Distinct Responses of Arabidopsis Telomeres and Transposable Elements to Zebularine Exposure

 and
and 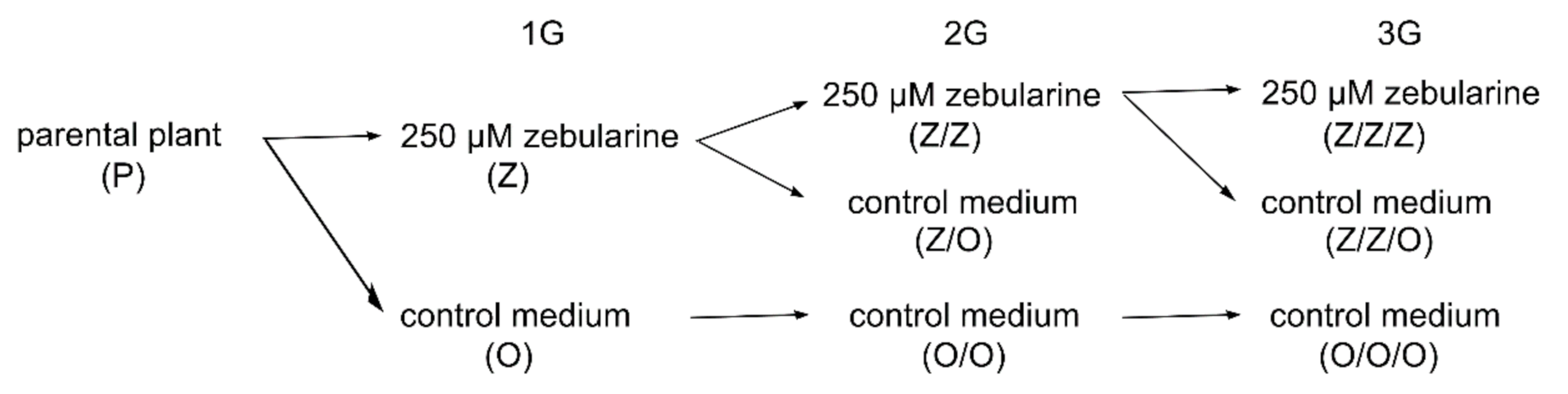{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
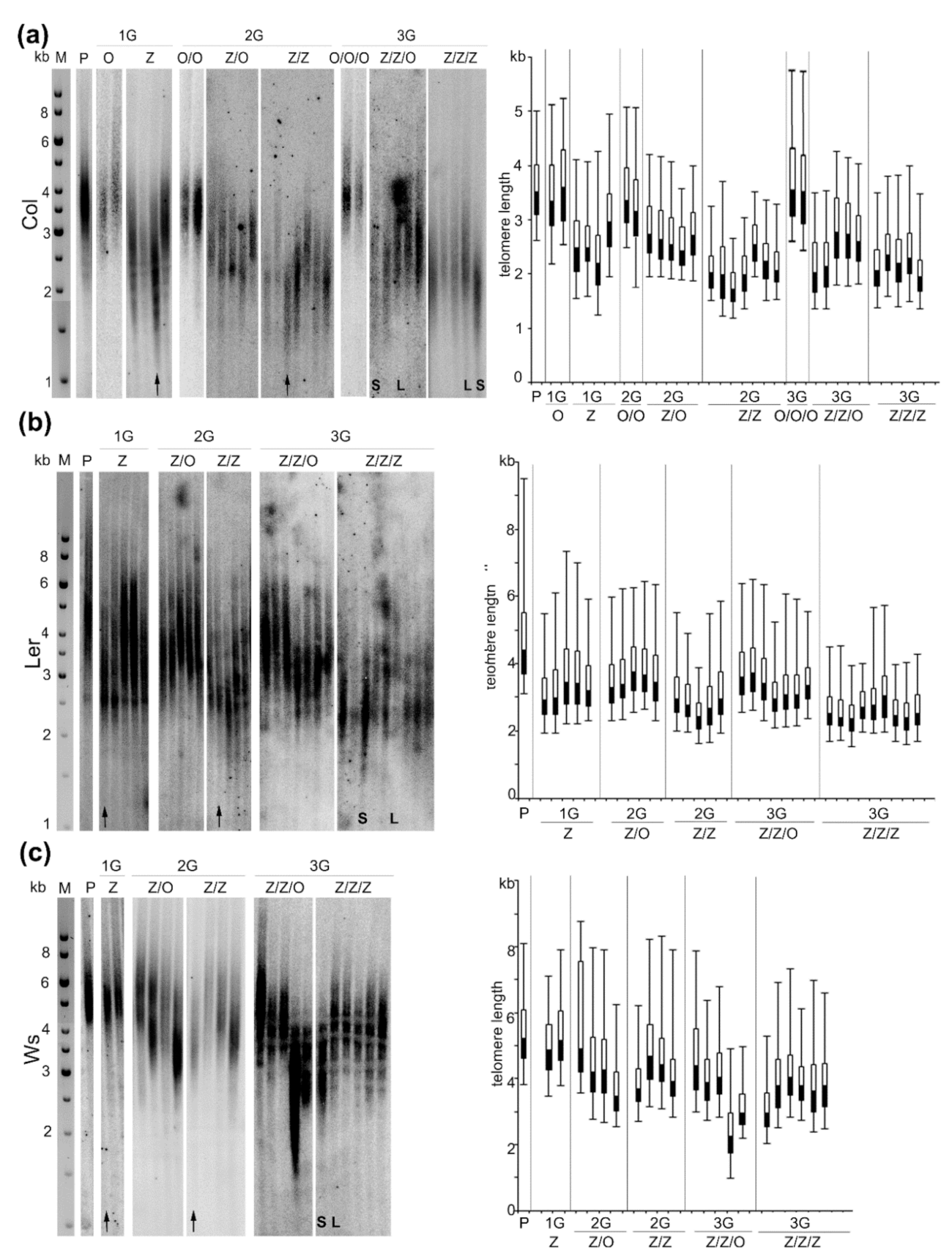
2.1. Zebularine-Induced Gradual Shortening of Telomeres in Arabidopsis Plants of Columbia-0 (Col), Wassilevskija (Ws), and Landsberg erecta (Ler) Ecotypes
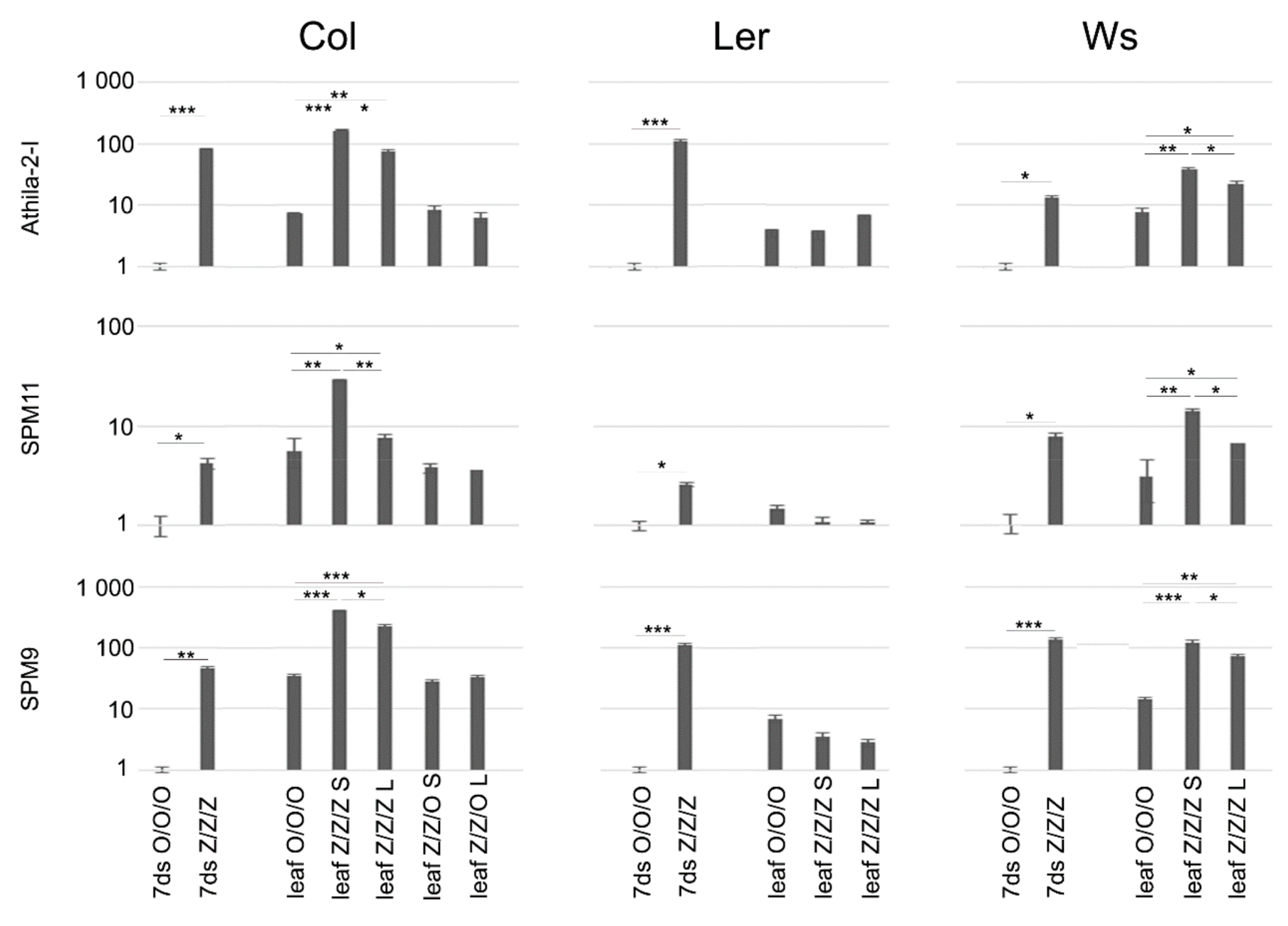
2.2. Extent of Changes in Telomere Lengths Correlated with Transcriptional Activation of Transposable Elements in Col and Ws Zebularine Plants
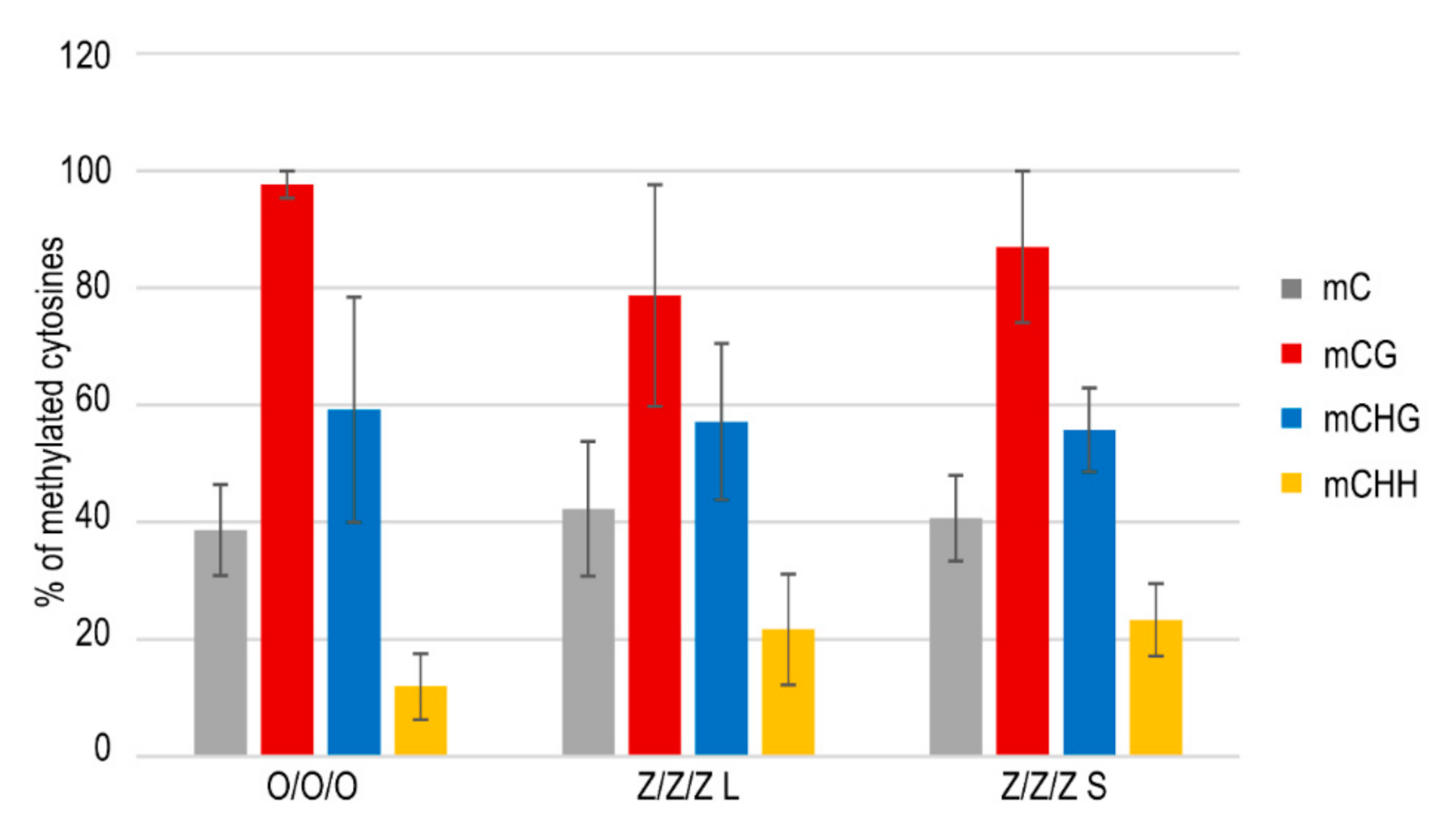
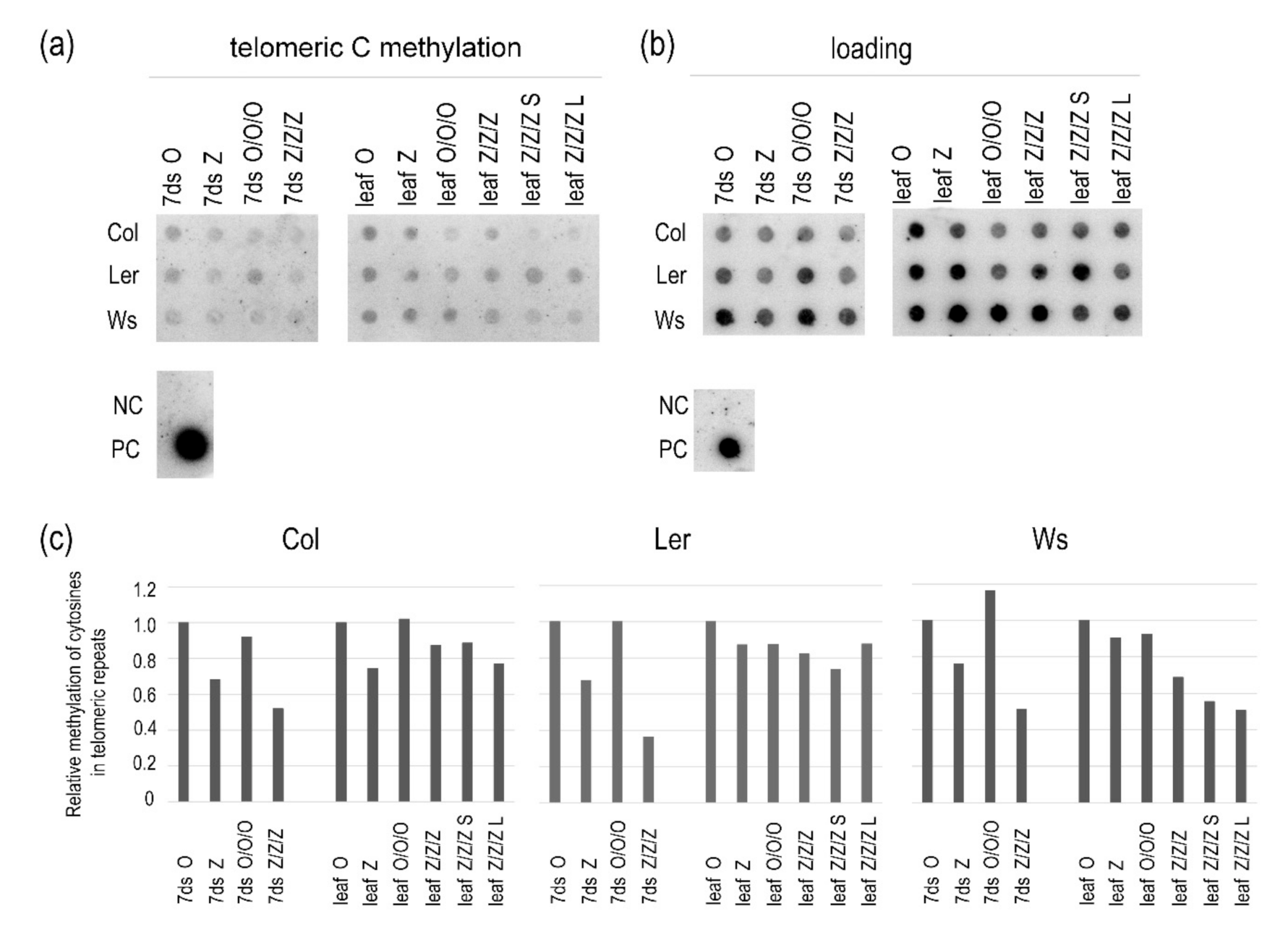
2.3. The Maintenance of the Short Telomere Phenotype and Transcriptional Activation of TEs in Leaves of Z/Z/Z Plants Are Not Tightly Related to the Loss of Methylated Cytosines in These Regions
3. Discussion
4. Materials and Methods
4.1. Cultivation of Plants
4.2. Analysis of Telomere Length by the Terminal Restriction Fragments (TRF) Method
4.3. Analysis of Transposable Elements Transcript Levels
4.4. Analysis of Methylation of TE Cytosines
4.5. Analysis of Methylation of Telomeric Cytosines
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HR | Homologous recombination |
| TE | Transposable Eeement |
| TRF | Terminal restriction fragment |
| Col | Columbia-0 |
| Ws | Wassilevskija |
| Ler | Landsberg erecta |
References
- Richards, E.J.; Ausubel, F.M. Isolation of a higher eukaryotic telomere from Arabidopsis thaliana. Cell 1988, 53, 127–136. [Google Scholar] [CrossRef]
