
Some Dietary Phenolic Compounds Can Activate Thyroid Peroxidase and Inhibit Lipoxygenase-Preliminary Study in the Model Systems

 , ,
, , 
Abstract
:1. Introduction
2. Results and Discussion
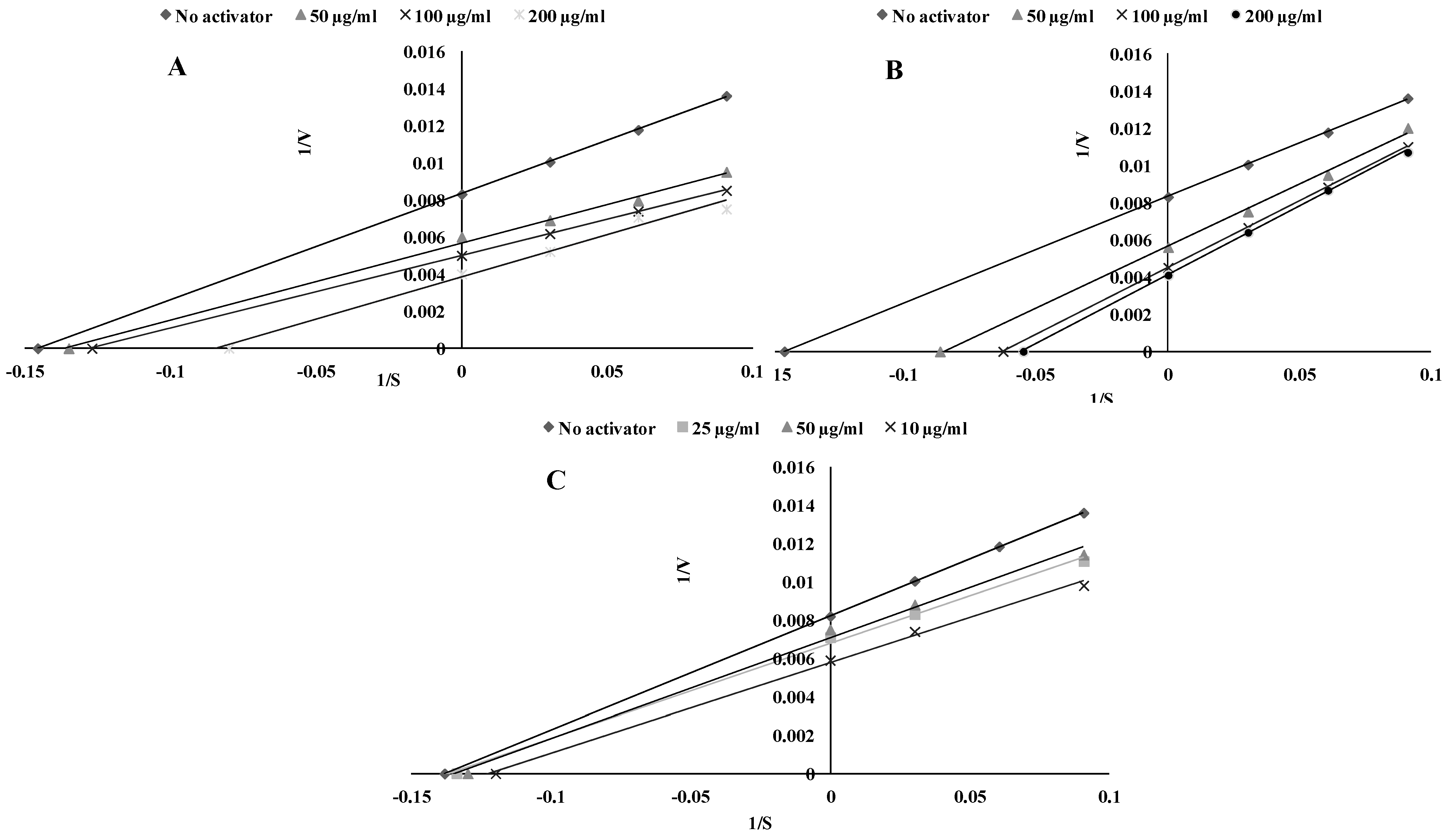
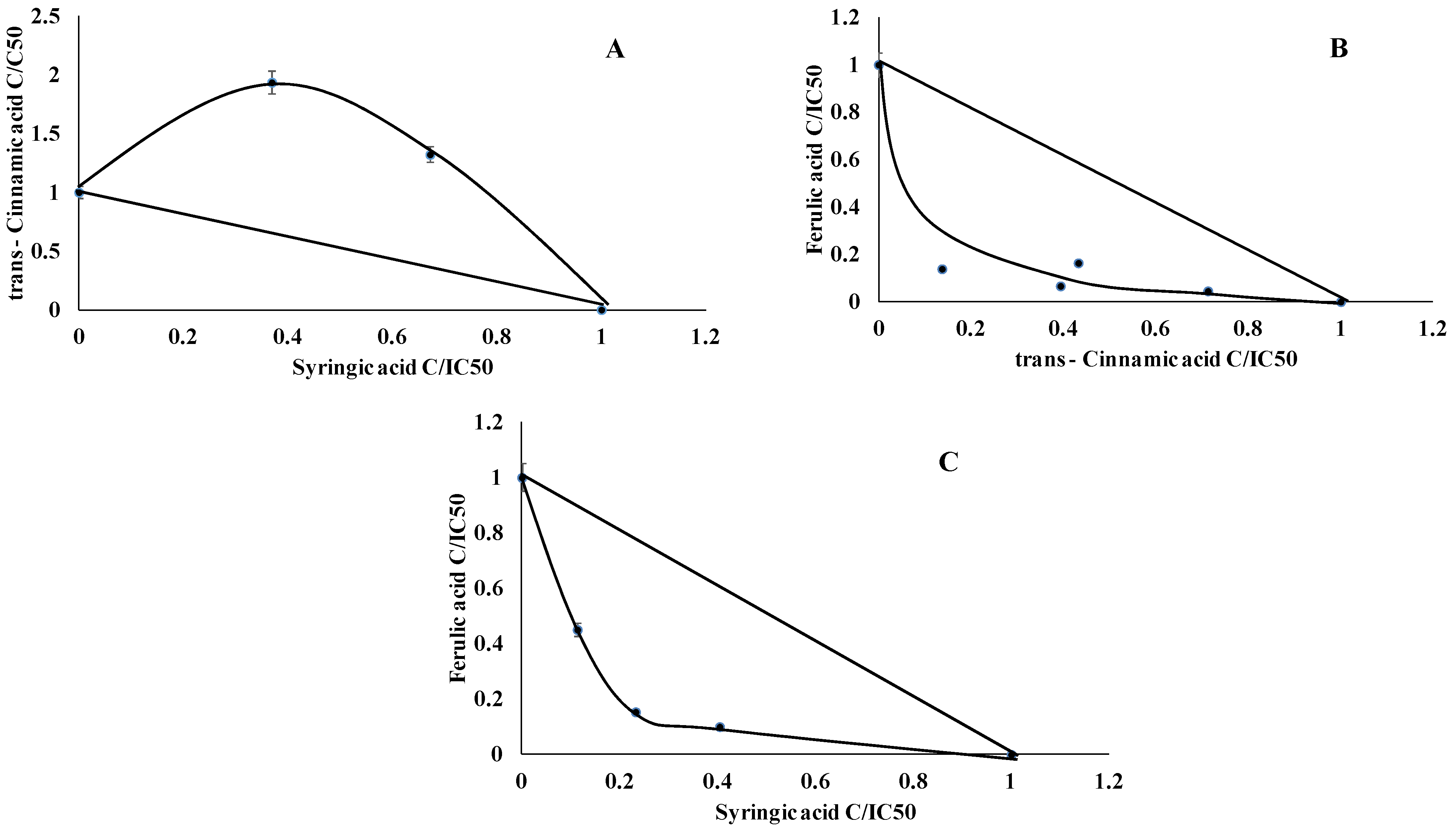
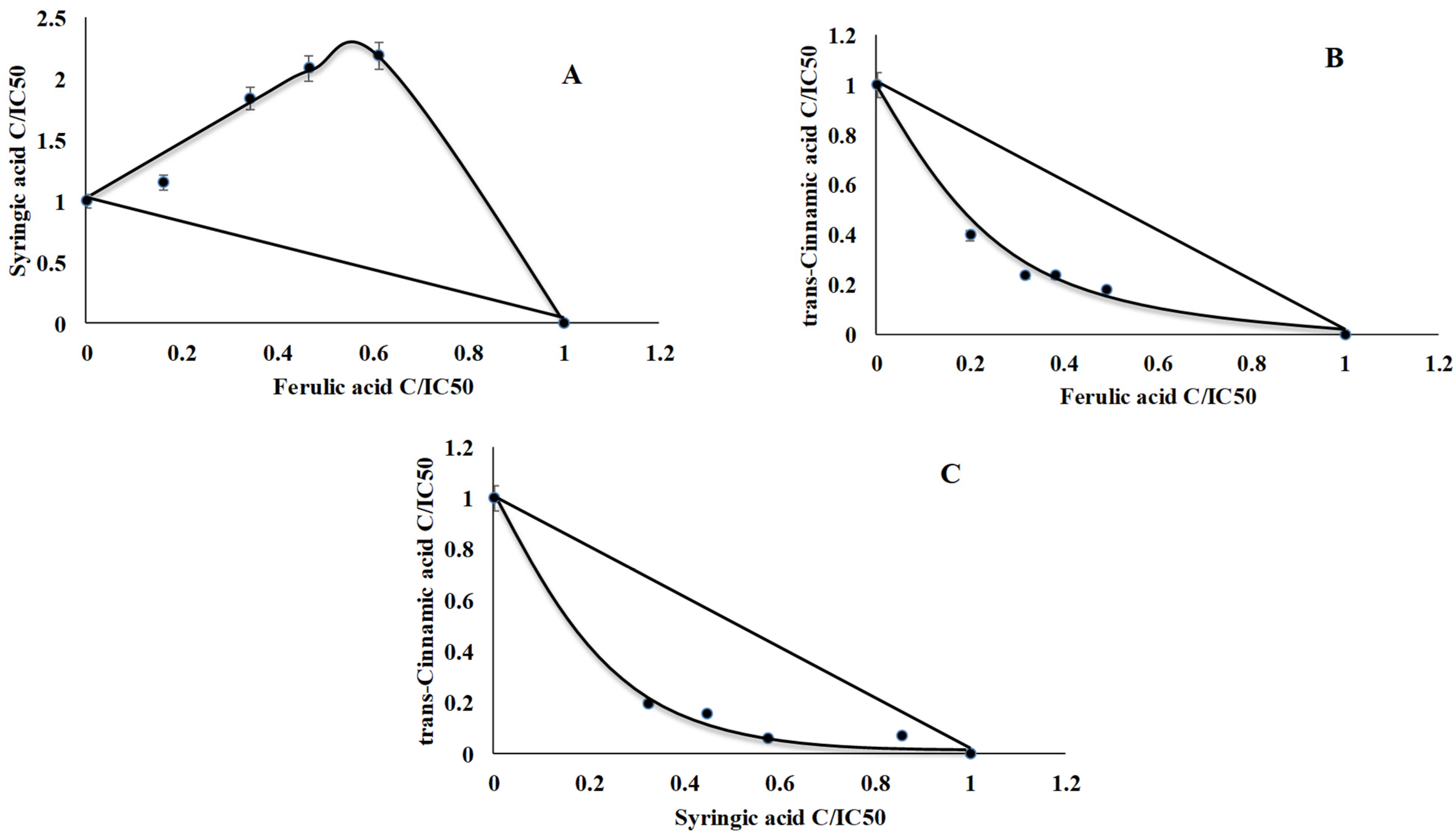
2.1. TPO Assay
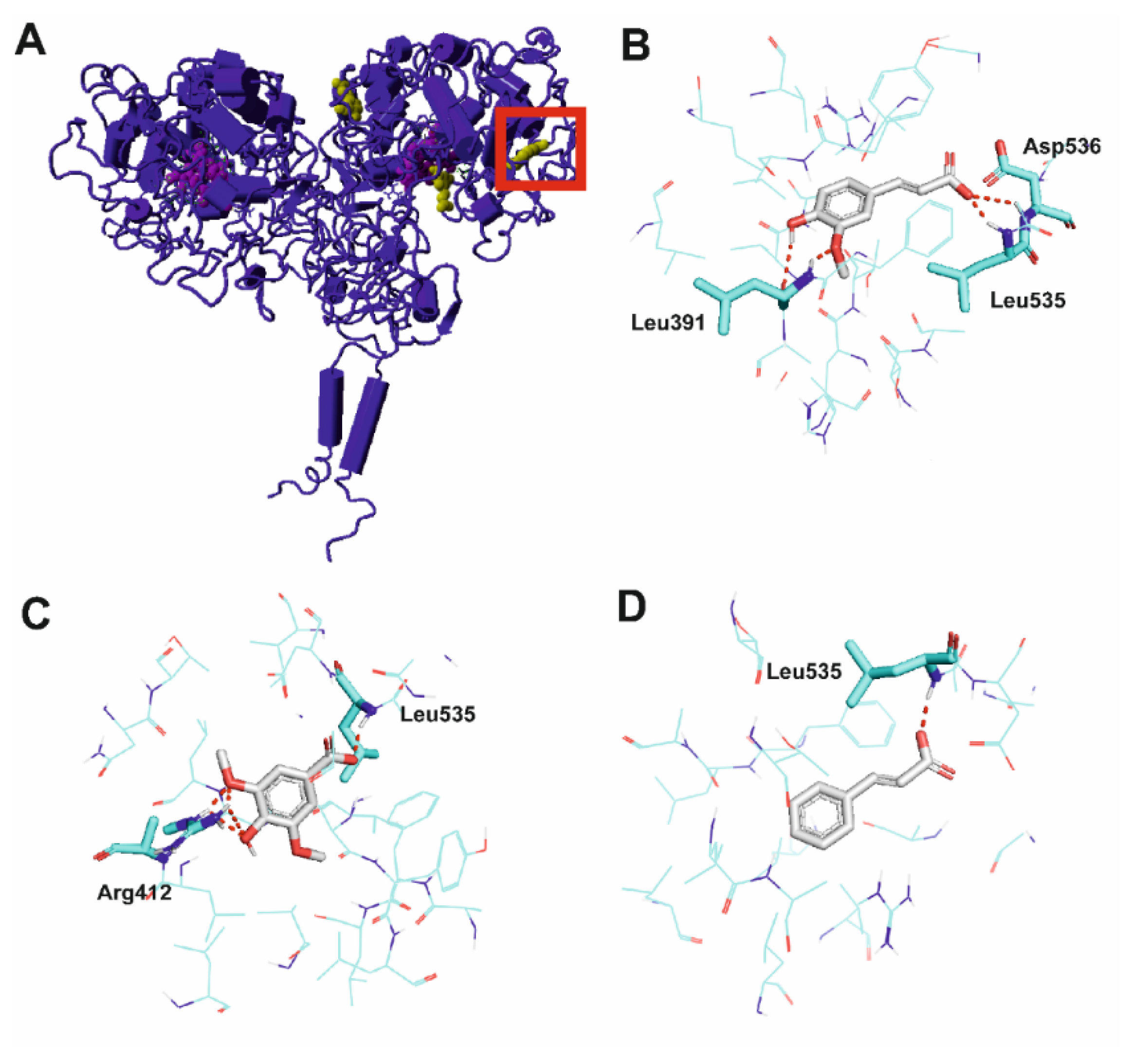
2.2. Molecular Modeling of TPO Activation
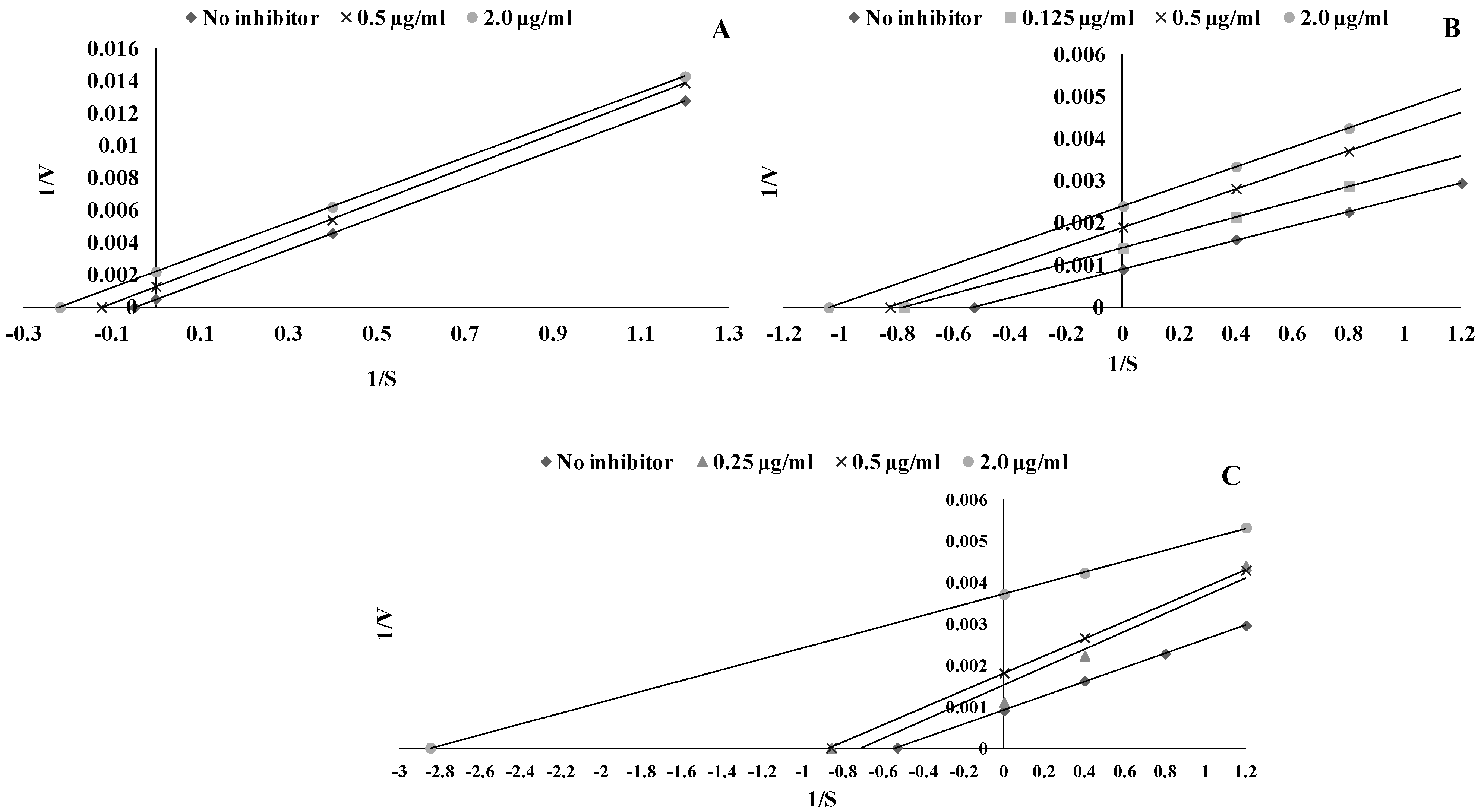
2.3. LOX Assay
2.4. Molecular Modeling of LOX Inhibition
2.5. Antiradical Analysis
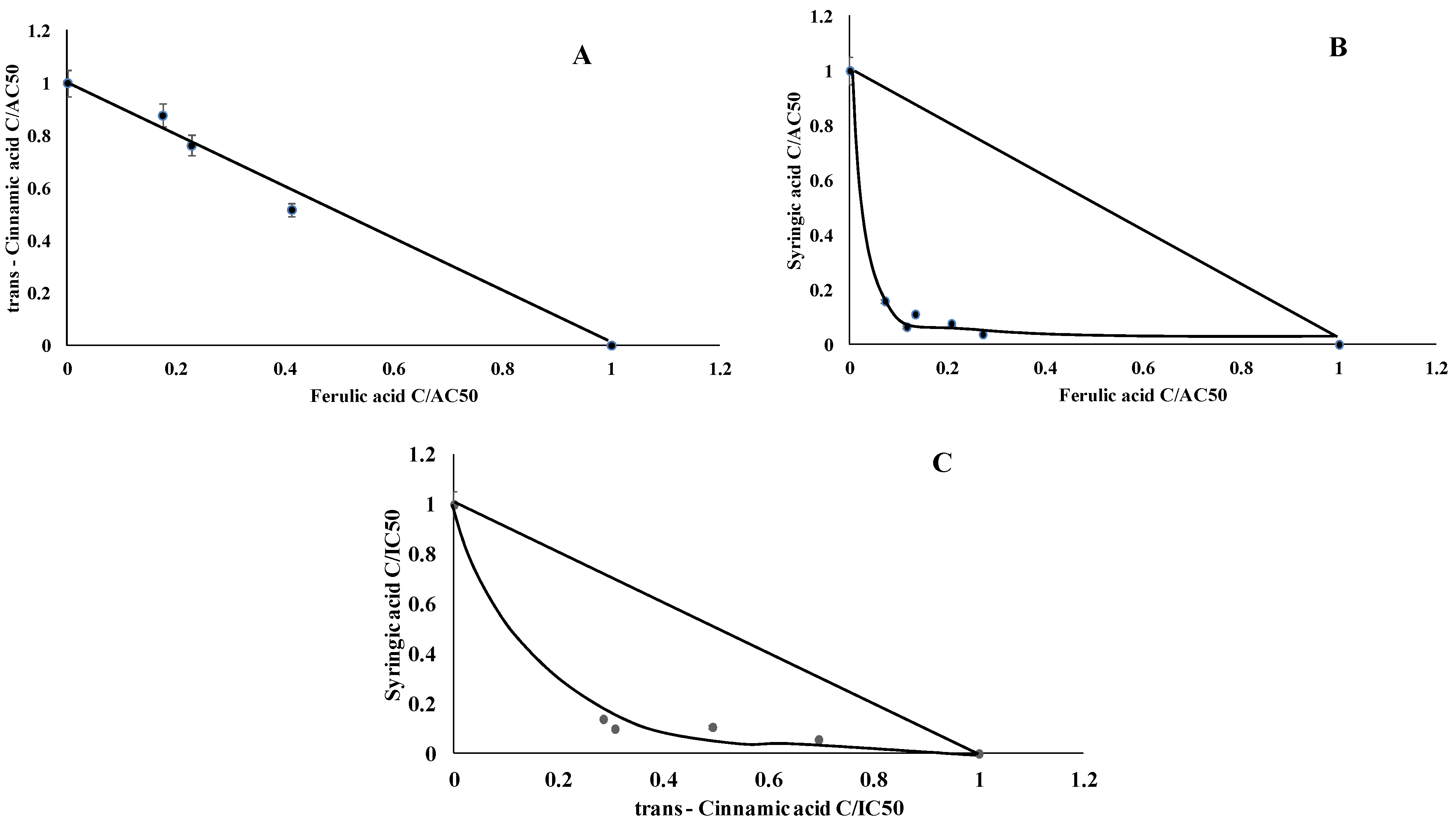
2.6. Interaction Assay
3. Materials and Methods
3.1. Chemicals
3.2. Preparation of Phenolic Acids Solutions
3.3. Molecular Modeling
3.3.1. Small Molecules Modeling
3.3.2. Protein Models
3.3.3. Binding Site Identification and Molecular Docking
3.4. Enzymatic Assay
3.4.1. TPO Preparation
3.4.2. TPO Assay
3.4.3. Inhibition of Lipoxygenase Activity (LOXI)
3.5. In Vitro Antiradical Capacity Assay
3.6. Isobolographic Analysis
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Y.K.; Lin, C.L.; Cheng, F.T.F.; Sung, F.C.; Kao, C.H. Cancer risk in patients with Hashimoto’s thyroiditis: A nationwide cohort study. Br. J. Cancer 2013, 109, 2496–2501. [Google Scholar] [CrossRef] [Green Version]
