Compensation Mechanism of the Photosynthetic Apparatus in Arabidopsis thaliana ch1 Mutants



Abstract
:1. Introduction
2. Results
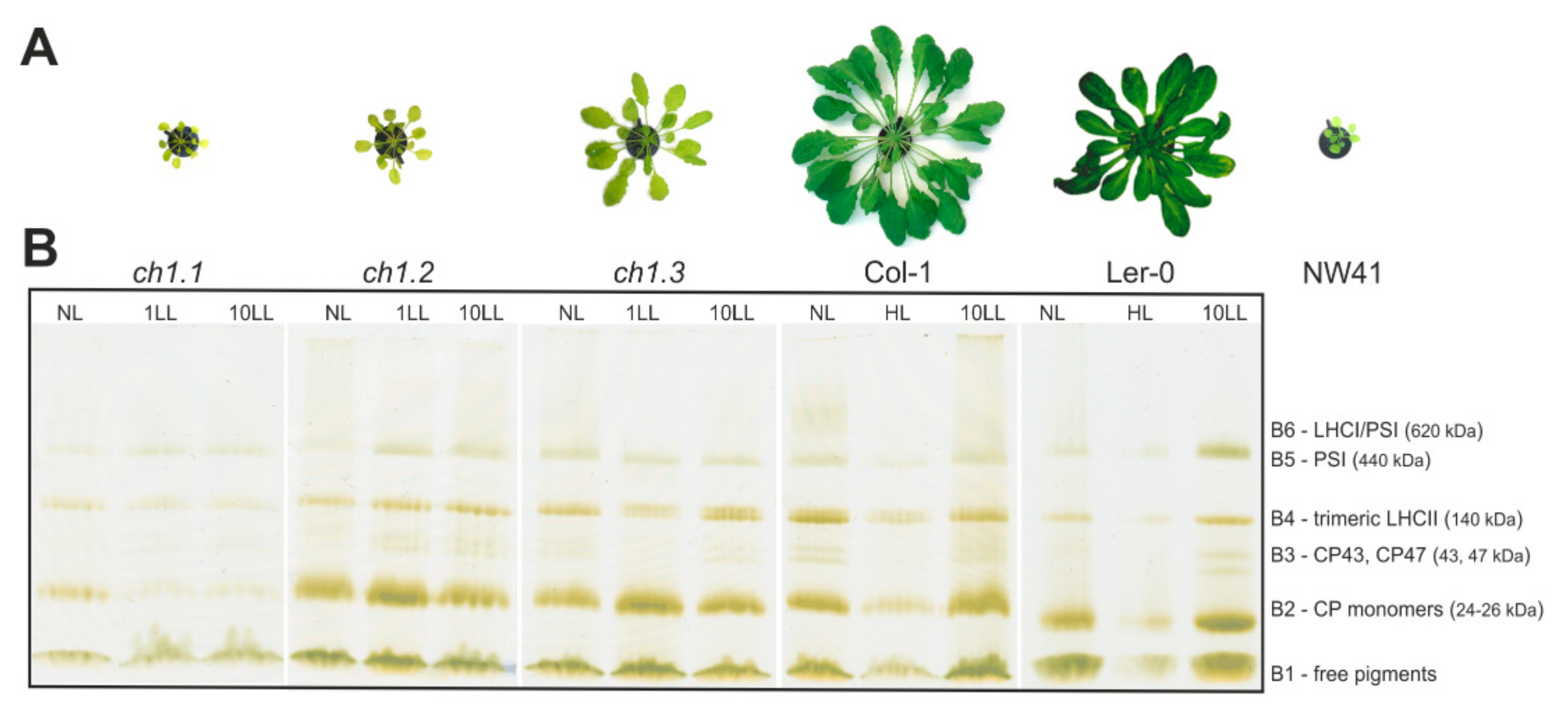
2.1. Pale Green Phenotypes
2.2. Organization and Composition of the Photosynthetic Apparatus and Analysis of PSI and PSII Protein Levels in Thylakoid Membranes
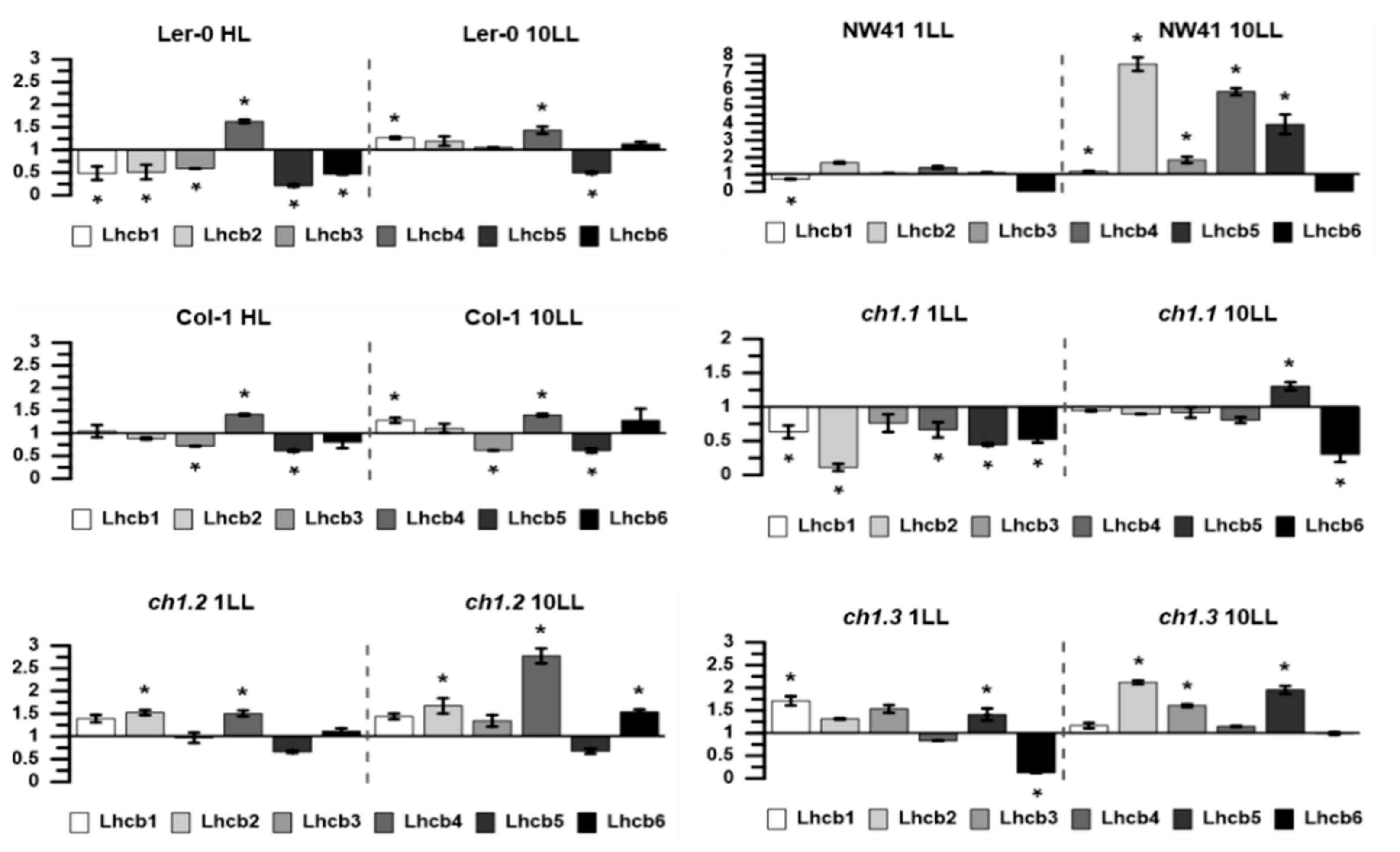
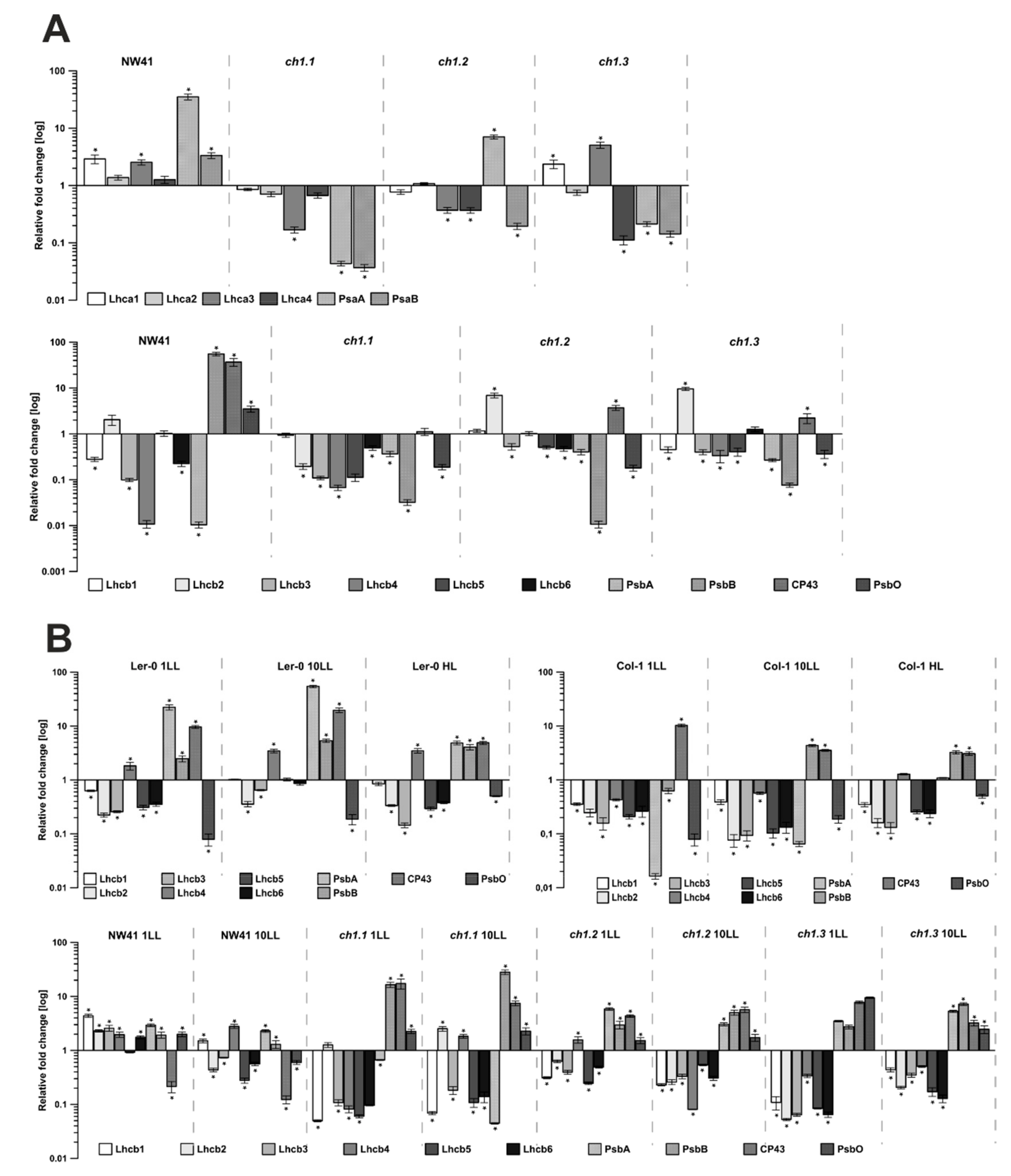
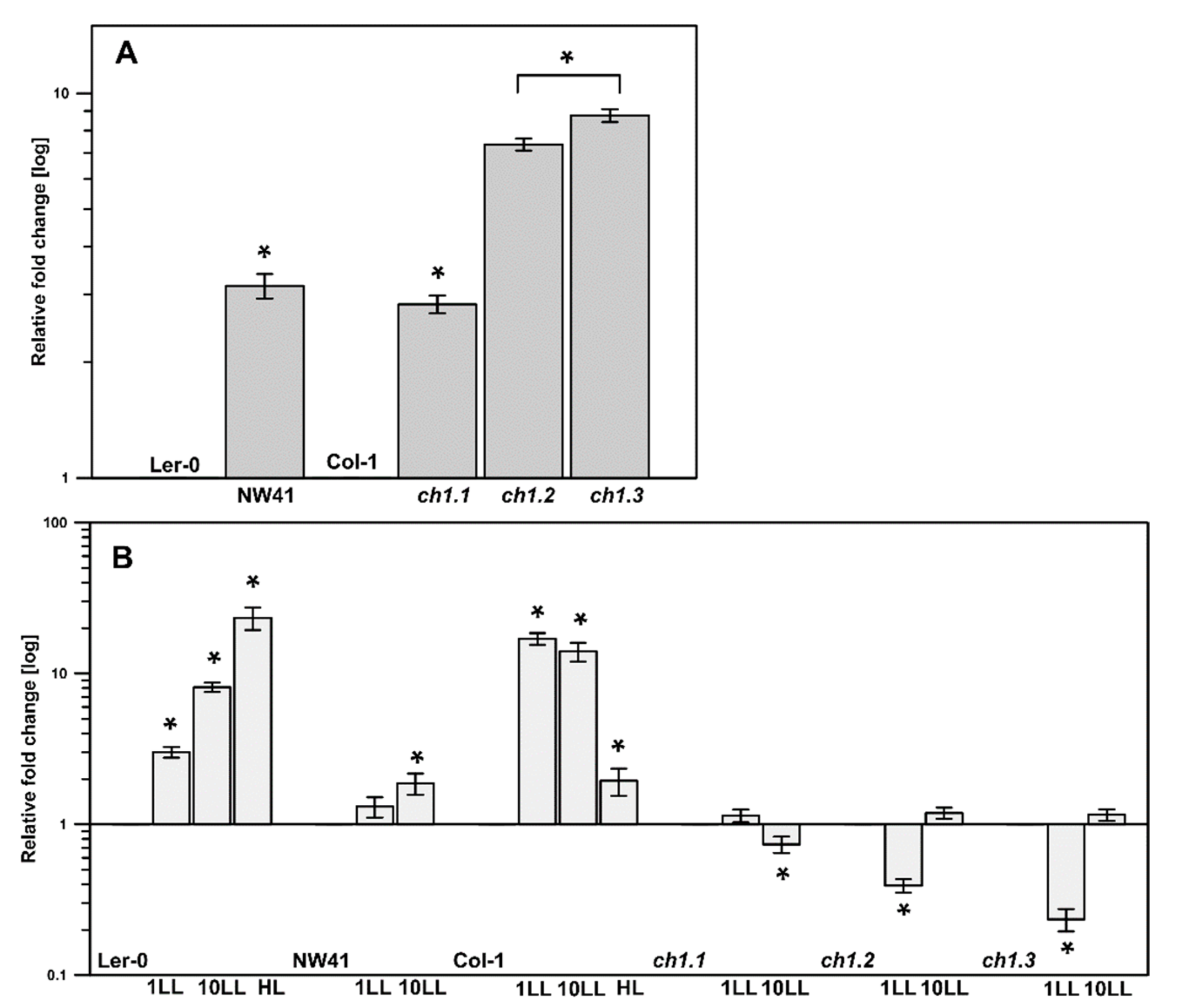
2.3. Gene Expression
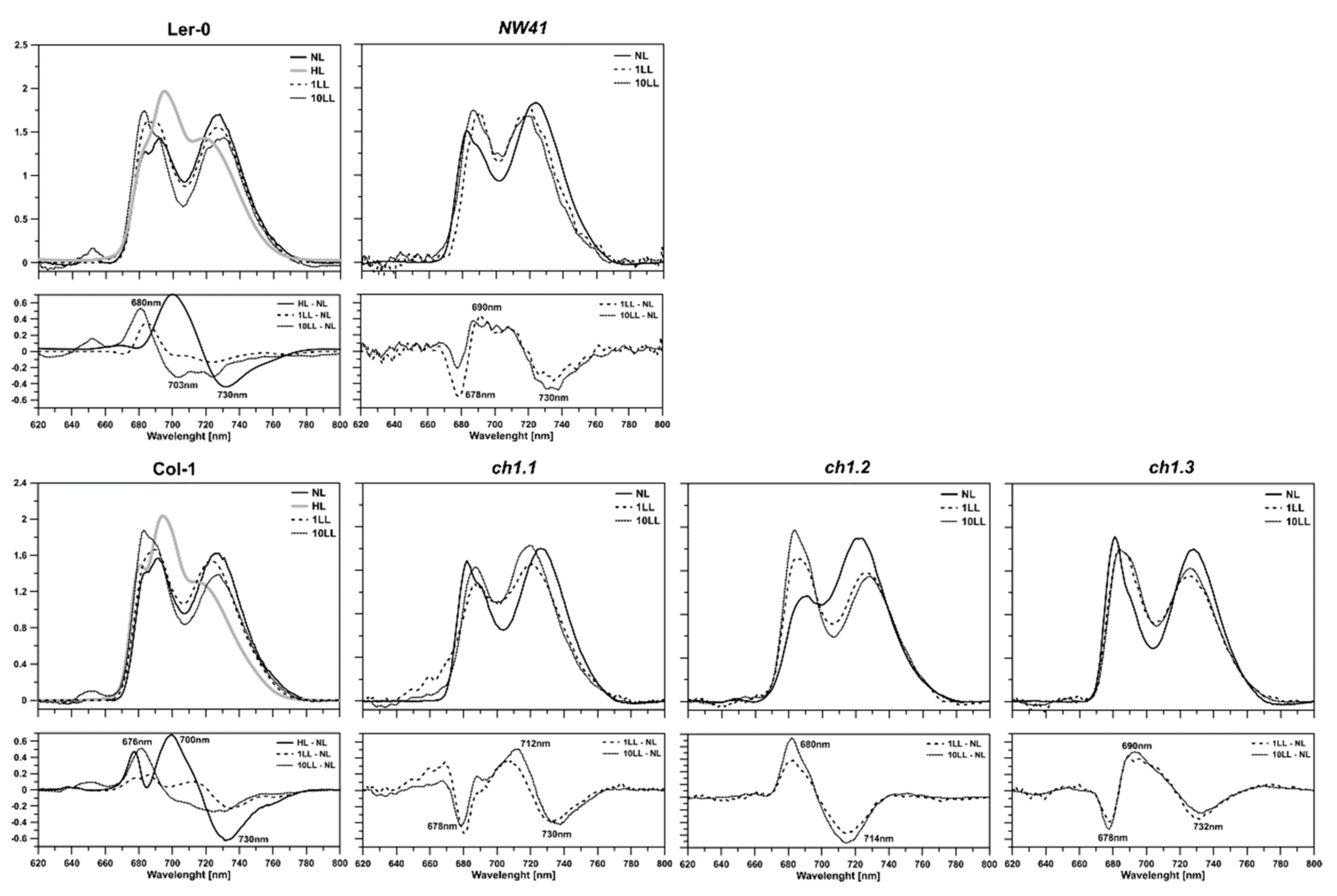
2.4. Efficiency of the Photosynthetic Apparatus
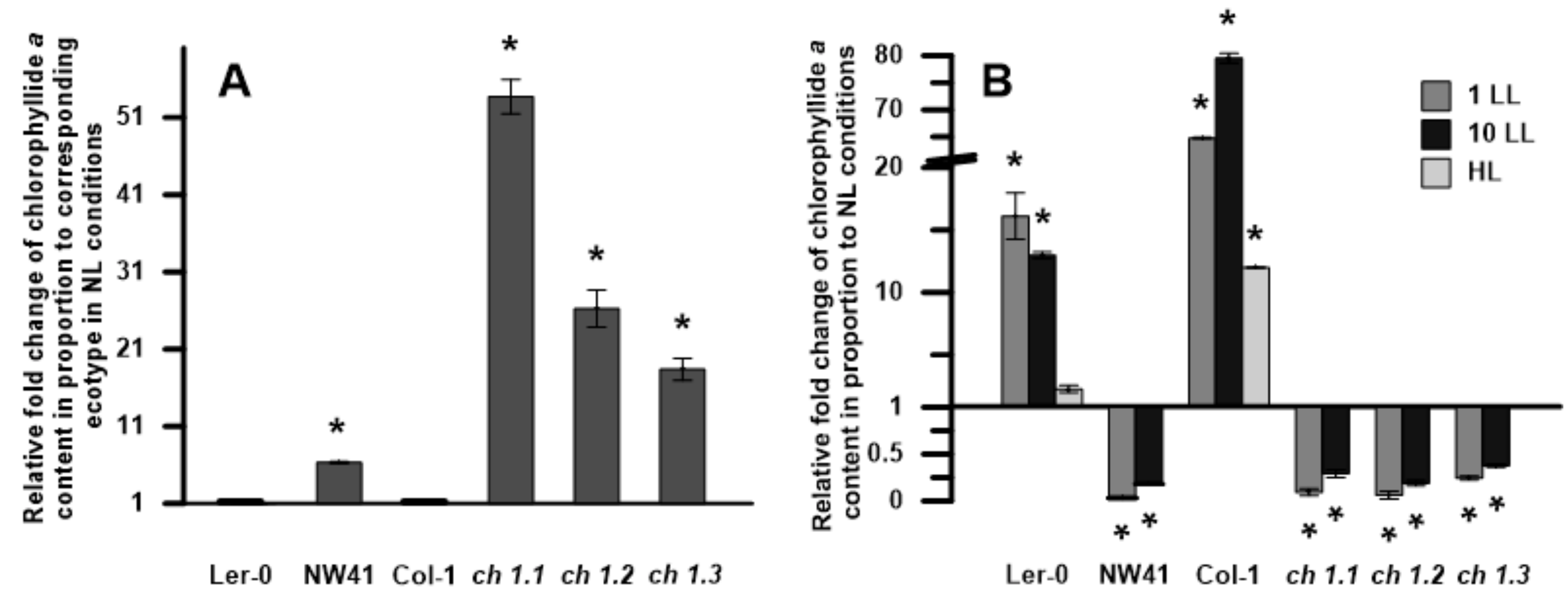
2.5. Pigment Composition
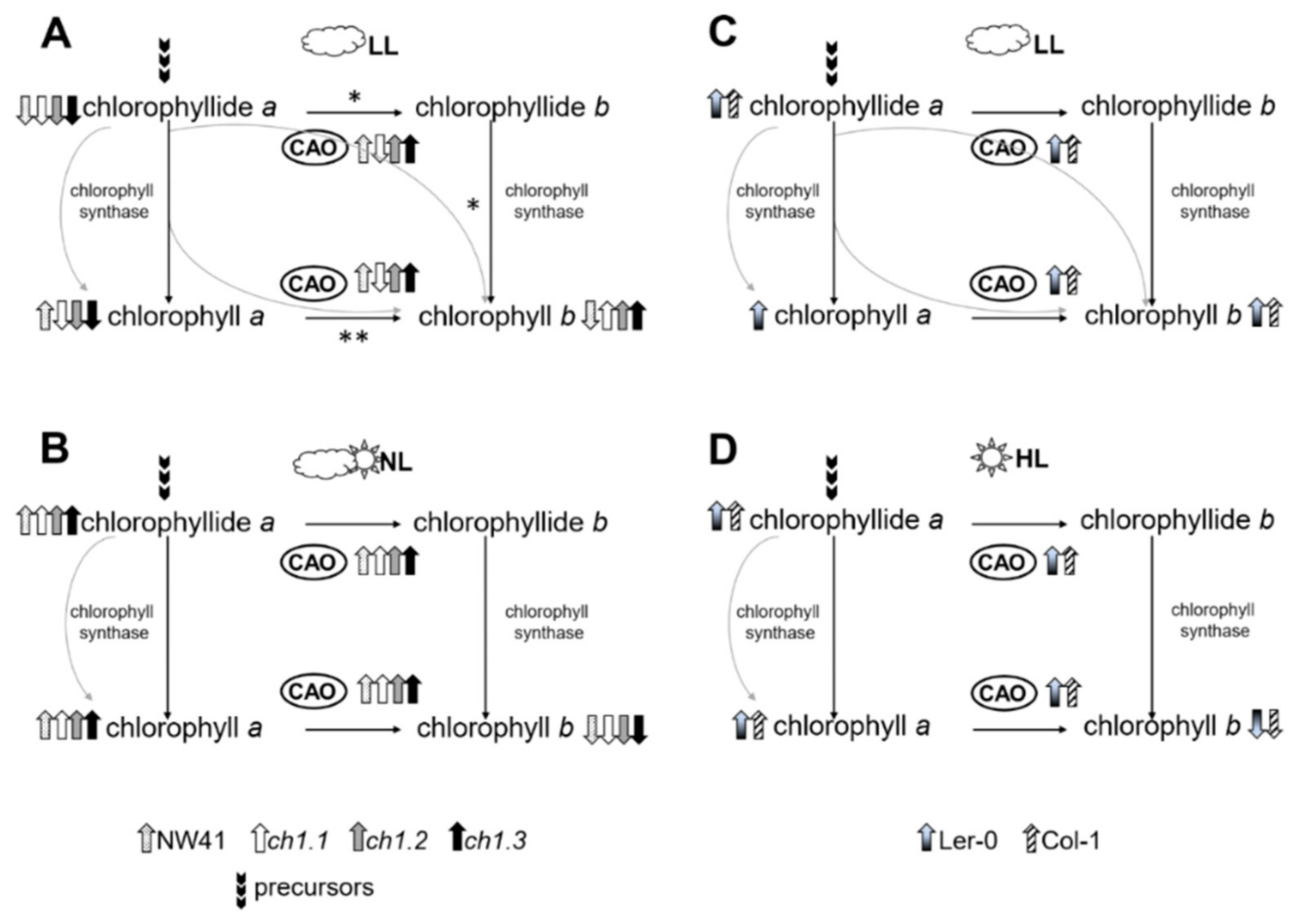
3. Discussion
3.1. Fifty Shades of Pale Green Phenotypes
3.2. Same Shortage, Different Statement (Coping with It)
3.3. Chlorina on the Crossroads
3.4. Natural Abilities to Rearrangements of Photosynthetic Apparatus
4. Materials and Methods
4.1. Plant Material, Growth Conditions, Stress Treatment and Photosynthetic Measurements
4.2. Preparation of Thylakoid Membranes
4.3. Protein Concentration
4.4. Low-Temperature (77 K) Fluorescence Measurements
4.5. Mild-Denaturing Electrophoresis, SDS-PAGE and Immunoblot Analysis
4.6. Extraction of Pigments
4.7. PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CAO | Chlorophyll a oxigenase |
| Chl | Chlorophyll |
| Chlide | Chlorophyllide |
| LHC | Light-harvesting complex |
References
- Kouril, R.; Dekker, J.P.; Boekema, E.J. Supramolecular organization of photosystem II in green plants. Biochim. Biophys. Acta 2012, 1817, 2–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouril, R.; Zygadlo, A.; Arteni, A.A.; de Wit, C.D.; Dekker, J.P.; Jensen, P.E.; Scheller, H.V.; Boekema, E.J. Structural characterization of a complex of photosystem I and light-harvesting complex II of Arabidopsis thaliana. Biochemistry 2005, 44, 10935–10940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caspy, I.; Nelson, N. Structure of the plant photosystem I. Biochem. Soc. Trans. 2018, 46, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Tanaka, R. The biochemistry, physiology, and evolution of the chlorophyll cycle. Adv. Bot. Res. 2019, 90, 183–212. [Google Scholar] [CrossRef]
