
Rutin Modulates MAPK Pathway Differently from Quercetin in Angiotensin II-Induced H9c2 Cardiomyocyte Hypertrophy



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
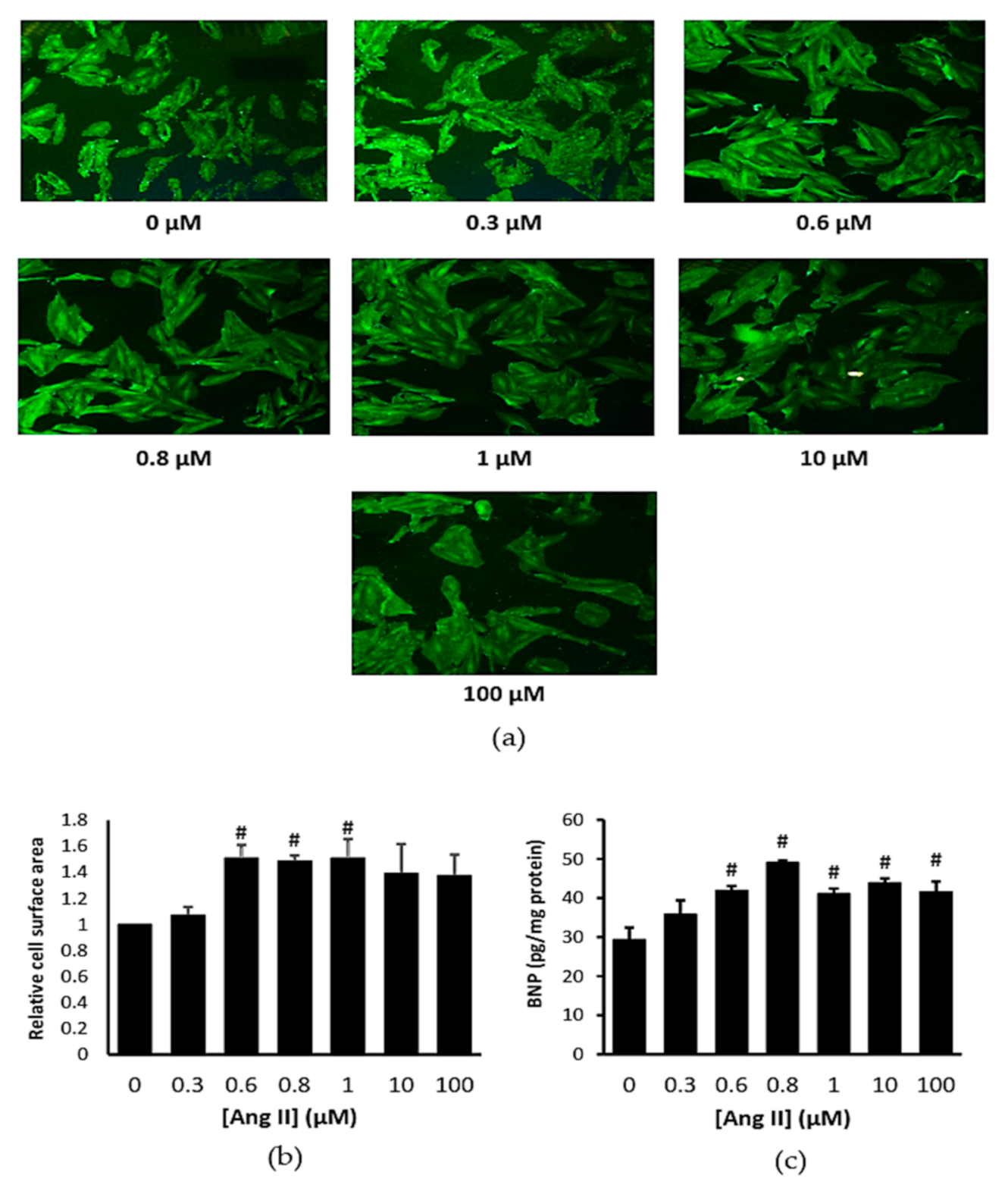
2.1. Ang II Increased Cardiomyocytes Cell Surface Area and B-Type Natriuretic Peptide Level
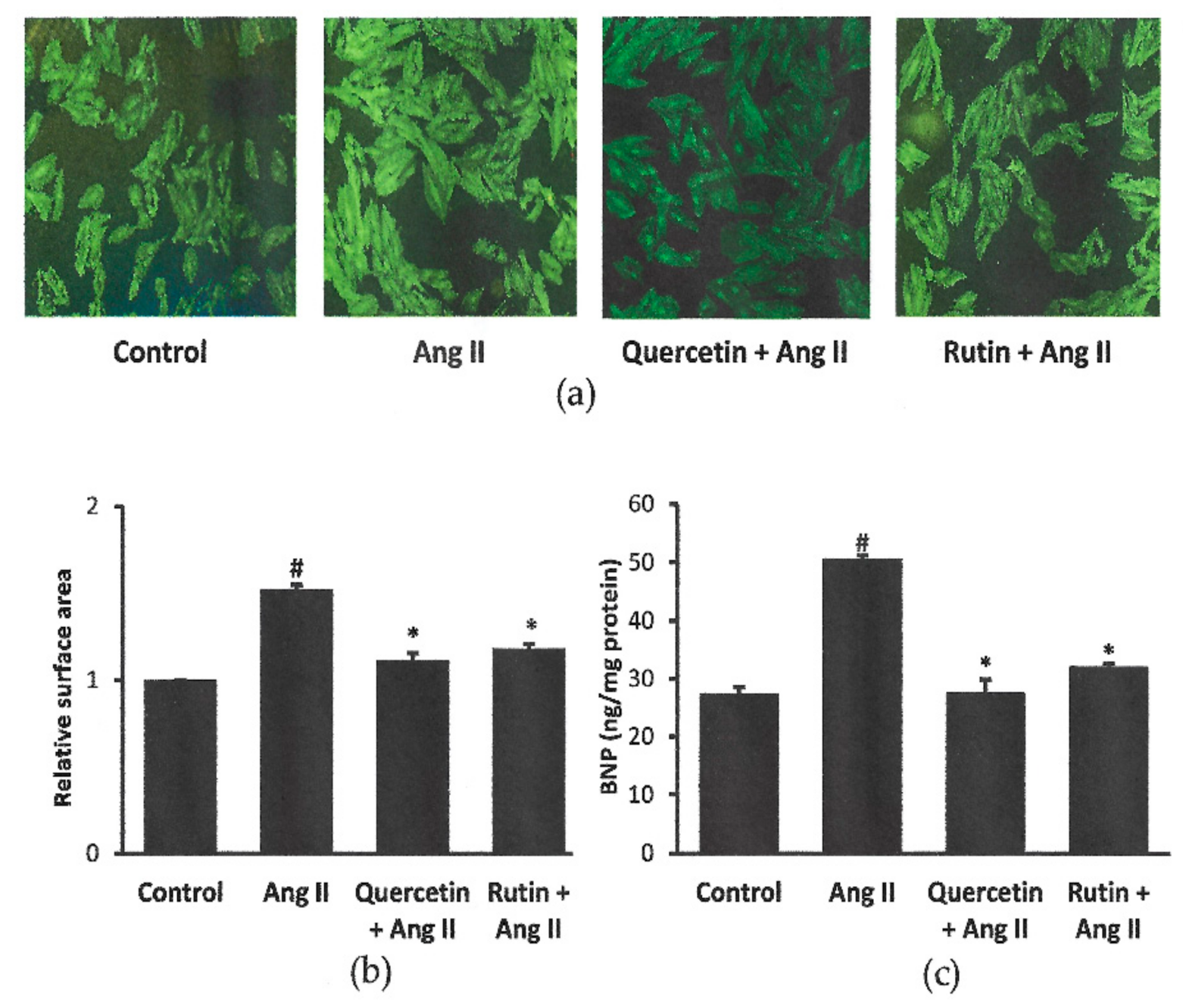
2.2. Rutin and Quercetin Inhibited Ang II-Induced Cardiomyocyte Hypertrophy and BNP Level
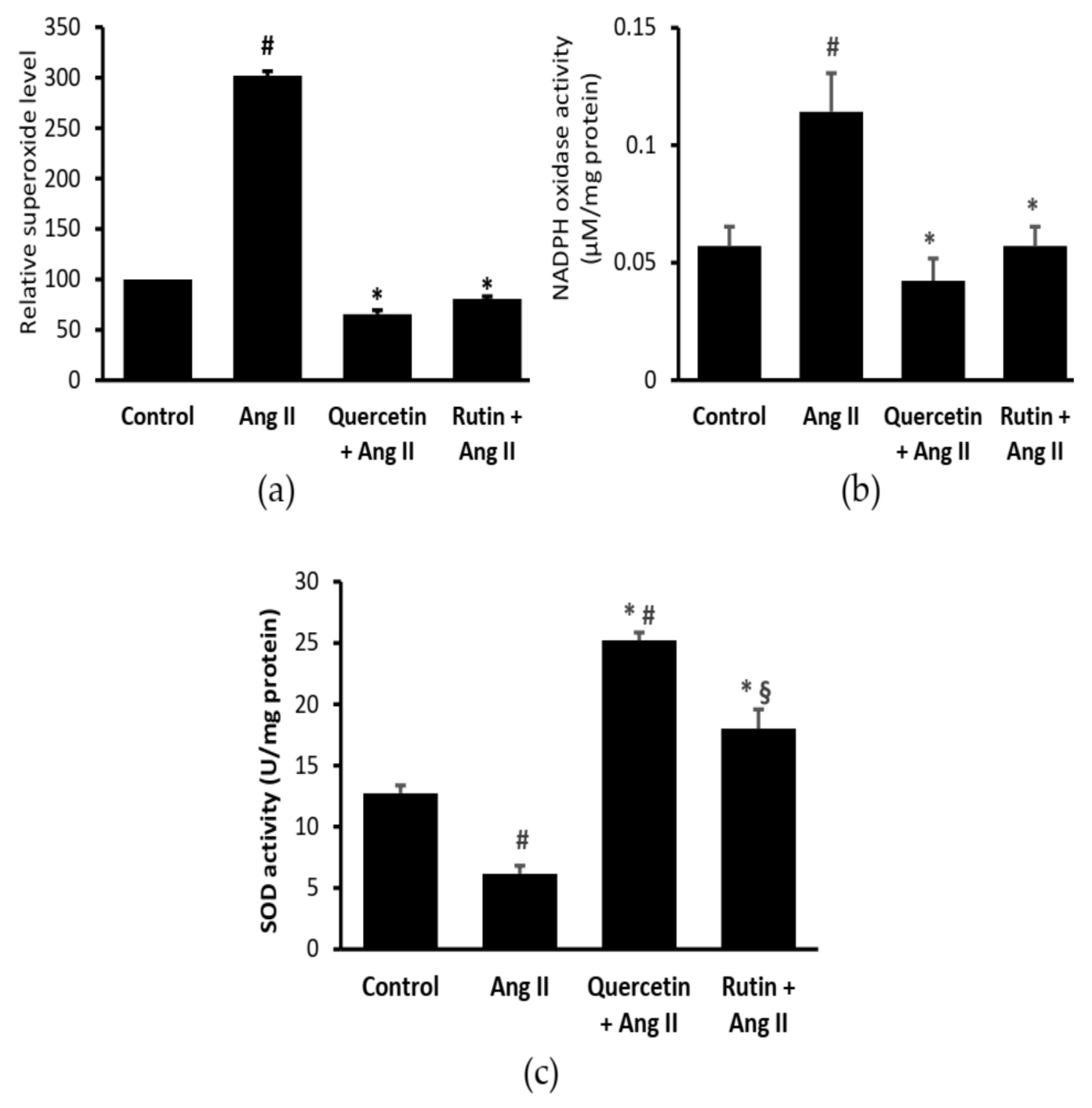
2.3. Rutin and Quercetin Inhibited Ang II-Induced Intracellular Superoxide, NADPH Oxidase, and SOD Activities
2.4. Quercetin and Rutin Reduced Cellular iNOS and NO Levels
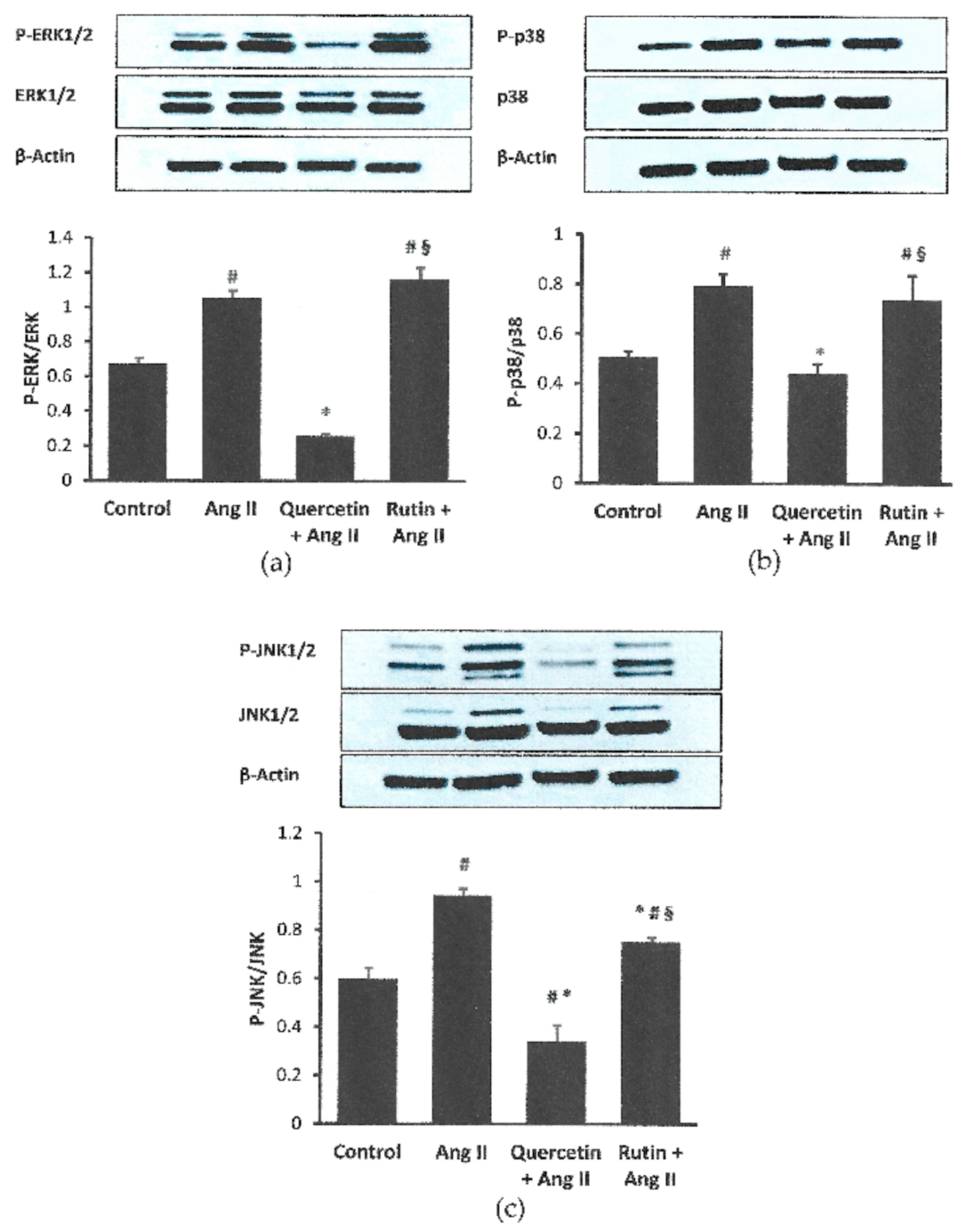
2.5. Quercetin and Rutin Affects MAPK Activation Differently
3. Discussion
4. Materials and Methods
4.1. Culture of Myocardial H9c2 Cell Line
4.2. Concentration-Response Studies of Ang II on Cardiomyocytes
4.3. Experimental Groups
4.4. Cell Size Measurement
4.5. Cellular BNP Level and iNOS Activity
4.6. Intracellular Superoxide Detection
4.7. NADPH Oxidase and SOD Activities
4.8. Cellular Nitrite
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shimizu, I.; Minamino, T. Physiological and pathological cardiac hypertrophy. J. Mol. Cell Cardiol. 2016, 97, 245–262. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Ma, B.; Han, X. The role of autophagy in angiotensin II-induced pathological cardiac hypertrophy. J. Mol. Endocrinol. 2016, 57, R143–R152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallo, S.; Vitacolonna, A.; Bonzano, A.; Comoglio, P.; Crepaldi, T. ERK: A key player in the pathophysiology of cardiac hypertrophy. Int. J. Mol. Sci. 2019, 20, 2164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, E.Y.; Kim, T.Y.; Hong, G.U.; Kang, H.; Lee, J.Y.; Park, J.Y.; Kim., S.C.; Kim, Y.H.; Chung, M.H.; Kwon, Y.I.; et al. Inhibitory effects of roseoside and icariside E4 isolated from a natural product mixture (No-ap) on the expression of angiotensin II receptor 1 and oxidative stress in angiotensin II-stimulated H9C2 cells. Molecules 2019, 24, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.L.; Li, Y.; Yin, P.P.; Kong, F.J.; Guo, J.J.; Shi, H.T.; Zhu, J.B.; Zou, Y.Z.; Ge, J.B. Hypertrophied myocardium is vulnerable to ischemia/reperfusion injury and refractory to rapamycin-induced protection due to increased oxidative/nitrative stress. Clin. Sci. 2018, 132, 93–110. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Rekep, M.; Wei, W.; Wu, Q.; Xue, Q.; Li, S.; Tian, J.; Yi, Q.; Zhang, G.; Zhang, G.; et al. Quercetin prevents in vivo and in vitro myocardial hypertrophy through the proteasome-GSK-3 pathway. Cardiovasc. Drugs Ther. 2018, 32, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Siti, H.N.; Jalil, J.; Asmadi, A.Y.; Kamisah, Y. Effects of quercetin on cardiac function in pressure overload and postischemic cardiac injury in rodents: A systematic review and meta-analysis. Cardiovasc. Drugs Ther. 2020. [Google Scholar] [CrossRef]
- Vázquez-Flores, L.F.; Casas-Grajales, S.; Herández-Aquino, E.; Vargas-Pozada, E.E.; Muriel, P. Antioxidant, antiinflammatory, and antifibrotic properties of quercetin in the liver. In Liver Pathophysiology: Therapies and Antioxidants; Muriel, P., Ed.; Academic Press: San Diego, CA, USA, 2017; pp. 653–674. [Google Scholar]
- Mbaveng, A.T.; Zhao, Q.; Kuete, V. Harmful and protective effects of phenolic compounds from African medicinal plants. In Toxicological Survey of African Medicinal Plants; Kuete, V., Ed.; Elsevier: London, UK, 2014; pp. 577–609. [Google Scholar]
- Singh, H.; Kaur, P.; Kaur, P.; Muthuraman, A.; Singh, G.; Kaur, M. Investigation of therapeutic potential and molecular mechanism of vitamin P and digoxin in I/R-induced myocardial infarction in rat. Naunyn Schmiedebergs Arch. Pharmacol. 2015, 388, 565–574. [Google Scholar] [CrossRef]
- Gui, J.; Mustafa, N.; Jalil, J.; Jubri, Z.; Kamisah, Y. Modulation of NOX4 and MAPK signalling pathways by Parkia speciosa empty pods in H9c2 cardiomyocytes exposed to H2O2. Indian J. Pharm. Sci. 2019, 81, 1029–1035. [Google Scholar] [CrossRef]
- Sghaier, B.M.; Pagano, A.; Mousslim, M.; Ammari, Y.; Kovacic, H.; Luis, J. Rutin inhibits proliferation, attenuates superoxide production and decreases adhesion and migration of human cancerous cells. Biomed. Pharmacother. 2016, 84, 1972–1978. [Google Scholar] [CrossRef]