- Carocho, M.; Ferreira, I.C.F.R. A review on antioxidants, prooxidants and related controversy: Natural and synthetic compounds, screening and analysis methodologies and future perspectives. Food Chem. Toxicol. 2013, 51, 15–25. [Google Scholar] [CrossRef]
- Mancini, A.; Di Segni, C.; Raimondo, S.; Olivieri, G.; Silvestrini, A.; Meucci, E.; Currò, D. Thyroid hormones, oxidative stress, and inflammation. Mediators Inflamm. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habza-Kowalska, E.; Kaczor, A.A.; Żuk, J.; Matosiuk, D.; Gawlik-Dziki, U. Thyroid peroxidase activity is inhibited by phenolic compounds-impact of interaction. Molecules 2019, 24, 2766. [Google Scholar] [CrossRef] [Green Version]
- Landberg, R.; Sunnerheim, K.; Dimberg, L.H. Avenanthramides as lipoxygenase inhibitors. Heliyon 2020, 6, e04304. [Google Scholar] [CrossRef]
- Grzesik, M.; Naparło, K.; Bartosz, G.; Sadowska-Bartosz, I. Antioxidant properties of catechins: Comparison with other antioxidants. Food Chem. 2018, 241, 480–492. [Google Scholar] [CrossRef] [PubMed]
- Gawlik-Dziki, U.; Bryda, J.; Dziki, D.; Swieca, M.; Habza-Kowalska, E.; Złotek, U. Impact of interactions between ferulic and chlorogenic acids on enzymatic and non-enzymatic lipids oxidation: An example of bread enriched with green coffee flour. Appl. Sci. 2019, 9, 568. [Google Scholar] [CrossRef] [Green Version]
