The Status of EGFR Modulates the Effect of miRNA-200c on ZEB1 Expression and Cell Migration in Glioblastoma Cells

 , ,
, ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
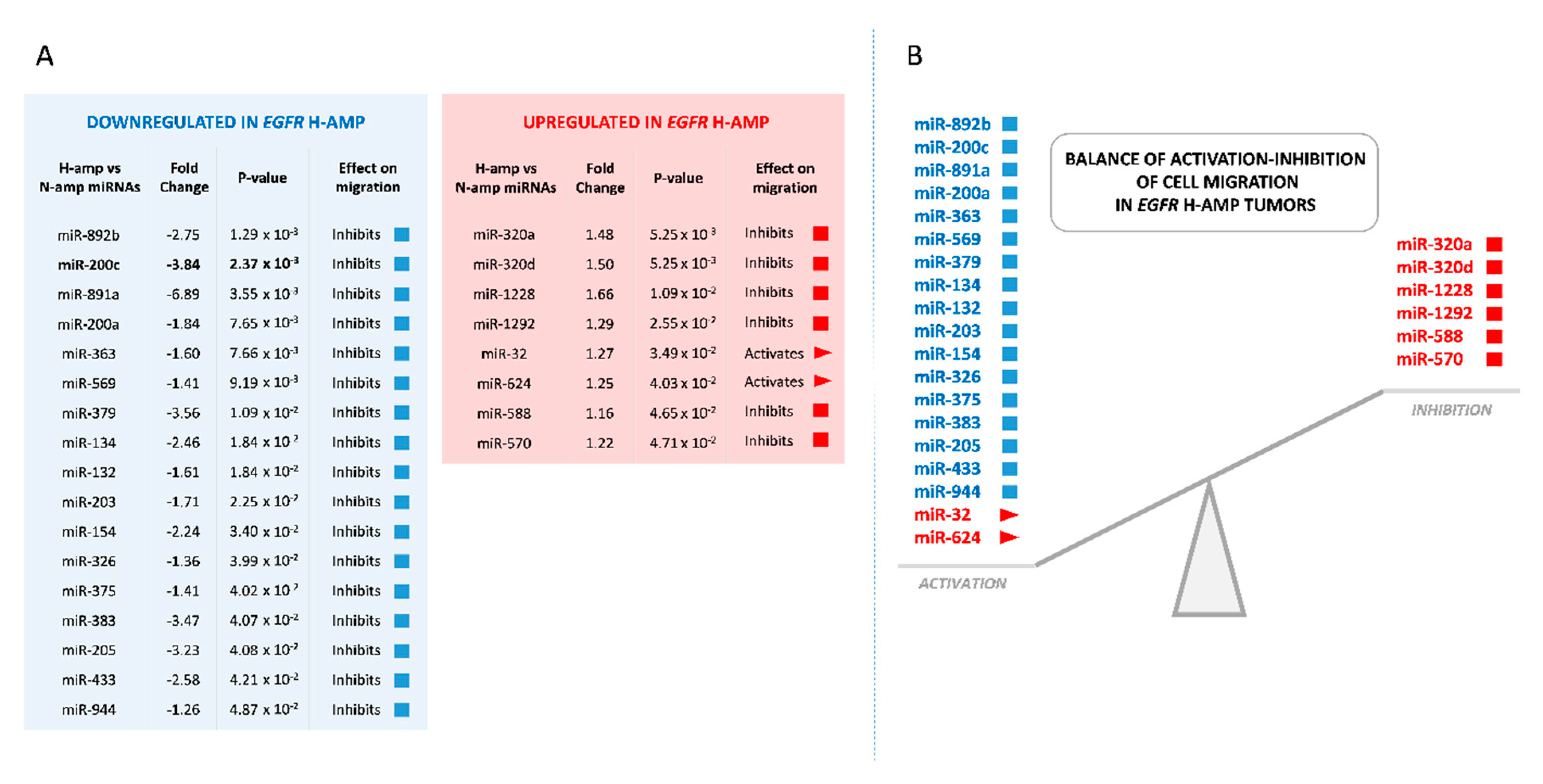
2.1. Whole-Genome miRNA Analysis and Cell Migration Biological Analysis
2.2. Downregulation of miR-200c in H-amp Tumors May Promote Cell Migration
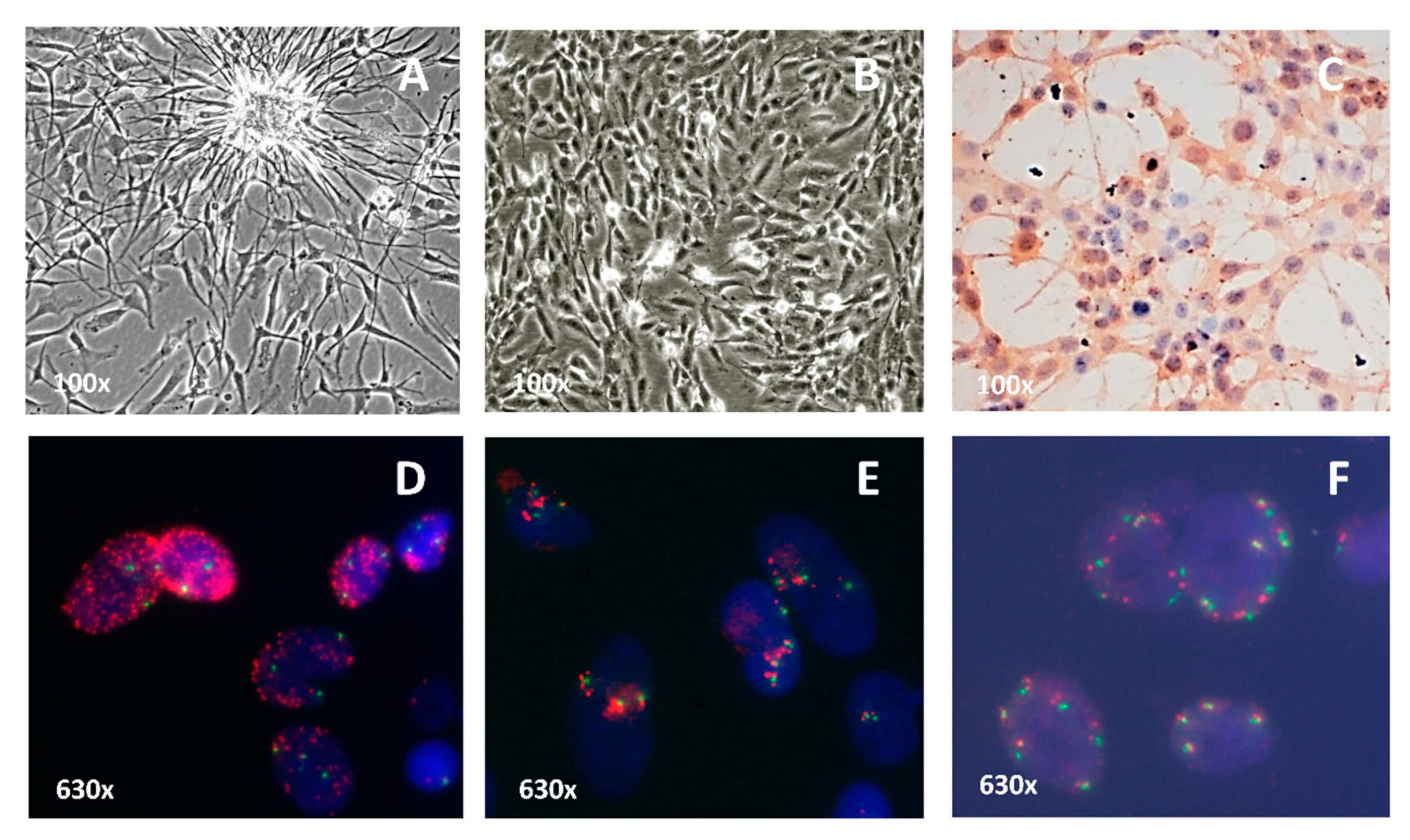
2.3. In Vitro Model of EGFR Amplification: Cell Culture Characterization
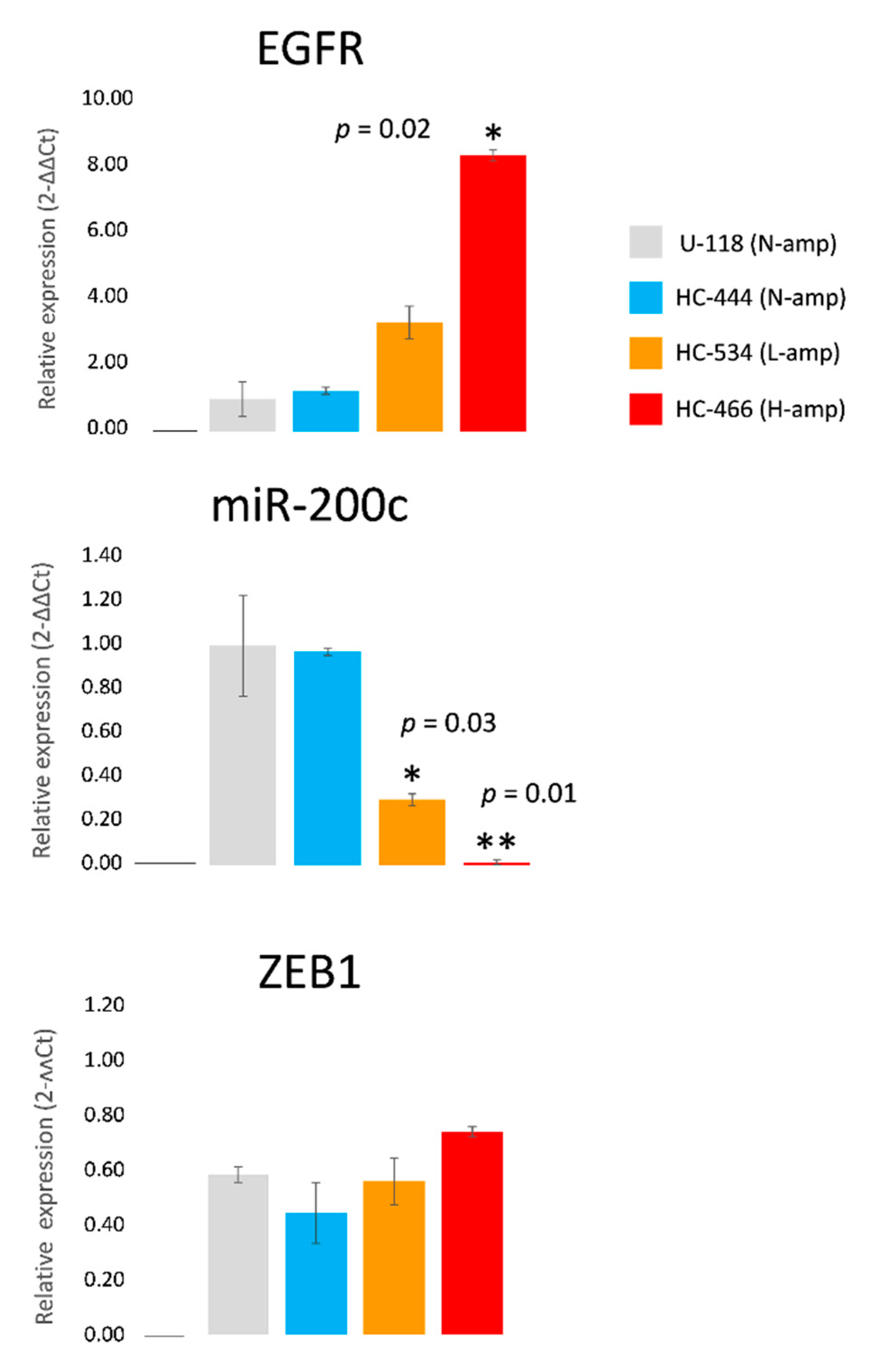
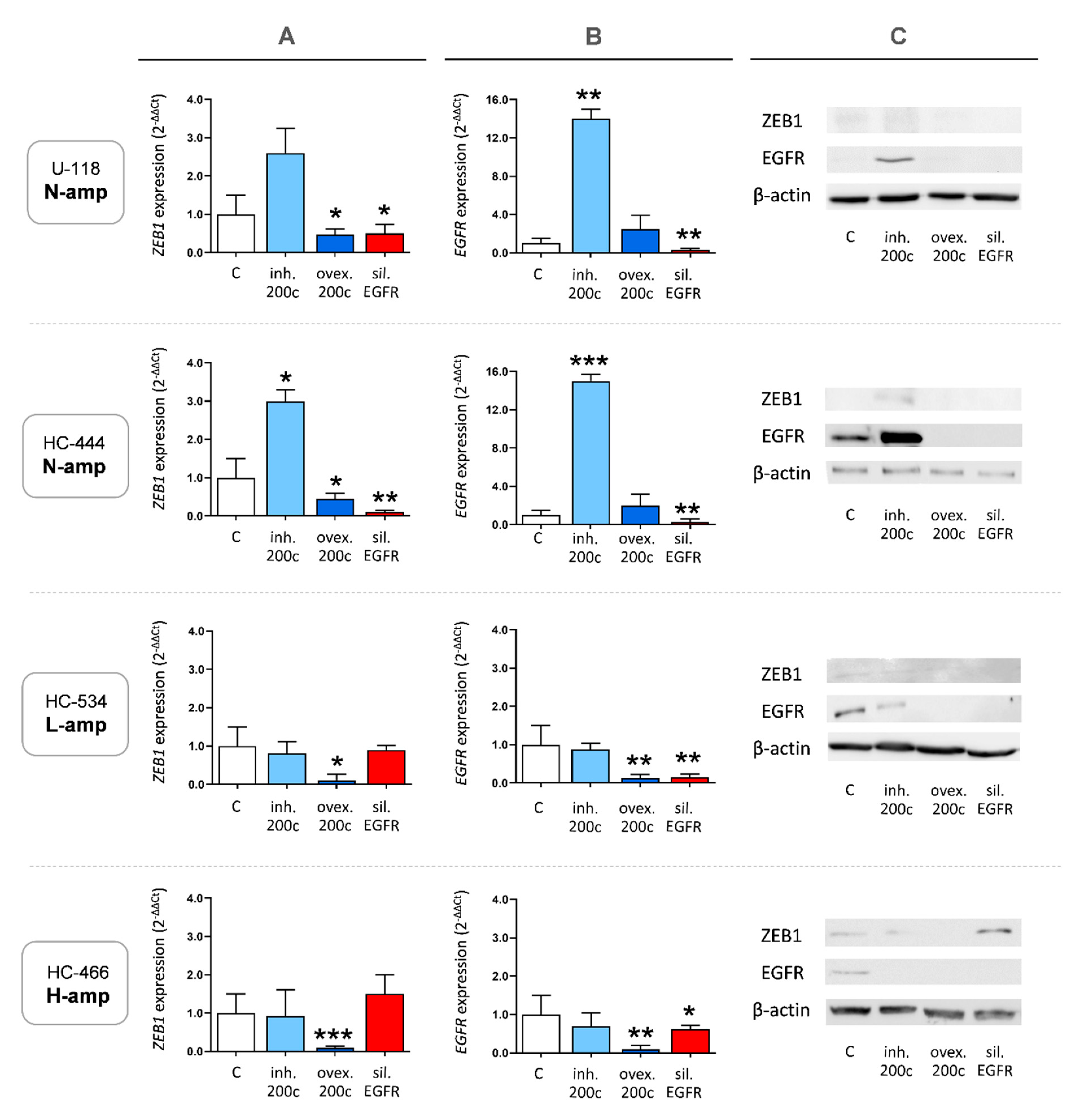
2.4. EGFR, miR-200c and ZEB1 Expression in Cell Cultures
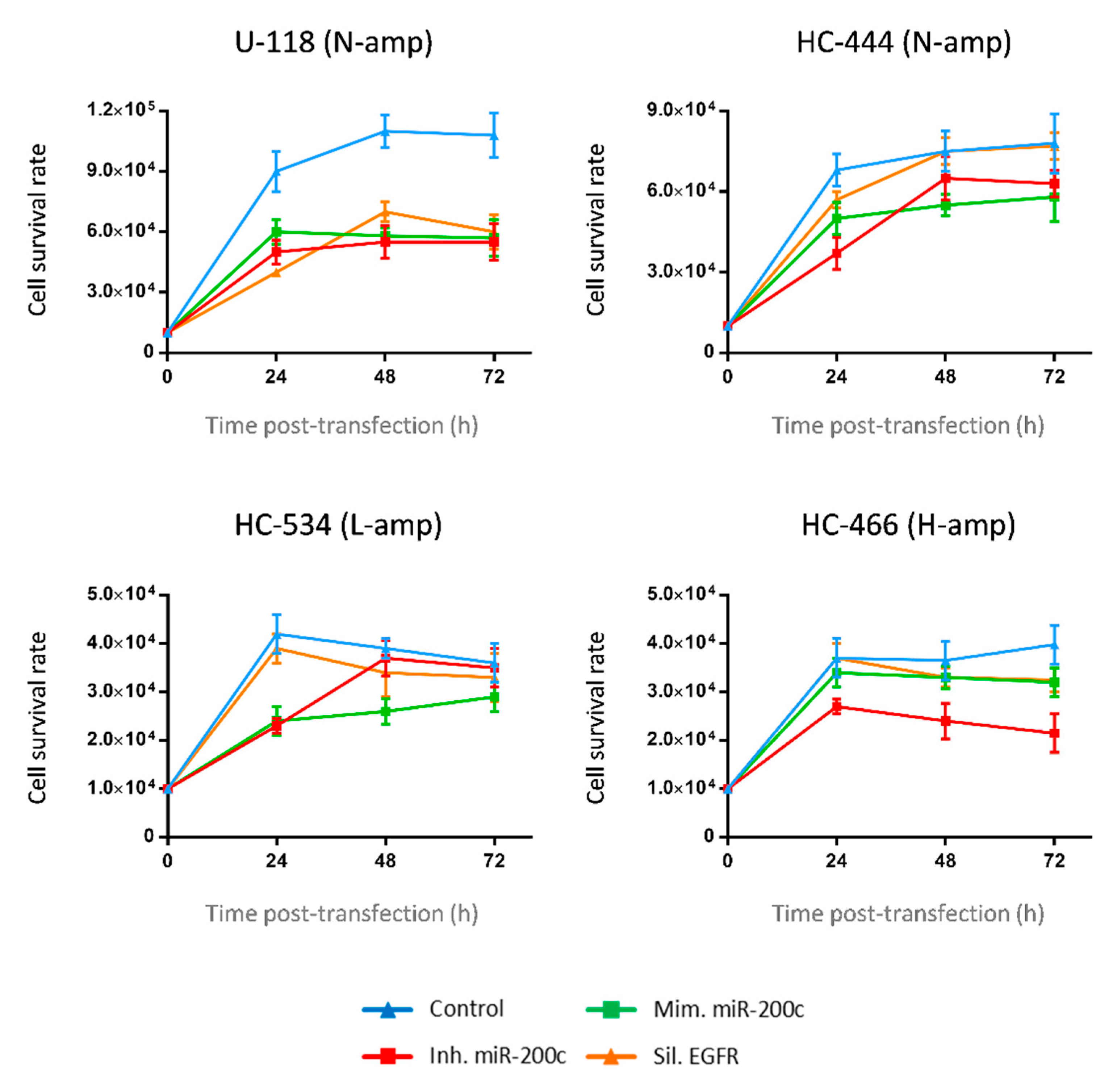
2.5. Transfection Efficiency and Cell Viability in GBM Transfected Cultures
2.6. MiR-200c Modulates ZEB1 and EGFR Expression in Cultures without EGFR Amplification
2.7. Overexpression of miR-200c Diminishes ZEB1 and EGFR Expression in Cultures with EGFR Amplification
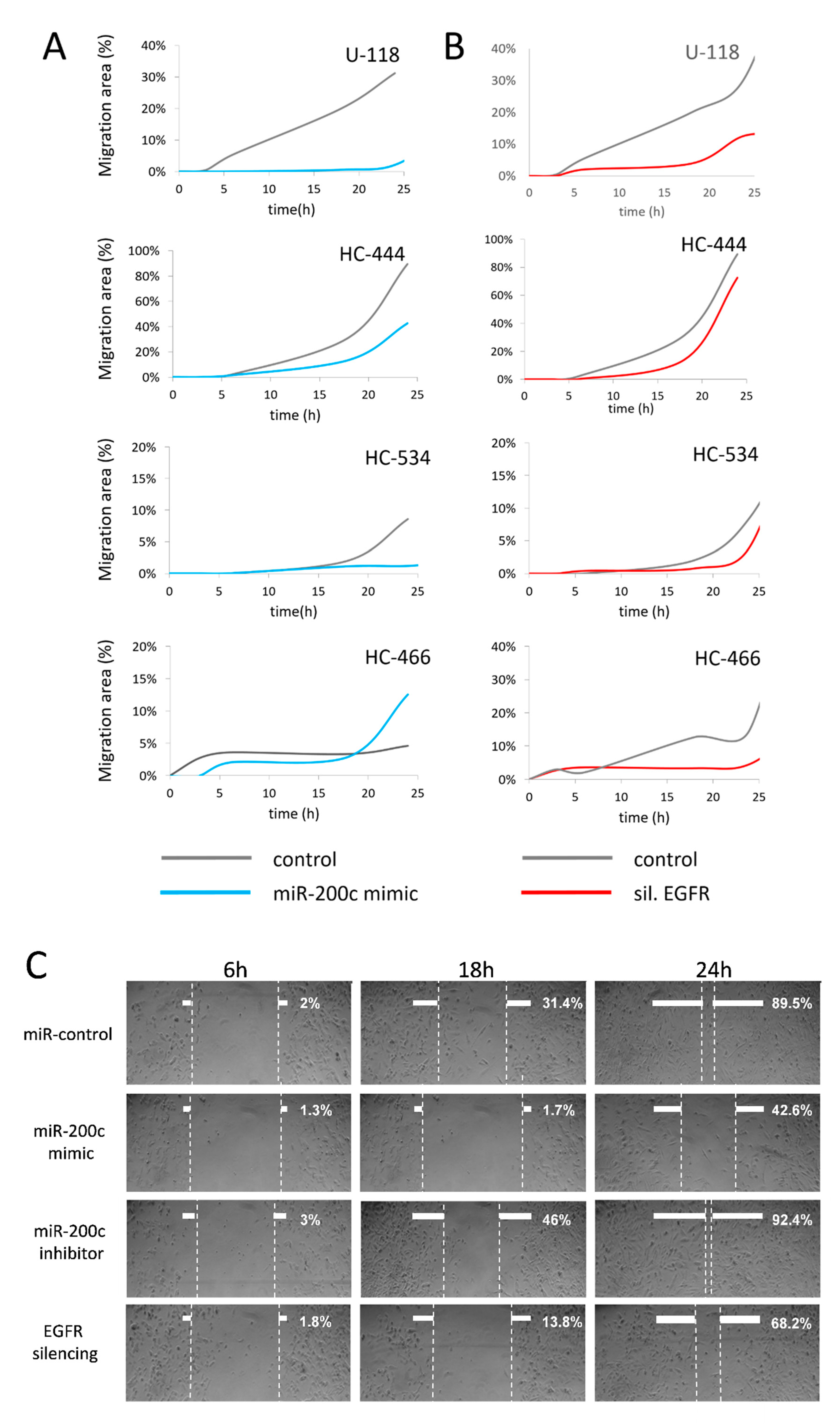
2.8. MiR-200c Upregulation in GBM Cell Cultures Inhibits Cell Migration
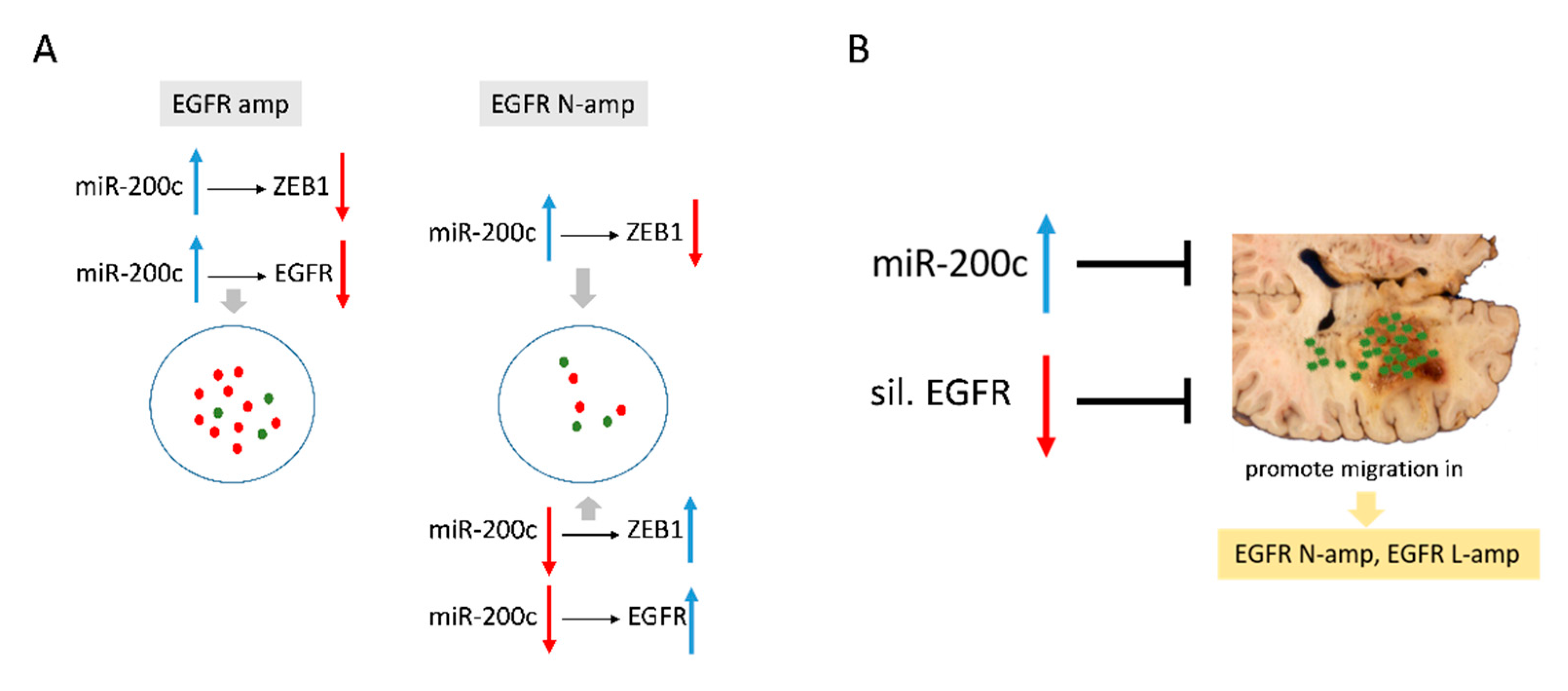
3. Discussion
4. Materials and Methods
4.1. Tumor Samples
4.2. MiRNA Expression Analysis in GBM Tumors
4.3. Cell Cultures and Treatment
4.4. Immunophenotypical Analysis
4.5. Conventional Cytogenetics and Fluorescence in Situ Hybridization (FISH)
4.6. Multiplex Ligation-Dependent Probe Amplification (MLPA)
4.7. RNA Extraction and Quantitative Real-Time PCR
4.8. Western Blot Analysis
4.9. Cell Transfection
4.10. Luminescence Cell Viability Assay
4.11. Wound Healing Assay
4.12. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behin, A.; Hoang-Xuan, K.; Carpentier, A.F.; Delattre, J.Y. Primary brain tumours in adults. Lancet 2003, 361, 323–331. [Google Scholar] [CrossRef]
- Hess, K. Extent of resection as a prognostic variable in the treatment of gliomas. Neurooncology 1999, 42, 227–301. [Google Scholar] [CrossRef] [PubMed]
