Metab. 2011, 96, 787–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanelli, F.; Mezzullo, M.; Repaci, A. Profiling plasma N-Acylethanolamine levels and their ratios as a biomarker of obesity and dysmetabolism. Mol. Metab. 2018, 14, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Capoccia, E.; Turco, F. Palmitoylethanolamide improves colon inflammation through an enteric glia/toll like receptor 4-dependent PPAR-alpha activation. Gut 2014, 63, 1300–1312. [Google Scholar] [CrossRef]
- Gasperi, V.; Avigliano, L.; Evangelista, D. 2-Arachidonoylglycerol enhances platelet formation from human megakaryoblasts. Cell Cycle. 2014, 13, 3938–3947. [Google Scholar] [CrossRef] [Green Version]
- Cabral, G.A.; Griffin-Thomas, L. Emerging role of the cannabinoid receptor CB2 in immune regulation: Therapeutic prospects for neuroinflammation. Expert Rev. Mol. Med. 2009, 11, e3. [Google Scholar] [CrossRef] [Green Version]
- Chiurchiu, V.; Battistini, L.; Maccarrone, M. Endocannabinoid signalling in innate and adaptive immunity. Immunology 2015, 144, 352–364. [Google Scholar] [CrossRef]
- Karwad, M.A.; Macpherson, T.; Wang, B. Oleoylethanolamine and palmitoylethanolamine modulate intestinal permeability in vitro via TRPV1 and PPARalpha. Faseb J. 2017, 31, 469–481. [Google Scholar] [CrossRef] [Green Version]
- Chiurchiu, V.; Leuti, A.; Smoum, R.; Mechoulam, R.; Maccarrone, M. Bioactive lipids ALIAmides differentially modulate inflammatory responses of distinct subsets of primary human T lymphocytes. Faseb J. 2018, 32, 5716–5723. [Google Scholar] [CrossRef]
- Jiang, S.; Fu, Y.; Avraham, H.K. Regulation of hematopoietic stem cell trafficking and mobilization by the endocannabinoid system. Transfusion 2011, 51, 65s–71s. [Google Scholar] [CrossRef]
- Patinkin, D.; Milman, G.; Breuer, A.; Fride, E.; Mechoulam, R. Endocannabinoids as positive or negative factors in hematopoietic cell migration and differentiation. Eur. J. Pharmacol. 2008, 595, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Galve-Roperh, I.; Chiurchiu, V.; Diaz-Alonso, J.; Bari, M.; Guzman, M.; Maccarrone, M. Cannabinoid receptor signaling in progenitor/stem cell proliferation and differentiation. Prog. Lipid Res. 2013, 52, 633–650. [Google Scholar] [CrossRef]
- Barbado, M.V.; Medrano, M.; Caballero-Velazquez, T. Cannabinoid derivatives exert a potent anti-myeloma activity both in vitro and in vivo. Int. J. Cancer 2017, 140, 674–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randall, M.D. Endocannabinoids and the haematological system. Br. J. Pharmacol. 2007, 152, 671–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giaginis, C.; Lakiotaki, E.; Korkolopoulou, P.; Konstantopoulos, K.; Patsouris, E.; Theocharis, S. Endocannabinoid System: A Promising Therapeutic Target for the Treatment of Haematological Malignancies? Curr. Med. Chem. 2016, 23, 2350–2362. [Google Scholar] [CrossRef]
- Joosten, M.; Valk, P.J.; Jorda, M.A. Leukemic predisposition of pSca-1/Cb2 transgenic mice. Exp. Hematol. 2002, 30, 142–149. [Google Scholar] [CrossRef]
- Jorda, M.A.; Rayman, N.; Tas, M. The peripheral cannabinoid receptor Cb2, frequently expressed on AML blasts, either induces a neutrophilic differentiation block or confers abnormal migration properties in a ligand-dependent manner. Blood 2004, 104, 526–534. [Google Scholar] [CrossRef] [Green Version]
- Jorda, M.A.; Rayman, N.; Valk, P.; De Wee, E.; Delwel, R. Identification, characterization, and function of a novel oncogene: The peripheral cannabinoid receptor Cb2. Ann. N. Y. Acad. Sci. 2003, 996, 10–16. [Google Scholar] [CrossRef]
