
Plasma Metabolomics Analysis of Polyvinyl Chloride Workers Identifies Altered Processes and Candidate Biomarkers for Hepatic Hemangiosarcoma and Its Development

 and
and
Abstract
:1. Introduction
2. Results
2.1. Demographics, Chemical Exposure and Clinical Chemistries of Hemangiosarcoma and Control Cohorts
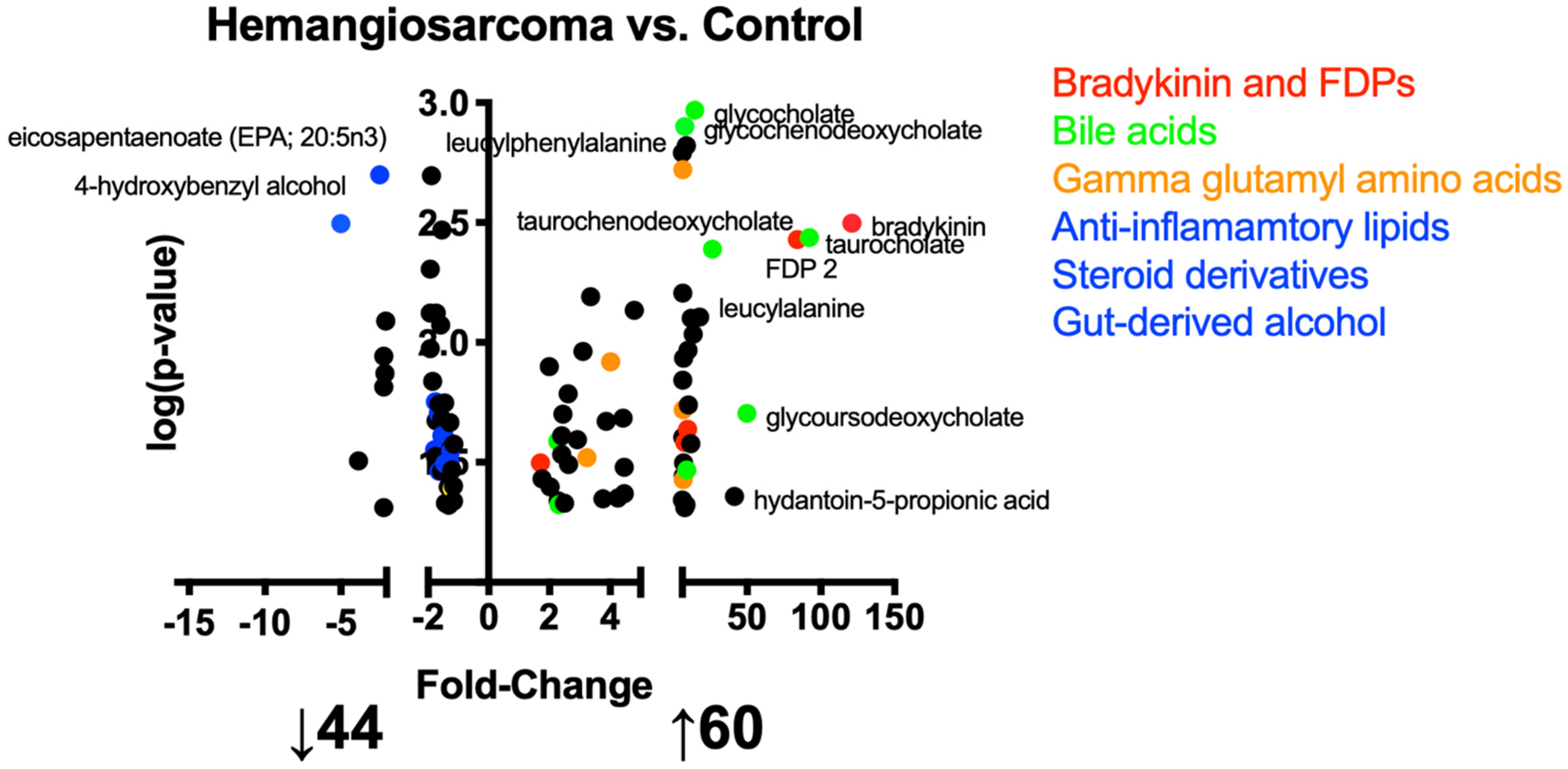
2.2. Metabolites Altered in the Development of vc-Mediated Hemangiosarcoma
2.3. Ontologies Implicated in the Development of VC-Mediated Hemangiosarcoma
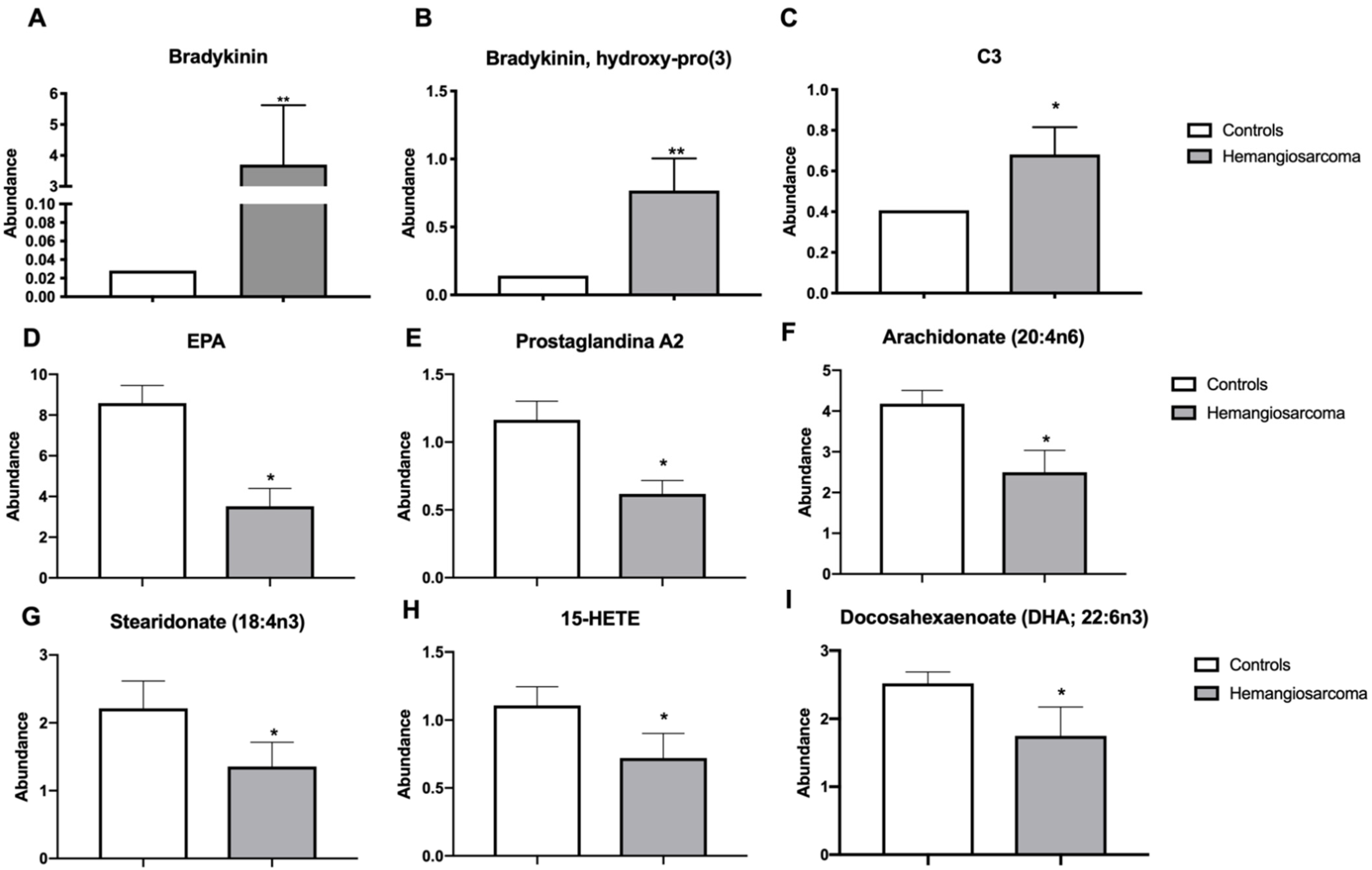
2.4. Serum Inflammatory Proteins Bradykinin and C3 Are Elevated while Anti-Inflammatory Lipids Are Diminished in VC-Mediated Hemangiosarcoma
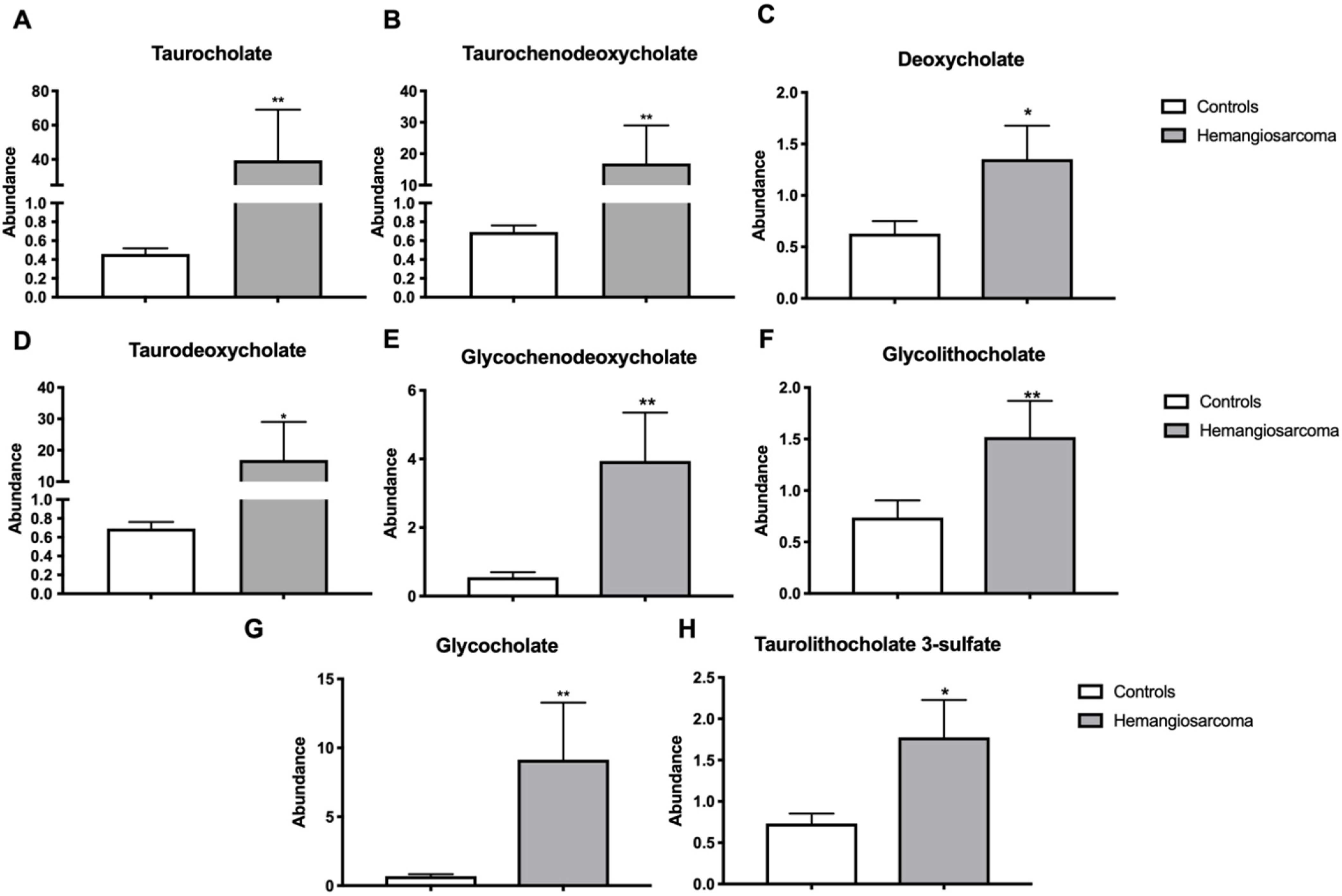
2.5. Serum Bile Acids Are Increased in VC-Mediated Hemangiosarcoma
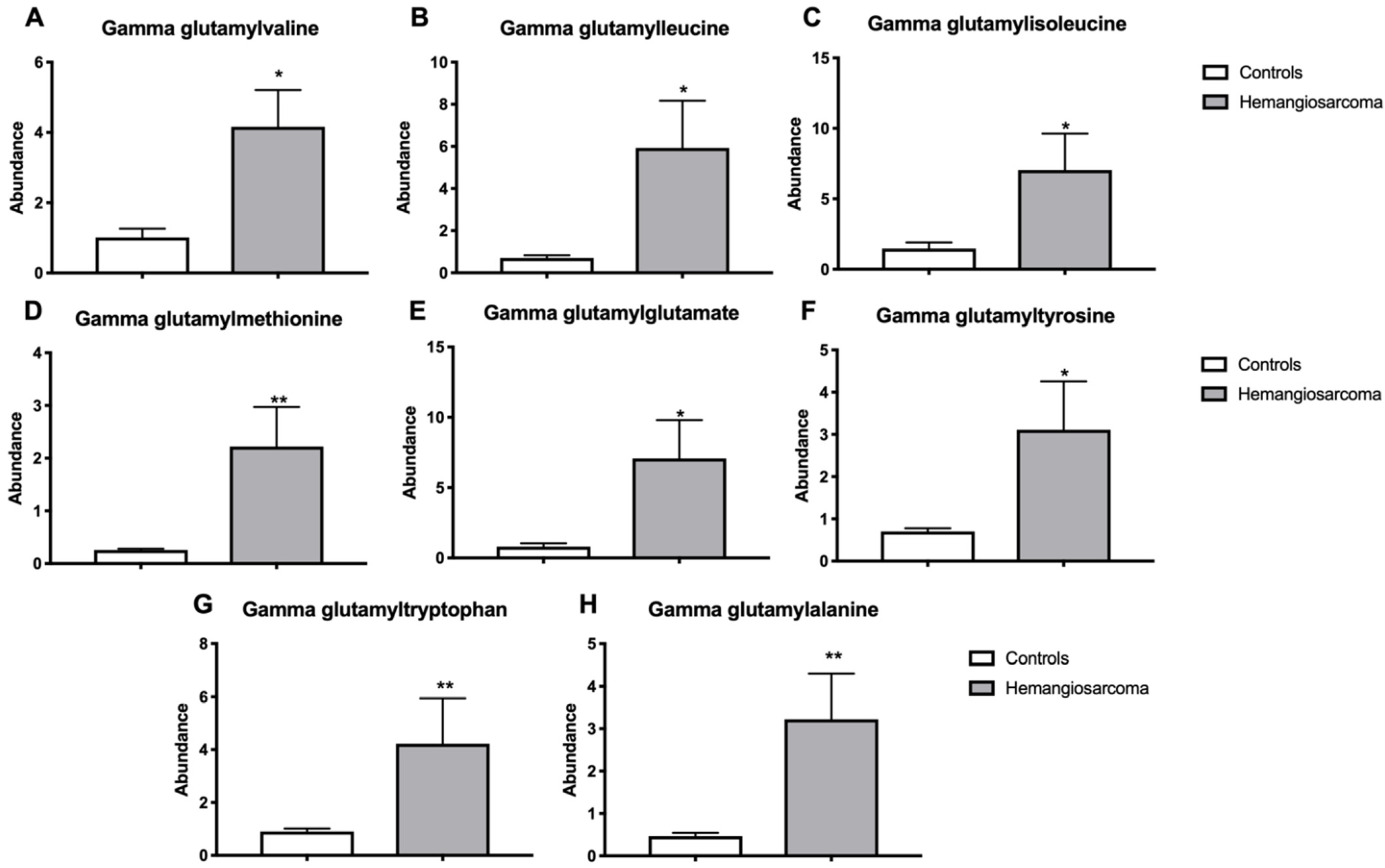
2.6. Serum Gamma-Glutamyl Amino Acids Are Increased in VC-Mediated Hemangiosarcoma
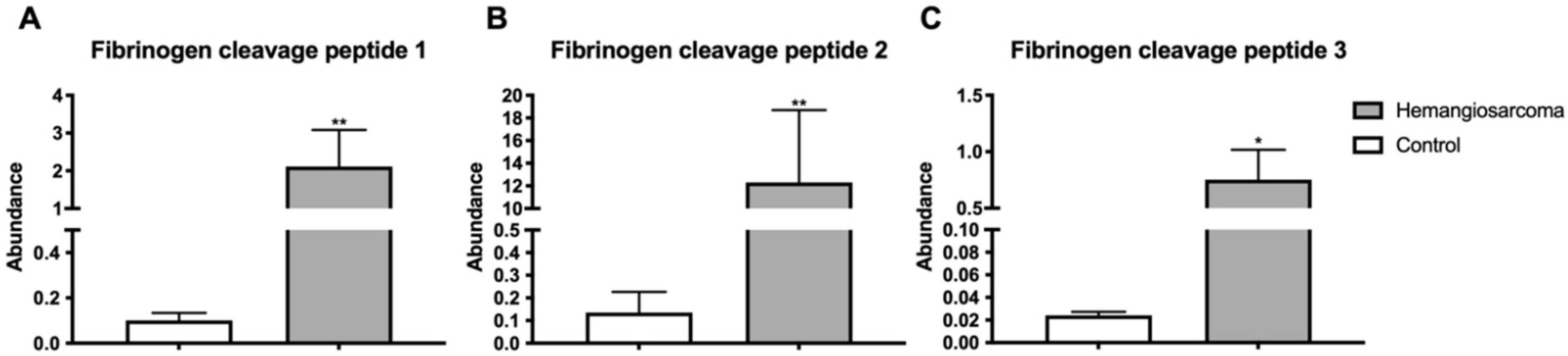
2.7. Fibrin Degradation Products Are Elevated in VC-Mediated Hemangiosarcoma
2.8. Steroid Hormones, Steroid Hormone Metabolites and Gut-Derived Phenol Gastrodigenin Are Diminished in VC-Mediated Hemangiosarcoma
2.9. Random Forest Analysis Demonstrates That Metabolites from the Hemangiosarcoma and Control Are Statistically Distinct
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Metabolomics
4.3. MetaCore Analysis
4.4. Random Forest Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Cohen, S.M.; Storer, R.D.; Criswell, K.A.; Doerrer, N.G.; Dellarco, V.L.; Pegg, D.G.; Wojcinski, Z.W.; Malarkey, D.E.; Jacobs, A.C.; Klaunig, J.E.; et al. Hemangiosarcoma in Rodents: Mode-of-Action Evaluation and Human Relevance. Toxicol. Sci. 2009, 111, 4–18. [Google Scholar] [CrossRef] [Green Version]
- Swenberg, J.A.; Lu, K.; Moeller, B.C.; Gao, L.; Upton, P.B.; Nakamura, J.; Starr, T.B. Endogenous versus exogenous DNA adducts: Their role in carcinogenesis, epidemiology, and risk assessment. Toxicol. Sci. Off. J. Soc. Toxicol. 2011, 120, 130–145. [Google Scholar] [CrossRef]
- Creech, J.L.; Johnson, M.N. Angiosarcoma of liver in the manufacture of polyvinyl chloride. J. Occup. Med. Off. Publ. Ind. Med. Assoc. 1974, 16, 150–152. [Google Scholar]
- Falk, H.; Creech, J.L., Jr.; Heath, C.W., Jr.; Johnson, M.N.; Key, M.M. Hepatic disease among workers at a vinyl chloride polymerization plant. JAMA 1974, 230, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Mundt, K.A.; Dell, L.D.; Crawford, L.; Gallagher, A.E. Quantitative estimated exposure to vinyl chloride and risk of angiosarcoma of the liver and hepatocellular cancer in the US industry-wide vinyl chloride cohort: Mortality update through 2013. Occup. Environ. Med. 2017, 74, 709–716. [Google Scholar] [CrossRef]
- Di Lorenzo, L.; Corfiati, M.; Catacchio, T. Liver angiosarcoma from past exposure to vinyl chloride: A case report. La Med. Del Lav. 2012, 103, 459–465. [Google Scholar]