- Oster, U.; Tanaka, R.; Tanaka, A.; Rudiger, W. Cloning and functional expression of the gene encoding the key enzyme for chlorophyll b biosynthesis (CAO) from Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2000, 21, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Chen, M. Chlorophyll Modifications and Their Spectral Extension in Oxygenic Photosynthesis. Annu. Rev. Biochem. 2014, 83, 317–340. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Yokono, M.; Akimoto, S.; Tanaka, R.; Tanaka, A. Deregulated chlorophyll b synthesis reduces the energy transfer rate between photosynthetic pigments and induces photodamage in Arabidopsis thaliana. Plant Cell Physiol. 2010, 51, 1055–1065. [Google Scholar] [CrossRef] [Green Version]
- Kunugi, M.; Takabayashi, A.; Tanaka, A. Evolutionary changes in chlorophyllide a oxygenase (CAO) structure contribute to the acquisition of a new light-harvesting complex in micromonas. J. Biol. Chem. 2013, 288, 19330–19341. [Google Scholar] [CrossRef] [Green Version]
- Yamasato, A.; Nagata, N.; Tanaka, R.; Tanaka, A. The N-terminal domain of chlorophyllide a oxygenase confers protein instability in response to chlorophyll B accumulation in Arabidopsis. Plant Cell 2005, 17, 1585–1597. [Google Scholar] [CrossRef] [Green Version]
- Reinbothe, C.; Bartsch, S.; Eggink, L.L.; Hoober, J.K.; Brusslan, J.; Andrade-Paz, R.; Monnet, J.; Reinbothe, S. A role for chlorophyllide a oxygenase in the regulated import and stabilization of light-harvesting chlorophyll a/b proteins. Proc. Natl. Acad. Sci. USA 2006, 103, 4777–4782. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, R.; Tanaka, A. Tetrapyrrole biosynthesis in higher plants. Annu. Rev. Plant Biol. 2007, 58, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, R.; Tanaka, A. Chlorophyll cycle regulates the construction and destruction of the light-harvesting complexes. Biochim. Biophys. Acta 2011, 1807, 968–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawara, E.; Sakuraba, Y.; Yamasato, A.; Tanaka, R.; Tanaka, A. Clp