- Mustafa, N.H.; Ugusman, A.; Jalil, J.; Kamisah, Y. Anti-inflammatory property of Parkia speciosa empty pod extract in human umbilical vein endothelial cells. J. Appl. Pharm. Sci. 2018, 8, 152–158. [Google Scholar]
- Gul, A.; Kunwar, B.; Mazhar, M.; Faizi, S.; Ahmed, D.; Shah, M.R.; Simjee, S.U. Rutin and rutin-conjugated gold nanoparticles ameliorate collagen-induced arthritis in rats through inhibition of NF-κB and iNOS activation. Int. Immunopharmacol. 2018, 59, 310–317. [Google Scholar] [CrossRef]
- Ditano-Vázquez, P.; Torres-Peña, J.D.; Galeano-Valle, F.; Pérez-Caballero, A.I.; Demelo-Rodríguez, P.; Lopez-Miranda, J.; Katsiki, N.; Delgado-Lista, J.; Alvarez-Sala-Walther, L.A. The fluid aspect of the Mediterranean diet in the prevention and management of cardiovascular disease and diabetes: The Role of polyphenol content in moderate consumption of wine and olive oil. Nutrients 2019, 11, 2833. [Google Scholar] [CrossRef] [Green Version]
- Ezzati, M.; Yousefi, B.; Velaei, K.; Safa, A. A review on anti-cancer properties of quercetin in breast cancer. Life Sci. 2020, 248, 117463. [Google Scholar] [CrossRef]
- Tang, S.M.; Deng, X.T.; Zhou, J.; Li, Q.P.; Ge, X.X.; Miao, L. Pharmacological basis and new insights of quercetin action in respect to its anti-cancer effects. Biomed. Pharmacother. 2020, 121, 109604. [Google Scholar] [CrossRef]
- Khan, F.; Pandey, P.; Upadhyay, T.K.; Jafri, A.; Jha, N.K.; Mishra, R.; Singh, V. Anti-cancerous effect of rutin against HPV-C33A cervical cancer cells via G0/G1 cell cycle arrest and apoptotic induction. Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Caparica, R.; Júlio, A.; Araújo, M.E.M.; Baby, A.R.; Fonte, P.; Costa, J.G.; Santos de Almeida, T. Anticancer activity of rutin and its combination with ionic liquids on renal cells. Biomolecules 2020, 10, 233. [Google Scholar] [CrossRef] [Green Version]
- Derosa, G.; Maffioli, P.; D’Angelo, A.; Di Pierro, F. A role for quercetin in coronavirus disease 2019 (COVID-19). Phytother. Res. 2021, 35, 1230–1236. [Google Scholar] [CrossRef]
- Kumari, A.; Rajput, V.S.; Nagpal, P.; Kukrety, H.; Grover, S.; Grover, A. Dual inhibition of SARS-CoV-2 spike and main protease through a repurposed drug, rutin. J. Biomol. Struct. Dyn. 2020, 2, 1–13. [Google Scholar]