- Graf, B.A.; Milbury, P.E.; Blumberg, J.B. Flavonols, flavones, flavanones, and human health: Epidemiological evidence. J. Med. Food 2005, 8, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Hossen, M.S.; Ali, M.Y.; Jahurul, M.H.A.; Abdel-Daim, M.M.; Gan, S.H.; Khalil, M.I. Beneficial roles of honey polyphenols against some human degenerative diseases: A review. Pharmacol. Rep. 2017, 69, 1194–1205. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.-C. Theoretical Basis, Experimental Design, and Computerized Simulation of Synergism and Antagonism in Drug Combination Studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef]
- Paunkov, A.; Chartoumpekis, D.V.; Ziros, P.G.; Chondrogianni, N.; Kensler, T.W.; Sykiotis, G.P. Impact of antioxidant natural compounds on the thyroid gland and implication of the Keap1/Nrf2 signaling pathway. Curr. Pharm. Des. 2019, 25, 1828–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.H. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr. 2003, 78, 517S–520S. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef]
- Gaitan, E.; Lindsay, R.H.; Reichert, R.D.; Ingbarf, S.H.; Cooksey, R.C.; Legan, J.; Meydrech, E.F.; Hill, J.; Kubota, K. Antithyroid and goitrogenic effects of millet: Role of C-Glycosylflavones. J. Clin. Endocrinol. Metab. 1989, 68, 707–714. [Google Scholar] [CrossRef]