- Lefranc, F.; Le Rhun, E.; Kiss, R.; Weller, M. Glioblastoma quo vadis: Will migration and invasiveness reemerge as therapeutic targets? Cancer Treat. Rev. 2018, 68, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Helson, L.; Majeed, M. Pleiotropic Chemotherapy to Abrogate Glioblastoma Multiforme Migration/Invasion. Anticancer. Res. 2019, 39, 3423–3427. [Google Scholar] [CrossRef] [PubMed]
- Huncharek, M.; Kupelnick, B. Epidermal growth factor receptor gene amplification as a prognostic marker in glioblastoma: Results of a meta-analysis. Oncol. Res. 2000, 12, 107–112. [Google Scholar] [CrossRef]
- Rao, R.; James, C. Altered molecular pathways in gliomas: An overview of clinically relevant issues. Semin. Oncol. 2004, 31, 595–604. [Google Scholar] [CrossRef]
- López-Ginés, L.; Gil-Benso, R.; Ferrer-Luna, R.; Benito, R.; Serna, E.; González-Darder, J.; Quilis, V.; Monleón, D.; Celda, B.; Cerdá-Nicolas, M. New pattern of EGFR amplification in glioblastoma and the relationship of gene copy number with gene expression profile. Mod. Pathol. 2010, 23, 856–865. [Google Scholar] [CrossRef]
- Márquez, A.; Wu, R.; Zhao, J.; Tao, J.; Shi, Z. Evaluation of epidermal growth factor receptor (EGFR) by chromogenic in situ hybridization (CISH) and immunohistochemistry (IHC) in archival gliomas using bright-field microscopy. Diagn Mol. Pathol. 2004, 13, 1–8. [Google Scholar] [CrossRef]
- Sauter, G.; Maeda, T.; Waldman, F.M.; Davis, R.L.; Feuerstein, B.G. Short communication: Patterns of epidermal growth factor receptor amplification in malignant gliomas. Am. J. Pathol. 1996, 148, 1047–1053. [Google Scholar]
- Kuwahara, Y.; Tanabe, C.; Ikeuchi, T.; Aoyagi, K.; Nishigaki, M.; Sakamoto, H.; Hoshinaga, K.; Yoshida, T.; Sasaki, H.; Terada, M. Alternative mechanisms of gene amplification in human cancers. Genes Chromosomes Cancer. 2004, 41, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.; Jung, S.; Keller, S.; Gregory, R.I.; Diederichs, S. Many roads to maturity: microRNA biogenesis pathways and their regulation. Nat Cell Biol. 2009, 11, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Ciafrè, S.A.; Galardi, S.; Mangiola, A.; Ferracin, M.; Liu, C.G.; Sabatino, G.; Negrini, M.; Maira, G.; Croce, C.M.; Farace, M.G. Extensive modulation of a set of microRNAs in primary glioblastoma. Biochem. Biophys. Res. Commun. 2005, 334, 1351–1358. [Google Scholar]