- Huynh, T.; Wang, H.; Luan, B. Structure-based lead optimization of herbal medicine rutin for inhibiting SARS-CoV-2’s main protease. Phys. Chem. Chem. Phys. 2020, 22, 25335–25343. [Google Scholar] [CrossRef]
- Colunga Biancatelli, R.M.L.; Berrill, M.; Catravas, J.D.; Marik, P.E. Quercetin and vitamin C: An experimental, synergistic therapy for the prevention and treatment of SARS-CoV-2 related disease (COVID-19). Front. Immunol. 2020, 11, 1451. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Liu, R.; Lin, N.; Luo, H.; Tang, J.; Huang, Q.; Sun, H.; Tang, L. NADPH oxidase hyperactivity contributes to cardiac dysfunction and apoptosis in rats with severe experimental pancreatitis through ROS-mediated MAPK signaling pathway. Oxid. Med. Cell. Longev. 2019, 2019, 4578175. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, T.M.; Monteiro, M.M.; Braga, V.A. Angiotensin-II-derived reactive oxygen species on baroreflex sensitivity during hypertension: New perspectives. Front. Physiol. 2013, 4, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cos, P.; Ying, L.; Calomme, M.; Hu, J.P.; Cimanga, K.; Van Poel, B.; Pieters, L.; Vlietinck, A.J.; Berghe, D.V. Structure-activity relationship and classification of flavonoids as inhibitors of xanthine oxidase and superoxide scavengers. J. Nat. Prod. 1998, 61, 71–76. [Google Scholar] [CrossRef]
- Zhuang, X.; Zhao, B.; Liu, S.; Song, F.; Cui, F.; Liu, Z.; Li, Y. Noncovalent interactions between superoxide dismutase and flavonoids studied by native mass spectrometry combined with molecular simulations. Anal. Chem. 2016, 88, 11720–11726. [Google Scholar] [CrossRef]
- Viskupicova, J.; Strosova, M.K.; Zizkova, P.; Majekova, M.; Horakova, L. Rutin stimulates sarcoplasmic reticulum Ca (2+)-ATPase activity (SERCA1) and protects SERCA1 from peroxynitrite mediated injury. Mol. Cell. Biochem. 2015, 402, 51–62. [Google Scholar] [CrossRef]
- Yelins’ka, A.M.; Liashenko, L.I.; Kostenko, V.O. Quercetin potentiates antiradical properties of epigallocatechin-3-gallate in periodontium of rats under systemic and local administration of lipopolisaccharide of Salmonella typhi. Wiad. Lek. 2019, 72, 1499–1503. [Google Scholar] [CrossRef]