- Tsolaki, E.; Eleftheriou, P.; Kartsev, V.; Geronikaki, A.; Saxena, A.K. Application of docking analysis in the prediction and biological evaluation of the lipoxygenase inhibitory action of thiazolyl derivatives of mycophenolic acid. Molecules 2018, 23, 1621. [Google Scholar] [CrossRef] [Green Version]
- Devi, N.S.; Ramanan, M.; Paragi-Vedanthi, P.; Doble, M. Phytochemicals as multi-target inhibitors of the inflammatory pathway- A modeling and experimental study. Biochem. Biophys. Res. Commun. 2017, 484, 467–473. [Google Scholar] [CrossRef]
- Saleem, H.; Zengin, G.; Locatelli, M.; Tartaglia, A.; Ferrone, V.; Htar, T.T.; Naidu, R.; Mahomoodally, M.F.; Ahemad, N. Filago germanica (L.) Huds. bioactive constituents: Secondary metabolites fingerprinting and in vitro biological assays. Ind. Crops Prod. 2020, 152, 112505. [Google Scholar] [CrossRef]
- Gilbert, N.C.; Gerstmeier, J.; Schexnaydre, E.E.; Börner, F.; Garscha, U.; Neau, D.B.; Werz, O.; Newcomer, M.E. Structural and mechanistic insights into 5-lipoxygenase inhibition by natural products. Nat. Chem. Biol. 2020, 16, 783–790. [Google Scholar] [CrossRef]
- Karbownik-Lewińska, M.; Kokoszko-Bilska, A. Oxidative damage to macromolecules in the thyroid-Experimental evidence. Thyroid Res. 2012, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Martysiak-Zurowska, D.; Wenta, W. A comparison of abts and dpph methods for assessing the total antioxidant capacity of human milk. Acta Sci. Pol. Technol. Aliment. 2012, 11, 83–89. [Google Scholar]