- Møller, H.G.; Rasmussen, A.P.; Andersen, H.H.; Johnsen, K.B.; Henriksen, M.; Duroux, M. A systematic review of microRNA in glioblastoma multiforme: Micromodulatorsin the mesenchymal mode of migration and invasion. Mol. Neurobiol. 2013, 47, 131–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serna, E.; López-Ginés, C.; Monleón, D.; Muñoz-Hidalgo, L.; Callaghan, R.C.; Gil-Benso, R.; Martinetto, H.; Gregori-Romero, A.; Gonzalez-Darder, J.; Cerdá-Nicolás, M. Correlation between EGFR amplification and the expression of microRNA-200c in primary glioblastoma. PLoS ONE 2014, 9, e102927. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.W.; Wang, X.; Yang, Y.; Mao, Q. Role of micro-RNA (miRNA) in pathogenesis of glioblastoma. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 1630–1639. [Google Scholar] [PubMed]
- Saadatpour, L.; Fadaee, E.; Fadaei, S.; Mansour, R.N.; Mohammadi, M.; Mousavi, S.M.; Goodharzi, M.; Verdi, J.; Mirzaei, H. Glioblastoma: Exosome and microRNA as novel diagnosis biomarkers. Cancer Gene Ther. 2016, 23, 415–418. [Google Scholar] [CrossRef]
- Kalkan, R.; Atli, E.I. The Impacts of miRNAs in Glioblastoma Progression. Crit. Rev. Eukaryot Gene Expr. 2016, 26, 137–142. [Google Scholar] [CrossRef]
- Korpal, M.; Lee, E.S.; Hu, G.; Kang, Y. The miR-200 family inhibitsepithelial-mesenchymal transition and cancer cell migration by direct targeting of E-cadherin transcriptional repressors ZEB1 and ZEB2. J. Biol. Chem. 2008, 283, 14910–14914. [Google Scholar] [CrossRef] [Green Version]
- Paterson, E.L.; Kolesnikoff, N.; Gregory, P.A.; Bert, A.G.; Khew-Goodall, Y.; Goodall, G.J. The microRNA-200 family regulates epithelial to mesenchymal transition. Sci. World J. 2008, 8, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Burk, U.; Schubert, J.; Wellner, U.; Schmalhofer, O.; Vincan, E.; Spaderna, S.; Brabletz, M. A reciprocal repression between ZEB1 and members of the miR-200 family promotes EMT and invasion in cancer cells. EMBO Rep. 2008, 9, 