- Fedeli, U.; Girardi, P.; Mastrangelo, G. Occupational exposure to vinyl chloride and liver diseases. World J. Gastroenterol 2019, 25, 4885–4891. [Google Scholar] [CrossRef]
- EASL Clinical Practice Guideline: Occupational liver diseases. J. Hepatol. 2019, 71, 1022–1037. [CrossRef] [PubMed] [Green Version]
- Zheng, Y.W.; Zhang, X.W.; Zhang, J.L.; Hui, Z.Z.; Du, W.J.; Li, R.M.; Ren, X.B. Primary hepatic angiosarcoma and potential treatment options. J. Gastroenterol. Hepatol. 2014, 29, 906–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cave, M.; Khan, R.; Reynolds, L.; Ray, M.; McClain, C.J. Vinyl Chloride Induced Hepatic Angiosarcoma: An Update of the Louisville Experience [abstract]. Gastroenterology 2007, 132, 133. [Google Scholar]
- Cave, M.; Falkner, K.C.; Ray, M.; Joshi-Barve, S.; Brock, G.; Khan, R.; Bon Homme, M.; McClain, C.J. Toxicant-associated steatohepatitis in vinyl chloride workers. Hepatology 2010, 51, 474–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guardiola, J.J.; Beier, J.I.; Falkner, K.C.; Wheeler, B.; Mcclain, C.J.; Cave, M. Occupational exposures at a polyvinyl chloride production facility are associated with significant changes to the plasma metabolome. Toxicol. Appl. Pharmacol. 2016, 313, 47–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golias, C.; Charalabopoulos, A.; Stagikas, D.; Charalabopoulos, K.; Batistatou, A. The kinin system--bradykinin: Biological effects and clinical implications. Multiple role of the kinin system—bradykinin. Hippokratia 2007, 11, 124–128. [Google Scholar] [PubMed]
- Xu, C.; Chen, Y.; Xu, L.; Miao, M.; Li, Y.; Yu, C. Serum complement C3 levels are associated with nonalcoholic fatty liver disease independently of metabolic features in Chinese population. Sci. Rep. 2016, 6, 23279. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Zhu, B.; Li, Y. Resolution of Cancer-Promoting Inflammation: A New Approach for Anticancer Therapy. Front. Immunol. 2017, 8, 71. [Google Scholar] [CrossRef] [Green Version]
- Jia, W.; Xie, G.; Jia, W. Bile acid–microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat. Rev. Gastroenterol. Hepatol. 2017, 15, 111–128. [Google Scholar] [CrossRef] [Green Version]
- Corti, A.; Franzini, M.; Paolicchi, A.; Pompella, A. Gamma-glutamyltransferase of cancer cells at the crossroads of tumor progression, drug resistance and drug targeting. Anticancer. Res. 2010, 30, 1169–1181. [Google Scholar]
- Kotronen, A.; Joutsi-Korhonen, L.; Sevastianova, K.; Bergholm, R.; Hakkarainen, A.; Pietiläinen, K.H.; Lundbom, N.; Rissanen, A.; Lassila, R.; Yki-Järvinen, H. Increased coagulation factor VIII, IX, XI and XII activities in non-alcoholic fatty liver disease. Liver Int. 2011, 31, 176–183. [Google Scholar] [CrossRef]
- He, J.; Cheng, Q.; Xie, W. Minireview: Nuclear Receptor-Controlled Steroid Hormone Synthesis and Metabolism. Mol. Endocrinol. 2010, 24, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Lou, S.; Balluff, B.; de Graaff, M.A.; Cleven, A.H.G.; Briaire—de Bruijn, I.; Bovée, J.V.M.G.; McDonnell, L.A. High-grade sarcoma diagnosis and prognosis: Biomarker discovery by mass spectrometry imaging. Proteomics 2016, 16, 1802–1813. [Google Scholar] [CrossRef]