- Kampa-Schittenhelm, K.M.; Salitzky, O.; Akmut, F.; Illing, B.; Kanz, L.; Salih, H.R.; Schittenhelm, M.M. Dronabinol has preferential antileukemic activity in acute lymphoblastic and myeloid leukemia with lymphoid differentiation patterns. BMC Cancer 2016, 16, 25. [Google Scholar] [CrossRef] [Green Version]
- Tefferi, A.; Vainchenker, W. Myeloproliferative Neoplasms: Molecular Pathophysiology, Essential Clinical Understanding, and Treatment Strategies. J. Clin. Oncol. 2011, 29, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Palandri, F.; Breccia, M.; Bonifacio, M. Life after ruxolitinib: Reasons for discontinuation, impact of disease phase, and outcomes in 218 patients with myelofibrosis. Cancer 2020, 126, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Passamonti, F.; Cervantes, F.; Vannucchi, A.M. A dynamic prognostic model to predict survival in primary myelofibrosis: A study by the IWG-MRT (International Working Group for Myeloproliferative Neoplasms Research and Treatment). Blood 2009, 115, 1703–1708. [Google Scholar] [CrossRef] [PubMed]
- Moreno, E.; Cavic, M.; Krivokuca, A.; Casado, V.; Canela, E. The Endocannabinoid System as a Target in Cancer Diseases: Are We There Yet? Front Pharmacol. 2019, 10, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sailler, S.; Schmitz, K.; Jäger, E. Regulation of circulating endocannabinoids associated with cancer and metastases in mice and humans. Oncoscience 2014, 1, 272–282. [Google Scholar] [CrossRef] [Green Version]
- Sollazzo, D.; Forte, D.; Polverelli, N. Crucial factors of the inflammatory microenvironment (IL-1beta/TNF-alpha/TIMP-1) promote the maintenance of the malignant hemopoietic clone of myelofibrosis: An in vitro study. Oncotarget 2016, 7, 43974–43988. [Google Scholar] [CrossRef] [Green Version]
- Catani, M.V.; Gasperi, V.; Evangelista, D.; Finazzi Agro, A.; Avigliano, L.; Maccarrone, M. Anandamide extends platelets survival through CB(1)-dependent Akt signaling. Cell Mol. Life Sci. 2010, 67, 601–610. [Google Scholar] [CrossRef]
- Bluher, M.; Engeli, S.; Kloting, N. Dysregulation of the peripheral and adipose tissue endocannabinoid system in human abdominal obesity. Diabetes 2006, 55, 3053–3060. [Google Scholar] [CrossRef] [Green Version]
- Turcotte, C.; Chouinard, F.; Lefebvre, J.S.; Flamand, N. Regulation of inflammation by cannabinoids, the endocannabinoids 2-arachidonoyl-glycerol and arachidonoyl-ethanolamide, and their metabolites. J. Leukoc. Biol. 2015, 97, 1049–1070. [Google Scholar] [CrossRef]
- Turcotte, C.; Blanchet, M.R.; Laviolette, M.; Flamand, N. The CB2 receptor and its role as a regulator of inflammation. Cell Mol. Life Sci. 2016, 73, 4449–4470. [Google Scholar] [CrossRef] [Green Version]
- Hillard, C.J. Circulating Endocannabinoids: From Whence Do They Come and Where are They Going? Neuropsychopharmacology 2018, 43, 155–172. [Google Scholar] [CrossRef]
- Sollazzo, D.; Forte, D.; Polverelli, N. Circulating Calreticulin Is Increased in Myelofibrosis: Correlation with Interleukin-6 Plasma Levels, Bone Marrow Fibrosis, and Splenomegaly. Mediat. Inflamm. 2016, 2016, 5860657. [Google Scholar] [CrossRef] [PubMed]
- Ciurea, S.O.; Merchant, D.; Mahmud, N. Pivotal contributions of megakaryocytes to the biology of idiopathic myelofibrosis. Blood 2007, 110, 986–993. [Google Scholar] [CrossRef]
- Malara, A.; Abbonante, V.; Zingariello, M.; Migliaccio, A.; Balduini, A. Megakaryocyte Contribution to Bone Marrow Fibrosis: Many Arrows in the Quiver. Mediterr. J. Hematol. Infect Dis. 2018, 10, e2018068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, H.; Adane, B.; Khan, N. Subversion of Systemic Glucose Metabolism as a Mechanism to Support the Growth of Leukemia Cells. Cancer Cell 2018, 34, 659–673.e6. [Google Scholar] [CrossRef] [Green Version]