- Moyzis, R.K.; Buckingham, J.M.; Cram, L.S.; Dani, M.; Deaven, L.L.; Jones, M.D.; Meyne, J.; Ratliff, R.L.; Wu, J.R. A highly conserved repetitive DNA sequence, (TTAGGG)n, present at the telomeres of human chromosomes. Proc. Natl. Acad. Sci. USA 1988, 85, 6622–6626. [Google Scholar] [CrossRef] [Green Version]
- Venkatesan, S.; Khaw, A.K.; Hande, M.P. Telomere biology-insights into an intriguing phenomenon. Cells 2017, 6, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, I.R.; Deleris, A.; Wong, W.; Zhong, X.H.; Chin, H.G.; Horwitz, G.A.; Kelly, K.A.; Pradhan, S.; Jacobsen, S.E. The de novo cytosine methyltransferase DRM2 requires intact UBA domains and a catalytically mutated paralog DRM3 during RNA-directed DNA methylation in Arabidopsis thaliana. PLoS Genet. 2010, 6, e1001182. [Google Scholar] [CrossRef] [Green Version]
- Cokus, S.J.; Feng, S.; Zhang, X.; Chen, Z.; Merriman, B.; Haudenschild, C.D.; Pradhan, S.; Nelson, S.F.; Pellegrini, M.; Jacobsen, S.E. Shotgun bisulphite sequencing of the Arabidopsis genome reveals DNA methylation patterning. Nature 2008, 452, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Ogrocka, A.; Polanska, P.; Majerova, E.; Janeba, Z.; Fajkus, J.; Fojtova, M. Compromised telomere maintenance in hypomethylated Arabidopsis thaliana plants. Nucleic Acids Res. 2014, 42, 2919–2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrbsky, J.; Akimcheva, S.; Watson, J.M.; Turner, T.L.; Daxinger, L.; Vyskot, B.; Aufsatz, W.; Riha, K. siRNA-mediated methylation of Arabidopsis telomeres. PLoS Genet. 2010, 6, e1000986. [Google Scholar] [CrossRef] [Green Version]
