Functional Coupling of Slack Channels and P2X3 Receptors Contributes to Neuropathic Pain Processing

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
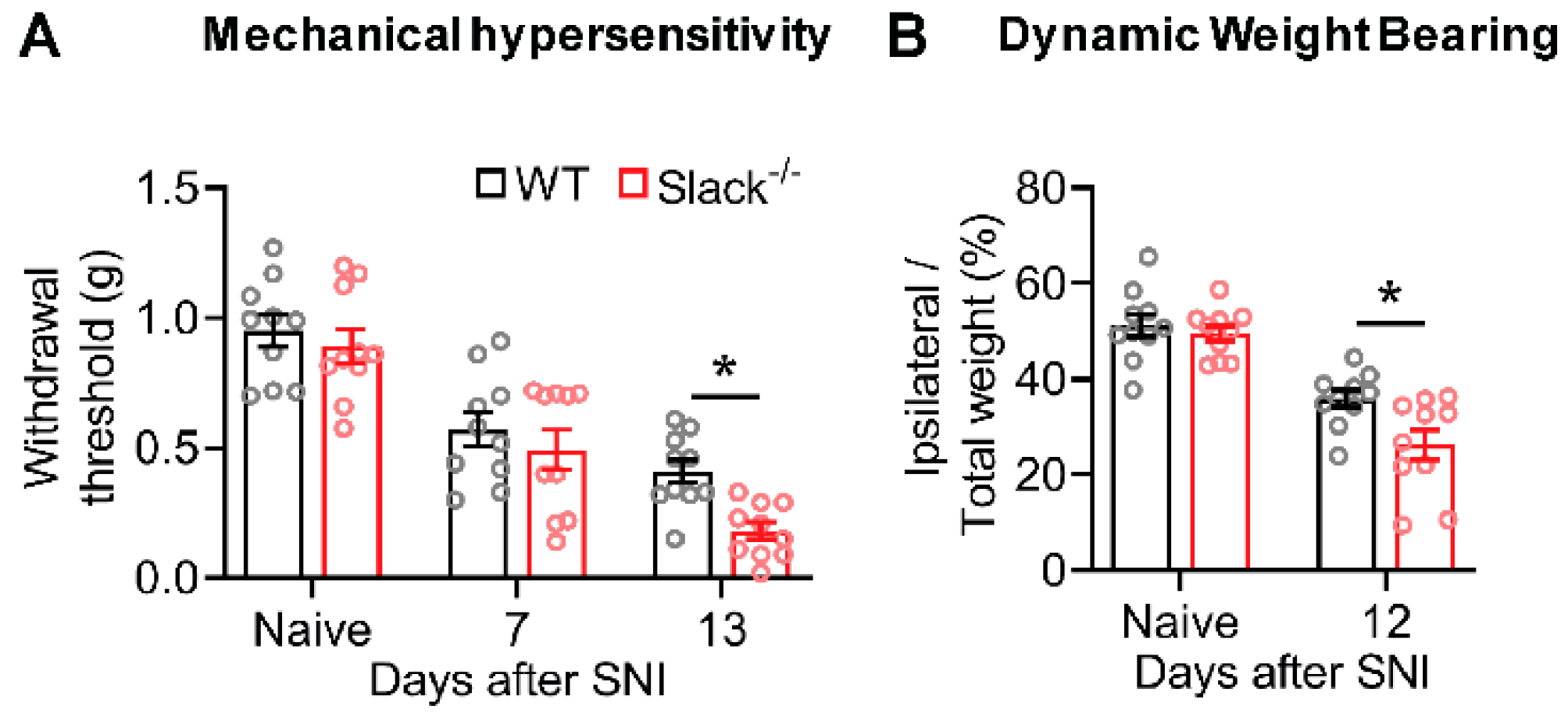
2.1. Slack-/- Mice Show Increased Neuropathic Pain Behavior after Peripheral Nerve Injury
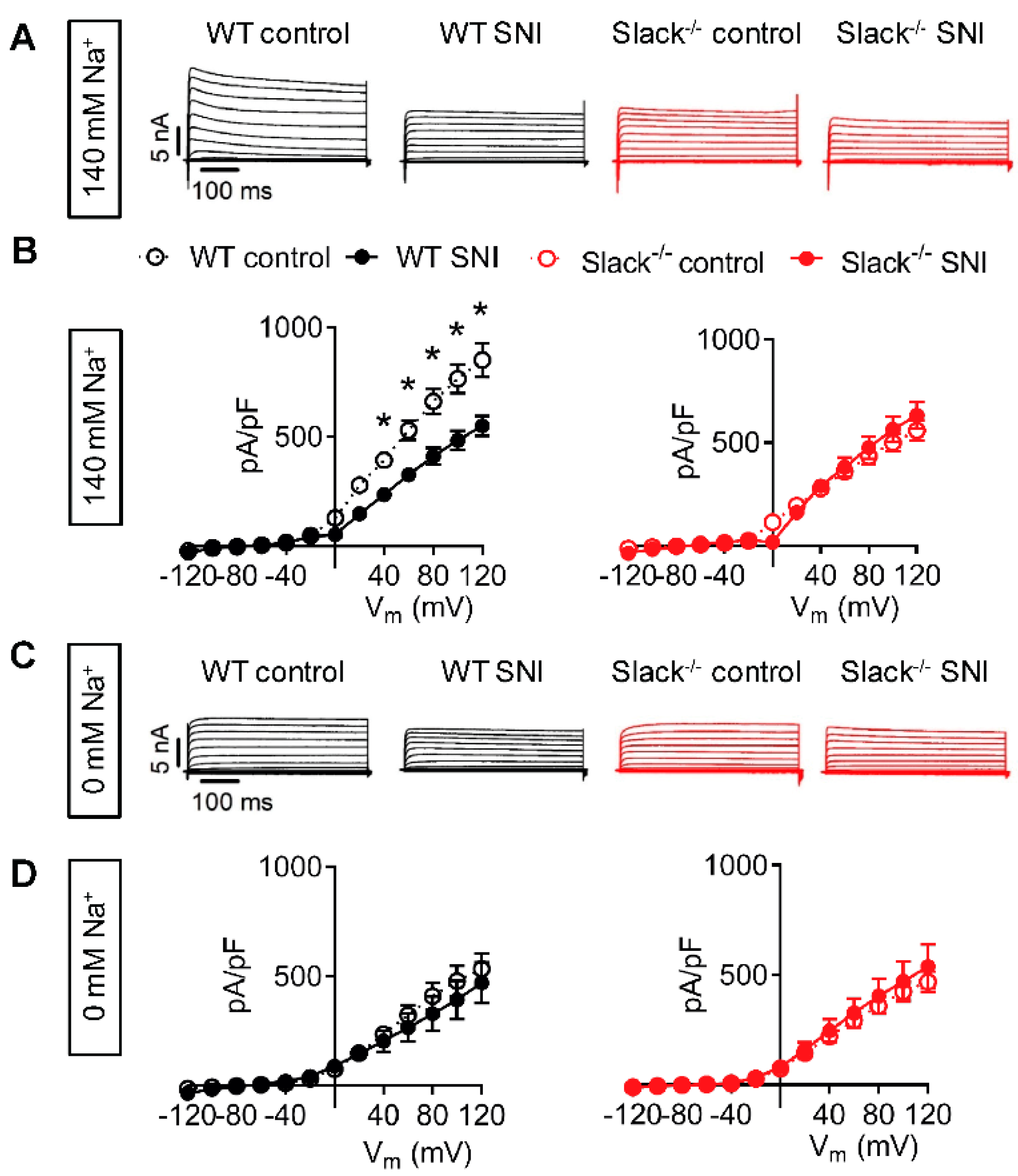
2.2. Slack-Mediated Potassium Currents in Sensory Neurons are Reduced after Nerve Injury
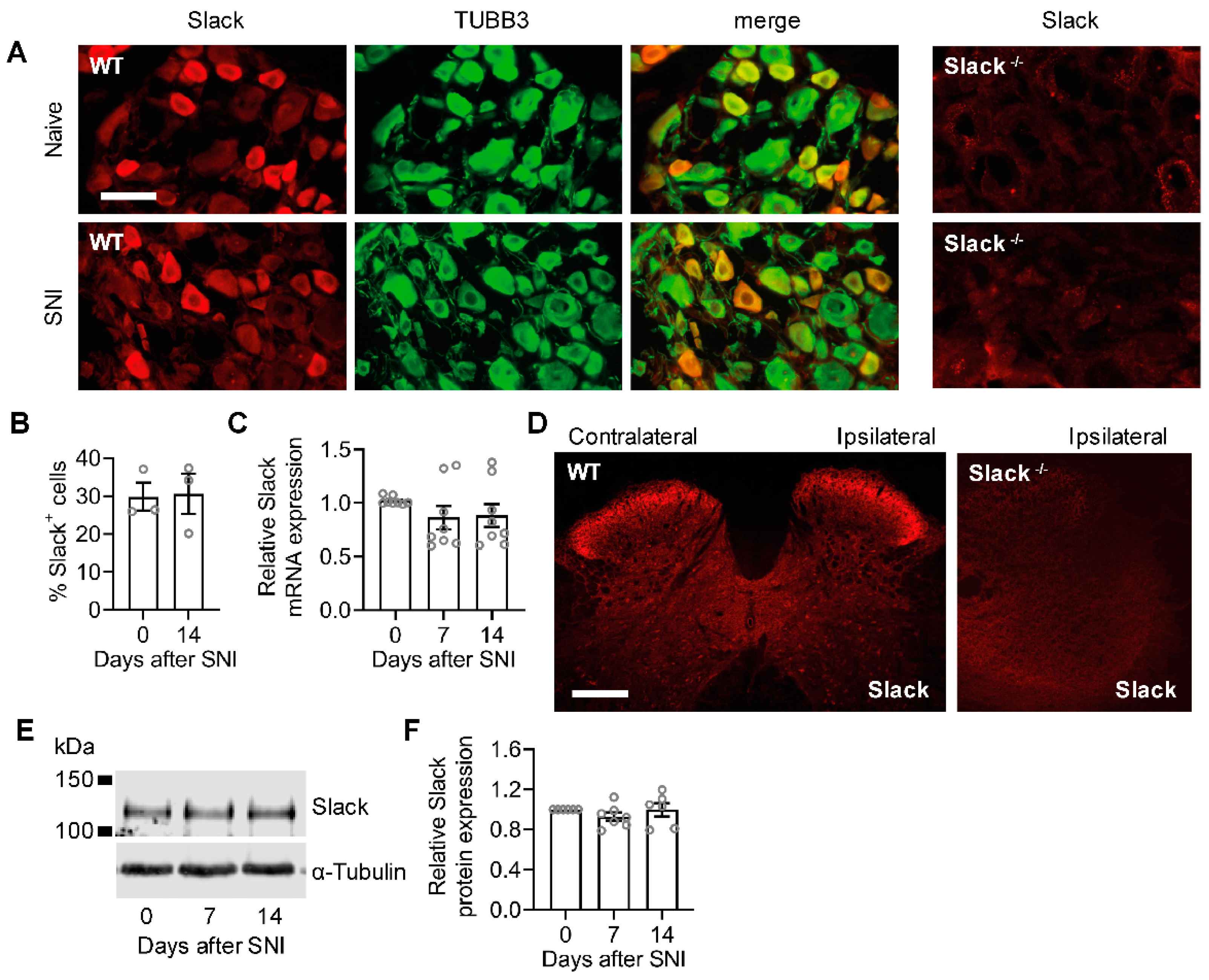
2.3. Unaltered Slack Expression in DRGs and the Spinal Cord after Spared Nerve Injury
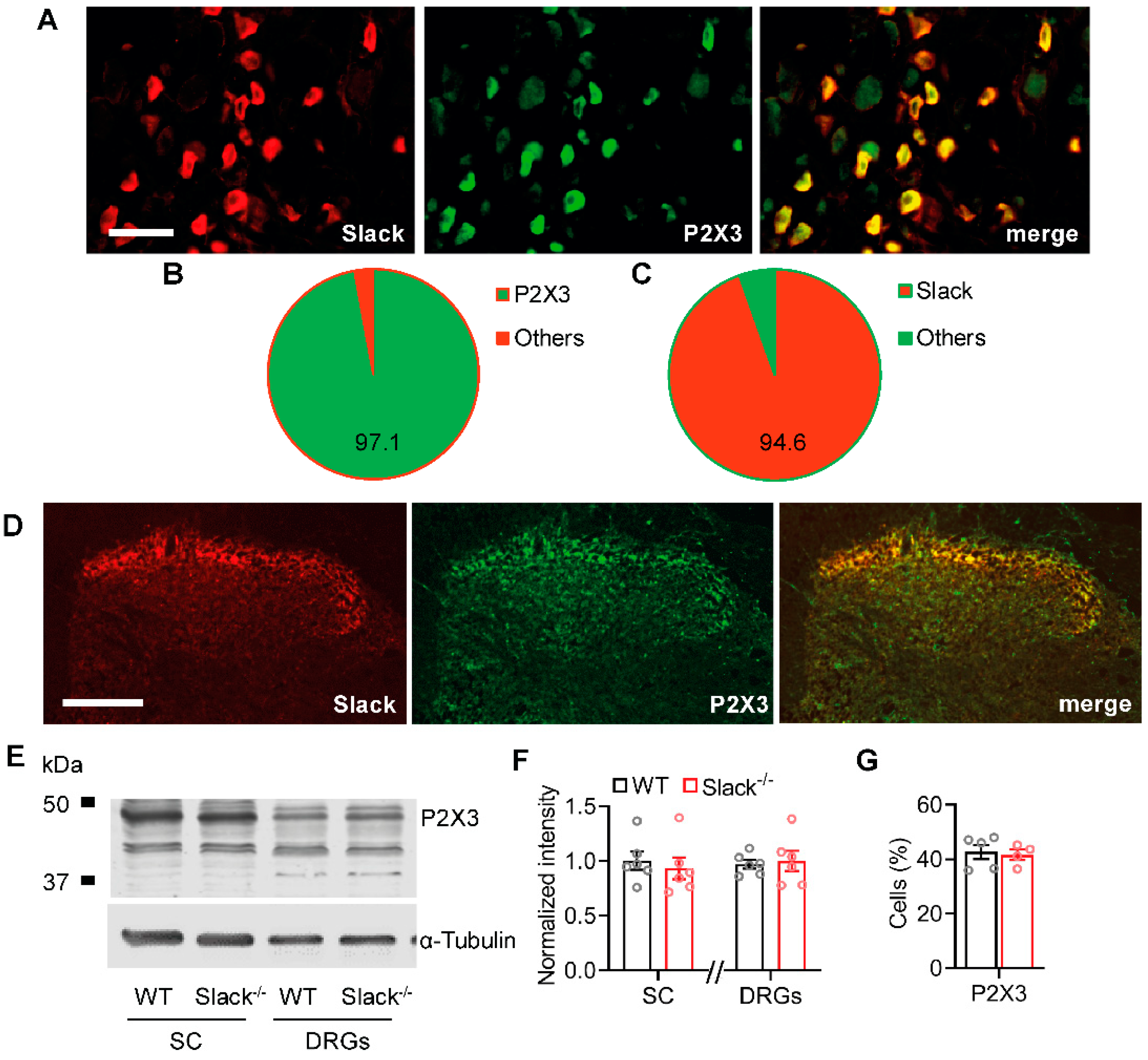
2.4. Slack Channels Colocalize with P2X3 Receptors in Sensory Neurons
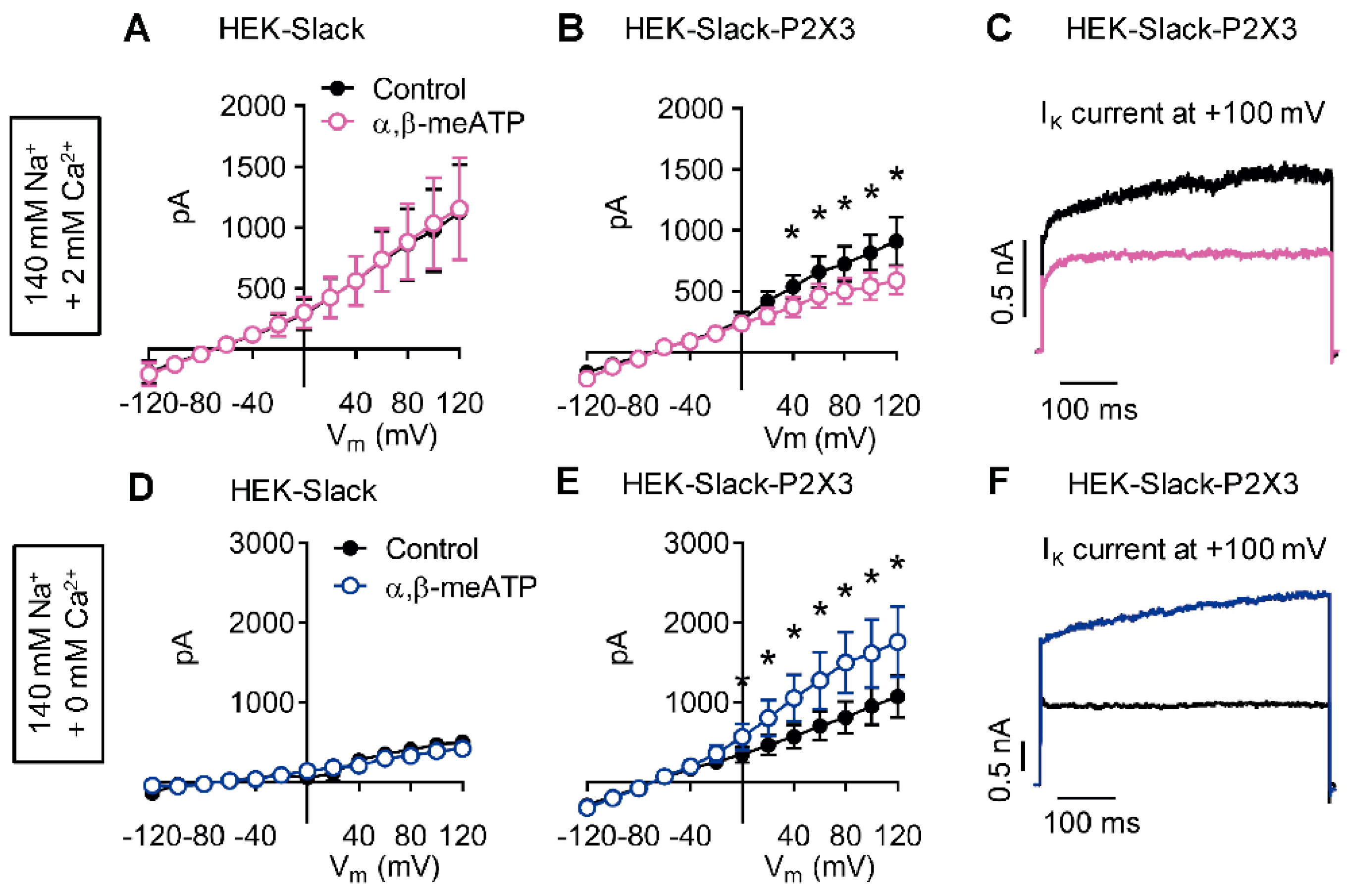
2.5. P2X3 Activation Modulates Slack-Mediated Potassium Currents In Vitro
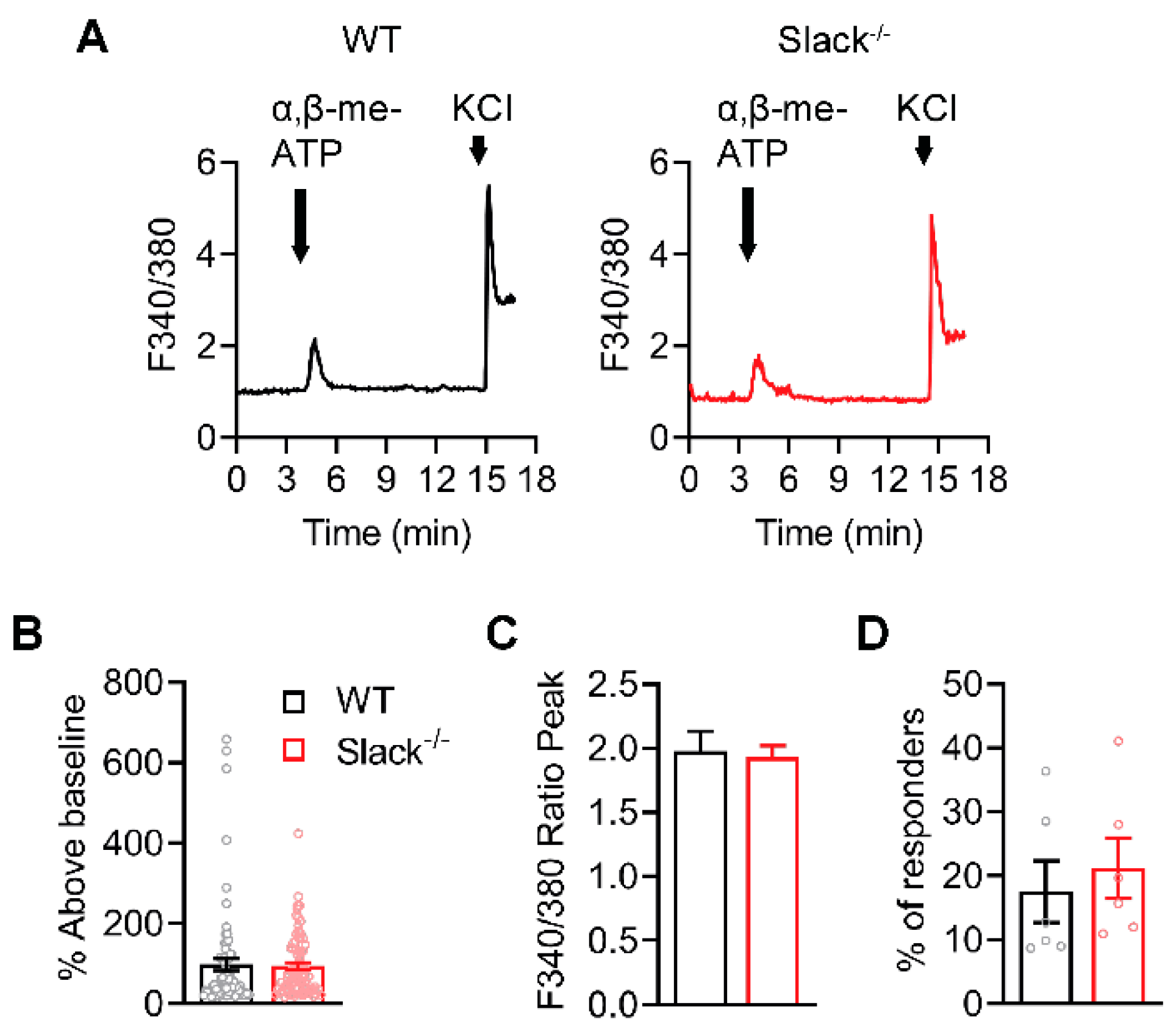
2.6. P2X3-Mediated Ca2+ Influx Is Unaltered in Sensory Neurons of Slack-/- Mice
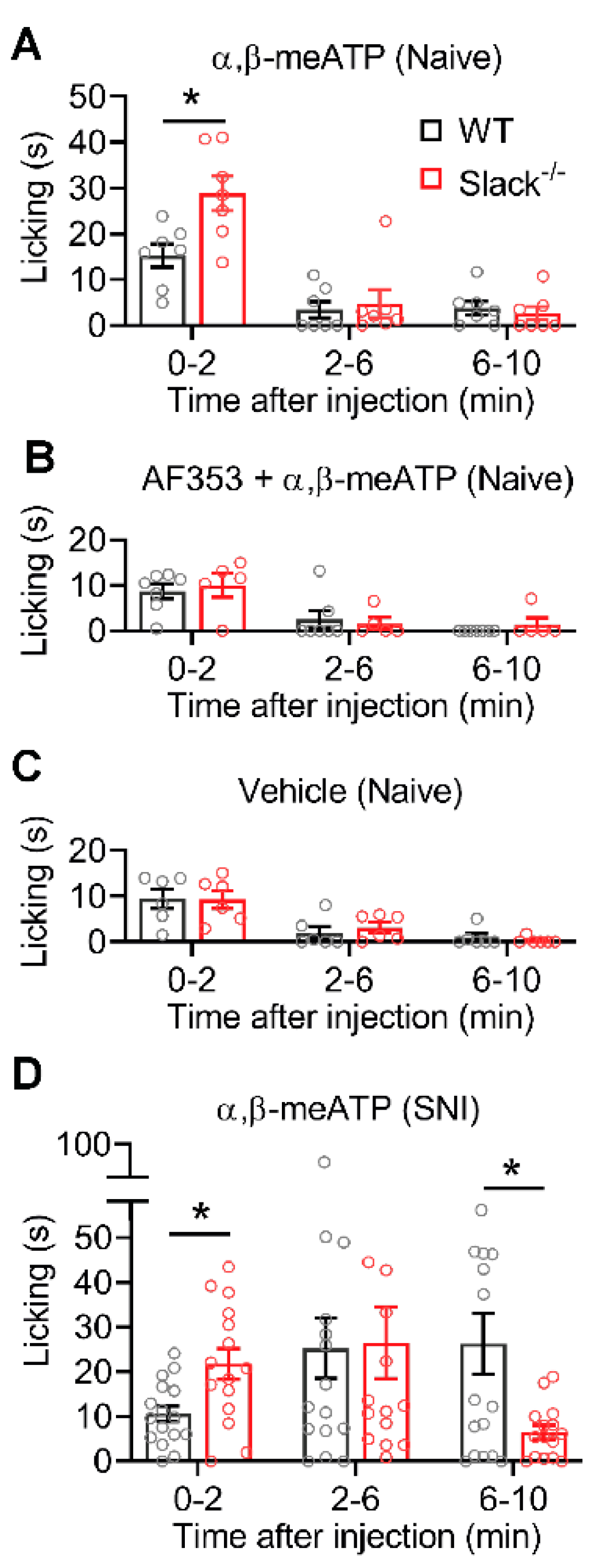
2.7. P2X3-Mediated Pain Behavior Is Altered in Slack-/- Mice
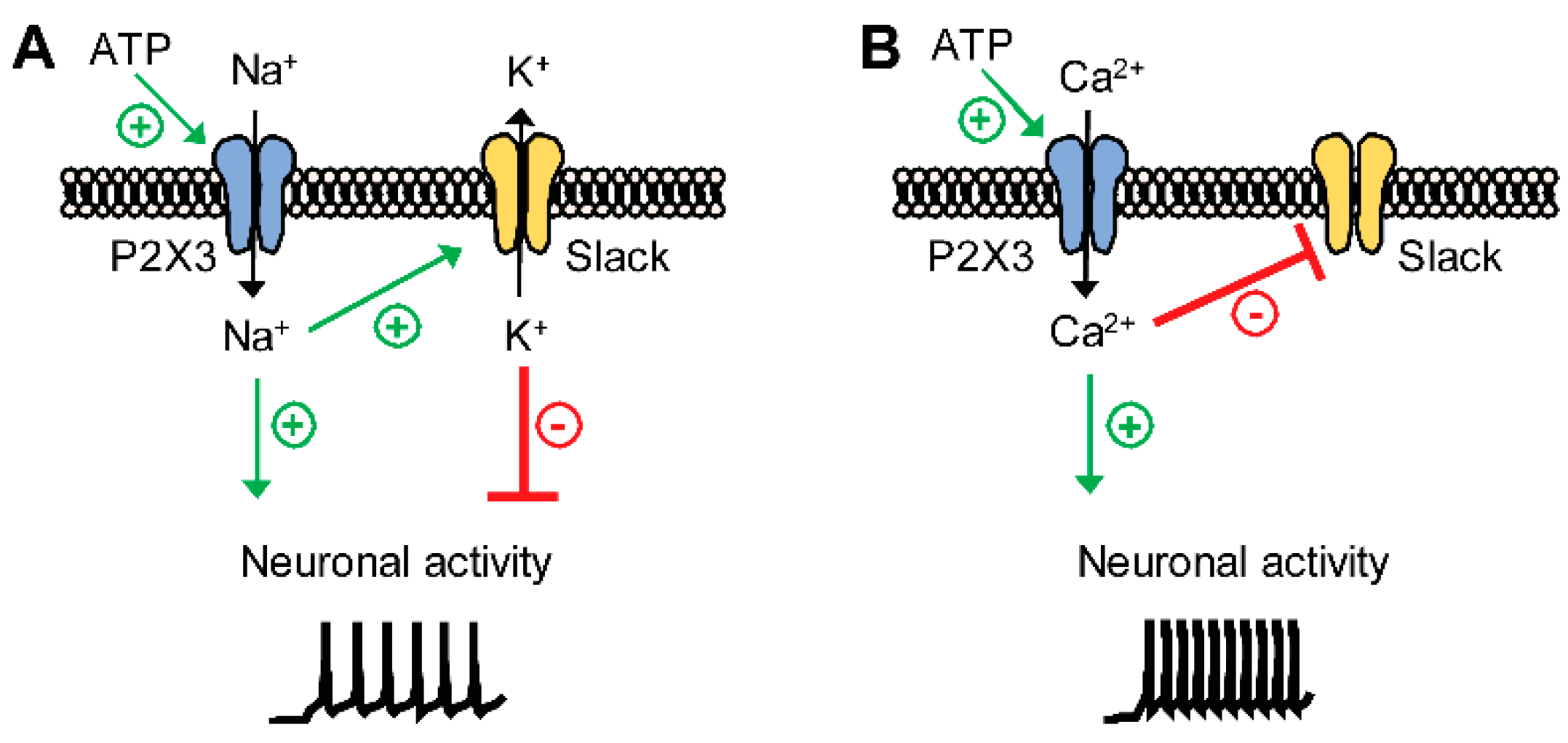
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Behavioral Testing
4.2.1. Spared Nerve Injury Model of Neuropathic Pain
4.2.2. α,β-meATP-Induced Paw Licking
4.3. Quantitative Real-Time Reverse Transcription PCR
4.4. Immunostaining
4.5. Western Blot
4.6. Patch Clamp Recordings
4.7. Calcium Imaging
4.8. Experimental Design and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| α,β-meATP | α,β-Methylene adenosine 5′-triphosphate |
| AF488 | Alexa Fluor 488 |
| CGRP | Calcitonin gene-related peptide |
| DRG | Dorsal root ganglion |
| GFP | Green fluorescent protein |
| IB4 | Griffonia simplicifolia isolectin B4 |
| IK | K+ current |
| IKNa | Na+-activated outward K+ current |
| I-V | Current-Voltage |
| NF200 | Neurofilament 200 |
| PBS | Phosphate-buffered saline |
| qRT-PCR | Quantitative real-time reverse transcription PCR |
| RIPA | Radioimmunoprecipitation assay |
| Slack | Sequence like a Ca2+-activated K+ channel |
| SNI | Spared nerve injury |
| TUBB3 | Tubulin β 3 |
| WT | Wild-type |
References
- van Hecke, O.; Austin, S.K.; Khan, R.A.; Smith, B.H.; Torrance, N. Neuropathic pain in the general population: A systematic review of epidemiological studies. Pain 2014, 155, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Finnerup, N.B.; Attal, N.; Haroutounian, S.; McNicol, E.; Baron, R.; Dworkin, R.H.; Gilron, I.; Haanpää, M.; Hansson, P.; Jensen, T.S.; et al. Pharmacotherapy for neuropathic pain in adults: A systematic review and meta-analysis. Lancet Neurol. 2015, 14, 162–173. [Google Scholar] [CrossRef] [Green Version]