- Restini, C.B.A.; Garcia, A.F.E.; Natalin, H.M.; Natalin, G.M.; Rizzi, E. Signaling pathways of cardiac remodeling related to angiotensin II. In Renin-Angiotensin System: Past, Present and Future; Tolekova, A.N., Ed.; InTech Open: London, UK, 2017; p. 51. [Google Scholar]
- Radi, R. Oxygen radicals, nitric oxide, and peroxynitrite: Redox pathways in molecular medicine. Proc. Natl. Acad. Sci. USA 2018, 115, 5839–5848. [Google Scholar] [CrossRef] [Green Version]
- Gui, J.S.; Jalil, J.; Kamisah, Y. Parkia speciosa empty pod extract exerts anti-inflammatory properties by modulating NFκB and MAPK pathways in cardiomyocytes exposed to tumor necrosis factor-α. Cytotechnology 2019, 71, 79–89. [Google Scholar] [CrossRef]
- Jones, H.S.; Gordon, A.; Magwenzi, S.G.; Naseem, K.; Atkin, S.L.; Courts, F.L. The dietary flavonol quercetin ameliorates angiotensin II-induced redox signaling imbalance in a human umbilical vein endothelial cell model of endothelial dysfunction via ablation of p47phox expression. Mol. Nutr. Food Res. 2016, 60, 787–797. [Google Scholar] [CrossRef]
- Qin, T.C.; Chen, L.; Yu, L.X.; Gu, Z.L. Inhibitory effect of quercetin on cultured neonatal rat cardiomyocytes hypertrophy induced by angiotensin. Acta Pharmacol. Sin. 2001, 22, 1103–1106. [Google Scholar]
- Wang, L.; Tan, A.; An, X.; Xia, Y.; Xie, Y. Quercetin dihydrate inhibition of cardiac fibrosis induced by angiotensin II in vivo and in vitro. Biomed. Pharmacother. 2020, 127, 110205. [Google Scholar] [CrossRef]
- Yan, L.; Zhang, J.D.; Wang, B.; Lv, Y.J.; Jiang, H.; Liu, G.L.; Qiao, Y.; Ren, M.; Guo, X.F. Quercetin inhibits left ventricular hypertrophy in spontaneously hypertensive rats and inhibits angiotensin II-induced H9C2 cells hypertrophy by enhancing PPAR-γ expression and suppressing AP-1 activity. PLoS ONE 2013, 8, e7254. [Google Scholar] [CrossRef] [Green Version]