- Vaquero-Sedas, M.I.; Gamez-Arjona, F.M.; Vega-Palas, M.A. Arabidopsis thaliana telomeres exhibit euchromatic features. Nucleic Acids Res. 2011, 39, 2007–2017. [Google Scholar] [CrossRef] [Green Version]
- Vega-Vaquero, A.; Bonora, G.; Morselli, M.; Vaquero-Sedas, M.I.; Rubbi, L.; Pellegrini, M.; Vega-Palas, M.A. Novel features of telomere biology revealed by the absence of telomeric DNA methylation. Genome Res. 2016, 26, 1047–1056. [Google Scholar] [CrossRef] [Green Version]
- Underwood, C.J.; Henderson, I.R.; Martienssen, R.A. Genetic and epigenetic variation of transposable elements in Arabidopsis. Curr. Opin. Plant. Biol. 2017, 36, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Creasey, K.M.; Zhai, J.X.; Borges, F.; Van Ex, F.; Regulski, M.; Meyers, B.C.; Martienssen, R.A. miRNAs trigger widespread epigenetically activated siRNAs from transposons in Arabidopsis. Nature 2014, 508, 411–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, A.; Yonebayashi, S.; Watanabe, K.; Toyama, T.; Shimada, H.; Kakutani, T. Mobilization of transposons by a mutation abolishing full DNA methylation in Arabidopsis. Nature 2001, 411, 212–214. [Google Scholar] [CrossRef] [PubMed]
- Singer, T.; Yordan, C.; Martienssen, R.A. Robertson’s Mutator transposons in A. thaliana are regulated by the chromatin-remodeling gene Decrease in DNA Methylation (DDM1). Gene Dev. 2001, 15, 591–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stroud, H.; Hale, C.J.; Feng, S.H.; Caro, E.; Jacob, Y.; Michaels, S.D.; Jacobsen, S.E. DNA methyltransferases are required to induce heterochromatic re-replication in Arabidopsis. PLoS Genet. 