- Lou, S.; Balluff, B.; Cleven, A.H.G.; Bovée, J.V.M.G.; McDonnell, L.A. Prognostic Metabolite Biomarkers for Soft Tissue Sarcomas Discovered by Mass Spectrometry Imaging. J. Am. Soc. Mass Spectrom. 2017, 28, 376–383. [Google Scholar] [CrossRef] [Green Version]
- Clapp, C.; Thebault, S.; Jeziorski, M.C.; Martinez De La Escalera, G. Peptide hormone regulation of angiogenesis. Physiol. Rev. 2009, 89, 1177–1215. [Google Scholar] [CrossRef]
- Ishihara, K.; Kamata, M.; Hayashi, I.; Yamashina, S.; Majima, M. Roles of bradykinin in vascular permeability and angiogenesis in solid tumor. Int. Immunopharmacol. 2002, 2, 499–509. [Google Scholar] [CrossRef]
- Yu, H.-S.; Wang, S.-W.; Chang, A.-C.; Tai, H.-C.; Yeh, H.-I.; Lin, Y.-M.; Tang, C.-H. Bradykinin promotes vascular endothelial growth factor expression and increases angiogenesis in human prostate cancer cells. Biochem. Pharmacol. 2014, 87, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Anders, L.C.; Lang, A.L.; Anwar-Mohamed, A.; Douglas, A.N.; Bushau, A.M.; Falkner, K.C.; Hill, B.G.; Warner, N.L.; Arteel, G.E.; Cave, M.; et al. Vinyl Chloride Metabolites Potentiate Inflammatory Liver Injury Caused by LPS in Mice. Toxicol. Sci. Off. J. Soc. Toxicol. 2016, 151, 312–323. [Google Scholar] [CrossRef] [Green Version]
- Bork, K.; Yasothan, U.; Kirkpatrick, P. Icatibant. Nat. Rev. Drug Discov. 2008, 7, 801–802. [Google Scholar] [CrossRef]
- Antonescu, C.R.; Yoshida, A.; Guo, T.; Chang, N.E.; Zhang, L.; Agaram, N.P.; Qin, L.X.; Brennan, M.F.; Singer, S.; Maki, R.G. KDR activating mutations in human angiosarcomas are sensitive to specific kinase inhibitors. Cancer Res. 2009, 69, 7175–7179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.-C.; Zhang, L.; Sung, Y.-S.; Chen, C.-L.; Kao, Y.-C.; Agaram, N.P.; Singer, S.; Tap, W.D.; D’Angelo, S.; Antonescu, C.R. Recurrent CIC Gene Abnormalities in Angiosarcomas: A Molecular Study of 120 Cases With Concurrent Investigation of PLCG1, KDR, MYC, and FLT4 Gene Alterations. Am. J. Surg. Pathol. 2016, 40, 645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Costa, P.L.N.; Sirois, P.; Tannock, I.F.; Chammas, R. The role of kinin receptors in cancer and therapeutic opportunities. Cancer Lett. 2014, 345, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Maeda, H.; Matsumura, Y.; Kato, H. Purification, and identification of [hydroxyprolyl3] bradykinin in ascitic fluid from a patient with gastric cancer. J. Biol. Chem. 1988, 263, 16051–16054. [Google Scholar] [CrossRef]
- Ravi, V.; Sanford, E.M.; Wang, W.L.; Ross, J.S.; Ramesh, N.; Futreal, A.; Patel, S.; Stephens, P.J.; Miller, V.A.; Ali, S.M. Antitumor Response of VEGFR2- and VEGFR3-Amplified Angiosarcoma to Pazopanib. J. Natl. Compr. Canc. Netw. 2016, 14, 499–502. [Google Scholar] [CrossRef]