- Tsantoulas, C.; Denk, F.; Signore, M.; Nassar, M.A.; Futai, K.; McMahon, S.B. Mice lacking Kcns1 in peripheral neurons show increased basal and neuropathic pain sensitivity. Pain 2018, 159, 1641–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.P.; Mao, J. Neuropathic pain: Mechanisms and their clinical implications. BMJ 2014, 348. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Mao, J. Update on neuropathic pain treatment: Ion channel blockers and gabapentinoids. Curr. Pain Headache Rep. 2013, 17. [Google Scholar] [CrossRef]
- Waxman, S.G.; Zamponi, G.W. Regulating excitability of peripheral afferents: Emerging ion channel targets. Nat. Neurosci. 2014, 17, 153–163. [Google Scholar] [CrossRef]
- Hite, R.K.; MacKinnon, R. Structural Titration of Slo2.2, a Na+-Dependent K+ Channel. Cell 2017, 168, 390–399.e11. [Google Scholar] [CrossRef] [Green Version]
- Bader, C.R.; Bernheim, L.; Bertrand, D. Sodium-activated potassium current in cultured avian neurones. Nature 1985, 317, 540–542. [Google Scholar] [CrossRef]
- Yuan, A.; Santi, C.M.; Wei, A.; Wang, Z.W.; Pollak, K.; Nonet, M.; Kaczmarek, L.; Crowder, C.M.; Salkoff, L. The sodium-activated potassium channel is encoded by a member of the Slo gene family. Neuron 2003, 37, 765–773. [Google Scholar] [CrossRef] [Green Version]
- Budelli, G.; Hage, T.A.; Wei, A.; Rojas, P.; Jong, Y.J.; O’Malley, K.; Salkoff, L. Na+-activated K+ channels express a large delayed outward current in neurons during normal physiology. Nat. Neurosci. 2009, 12, 745–750. [Google Scholar] [CrossRef] [Green Version]
- Budelli, G.; Sun, Q.; Ferreira, J.; Butler, A.; Santi, C.M.; Salkoff, L. SLO2 Channels Are Inhibited by All Divalent Cations That Activate SLO1 K+ Channels. J. Biol. Chem. 2016, 291, 7347–7356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamsett, T.J.; Picchione, K.E.; Bhattacharjee, A. NAD+ activates KNa channels in dorsal root ganglion neurons. J. Neurosci. 2009, 29, 5127–5134. [Google Scholar] [CrossRef] [PubMed]
- Nuwer, M.O.; Picchione, K.E.; Bhattacharjee, A. PKA-induced internalization of slack KNa channels produces dorsal root ganglion neuron hyperexcitability. J. Neurosci. 2010, 30, 14165–14172. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Wang, X.; Ostertag, E.M.; Nuwal, T.; Huang, B.; Jan, Y.N.; Basbaum, A.I.; Jan, L.Y. TMEM16C facilitates Na+-activated K+ currents in rat sensory neurons and regulates pain processing. Nat. Neurosci. 2013, 16, 1284–1290. [Google Scholar] [CrossRef]