protease controls chlorophyll b synthesis by regulating the level of chlorophyllide a oxygenase. Plant J. Cell Mol. Biol. 2007, 49, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Pribil, M.; Labs, M.; Leister, D. Structure and dynamics of thylakoids in land plants. J. Exp. Bot. 2014, 65, 1955–1972. [Google Scholar] [CrossRef] [Green Version]
- Horie, Y.; Ito, H.; Kusaba, M.; Tanaka, R.; Tanaka, A. Participation of chlorophyll b reductase in the initial step of the degradation of light-harvesting chlorophyll a/b-protein complexes in Arabidopsis. J. Biol. Chem. 2009, 284, 17449–17456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbstova, M.; Tietz, S.; Kinzel, C.; Turkina, M.V.; Kirchhoff, H. Architectural switch in plant photosynthetic membranes induced by light stress. Proc. Natl. Acad. Sci. USA 2012, 109, 20130–20135. [Google Scholar] [CrossRef] [Green Version]
- Leong, T.Y.; Anderson, J.M. Adaptation of the Thylakoid Membranes of Pea-Chloroplasts to Light Intensities. II. Regulation of Electron-Transport Capacities, Electron Carriers, Coupling Factor (CF1) Activity and Rates of Photosynthesis. Photosynth. Res. 1984, 5, 117–128. [Google Scholar] [CrossRef]
- Jia, T.; Ito, H.; Tanaka, A. Simultaneous regulation of antenna size and photosystem I/II stoichiometry in Arabidopsis thaliana. Planta 2016, 244, 1041–1053. [Google Scholar] [CrossRef] [Green Version]
- Bielczynski, L.W.; Schansker, G.; Croce, R. Effect of Light Acclimation on the Organization of Photosystem II Super- and Sub-Complexes in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 105. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, R.; Koshino, Y.; Sawa, S.; Ishiguro, S.; Okada, K.; Tanaka, A. Overexpression of chlorophyllide a oxygenase (CAO) enlarges the antenna size of photosystem II in Arabidopsis thaliana. Plant J. 2001, 26, 365–373. [Google Scholar] [CrossRef]