- Hoang, N.T.; Acevedo, L.A.; Mann, M.J.; Tolani, B. A review of soft-tissue sarcomas: Translation of biological advances into treatment measures. Cancer Manag. Res. 2018, 10, 1089–1114. [Google Scholar] [CrossRef] [Green Version]
- Radons, J. The Role of Inflammation in Sarcoma. Adv. Exp. Med. Biol. 2014, 816, 259–313. [Google Scholar] [PubMed]
- Kwaan, H.; Keer, H. Fibrinolysis and Cancer. Semin. Thromb. Hemost. 1990, 16, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Maragoudakis, M.E.; Tsopanoglou, N.E.; Andriopoulou, P. Mechanism of thrombin-induced angiogenesis. Biochem. Soc. Trans. 2002, 30, 172–177. [Google Scholar] [CrossRef]
- Tang, L.; Han, X. The urokinase plasminogen activator system in breast cancer invasion and metastasis. Biomed. Pharmacother. 2013, 67, 179–182. [Google Scholar] [CrossRef]
- Thompson, W.D.; Smith, E.B.; Stirk, C.M.; Marshall, F.I.; Stout, A.J.; Kocchar, A. Angiogenic activity of fibrin degradation products is located in fibrin fragment E. J. Pathol. 1992, 168, 47–53. [Google Scholar] [CrossRef]
- Astedt, B.; Svanberg, L.; Nilsson, I.M. Fibrin degradation products and ovarian tumours. Br. Med. J. 1971, 4, 458–459. [Google Scholar] [CrossRef] [Green Version]
- Kerber, A.; Trojan, J.; Herrlinger, K.; Zgouras, D.; Caspary, W.F.; Braden, B. The new DR-70 immunoassay detects cancer of the gastrointestinal tract: A validation study. Aliment. Pharmacol. Ther. 2004, 20, 983–987. [Google Scholar] [CrossRef]
- Lin, S.-Z.; Chen, C.-C.; Lee, K.-C.; Tseng, C.-W.; Lin, H.-Y.; Chen, Y.-C.; Lin, H.-C. DR-70 immunoassay for the surveillance of hepatocellular carcinoma. J. Gastroenterol. Hepatol. 2012, 27, 547–552. [Google Scholar] [CrossRef]
- So, H.J.; Hong, S.-I.; Lee, J.K.; Chang, Y.H.; Kang, S.J.; Hong, Y.J. Comparison of the serum fibrin-fibrinogen degradation products with cytokeratin 19 fragment as biomarkers in patients with lung cancer. Biomed. Rep. 2014, 2, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Lang, A.L.; Chen, L.; Poff, G.D.; Ding, W.-X.; Barnett, R.A.; Arteel, G.E.; Beier, J.I. Vinyl chloride dysregulates metabolic homeostasis and enhances diet-induced liver injury in mice. Hepatol. Commun. 2018, 2, 270–284. [Google Scholar] [CrossRef] [Green Version]
- Hammer, A.S.; Couto, C.G.; Swardson, C.; Getzy, D. Hemostatic abnormalities in dogs with hemangiosarcoma. J. Vet. Intern. Med. 1991, 5, 11–14. [Google Scholar] [CrossRef]
- Liss, G.M.; Greenberg, R.A.; Tamburro, C.H. Use of serum bile acids in the identification of vinyl chloride hepatotoxicity. Am. J. Med. 1985, 78, 68–76. [Google Scholar] [CrossRef]