- Samsonowicz, M.; Regulska, E.; Karpowicz, D.; Le, B. Antioxidant properties of co ff ee substitutes rich in polyphenols and minerals. Food Chem. 2019, 278, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.Y.; Pei, L.; Liu, Q.; Chen, S.; Dou, H.; Shu, G.; Yuan, Z.X.; Lin, J.; Peng, G.; Zhang, W.; et al. Isobologram analysis: A comprehensive review of methodology and current research. Front. Pharmacol. 2019, 10, 10. [Google Scholar] [CrossRef] [PubMed]
- Gawlik-Dziki, U. Application of Isobolographic Analysis for the Evaluation of Interactions between Bioactive Constituents from Dandelion (Taraxaci Flos) and Lime (Tiliae Flos); Annales Universitatis Mariae Curie-Skłodowska: Lublin, Poland, 2011. [Google Scholar]
- Habza-Kowalska, E.; Gawlik-Dziki, U.; Dziki, D. Mechanism of action and interactions between thyroid peroxidase and lipoxygenase inhibitors derived from plant sources. Biomolecules 2019, 9, 663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durak, A.; Gawlik-Dziki, U.; Pecio, Ł. Coffee with cinnamon-Impact of phytochemicals interactions on antioxidant and anti-inflammatory in vitro activity. Food Chem. 2014, 162, 81–88. [Google Scholar] [CrossRef]