- Takabayashi, A.; Kurihara, K.; Kuwano, M.; Kasahara, Y.; Tanaka, R.; Tanaka, A. The oligomeric states of the photosystems and the light-harvesting complexes in the Chl b-less mutant. Plant Cell Physiol. 2011, 52, 2103–2114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Highkin, H.R.; Frenkel, A.W. Studies of growth & metabolism of a barley mutant lacking chlorophyll b. Plant Physiol. 1962, 37, 814–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.; Peng, C.; Xu, X.; Lin, G.; Zhang, J. Thermostability of photosynthesis in two new chlorophyll b-less rice mutants. Sci. China. Ser. C Life Sci. 2005, 48, 139–147. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J.H.; Yoo, E.S.; Lee, C.H.; Hirochika, H.; An, G. Differential regulation of chlorophyll a oxygenase genes in rice. Plant Mol. Biol. 2005, 57, 805–818. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Li, X.P.; Razeghifard, R.; Anderson, J.M.; Niyogi, K.K.; Pogson, B.J.; Chow, W.S. The multiple roles of light-harvesting chlorophyll a/b-protein complexes define structure and optimize function of Arabidopsis chloroplasts: A study using two chlorophyll b-less mutants. Biochim. Biophys. Acta 2009, 1787, 973–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouril, R.; Wientjes, E.; Bultema, J.B.; Croce, R.; Boekema, E.J. High-light vs. low-light: Effect of light acclimation on photosystem II composition and organization in Arabidopsis thaliana. Biochim. Biophys. Acta 2013, 1827, 411–419. [Google Scholar] [CrossRef] [Green Version]
- Espineda, C.E.; Linford, A.S.; Devine, D.; Brusslan, J.A. The AtCAO gene, encoding chlorophyll a oxygenase, is required for chlorophyll b synthesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1999, 96, 10507–10511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skupien, J.; Wojtowicz, J.; Kowalewska, L.; Mazur, R.; Garstka, M.; Gieczewska, K.; Mostowska, A. Dark-chilling induces substantial structural changes and modifies galactolipid and carotenoid composition during chloroplast biogenesis in cucumber (Cucumis sativus L.) cotyledons. Plant Physiol. Biochem. 2017, 111, 107–118. [Google Scholar] [CrossRef]