- Lu, R.; Bausch, A.E.; Kallenborn-Gerhardt, W.; Stoetzer, C.; Debruin, N.; Ruth, P.; Geisslinger, G.; Leffler, A.; Lukowski, R.; Schmidtko, A. Slack channels expressed in sensory neurons control neuropathic pain in mice. J. Neurosci. 2015, 35, 1125–1135. [Google Scholar] [CrossRef]
- Evely, K.M.; Pryce, K.D.; Bausch, A.E.; Lukowski, R.; Ruth, P.; Haj-Dahmane, S.; Bhattacharjee, A. Slack KNa Channels Influence Dorsal Horn Synapses and Nociceptive Behavior. Mol. Pain 2017, 13. [Google Scholar] [CrossRef] [Green Version]
- Gururaj, S.; Evely, K.M.; Pryce, K.D.; Li, J.; Qu, J.; Bhattacharjee, A. Protein kinase A-induced internalization of Slack channels from the neuronal membrane occurs by adaptor protein-2/clathrin-mediated endocytosis. J. Biol. Chem. 2017, 292, 19304–19314. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Espinosa, P.L.; Wu, J.; Yang, C.; Gonzalez-Perez, V.; Zhou, H.; Liang, H.; Xia, X.M.; Lingle, C.J. Knockout of Slo2.2 enhances itch, abolishes KNa current, and increases action potential firing frequency in DRG neurons. eLife 2015, 4. [Google Scholar] [CrossRef] [Green Version]
- Schmiedl, S.; Peters, D.; Schmalz, O.; Mielke, A.; Rossmanith, T.; Diop, S.; Piefke, M.; Thurmann, P.; Schmidtko, A. Loxapine for Treatment of Patients With Refractory, Chemotherapy-Induced Neuropathic Pain: A Prematurely Terminated Pilot Study Showing Efficacy But Limited Tolerability. Front. Pharmacol. 2019, 10, 838. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic Mechanisms and Pain. Adv. Pharmacol. 2016, 75, 91–137. [Google Scholar]
- Krajewski, J.L. P2X3-Containing Receptors as Targets for the Treatment of Chronic Pain. Neurotherapeutics 2020, 17, 826–838. [Google Scholar] [CrossRef] [PubMed]
- Pryce, K.D.; Powell, R.; Agwa, D.; Evely, K.M.; Sheehan, G.D.; Nip, A.; Tomasello, D.L.; Gururaj, S.; Bhattacharjee, A. Magi-1 scaffolds NaV1.8 and Slack KNa channels in dorsal root ganglion neurons regulating excitability and pain. FASEB J. 2019, 33, 7315–7330. [Google Scholar] [CrossRef] [PubMed]