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.C.; Matsen, C.B.; Gonzales, F.A.; Ye, W.; Greer, S.; Marquez, V.E.; Jones, P.A.; Selker, E.U. Inhibition of DNA methylation and reactivation of silenced genes by zebularine. J. Natl. Cancer Inst. 2003, 95, 399–409. [Google Scholar] [CrossRef]
- Champion, C.; Guianvarc’h, D.; Senamaud-Beaufort, C.; Jurkowska, R.Z.; Jeltsch, A.; Ponger, L.; Arimondo, P.B.; Guieysse-Peugeot, A.L. Mechanistic insights on the inhibition of C5 DNA methyltransferases by zebularine. PLoS ONE 2010, 5, e12388. [Google Scholar] [CrossRef] [Green Version]
- Ben-Kasus, T.; Ben-Zvi, Z.; Marquez, V.E.; Kelley, J.A.; Agbaria, R. Metabolic activation of zebularine, a novel DNA methylation inhibitor, in human bladder carcinoma cells. Biochem. Pharmacol. 2005, 70, 121–133. [Google Scholar] [CrossRef]
- Liu, C.H.; Finke, A.; Diaz, M.; Rozhon, W.; Poppenberger, B.; Baubec, T.; Pecinka, A. Repair of DNA damage induced by the cytidine analog zebularine requires ATR and ATM in Arabidopsis. Plant Cell 2015, 27, 1788–1800. [Google Scholar] [CrossRef] [Green Version]
- Baubec, T.; Pecinka, A.; Rozhon, W.; Mittelsten Scheid, O. Effective, homogeneous and transient interference with cytosine methylation in plant genomic DNA by zebularine. Plant J. 2009, 57, 542–554. [Google Scholar] [CrossRef] [Green Version]