- Pfundel, E.E.; Klughammer, C.; Meister, A.; Cerovic, Z.G. Deriving fluorometer-specific values of relative PSI fluorescence intensity from quenching of F(0) fluorescence in leaves of Arabidopsis thaliana and Zea mays. Photosynth. Res. 2013, 114, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Rudowska, L.; Gieczewska, K.; Mazur, R.; Garstka, M.; Mostowska, A. Chloroplast biogenesis—Correlation between structure and function. Biochim. Biophys. Acta 2012, 1817, 1380–1387. [Google Scholar] [CrossRef] [Green Version]
- Mueller, A.H.; Dockter, C.; Gough, S.P.; Lundqvist, U.; von Wettstein, D.; Hansson, M. Characterization of mutations in barley fch2 encoding chlorophyllide a oxygenase. Plant Cell Physiol. 2012, 53, 1232–1246. [Google Scholar] [CrossRef]
- Hirono, Y.; Redei, G.P. Induced Premeiotic Exchange of Linked Markers in the Angiosperm Arabidopsis. Genetics 1965, 51, 519–526. [Google Scholar] [PubMed]
- Dall’Osto, L.; Cazzaniga, S.; Havaux, M.; Bassi, R. Enhanced photoprotection by protein-bound vs free xanthophyll pools: A comparative analysis of chlorophyll b and xanthophyll biosynthesis mutants. Mol. Plant 2010, 3, 576–593. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Grimm, B. Comparative Analysis of Light-Harvesting Antennae and State Transition in chlorina and cpSRP Mutants. Plant Physiol. 2016, 172, 1519–1531. [Google Scholar] [CrossRef] [Green Version]
- Dall’Osto, L.; Cazzaniga, S.; North, H.; Marion-Poll, A.; Bassi, R. The Arabidopsis aba4-1 mutant reveals a specific function for neoxanthin in protection against photooxidative stress. Plant Cell 2007, 19, 1048–1064. [Google Scholar] [CrossRef] [Green Version]
- Havaux, M.; Dall’Osto, L.; Cuine, S.; Giuliano, G.; Bassi, R. The effect of zeaxanthin as the only xanthophyll on the structure and function of the photosynthetic apparatus in Arabidopsis thaliana. J. Biol. Chem. 2004, 279, 13878–13888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havaux, M.; Dall’osto, L.; Bassi, R. Zeaxanthin has enhanced antioxidant capacity with respect to all other xanthophylls in Arabidopsis leaves and functions independent of binding to PSII antennae. Plant Physiol. 2007, 145, 1506–1520. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T.; Tanaka, A.; Melis, A. Chlorophyll antenna size adjustments by irradiance in Dunaliella salina involve coordinate regulation of chlorophyll a oxygenase (CAO) and Lhcb gene expression. Plant Mol. Biol. 2003, 51, 757–771. [Google Scholar] [CrossRef]