582–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.M.; Gaur, A.B.; Lengyel, E.; Peter, M.E. The miR-200 family determines the epithelial phenotype of cancer cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes Dev. 2008, 22, 894–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utsuki, S.; Sato, Y.; Oka, H.; Tsuchiya, B.; Suzuki, S.; Fujii, K. Relationship between the expression of E-, N-cadherins and beta-catenin and tumor grade in astrocytomas. J. Neurooncol. 2002, 57, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Lewis-Tuffin, L.J.; Rodríguez, F.; Giannini, C.; Scheithauer, B.; Necela, B.M.; Sarkaria, J.N.; Anastasiadis, P.Z. Misregulated E-cadherin expression associated with an aggressive brain tumor phenotype. PLoS ONE 2010, 5, e13665. [Google Scholar] [CrossRef]
- Edwards, L.A.; Woolard, K.; Son, M.J.; Li, A.; Lee, J.; Ene, C.; Mantey, S.A.; Maric, D.; Song, H.; Belova, G.; et al. Effect of brain and tumor-derived connective tissue growth factor on glioma invasion. J. Natl. Cancer Inst. 2011, 103, 1162–1178. [Google Scholar] [CrossRef]
- Kahlert, U.D.; Maciaczyk, D.; Doostkam, S.; Orr, B.A.; Simons, B.; Bogiel, T.; Reithmeier, T.; Prinz, M.; Schubert, J.; Niedermann, G.; et al. Activation of canonical WNT/b-catenin signaling enhances in vitro motility of glioblastoma cells by activation of ZEB1 and other activators of epithelial-to mesenchymal transition. Cancer Lett. 2012, 325, 42–53. [Google Scholar] [CrossRef]
- Siebzehnrubl, F.A.; Silver, D.J.; Tugertimur, B.; Deleyrolle, L.P.; Siebzehnrubl, D.; Sarkisian, M.R.; Devers, K.G.; Yachnis, A.T.; Kupper, M.D.; Neal, D.; et al. The ZEB1 pathway links glioblastoma initiation, invasion and chemoresistance. EMBO Mol. Med. 2013, 5, 1196–1212. [Google Scholar] [CrossRef]
- Guo, E.; Wang, Z.; Wang, S. MiR-200c and miR-141 inhibit ZEB1 synergistically and suppress glioma cell growth and migration. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 3385–3391. [Google Scholar]