- Usoskin, D.; Furlan, A.; Islam, S.; Abdo, H.; Lonnerberg, P.; Lou, D.; Hjerling-Leffler, J.; Haeggstrom, J.; Kharchenko, O.; Kharchenko, P.V.; et al. Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat. Neurosci. 2015, 18, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Vulchanova, L.; Riedl, M.S.; Shuster, S.J.; Stone, L.S.; Hargreaves, K.M.; Buell, G.; Surprenant, A.; North, R.A.; Elde, R. P2X3 is expressed by DRG neurons that terminate in inner lamina II. Eur. J. Neurosci. 1998, 10, 3470–3478. [Google Scholar] [CrossRef] [PubMed]
- Fabbretti, E. ATP P2X3 receptors and neuronal sensitization. Front. Cell Neurosci. 2013, 7, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Li, G.W.; Wang, C.; Gu, Y.; Huang, L.Y. Mechanisms underlying enhanced P2X receptor-mediated responses in the neuropathic pain state. Pain 2005, 119, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.; Xiong, Y.; Yan, N.; Li, X.; Mao, Y.; Ni, X.; He, C.; LaMotte, R.H.; Burnstock, G.; Sun, J. Functional up-regulation of P2X3 receptors in the chronically compressed dorsal root ganglion. Pain 2008, 140, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Richards, D.; Gever, J.R.; Ford, A.P.; Fountain, S.J. Action of MK-7264 (gefapixant) at human P2X3 and P2X2/3 receptors and in vivo efficacy in models of sensitisation. Br. J. Pharmacol. 2019, 176, 2279–2291. [Google Scholar] [CrossRef] [Green Version]
- Hausmann, R.; Bodnar, M.; Woltersdorf, R.; Wang, H.; Fuchs, M.; Messemer, N.; Qin, Y.; Gunther, J.; Riedel, T.; Grohmann, M.; et al. ATP binding site mutagenesis reveals different subunit stoichiometry of functional P2X2/3 and P2X2/6 receptors. J. Biol. Chem. 2012, 287, 13930–13943. [Google Scholar] [CrossRef] [Green Version]
- Virginio, C.; North, R.A.; Surprenant, A. Calcium permeability and block at homomeric and heteromeric P2X2 and P2X3 receptors, and P2X receptors in rat nodose neurones. J. Physiol. 1998, 510, 27–35. [Google Scholar] [CrossRef]
- Simonetti, M.; Fabbro, A.; D’Arco, M.; Zweyer, M.; Nistri, A.; Giniatullin, R.; Fabbretti, E. Comparison of P2X and TRPV1 receptors in ganglia or primary culture of trigeminal neurons and their modulation by NGF or serotonin. Mol. Pain 2006, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grundy, L.; Daly, D.M.; Chapple, C.; Grundy, D.; Chess-Williams, R. TRPV1 enhances the afferent response to P2X receptor activation in the mouse urinary bladder. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaan, T.K.; Yip, P.K.; Patel, S.; Davies, M.; Marchand, F.; Cockayne, D.A.; Nunn, P.A.; Dickenson, A.H.; Ford, A.P.; Zhong, Y.; et al. Systemic blockade of P2X3 and P2X2/3 receptors attenuates bone cancer pain behaviour in rats. Brain 2010, 133, 2549–2564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gever, J.R.; Soto, R.; Henningsen, R.A.; Martin, R.S.; Hackos, D.H.; Panicker, S.; Rubas, W.; Oglesby, I.B.; Dillon, M.P.; Milla, M.E.; et al. AF-353, a novel, potent and orally bioavailable P2X3/P2X2/3 receptor antagonist. Br. J. Pharmacol. 