- Yin, P.; Wan, D.; Zhao, C.; Chen, J.; Zhao, X.; Wang, W.; Lu, X.; Yang, S.; Gu, J.; Xu, G. A metabonomic study of hepatitis B-induced liver cirrhosis and hepatocellular carcinoma by using RP-LC and HILIC coupled with mass spectrometry. Mol. Biosyst. 2009, 5, 868. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Xie, G.; Wang, X.; Fan, J.; Qiu, Y.; Zheng, X.; Qi, X.; Cao, Y.; Su, M.; Wang, X.; et al. Serum and urine metabolite profiling reveals potential biomarkers of human hepatocellular carcinoma. Mol. Cell. Proteom. 2011, 10, M110.004945. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Zhao, R.; Zhou, X.; Liang, X.; Campbell, D.J.W.; Zhang, X.; Zhang, L.; Shi, R.; Wang, G.; Pandak, W.M.; et al. Conjugated bile acids promote cholangiocarcinoma cell invasive growth through activation of sphingosine 1-phosphate receptor 2. Hepatology 2014, 60, 908–918. [Google Scholar] [CrossRef] [PubMed]
- Strom, B.L.; Soloway, R.D.; Rios-Dalenz, J.; Rodriguez-Martinez, H.A.; West, S.L.; Kinman, J.L.; Crowther, R.S.; Taylor, D.; Polansky, M.; Berlin, J.A. Biochemical epidemiology of gallbladder cancer. Hepatology 1996, 23, 1402–1411. [Google Scholar] [CrossRef]
- Salmon, R.J.; Laurent, M.; Thierry, J.P. Effect of taurocholic acid feeding on methyl-nitro-N-nitroso-guanidine induced gastric tumors. Cancer Lett. 1984, 22, 315–320. [Google Scholar] [CrossRef]
- Dent, P.; Fang, Y.; Gupta, S.; Studer, E.; Mitchell, C.; Spiegel, S.; Hylemon, P.B. Conjugated bile acids promote ERK1/2 and AKT activation via a pertussis toxin-sensitive mechanism in murine and human hepatocytes. Hepatology 2005, 42, 1291–1299. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.-H.; Ancrile, B.B.; Kashatus, D.F.; Counter, C.M. Tumour maintenance is mediated by eNOS. Nature 2008, 452, 646–649. [Google Scholar] [CrossRef] [Green Version]
- Weninger, W.; Rendl, M.; Pammer, J.; Mildner, M.; Tschugguel, W.; Schneeberger, C.; Stürzl, M.; Tschachler, E. Nitric oxide synthases in Kaposi’s sarcoma are expressed predominantly by vessels and tissue macrophages. Lab. Investig. J. Tech. Methods Pathol. 1998, 78, 949–955. [Google Scholar]
- Suzuki, H.; Yamada, C.; Kato, K. g-Glutamyl compounds and their enzymatic production using bacterial g-glutamyltranspeptidase. Amino Acids 2007, 32, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Bo, S.; Gambino, R.; Durazzo, M.; Guidi, S.; Tiozzo, E.; Ghione, F.; Gentile, L.; Cassader, M.; Pagano, G.F. Associations between gamma-glutamyl transferase, metabolic abnormalities and inflammation in healthy subjects from a population-based cohort: A possible implication for oxidative stress. World J. Gastroenterol. 2005, 11, 7109–7117. [Google Scholar] [CrossRef]
- Soga, T.; Sugimoto, M.; Honma, M.; Mori, M.; Igarashi, K.; Kashikura, K.; Ikeda, S.; Hirayama, A.; Yamamoto, T.; Yoshida, H.; et al. Serum metabolomics reveals γ-glutamyl dipeptides as biomarkers for discrimination among different forms of liver disease. J. Hepatol. 2011, 55, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Joyce, A.; Yates, K.; Aouizerat, B.; Sanyal, A.J. Metabolomic Profiling to Identify Predictors of Response to Vitamin E for Non-Alcoholic Steatohepatitis (NASH). PLoS ONE 2012, 7, e44106. [Google Scholar] [CrossRef]