- Giacomelli, C.; Daniele, S.; Natali, L.; Iofrida, C.; Flamini, G.; Braca, A.; Trincavelli, M.L.; Martini, C. Carnosol controls the human glioblastoma stemness features through the epithelial-mesenchymal transition modulation and the induction of cancer stem cell apoptosis. Sci. Rep. 2017, 7, 15174. [Google Scholar] [CrossRef]
- Singh, D.K.; Kollipara, R.K.; Vemireddy, V.; Yang, X.L.; Sun, Y.; Regmi, N.; Klingler, S.; Hatanpaa, K.J.; Raisanen, J.; Cho, S.K. Oncogenes activate an autonomous transcriptional regulatory circuit that drives glioblastoma. Cell Rep. 2017, 18, 961–976. [Google Scholar] [CrossRef]
- Hurteau, G.J.; Carlson, J.A.; Spivack, S.D.; Brock, G.J. Overexpression of the microRNA hsa-miR-200c leads to reduced expression of transcription factor 8 and increased expression of E-cadherin. Cancer Res. 2007, 67, 7972–7976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosmaninho, P.; Mükusch, S.; Piscopo, V.; Teixeira, V.; Raposo, A.A.; Warta, R.; Bennewitz, R.; Tang, Y.; Herold-Mende, C.; Stifani, S. Zeb1 potentiates genome-wide gene transcription with Lef1 to promote glioblastoma cell invasion. EMBO J. 2018, 1, e97115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, Z.B.; Ren, W.M.; Ye, X.G.; Zhang, Y.Y. The miR-200 family regulates the epithelial-mesenchymal transition induced by EGF/EGFR in anaplastic thyroid cancer cells. Int. J. Mol. Med. 2012, 30, 856–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- TCGA. Molecular Profiling Reveals Biologically Discrete Subsets and Pathways of Progression in Diffuse Glioma. Cell 2016, 164, 550–563. [Google Scholar] [CrossRef] [PubMed]
- Kefas, B.; Godlewski, J.; Comeau, L.; Li, Y.; Abounader, R.; Hawkinson, M.; Lee, J.; Fine, H.; Chiocca, E.A.; Lawler, S.; et al. MicroRNA-7 inhibits the epidermal growth factor receptor and the Akt pathway and is down-regulated in glioblastoma. Cancer Res. 2008, 68, 3566–3572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchini, S.; Cavalieri, D.; Fruscio, R.; Calura, E.; Garavaglia, D.; Fuso, I.; Mangioni, C.; Cattoretti, G.; Clivio, L.; Beltrame, L.; et al. Association between miR-200c and the survival of patients with stage I epithelial ovarian cancer: A retrospective study of two independent tumour tissue collections. Lancet Oncol. 2011, 12, 273–285. [Google Scholar] [CrossRef]