2010, 160, 1387–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbretti, E. P2X3 receptors are transducers of sensory signals. Brain Res. Bull. 2019, 151, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, A.; Hochgerner, H.; Lonnerberg, P.; Johnsson, A.; Memic, F.; van der Zwan, J.; Haring, M.; Braun, E.; Borm, L.E.; La Manno, G.; et al. Molecular Architecture of the Mouse Nervous System. Cell 2018, 174, 999–1014.e22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsantoulas, C.; McMahon, S.B. Opening paths to novel analgesics: The role of potassium channels in chronic pain. Trends Neurosci. 2014, 37, 146–158. [Google Scholar] [CrossRef] [Green Version]
- Tsantoulas, C.; Zhu, L.; Shaifta, Y.; Grist, J.; Ward, J.P.; Raouf, R.; Michael, G.J.; McMahon, S.B. Sensory neuron downregulation of the Kv9.1 potassium channel subunit mediates neuropathic pain following nerve injury. J. Neurosci. 2012, 32, 17502–17513. [Google Scholar] [CrossRef] [Green Version]
- Liang, L.; Gu, X.; Zhao, J.Y.; Wu, S.; Miao, X.; Xiao, J.; Mo, K.; Zhang, J.; Lutz, B.M.; Bekker, A.; et al. G9a participates in nerve injury-induced Kcna2 downregulation in primary sensory neurons. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Calvo, M.; Richards, N.; Schmid, A.B.; Barroso, A.; Zhu, L.; Ivulic, D.; Zhu, N.; Anwandter, P.; Bhat, M.A.; Court, F.A.; et al. Altered potassium channel distribution and composition in myelinated axons suppresses hyperexcitability following injury. eLife 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Nanou, E.; Kyriakatos, A.; Bhattacharjee, A.; Kaczmarek, L.K.; Paratcha, G.; El Manira, A. Na+-mediated coupling between AMPA receptors and KNa channels shapes synaptic transmission. Proc. Natl. Acad. Sci. USA 2008, 105, 20941–20946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wildman, S.S.; Brown, S.G.; Rahman, M.; Noel, C.A.; Churchill, L.; Burnstock, G.; Unwin, R.J.; King, B.F. Sensitization by extracellular Ca2+ of rat P2X5 receptor and its pharmacological properties compared with rat P2X1. Mol. Pharmacol. 