- Huang, J.; Mondul, A.M.; Weinstein, S.J.; Koutros, S.; Derkach, A.; Karoly, E.; Sampson, J.N.; Moore, S.C.; Berndt, S.I.; Albanes, D. Serum metabolomic profiling of prostate cancer risk in the prostate, lung, colorectal, and ovarian cancer screening trial. Br. J. Cancer 2016, 115, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Lin, C.-C.; Spasojevic, I.; Iversen, E.S.; Chi, J.-T.; Marks, J.R. A joint analysis of metabolomics and genetics of breast cancer. Breast Cancer Res. 2014, 16, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mark, L.; Delmore, F.; Creech, J.L.; Ogden, L.L., II; Fadell, E.H.; Songster, C.L.; Clanton, J.; Johnson, M.N.; Christopherson, W.M. Clinical and morphologic features of hepatic angiosarcoma in vinyl chloride workers. Cancer 1976, 37, 149–163. [Google Scholar]
- Garcia, A.R.; Ribeiro, J.; Gervásio, H.; Sousa, F.C.E. Hepatic Angiosarcoma Masquerading as Hemangioma: A Challenging Differential Diagnosis. Acta Médica Port. 2017, 30, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, R.H. 4-Hydroxybenzyl alcohol. A metabolite produced during the biosynthesis of thiamine in Escherichia coli. Biochim. Biophys. Acta 1979, 583, 55–62. [Google Scholar] [CrossRef]
- Lim, E.-J.; Kang, H.-J.; Jung, H.-J.; Park, E.-H. Anti-angiogenic, anti-inflammatory and anti-nociceptive activity of 4-hydroxybenzyl alcohol. J. Pharm. Pharm. 2007, 59, 1235–1240. [Google Scholar] [CrossRef]
- Luo, L.; Kim, S.-W.; Lee, H.-K.; Kim, I.-D.; Lee, H.; Lee, J.-K. Anti-oxidative effects of 4-hydroxybenzyl alcohol in astrocytes confer protective effects in autocrine and paracrine manners. PLoS ONE 2017, 12, e0177322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberto De Oliveira, M.; Bittencourt Brasil, F.; Fürstenau, C.R. Evaluation of the Mitochondria-Related Redox and Bioenergetics Effects of Gastrodin in SH-SY5Y Cells Exposed to Hydrogen Peroxide. J. Mol. Neurosci. 2018, 64, 242–251. [Google Scholar] [CrossRef]
- Laschke, M.W.; Vorsterman van Oijen, A.E.; Scheuer, C.; Menger, M.D. In vitro and in vivo evaluation of the anti-angiogenic actions of 4-hydroxybenzyl alcohol. Br. J. Pharmacol. 2011, 163, 835–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Eliseo, D.; Velotti, F. Omega-3 Fatty Acids and Cancer Cell Cytotoxicity: Implications for Multi-Targeted Cancer Therapy. J. Clin. Med. 2016, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Edwards, I.J.; Sun, H.; Hu, Y.; Berquin, I.M.; O’Flaherty, J.T.; Cline, J.M.; Rudel, L.L.; Chen, Y.Q. In Vivo and in Vitro Regulation of Syndecan 1 in Prostate Cells by n -3 Polyunsaturated Fatty Acids. J. Biol. Chem. 2008, 283, 18441–18449. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Berquin, I.M.; Owens, R.T.; O’Flaherty, J.T.; Edwards, I.J. Peroxisome proliferator-activated receptor gamma-mediated up-regulation of syndecan-1 by n-3 fatty acids promotes apoptosis of human breast cancer cells. Cancer Res. 2008, 68, 2912–2919. [Google Scholar] [CrossRef] [Green Version]