- Peter, A.K.; Bjerke, M.A.; Leinwand, L.A. Biology of the cardiac myocyte in heart disease. Mol. Biol. Cell 2016, 27, 2149–2160. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Sun, B.; Tong, Q.; Ren, L. Rutin protects against pirarubicin-induced cardiotoxicity through TGF-β1-p38 MAPK signaling pathway. Evid. Based Complement. Altern. Med. 2017, 2017, 1759385. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-mediated cellular signaling. Oxid. Med. Cell. Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angeloni, C.; Hrelia, S. Quercetin reduces inflammatory responses in LPS-stimulated cardiomyoblasts. Oxid. Med. Cell. Longev. 2012, 2012, 837104. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Cheong, Y.K.; Kim, N.H.; Chung, H.T.; Kang, D.G.; Pae, H.O. Mitogen-activated protein kinases and reactive oxygen species: How can ROS activate MAPK pathways? J. Signal Transduct. 2011, 2011, 792639. [Google Scholar] [CrossRef]
- Sriramula, S.; Francis, J. Tumor necrosis factor-alpha is essential for angiotensin II-induced ventricular remodeling: Role for oxidative stress. PLoS ONE 2015, 10, e0138372. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Behm, D.J.; Nerurkar, S.S.; Ao, Z.; Bentley, R.; Mirabile, R.C.; Johns, D.G.; Woods, T.N.; Doe, C.P.; Coatney, R.W.; et al. Effects of p38 MAPK Inhibitor on angiotensin II-dependent hypertension, organ damage, and superoxide anion production. J. Cardiovasc. Pharmacol. 2007, 49, 362–368. [Google Scholar] [CrossRef]
- Yokota, T.; Wang, Y. p38 MAP kinases in the heart. Gene 2016, 575, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Elimban, V.; Nijjar, M.S.; Gupta, S.K.; Dhalla, N.S. Role of mitogen-activated protein kinase in cardiac hypertrophy and heart failure. Exp. Clin. Cardiol. 2003, 8, 173–183. [Google Scholar]
- Plotnikov, A.; Zehorai, E.; Procaccia, S.; Seger, R. The MAPK cascades: Signaling components, nuclear roles and mechanisms of nuclear translocation. Biochim. Biophys. Acta 2011, 1813, 1619–1633. [Google Scholar] [CrossRef] [Green Version]
- Majalahti, T.; Suo-Palosaari, M.; Sármán, B.; Hautala, N.; Pikkarainen, S.; Tokola, H.; Vuolteenaho, O.; Wang, J.; Paradis, P.; Nemer, M.; et al. Cardiac BNP gene activation by angiotensin II in vivo. Mol. Cell. Endocrinol. 2007, 273, 59–67. [Google Scholar] [CrossRef]
- Nayer, J.; Aggarwal, P.; Galwankar, S. Utility of point-of-care testing of natriuretic peptides (brain natriuretic peptide and n-terminal pro-brain natriuretic peptide) in the emergency department. Int. J. Crit. Illn. Inj. Sci. 2014, 4, 209–215. [Google Scholar]