2002, 62, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Mo, G.; Bernier, L.P.; Zhao, Q.; Chabot-Dore, A.J.; Ase, A.R.; Logothetis, D.; Cao, C.Q.; Seguela, P. Subtype-specific regulation of P2X3 and P2X2/3 receptors by phosphoinositides in peripheral nociceptors. Mol. Pain 2009, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Rosenhouse-Dantsker, A.; Tang, Q.Y.; Noskov, S.; Logothetis, D.E. The RCK2 domain uses a coordination site present in Kir channels to confer sodium sensitivity to Slo2.2 channels. J. Neurosci. 2010, 30, 7554–7562. [Google Scholar] [CrossRef] [Green Version]
- Hullugundi, S.K.; Ferrari, M.D.; van den Maagdenberg, A.M.; Nistri, A. The mechanism of functional up-regulation of P2X3 receptors of trigeminal sensory neurons in a genetic mouse model of familial hemiplegic migraine type 1 (FHM-1). PLoS ONE 2013, 8, e60677. [Google Scholar] [CrossRef] [Green Version]
- Khakh, B.S.; Zhou, X.; Sydes, J.; Galligan, J.J.; Lester, H.A. State-dependent cross-inhibition between transmitter-gated cation channels. Nature 2000, 406, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Barajas-Lopez, C.; Montano, L.M.; Espinosa-Luna, R. Inhibitory interactions between 5-HT3 and P2X channels in submucosal neurons. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, G1238–G1248. [Google Scholar] [CrossRef] [Green Version]
- Boue-Grabot, E.; Emerit, M.B.; Toulme, E.; Seguela, P.; Garret, M. Cross-talk and co-trafficking between rho1/GABA receptors and ATP-gated channels. J. Biol. Chem. 2004, 279, 6967–6975. [Google Scholar] [CrossRef] [Green Version]
- Xia, R.; Mei, Z.Z.; Milligan, C.; Jiang, L.H. Inhibitory interaction between P2X4 and GABAC ρ1 receptors. Biochem. Biophys. Res. Commun. 2008, 375, 38–43. [Google Scholar] [CrossRef]
- Khakh, B.S.; Fisher, J.A.; Nashmi, R.; Bowser, D.N.; Lester, H.A. An angstrom scale interaction between plasma membrane ATP-gated P2X2 and α4β2 nicotinic channels measured with fluorescence resonance energy transfer and total internal reflection fluorescence microscopy. J. Neurosci. 2005, 25, 6911–6920. [Google Scholar] [CrossRef] [Green Version]
- Stephan, G.; Huang, L.; Tang, Y.; Vilotti, S.; Fabbretti, E.; Yu, Y.; Norenberg, W.; Franke, H.; Goloncser, F.; Sperlagh, B.; et al. Author Correction: The ASIC3/P2X3 cognate receptor is a pain-relevant and ligand-gated cationic channel. Nat. Commun. 2018, 9, 3351. [Google Scholar] [CrossRef] [PubMed]