- Berdyshev, E.V.; Schmid, P.C.; Krebsbach, R.J.; Schmid, H.H. Activation of PAF receptors results in enhanced synthesis of 2-arachidonoylglycerol (2-AG) in immune cells. Faseb J. 2001, 15, 2171–2178. [Google Scholar] [CrossRef] [PubMed]
- Romano, M.; Sollazzo, D.; Trabanelli, S. Mutations in JAK2 and Calreticulin genes are associated with specific alterations of the immune system in myelofibrosis. Oncoimmunology 2017, 6, e1345402. [Google Scholar] [CrossRef] [Green Version]
- Geyer, H.L.; Dueck, A.C.; Scherber, R.M.; Mesa, R.A. Impact of Inflammation on Myeloproliferative Neoplasm Symptom Development. Mediat. Inflamm. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Geyer, H.L.; Kosiorek, H.; Dueck, A.C. Associations between gender, disease features and symptom burden in patients with myeloproliferative neoplasms: An analysis by the MPN QOL International Working Group. Haematologica 2017, 102, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Barraco, D.; Mora, B.; Guglielmelli, P. Gender effect on phenotype and genotype in patients with post- polycythemia vera and post-essential thrombocythemia myelofibrosis: Results from the MYSEC project. Blood Cancer J. 2018, 8, 89. [Google Scholar] [CrossRef]
- Gasperi, V.; Evangelista, D.; Savini, I.; Del Principe, D.; Avigliano, L.; Maccarrone, M.; Catani, M.V. Downstream effects of endocannabinoid on blood cells: Implications for health and disease. Cell Mol. Life Sci. 2015, 72, 3235–3252. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Descriptive Parameters | HC (n = 10) | ET (n = 5) | PV (n = 9) | MF (n = 41) |
|---|---|---|---|---|
| Sex (F/M) | 5/5 | 2/3 | 3/6 | 19/22 |
| Age (years) | 59 (31–73) | 52 (42–57) ** | 57 (26–71) ## | 72 (46–89) + |
| WBC (103/μL) | 6.1 (4.3–9.0) | 7.5 (7.4–10.3) | 8.1 (7.4–14.9) | 9.9 (1.6–38.6) |
| PLT (103/µL) | 261 (159–306) | 463 (330–656) * | 438 (229–762) | 121 (38–632) |
| Hgb (g/dL) | 14.1 (12.9–15.5) | 14.2 (14–15.4) *** | 13.9 (11–16.20) ## | 9.9 (7.2–15.28) +++ |
| RBC (106/µL) | 4.6 (4.1–5.3) | 5.59 (4.6–5.65) | 5.5 (3.38–7.4) # | 3.7(2.4–6.2) |
| Hct (%) | 41.5 (37.6–46.7) | 44.7(43–47.98) *** | 46.5 (41.8–49.5) ### | 30.87 (24.16–50.69) ++ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forte, D.; Fanelli, F.; Mezzullo, M.; Barone, M.; Corradi, G.; Auteri, G.; Bartoletti, D.; Martello, M.; Ottaviani, E.; Terragna, C.; et al. Disease-Specific Derangement of Circulating Endocannabinoids and N-Acylethanolamines in Myeloproliferative Neoplasms. Int. J. Mol. Sci. 2020, 21, 3399. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093399
Forte D, Fanelli F, Mezzullo M, Barone M, Corradi G, Auteri G, Bartoletti D, Martello M, Ottaviani E, Terragna C, et al. Disease-Specific Derangement of Circulating Endocannabinoids and N-Acylethanolamines in Myeloproliferative Neoplasms. International Journal of Molecular Sciences. 2020; 21(9):3399. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093399
Chicago/Turabian StyleForte, Dorian, Flaminia Fanelli, Marco Mezzullo, Martina Barone, Giulia Corradi, Giuseppe Auteri, Daniela Bartoletti, Marina Martello, Emanuela Ottaviani, Carolina Terragna, and et al. 2020. "Disease-Specific Derangement of Circulating Endocannabinoids and N-Acylethanolamines in Myeloproliferative Neoplasms" International Journal of Molecular Sciences 21, no. 9: 3399. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21093399