- Yancy, C.W.; Jessup, M.; Bozkurt, B.; Butler, J.; Casey, D.E., Jr.; Colvin, M.M.; Drazner, M.H.; Filippatos, G.S.; Fonarow, G.C.; Givertz, M.M.; et al. 2017 ACC/AHA/HFSA focused update of the 2013 ACCF/AHA guideline for the management of heart failure: A report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines and the Heart Failure Society of America. J. Am. Coll. Cardiol. 2017, 70, 776–803. [Google Scholar]
- Chu, J.X.; Li, G.M.; Gao, X.J.; Wang, J.X.; Han, S.Y. Buckwheat rutin inhibits AngII-induced cardiomyocyte hypertrophy via blockade of CaN-dependent signal pathway. Iran. J. Pharm. Res. 2014, 13, 1347–1355. [Google Scholar]
- Li, M.; Jiang, Y.; Jing, W.; Sun, B.; Miao, C.; Ren, L. Quercetin provides greater cardioprotective effect than its glycoside derivative rutin on isoproterenol-induced cardiac fibrosis in the rat. Can. J. Physiol. Pharmacol. 2013, 91, 951–959. [Google Scholar] [CrossRef]
- Mladenka, P.; Zatloukalová, L.; Simůnek, T.; Bobrovová, Z.; Semecký, V.; Nachtigal, P.; Hasková, P.; Macková, E.; Vávrová, J.; Holecková, M.; et al. Direct administration of rutin does not protect against catecholamine cardiotoxicity. Toxicology 2009, 255, 25–32. [Google Scholar] [CrossRef]
- Siti, H.N.; Jalil, J.; Asmadi, A.Y.; Kamisah, Y. Roles of rutin in cardiac remodeling. J. Funct. Foods 2020, 64, 103606. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, Y.; Lighthouse, J.K.; Mickelsen, D.M.; Wu, J.; Yao, P.; Small, E.M.; Yan, C. A novel role of cyclic nucleotide phosphodiesterase 10A in pathological cardiac remodeling and dysfunction. Circulation 2020, 141, 217–233. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.L.; Huang, H.L.; Chien, H.C.; Chen, C.M.; Lin, C.N.; Ko, W.C. Inhibitory effects of quercetin derivatives on phosphodiesterase isozymes and high-affinity [(3) H]-rolipram binding in guinea pig tissues. Investig. New Drugs 2008, 26, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Orhan, I.E.; Ertas, A.; Temel, H.; Hadda, T.B.; Saleem, M.; Raza, M.; Khan, H. Elucidation of phosphodiesterase-1 inhibitory effect of