- Krol, M.; Ivanov, A.G.; Jansson, S.; Kloppstech, K.; Huner, N.P. Greening under high light or cold temperature affects the level of xanthophyll-cycle pigments, early light-inducible proteins, and light-harvesting polypeptides in wild-type barley and the chlorina f2 mutant. Plant Physiol. 1999, 120, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Andersson, J.; Wentworth, M.; Walters, R.G.; Howard, C.A.; Ruban, A.V.; Horton, P.; Jansson, S. Absence of the Lhcb1 and Lhcb2 proteins of the light-harvesting complex of photosystem II—Effects on photosynthesis, grana stacking and fitness. Plant J. Cell Mol. Biol. 2003, 35, 350–361. [Google Scholar] [CrossRef]
- Puthiyaveetil, S.; Tsabari, O.; Lowry, T.; Lenhert, S.; Lewis, R.R.; Reich, Z.; Kirchhoff, H. Compartmentalization of the protein repair machinery in photosynthetic membranes. Proc. Natl. Acad. Sci. USA 2014, 111, 15839–15844. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y. Quality Control of Photosystem II: The Mechanisms for Avoidance and Tolerance of Light and Heat Stresses are Closely Linked to Membrane Fluidity of the Thylakoids. Front. Plant Sci. 2016, 7, 1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Lu, Y.; Hua, W.; Last, R.L. A New Light on Photosystem II Maintenance in Oxygenic Photosynthesis. Front. Plant Sci. 2019, 10, 975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dilnawaz, F.; Mohapatra, P.; Misra, M.; Ramaswamy, N.K.; Misra, A.N. The distinctive pattern of photosystem 2 activity, photosynthetic pigment accumulation, and ribulose-1,5-bisphosphate carboxylase/oxygenase content of chloroplasts along the axis of primary wheat leaf lamina. Photosynthetica 2001, 39, 557–563. [Google Scholar] [CrossRef]