- Majerova, E.; Fojtova, M.; Mozgova, I.; Bittova, M.; Fajkus, J. Hypomethylating drugs efficiently decrease cytosine methylation in telomeric DNA and activate telomerase without affecting telomere lengths in tobacco cells. Plant Mol. Biol. 2011, 77, 371–380. [Google Scholar] [CrossRef]
- Griffin, P.T.; Niederhuth, C.E.; Schmitz, R.J. A comparative analysis of 5-azacytidine- and zebularine-induced DNA demethylation. G3 Genes Genomes Genet. 2016, 6, 2773–2780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakirov, E.V.; Shippen, D.E. Length regulation and dynamics of individual telomere tracts in wild-type Arabidopsis. Plant Cell 2004, 16, 1959–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzke, M.A.; Mette, M.F.; Aufsatz, W.; Jakowitsch, J.; Matzke, A.J.M. Host defenses to parasitic sequences and the evolution of epigenetic control mechanisms. Genetica 1999, 107, 271–287. [Google Scholar] [CrossRef] [PubMed]
- Yoder, J.A.; Walsh, C.P.; Bestor, T.H. Cytosine methylation and the ecology of intragenomic parasites. Trends Genet. 1997, 13, 335–340. [Google Scholar] [CrossRef]
- Kato, M.; Miura, A.; Bender, J.; Jacobsen, S.E.; Kakutani, T. Role of CG and non-CG methylation in immobilization of transposons in Arabidopsis. Curr. Biol. 2003, 13, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Baubec, T.; Finke, A.; Scheid, O.M.; Pecinka, A. Meristem-specific expression of epigenetic regulators safeguards transposon silencing in Arabidopsis. EMBO Rep. 2014, 15, 446–452. [Google Scholar] [CrossRef] [Green Version]
- Hetzl, J.; Foerster, A.M.; Raidl, G.; Scheid, O.M. CyMATE: A new tool for methylation analysis of plant genornic DNA after bisulphite sequencing. Plant J. 2007, 51, 526–536. [Google Scholar] [CrossRef]