- Vacca, F.; Amadio, S.; Sancesario, G.; Bernardi, G.; Volonte, C. P2X3 receptor localizes into lipid rafts in neuronal cells. J. Neurosci. Res. 2004, 76, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Vacca, F.; D’Ambrosi, N.; Nestola, V.; Amadio, S.; Giustizieri, M.; Cucchiaroni, M.L.; Tozzi, A.; Velluz, M.C.; Mercuri, N.B.; Volonte, C. N-Glycans mutations rule oligomeric assembly and functional expression of P2X3 receptor for extracellular ATP. Glycobiology 2011, 21, 634–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decosterd, I.; Woolf, C.J. Spared nerve injury: An animal model of persistent peripheral neuropathic pain. Pain 2000, 87, 149–158. [Google Scholar] [CrossRef]
- Bourquin, A.F.; Suveges, M.; Pertin, M.; Gilliard, N.; Sardy, S.; Davison, A.C.; Spahn, D.R.; Decosterd, I. Assessment and analysis of mechanical allodynia-like behavior induced by spared nerve injury (SNI) in the mouse. Pain 2006, 122, 14.e1–14.e14. [Google Scholar] [CrossRef] [Green Version]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Dixon, W.J. Efficient analysis of experimental observations. Annu. Rev. Pharmacol. Toxicol. 1980, 20, 441–462. [Google Scholar] [CrossRef]
- Christensen, S.L.; Hansen, R.B.; Storm, M.A.; Olesen, J.; Hansen, T.F.; Ossipov, M.; Izarzugaza, J.M.G.; Porreca, F.; Kristensen, D.M. Von Frey testing revisited: Provision of an online algorithm for improved accuracy of 50% thresholds. Eur. J. Pain. 2020, 24, 783–790. [Google Scholar] [CrossRef]
- Pitzer, C.; Kuner, R.; Tappe-Theodor, A. EXPRESS: Voluntary and evoked behavioral correlates in neuropathic pain states under different housing conditions. Mol. Pain 2016, 12. [Google Scholar] [CrossRef]
- Stark, A.; Schwenk, R.; Wack, G.; Zuchtriegel, G.; Hatemler, M.G.; Brautigam, J.; Schmidtko, A.; Reichel, C.A.; Bischoff, I.; Furst, R. Narciclasine exerts anti-inflammatory actions by blocking leukocyte-endothelial cell interactions and down-regulation of the endothelial TNF receptor 1. FASEB J. 2019, 33, 8771–8781. [Google Scholar] [CrossRef]
- Majuta, L.A.; Longo, G.; Fealk, M.N.; McCaffrey, G.; Mantyh, P.W. Orthopedic surgery and bone fracture pain are both significantly attenuated by sustained blockade of nerve growth factor. Pain 2015, 156, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, L.; Ma, C.; Liu, Q.; Weng, H.J.; Cui, Y.; Tang, Z.; Kim, Y.; Nie, H.; Qu, L.; Patel, K.N.; et al. A subpopulation of nociceptors specifically linked to itch. Nat. Neurosci. 