- Subbarayan, V.; Xu, X.-C.; Kim, J.; Yang, P.; Hoque, A.; Sabichi, A.L.; Llansa, N.; Mendoza, G.; Logothetis, C.J.; Newman, R.A.; et al. Inverse relationship between 15-lipoxygenase-2 and PPAR-gamma gene expression in normal epithelia compared with tumor epithelia. Neoplasia 2005, 7, 280–293. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Li, M.-Y.; Ma, L.T.; Hsin, M.K.Y.; Mok, T.S.K.; Underwood, M.J.; Chen, G.G. 5-Lipoxygenases and its metabolites 15(S)-HETE and 13(S)-HODE in the development of non-small cell lung cancer. Thorax 2010, 65, 321–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shappell, S.B.; Gupta, R.A.; Manning, S.; Whitehead, R.; Boeglin, W.E.; Schneider, C.; Case, T.; Price, J.; Jack, G.S.; Wheeler, T.M.; et al. 15S-Hydroxyeicosatetraenoic acid activates peroxisome proliferator-activated receptor gamma and inhibits proliferation in PC3 prostate carcinoma cells. Cancer Res. 2001, 61, 497–503. [Google Scholar]
- Chen, G.G.; Xu, H.; Lee, J.F.Y.; Subramaniam, M.; Leung, K.L.; Wang, S.H.; Chan, U.P.F.; Spelsberg, T.C. 15-hydroxy-eicosatetraenoic acid arrests growth of colorectal cancer cellsvia a peroxisome proliferator-activated receptor gamma-dependent pathway. Int. J. Cancer 2003, 107, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Mariani, F.; Sena, P.; Roncucci, L. Inflammatory pathways in the early steps of colorectal cancer development. World J. Gastrenterol. 2014, 20, 9716–9731. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, R.A.; Tamburro, C.H. Exposure indices for epidemiological surveillance of carcinogenic agents in an industrial chemical environment. J. Occup. Med. Off. Publ. Ind. Med. Assoc. 1981, 23, 353–358. [Google Scholar]
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Controls | Hemangiosarcoma |
|---|---|---|
| Number (unless otherwise noted) | 17 | 15 |
| Age at sampling (years) | 50.5 ± 6.3 | 56.2 ± 10.9 |
| BMI (kg/m2) | 27.1 ± 3.4 (n = 16) | 25.3 ± 2.1 (n = 12) |
| Employment duration at sampling (years) | 25.5 ± 5.7 | 31.9 ± 12.0 |
| Cumulative Exposure Rank Month (CERM) | 1156 ± 420.4 | 1266 ± 541.2 |
| Duration of sample storage (years) | 35.0 ± 5.1 | 27.5 ± 12.0 a |
| Analyte (Reference Range) | Controls | Hemangiosarcoma |
|---|---|---|
| Number (unless otherwise specified) | 17 | 12 |
| Albumin (3.5–5.0 mg/dL) | 4.6 ± 0.2 | 4.4 ± 0.2 |
| Total bilirubin (0.2–1.0 mg/dL) | 0.6 ± 0.2 | 0.6 ± 0.2 |
| Alkaline phosphatase (38–126 U/L) | 70.5 ± 15.0 | 68.3 ± 36.2 |
| AST (10–50 U/L) | 24.1 ± 15.6 | 27.8 ± 12.1 |
| ALT (20–70 U/L) | 24.9 ± 18.4 | 19.8 ± 8.3 |
| Triglycerides (10–190 mg/dL) | 153 ± 49.6 | 201 ± 101.6 (n = 8) |
| Total cholesterol (120–200 mg/dL) | 213 ± 32.6 | 217 ± 32.5 (n = 8) |
| Glucose (70–110 mg/dL) | 101 ± 17.3 | 105 ± 16.1 (n = 8) |
| Ontology | p-Value | FDR |
|---|---|---|
| Bile acid biosynthesis | 3.45 × 10−7 | 1.17 × 10−5 |
| Taurine and hypotaurine metabolism | 5.74 × 10−6 | 9.76 × 10−5 |