- Garstka, M.; Drozak, A.; Rosiak, M.; Venema, J.H.; Kierdaszuk, B.; Simeonova, E.; van Hasselt, P.R.; Dobrucki, J.; Mostowska, A. Light-dependent reversal of dark-chilling induced changes in chloroplast structure and arrangement of chlorophyll-protein complexes in bean thylakoid membranes. BBA Bioenerg. 2005, 1710, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hipkins, M.F.; Baker, N.R. Photosynthetic Energy Transduction: A Practical Approach, 1st ed.; Pract. Approach Ser. 17; Oxford University Press: Oxford, UK, 1986; pp. 63–64. [Google Scholar]
- Kowalewska, L.; Mazur, R.; Suski, S.; Garstka, M.; Mostowska, A. Three-Dimensional Visualization of the Tubular-Lamellar Transformation of the Internal Plastid Membrane Network during Runner Bean Chloroplast Biogenesis. Plant Cell 2016, 28, 875–891. [Google Scholar] [CrossRef] [Green Version]
- Rumak, I.; Mazur, R.; Gieczewska, K.; Koziol-Lipinska, J.; Kierdaszuk, B.; Michalski, W.P.; Shiell, B.J.; Venema, J.H.; Vredenberg, W.J.; Mostowska, A.; et al. Correlation between spatial (3D) structure of pea and bean thylakoid membranes and arrangement of chlorophyll-protein complexes. BMC Plant Biol. 2012, 12, 72. [Google Scholar] [CrossRef] [Green Version]
- Szalonek, M.; Sierpien, B.; Rymaszewski, W.; Gieczewska, K.; Garstka, M.; Lichocka, M.; Sass, L.; Paul, K.; Vass, I.; Vankova, R.; et al. Potato Annexin STANN1 Promotes Drought Tolerance and Mitigates Light Stress in Transgenic Solanum tuberosum L. Plants. PLoS ONE 2015, 10, e0132683. [Google Scholar] [CrossRef]
- Sztatelman, O.; Grzyb, J.; Gabrys, H.; Banas, A.K. The effect of UV-B on Arabidopsis leaves depends on light conditions after treatment. BMC Plant Biol. 2015, 15, 281. [Google Scholar] [CrossRef] [Green Version]
- Mazur, R.; Mostowska, A.; Szach, J.; Gieczewska, K.; Wojtowicz, J.; Bednarska, K.; Garstka, M.; Kowalewska, L. Galactolipid deficiency disturbs spatial arrangement of the thylakoid network in Arabidopsis thaliana plants. J. Exp. Bot. 2019, 70, 4689–4703. [Google Scholar] [CrossRef] [Green Version]