- Cesarini, V.; Silvestris, D.A.; Tassinari, V.; Tomaselli, S.; Alon, S.; Eisenberg, E.; Locatelli, F.; Gallo, A. ADAR2/miR-589-3p axis controls glioblastoma cell migration/invasion. Nucleic Acids Res. 2018, 46, 2045–2059. [Google Scholar] [CrossRef]
- Xu, T.J.; Qiu, P.; Zhang, Y.B.; Yu, S.Y.; Xu, G.M.; Yang, W. MiR-148a inhibits the proliferation and migration of glioblastoma by targeting ITGA9. Hum. Cell. 2019, 32, 548–556. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, J.; Hui, X.; Wang, W.; Fang, D.; Ding, L. Mir-758-5p Suppresses Glioblastoma Proliferation, Migration and Invasion by Targeting ZBTB20. Cell Physiol. Biochem. 2018, 48, 2074–2083. [Google Scholar] [CrossRef]
- Adam, L.; Zhong, M.; Choi, W.; Qi, W.; Nicoloso, M.; Arora, A.; Calin, G.; Wang, H.; Siefker-Radtke, A.; McConkey, D.; et al. MiR-200 expression regulates epithelial-to-mesenchymal transition in bladder cancer cells and reverses resistance to epidermal growth factor receptor therapy. Clin. Cancer Res. 2009, 15, 5060–5072. [Google Scholar] [CrossRef] [Green Version]
- Bockmeyer, C.L.; Christgen, M.; Müller, M.; Fischer, S.; Ahrens, P.; Länger, F.; Kreipe, H.; Lehmann, U. MicroRNA profiles of healthy basal and luminal mammary epithelial cells are distinct and reflected in different breast cancer subtypes. Breast Cancer Res. Treat. 2011, 130, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Men, D.; Liang, Y.; Chen, L. Decreased expression ofmicroRNA-200b is an independent unfavorable prognostic factor for glioma patients. Cancer Epidemiol. 2014, 38, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Tang, H.; Liu, X.; Liao, Y.; Li, H.; Zhao, Z.; Yuan, X.; Jiang, W. MiR-200b as a prognostic factor targets multiple members of RAB family in glioma. Med. Oncol. 2014, 31, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, L.; He, L.; Wang, J.; Shi, X.; Li, Z.; Hou, K.; Teng, Y.; Qu, X. MiR-891a-5p as a prognostic marker and therapeutic target for hormone receptor-positive breast cancer. J. Cancer 2020, 11, 3771–3782. [Google Scholar] [CrossRef]