| Cholesterol metabolism | 1.08 × 10−5 | 1.22 × 10−4 |
| Cholesterol and bile acid homeostasis | 4.56 × 10−5 | 1.10 × 10−3 |
| Renal secretion of inorganic electrolytes | 1.17 × 10−4 | 9.91 × 10−4 |
| Renal secretion of organic electrolytes/rodent version | 2.57 × 10−4 | 1.75 × 10−3 |
| Regulation of lipid metabolism_Regulation of acetyl-CoA carboxylase 1 activity | 3.88 × 10−4 | 2.20 × 10−3 |
| Medium-chain saturated fatty acids synthesis | 5.72 × 10−4 | 2.78 × 10−3 |
| Upregulation of IL-8 expression in colorectal cancer | 1.24 × 10−3 | 5.27 × 10−3 |
| Carbohydrate metabolism_TCA and tricarboxylic acid transport | 1.97 × 10−3 | 4.74 × 10−2 |
| Oxidative phosphorylation | 2.03 × 10−3 | 7.68 × 10−3 |
| Metabolic diseases | 3.28 × 10−3 | 3.93 × 10−2 |
| Oxidative stress Role of Sirtuin1 and PGC1-alpha in activation of antioxidant defense system | 4.31 × 10−3 | 1.46 × 10−2 |
| Ontology | p-Value | FDR |
|---|---|---|
| Nicotine signaling in cholinergic neurons | 1.65 × 10−5 | 4.94 × 10−4 |
| L-Lysine metabolism | 1.06 × 10−4 | 1.20 × 10−3 |
| L-Tryptophan metabolism (part 2) | 1.20 × 10−4 | 1.20 × 10−3 |
| Nicotine action | 1.48 × 10−4 | 5.02 × 10−3 |
| (L)-carnitine pathway | 3.12 × 10−4 | 3.12 × 10−3 |
| N-Acylethanolamines, HSRL5-transacylation pathway | 3.44 × 10−4 | 2.58 × 10−3 |
| Development_Neuromuscular junction | 1.25 × 10−3 | 3.76 × 10−3 |
| Neurophysiological process_Transmission of nerve impulse | 2.60 × 10−3 | 3.89 × 10−3 |
| Renal secretion of organic electrolytes/Rodent version | 2.79 × 10−3 | 1.67 × 10−2 |
| Cholesterol biosynthesis | 4.93 × 10−3 | 2.46 × 10−2 |
| Steroid metabolism_Cholesterol biosynthesis | 9.27 × 10−3 | 4.64 × 10−2 |
| Nicotine signaling in cholinergic neurons | 1.65 × 10−5 | 4.94 × 10−4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guardiola, J.J.; Hardesty, J.E.; Beier, J.I.; Prough, R.A.; McClain, C.J.; Cave, M.C. Plasma Metabolomics Analysis of Polyvinyl Chloride Workers Identifies Altered Processes and Candidate Biomarkers for Hepatic Hemangiosarcoma and Its Development. Int. J. Mol. Sci. 2021, 22, 5093. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105093
Guardiola JJ, Hardesty JE, Beier JI, Prough RA, McClain CJ, Cave MC. Plasma Metabolomics Analysis of Polyvinyl Chloride Workers Identifies Altered Processes and Candidate Biomarkers for Hepatic Hemangiosarcoma and Its Development. International Journal of Molecular Sciences. 2021; 22(10):5093. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105093
Chicago/Turabian StyleGuardiola, John J., Josiah E. Hardesty, Juliane I. Beier, Russell A. Prough, Craig J. McClain, and Matthew C. Cave. 2021. "Plasma Metabolomics Analysis of Polyvinyl Chloride Workers Identifies Altered Processes and Candidate Biomarkers for Hepatic Hemangiosarcoma and Its Development" International Journal of Molecular Sciences 22, no. 10: 5093. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105093