- Durak, A.; Gawlik-Dziki, U.; Kowalska, I. Evaluation of interactions between coffee and cardamom, their type, and strength in relation to interactions in a model system. CyTA-J. Food 2017, 15, 266–276. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Durak, A.; Gawlik-Dziki, U.; Kowlska, I. Coffee with ginger-interactions of biologically active phytochemicals in the model system. Food Chem. 2015, 166, 261–269. [Google Scholar] [CrossRef]
- Dziki, D.; Swieca, M.; Jakubczyk, A.; Gawlik-Dziki, U. Interactions between ferulic and chlorogenic acids as a main factor determining antiradical activity of wholemeal wheat bread enriched with green coffee. Planta Med. 2015, 81, 1486–1487. [Google Scholar] [CrossRef]
- Li, W.; Zhao, X.; Sun, X.; Zu, Y.; Liu, Y.; Ge, Y. Evaluation of antioxidant ability in vitro and bioavailability of trans -cinnamic acid nanoparticle by liquid antisolvent precipitate. J. Nanomater. 2016, 2016, 1–11. [Google Scholar]
- Garrait, G.; Jarrige, J.F.; Blanquet, S.; Beyssac, E.; Cardot, J.M.; Alric, M. Gastrointestinal absorption and urinary excretion of trans-cinnamic and p-coumaric acids in rats. J. Agric. Food Chem. 2006, 54, 2944–2950. [Google Scholar] [CrossRef]
- Adisakwattana, S. Cinnamic acid and its derivatives: Mechanisms for prevention and management of diabetes and its complications. Nutrients 2017, 9, 163. [Google Scholar] [CrossRef] [Green Version]