some selected natural polyphenolics using in vitro and in silico methods. Curr. Top. Med. Chem. 2017, 17, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Prathapan, A.; Vineetha, V.P.; Abhilash, P.A.; Raghu, K.G. Boerhaavia diffusa L. attenuates angiotensin II-induced hypertrophy in H9c2 cardiac myoblast cells via modulating oxidative stress and down-regulating NF-κβ and transforming growth factor β1. Br. J. Nutr. 2013, 110, 1201–1210. [Google Scholar] [CrossRef] [Green Version]
- Jeong, M.H.; Kim, S.J.; Kang, H.; Park, K.W.; Park, W.J.; Yang, S.Y.; Yang, D.K. Cucurbitacin I attenuates cardiomyocyte hypertrophy via inhibition of connective tissue growth factor (CCN2) and TGF-β/Smads signalings. PLoS ONE 2015, 10, e0136236. [Google Scholar] [CrossRef] [Green Version]
- Mustapha, N.M.; Tarr, J.M.; Kohner, E.M.; Chibber, R. NADPH oxidase versus mitochondria-derived ROS in glucose-induced apoptosis of pericytes in early diabetic retinopathy. J. Ophthamol. 2010, 2010, 746978. [Google Scholar]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Siti, H.N.; Kamisah, Y.; Mohamed, S.; Jaarin, K. Effects of citrus leaf extract on aortic vascular reactivity in hypertensive rats fed repeatedly heated vegetable oil. Appl. Physiol. Nutr. Metab. 2019, 44, 373–380. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siti, H.N.; Jalil, J.; Asmadi, A.Y.; Kamisah, Y. Rutin Modulates MAPK Pathway Differently from Quercetin in Angiotensin II-Induced H9c2 Cardiomyocyte Hypertrophy. Int. J. Mol. Sci. 2021, 22, 5063. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105063
Siti HN, Jalil J, Asmadi AY, Kamisah Y. Rutin Modulates MAPK Pathway Differently from Quercetin in Angiotensin II-Induced H9c2 Cardiomyocyte Hypertrophy. International Journal of Molecular Sciences. 2021; 22(10):5063. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105063
Chicago/Turabian StyleSiti, Hawa Nordin, Juriyati Jalil, Ahmad Yusof Asmadi, and Yusof Kamisah. 2021. "Rutin Modulates MAPK Pathway Differently from Quercetin in Angiotensin II-Induced H9c2 Cardiomyocyte Hypertrophy" International Journal of Molecular Sciences 22, no. 10: 5063. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105063