- Shin, S.S.; Park, S.S.; Hwang, B.; Moon, B.; Kim, W.T.; Kim, W.J.; Moon, S.K. MicroRNA-892b influences proliferation, migration and invasion of bladder cancer cells by mediating the p19ARF/cyclin D1/CDK6 and Sp-1/MMP-9 pathways. Oncol. Rep. 2016, 36, 2313–2320. [Google Scholar] [CrossRef]
- Zhen, Q.; Liu, J.; Gao, L.; Liu, J.; Wang, R.; Chu, W.; Zhang, Y.; Tan, G.; Zhao, X.; Lv, B. MicroRNA-200a Targets EGFR and c-Met to Inhibit Migration, Invasion, and Gefitinib Resistance in Non-Small Cell Lung Cancer. Cytogenet. Genome Res. 2015, 146, 1–8. [Google Scholar] [CrossRef]
- Wu, X.; Wu, G.; Wu, Z.; Yao, X.; Li, G. MiR-200a Suppresses the Proliferation and Metastasis in Pancreatic Ductal Adenocarcinoma through Downregulation of DEK Gene. Transl. Oncol. 2016, 9, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sun, J.; Cai, Y.; Jiang, Y.; Wang, X.; Huang, X.; Yin, Y.; Li, H. MiR-200a acts as an oncogene in colorectal carcinoma by targeting PTEN. Exp. Mol. Pathol. 2016, 101, 308–313. [Google Scholar] [CrossRef]
- Bendoraite, A.; Knouf, E.C.; Garg, K.S.; Parkin, R.K.; Kroh, E.M.; O’Briant, K.C.; Ventura, A.P.; Godwin, A.K.; Karlan, B.Y.; Drescher, C.W.; et al. Regulation of miR-200 family microRNAs and ZEB transcription factors in ovarian cancer: Evidence supporting a mesothelial-to-epithelial transition. Gynecol. Oncol. 2010, 116, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Kahlert, U.D.; Nikkhah, G.; Maciaczyk, J. Epithelial-to-mesenchymal(-like) transition as a relevant molecular event in malignant gliomas. Cancer Lett. 2013, 331, 131–138. [Google Scholar] [CrossRef]
- Han, S.P.; Kim, J.H.; Han, M.E.; Sim, H.E.; Kim, K.S.; Yoon, S.; Baek, S.Y.; Kim, B.S.; Oh, S.O. SNAI1 is involved in the proliferation and migration of glioblastoma cells. Cell Mol. Neurobiol. 2011, 31, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Kahlert, U.D.; Suwala, A.K.; Raabe, E.H.; Siebzehnrubl, F.A.; Suarez, M.J.; Orr, B.A.; Bar, E.E.; Maciaczyk, J.; Eberhart, C.G. ZEB1 Promotes invasion in human fetal neural stem cells and hypoxic glioma neurospheres. Brain Pathol. 