- Czechowski, T.; Stitt, M.; Altmann, T.; Udvardi, M.K.; Scheible, W.R. Genome-wide identification and testing of superior reference genes for transcript normalization in Arabidopsis. Plant Physiol. 2005, 139, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.H.; Liu, R.; Yan, L.; Liu, Z.Q.; Jiang, S.C.; Shen, Y.Y.; Wang, X.F.; Zhang, D.P. Light-harvesting chlorophyll a/b-binding proteins are required for stomatal response to abscisic acid in Arabidopsis. J. Exp. Bot. 2012, 63, 1095–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ler-0 NL | NW41 NL | Col-1 NL | ch1.1 NL | ch1.2 NL | ch1.3 NL | |
|---|---|---|---|---|---|---|
| Chla/b | 2.26 | 9.11 * | 2.88 | ND * | ND * | 3.37 * |
| ±0.16 | ±0.55 | ±0.04 | - | - | ±0.08 | |
| Fv/Fm | 0.815 | 0.747 * | 0.825 | 0.76 * | 0.734 * | 0.776 * |
| ±0.01 | ±0.08 | ±0.01 | ±0.01 | ±0.02 | ±0.02 | |
| F0 | 0.435 | 0.595 | 0.513 | 0.146 * | 0.255 * | 0.587 |
| ±0.10 | ±0.16 | ±0.08 | ±0.08 | ±0.08 | ±0.09 | |
| Fm | 2.345 | 2.408 | 2.86 | 0.631 * | 0.894 * | 2.553 |
| ±0.61 | ±0.48 | ±0.42 | ±0.06 | ±0.17 | ±0.05 | |
| Y(II) | 0.494 | 0.300 * | 0.435 | 0.601 | 0.542 | 0.256 |
| ±0.01 | ±0.17 | ±0.03 | ±0.09 | ±0.10 | ±0.04 | |
| Y(I) | 0.803 | 0.552 * | 0.723 | 0.767 | 0.597 | 0.640 |
| ±0.02 | ±0.10 | ±0.03 | ±0.02 | ±0.06 | ±0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wójtowicz, J.; Jagielski, A.K.; Mostowska, A.; Gieczewska, K.B. Compensation Mechanism of the Photosynthetic Apparatus in Arabidopsis thaliana ch1 Mutants. Int. J. Mol. Sci. 2021, 22, 221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010221
Wójtowicz J, Jagielski AK, Mostowska A, Gieczewska KB. Compensation Mechanism of the Photosynthetic Apparatus in Arabidopsis thaliana ch1 Mutants. International Journal of Molecular Sciences. 2021; 22(1):221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010221
Chicago/Turabian StyleWójtowicz, Joanna, Adam K. Jagielski, Agnieszka Mostowska, and Katarzyna B. Gieczewska. 2021. "Compensation Mechanism of the Photosynthetic Apparatus in Arabidopsis thaliana ch1 Mutants" International Journal of Molecular Sciences 22, no. 1: 221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010221