- Bourne, L.; Paganga, G.; Baxter, D.; Hughes, P.; Rice-Evans, C. Absorption of ferulic acid from low-alcohol beer. Free Radic. Res. 2000, 32, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Mateo Anson, N.; van den Berg, R.; Havenaar, R.; Bast, A.; Haenen, G.R.M.M. Bioavailability of ferulic acid is determined by its bioaccessibility. J. Cereal Sci. 2009, 49, 296–300. [Google Scholar] [CrossRef]
- Srinivasulu, C.; Ramgopal, M.; Ramanjaneyulu, G.; Anuradha, C.M.; Suresh Kumar, C. Syringic acid (SA) ‒ a review of its occurrence, biosynthesis, pharmacological and industrial importance. Biomed. Pharmacother. 2018, 108, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Available online: Https://Pubchem.Ncbi.Nlm.Nih.Gov/Compound/10742 (accessed on 21 April 2021).
- Available online: Https://Pubchem.Ncbi.Nlm.Nih.Gov/Compound/445858 (accessed on 21 April 2021).
- Available online: Https://Pubchem.Ncbi.Nlm.Nih.Gov/Compound/444539 (accessed on 21 April 2021).
- Schrödinger Release 2020–4: LigPrep; Schrödinger, LLC: New York, NY, USA, 2020.
- Schrödinger Release 2020–4: Epik; Schrödinger, LLC: New York, NY, USA, 2020.
- Webb, B.; Sali, A. Comparative protein structure modeling using MODELLER. Curr. Protoc. Bioinform. 2016, 54, 5–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabler, F.; Nam, S.Z.; Till, S.; Mirdita, M.; Steinegger, M.; Söding, J.; Lupas, A.N.; Alva, V. Protein Sequence Analysis Using the MPI Bioinformatics Toolkit. Curr. Protoc. Bioinform. 2020, 72, e108. [Google Scholar] [CrossRef] [PubMed]