2014, 25, 724–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, F.; Kubota, Y.; Natsuizaka, M.; Maehara, O.; Hatanaka, Y.; Marukawa, K.; Terashita, K.; Suda, G.; Ohnishi, S.; Shimizu, Y.; et al. EGFR inhibitors prevent induction of cancer stem-like cells in esophageal squamous cell carcinoma by suppressing epithelial-mesenchymal transition. Cancer Biol. Ther. 2015, 16, 933–940. [Google Scholar] [CrossRef] [PubMed]
- McGowan-Jordan, J.; Simons, A.; Schmid, M. ISCN: An International System for Human Nomenclature; Karger: Basel, Switzerland, 2020. [Google Scholar]
- Layfield, L.J.; Willmore, C.; Tripp, S.; Jones, C.; Jensen, R.L. Epidermal growth factor receptor in amplification and and protein expression in glioblastoma multiforme: Prognostic significance and relationship to other prognostic factors. Appl. Immunohistochem. Mol. Morphol. 2006, 91, 91–96. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz-Hidalgo, L.; San-Miguel, T.; Megías, J.; Serna, E.; Calabuig-Fariñas, S.; Monleón, D.; Gil-Benso, R.; Cerdá-Nicolás, M.; López-Ginés, C. The Status of EGFR Modulates the Effect of miRNA-200c on ZEB1 Expression and Cell Migration in Glioblastoma Cells. Int. J. Mol. Sci. 2021, 22, 368. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010368
Muñoz-Hidalgo L, San-Miguel T, Megías J, Serna E, Calabuig-Fariñas S, Monleón D, Gil-Benso R, Cerdá-Nicolás M, López-Ginés C. The Status of EGFR Modulates the Effect of miRNA-200c on ZEB1 Expression and Cell Migration in Glioblastoma Cells. International Journal of Molecular Sciences. 2021; 22(1):368. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010368
Chicago/Turabian StyleMuñoz-Hidalgo, Lisandra, Teresa San-Miguel, Javier Megías, Eva Serna, Silvia Calabuig-Fariñas, Daniel Monleón, Rosario Gil-Benso, Miguel Cerdá-Nicolás, and Concha López-Ginés. 2021. "The Status of EGFR Modulates the Effect of miRNA-200c on ZEB1 Expression and Cell Migration in Glioblastoma Cells" International Journal of Molecular Sciences 22, no. 1: 368. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010368