2013, 16, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Lukowski, R.; Sausbier, M.; Zhang, D.D.; Sisignano, M.; Schuh, C.D.; Kuner, R.; Ruth, P.; Geisslinger, G.; Schmidtko, A. BKCa channels expressed in sensory neurons modulate inflammatory pain in mice. Pain 2014, 155, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.; Mergia, E.; Kennel, L.; Drees, O.; Steubing, R.D.; Real, C.I.; Kallenborn-Gerhardt, W.; Lu, R.; Friebe, A.; Koesling, D.; et al. Distinct functions of soluble guanylyl cyclase isoforms NO-GC1 and NO-GC2 in inflammatory and neuropathic pain processing. Pain 2018, 160, 607–618. [Google Scholar] [CrossRef]
- Kallenborn-Gerhardt, W.; Hohmann, S.W.; Syhr, K.M.; Schroder, K.; Sisignano, M.; Weigert, A.; Lorenz, J.E.; Lu, R.; Brune, B.; Brandes, R.P.; et al. Nox2-dependent signaling between macrophages and sensory neurons contributes to neuropathic pain hypersensitivity. Pain 2014, 155, 2161–2170. [Google Scholar] [CrossRef]
- Schmidtko, A.; Gao, W.; Sausbier, M.; Rauhmeier, I.; Sausbier, U.; Niederberger, E.; Scholich, K.; Huber, A.; Neuhuber, W.; Allescher, H.D.; et al. Cysteine-rich protein 2, a novel downstream effector of cGMP/cGMP-dependent protein kinase I-mediated persistent inflammatory pain. J. Neurosci. 2008, 28, 1320–1330. [Google Scholar] [CrossRef]
- Everill, B.; Kocsis, J.D. Nerve growth factor maintains potassium conductance after nerve injury in adult cutaneous afferent dorsal root ganglion neurons. Neuroscience 2000, 100, 417–422. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, R.; Metzner, K.; Zhou, F.; Flauaus, C.; Balzulat, A.; Engel, P.; Petersen, J.; Ehinger, R.; Bausch, A.; Ruth, P.; et al. Functional Coupling of Slack Channels and P2X3 Receptors Contributes to Neuropathic Pain Processing. Int. J. Mol. Sci. 2021, 22, 405. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010405
Lu R, Metzner K, Zhou F, Flauaus C, Balzulat A, Engel P, Petersen J, Ehinger R, Bausch A, Ruth P, et al. Functional Coupling of Slack Channels and P2X3 Receptors Contributes to Neuropathic Pain Processing. International Journal of Molecular Sciences. 2021; 22(1):405. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010405
Chicago/Turabian StyleLu, Ruirui, Katharina Metzner, Fangyuan Zhou, Cathrin Flauaus, Annika Balzulat, Patrick Engel, Jonas Petersen, Rebekka Ehinger, Anne Bausch, Peter Ruth, and et al. 2021. "Functional Coupling of Slack Channels and P2X3 Receptors Contributes to Neuropathic Pain Processing" International Journal of Molecular Sciences 22, no. 1: 405. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010405