- Sovakova, P.P.; Magdolenova, A.; Konecna, K.; Rajecka, V.; Fajkus, J.; Fojtova, M. Telomere elongation upon transfer to callus culture reflects the reprogramming of telomere stability control in Arabidopsis. Plant Mol. Biol. 2018, 98, 81–99. [Google Scholar] [CrossRef]
- Xie, X.Y.; Shippen, D.E. DDM1 guards against telomere truncation in Arabidopsis. Plant Cell Rep. 2018, 37, 501–513. [Google Scholar] [CrossRef] [Green Version]
- Anteková, K. Analysis of Levels of Transposable Elements Transcripts in Arabidopsis Plants Exposed to the Zebularine during Germination; Unpublished Results; Masaryk University: Brno, Czech Republic, 2018. [Google Scholar]
- Suzuki, M.M.; Bird, A. DNA methylation landscapes: Provocative insights from epigenomics. Nat. Rev. Genet. 2008, 9, 465–476. [Google Scholar] [CrossRef]
- Polanská, P. Telomeres and Telomerase in Plants with Hypomethylated Genomes. Diploma Thesis, Masaryk University, Brno, Czech Republic, 2013. [Google Scholar]
- Vaquero-Sedas, M.I.; Vega-Palas, M.A. Determination of Arabidopsis thaliana telomere length by PCR. Sci. Rep. 2014, 4, 5540. [Google Scholar] [CrossRef] [PubMed]
- Fojtová, M.; Fajkus, P.; Polanská, P.; Fajkus, J. Terminal restriction fragments (TRF) method to analyze telomere lengths. Bio Protoc. 2015, 5, e1671. [Google Scholar] [CrossRef]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Adamusova, K.; Khosravi, S.; Fujimoto, S.; Houben, A.; Matsunaga, S.; Fajkus, J.; Fojtova, M. Two combinatorial patterns of telomere histone marks in plants with canonical and non-canonical telomere repeats. Plant J. 2020, 102, 678–687. [Google Scholar] [CrossRef] [PubMed]
- Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. Improved telomere detection using a telomere repeat probe (TTAGGG)n Generated by PCR. Nucleic Acids Res. 1991, 19, 4780. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. Quantification Strategies in Real-Time PCR. In A-Z of Quantitative PCR; Bustin, S.A., Ed.; International University Line: La Jola, CA, USA, 2004; pp. 87–112. [Google Scholar]
- Clark, S.J.; Harrison, J.; Paul, C.L.; Frommer, M. High-sensitivity mapping of methylated cytosines. Nucleic Acids Res. 1994, 22, 2990–2997. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konečná, K.; Sováková, P.P.; Anteková, K.; Fajkus, J.; Fojtová, M. Distinct Responses of Arabidopsis Telomeres and Transposable Elements to Zebularine Exposure. Int. J. Mol. Sci. 2021, 22, 468. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010468
Konečná K, Sováková PP, Anteková K, Fajkus J, Fojtová M. Distinct Responses of Arabidopsis Telomeres and Transposable Elements to Zebularine Exposure. International Journal of Molecular Sciences. 2021; 22(1):468. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010468
Chicago/Turabian StyleKonečná, Klára, Pavla Polanská Sováková, Karin Anteková, Jiří Fajkus, and Miloslava Fojtová. 2021. "Distinct Responses of Arabidopsis Telomeres and Transposable Elements to Zebularine Exposure" International Journal of Molecular Sciences 22, no. 1: 468. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010468