- Le, S.N.; Porebski, B.T.; McCoey, J.; Fodor, J.; Riley, B.; Godlewska, M.; Góra, M.; Czarnocka, B.; Paul Banga, J.; Hoke, D.E.; et al. Modelling of thyroid peroxidase reveals insights into its enzyme function and autoantigenicity. PLoS ONE 2015, 10, e0142615. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Anishchenko, I.; Park, H.; Peng, Z.; Ovchinnikov, S.; Baker, D. Improved protein structure prediction using predicted interresidue orientations. Proc. Natl. Acad. Sci. USA 2020, 117, 1496–1503. [Google Scholar] [CrossRef]
- Lomize, M.A.; Lomize, A.L.; Pogozheva, I.D.; Mosberg, H.I. OPM: Orientations of proteins in membranes database. Bioinformatics 2006, 22, 623–625. [Google Scholar] [CrossRef]
- Lindahl, E.; Abraham, M.J.; Hess, B.; van der Spoel, D. GROMACS 2020 Source Code. Zenodo 2018. [Google Scholar] [CrossRef]
- Lee, J.; Cheng, X.; Swails, J.M.; Yeom, M.S.; Eastman, P.K.; Lemkul, J.A.; Wei, S.; Buckner, J.; Jeong, J.C.; Qi, Y.; et al. CHARMM-GUI Input Generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM simulations using the CHARMM36 additive force field. J. Chem. Theory Comput. 2016, 12, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger Release 2020–4: Maestro; Schrödinger, LLC: New York, NY, USA, 2020.
- Patel, J.Z.; Parkkari, T.; Laitinen, T.; Kaczor, A.A.; Saario, S.M.; Savinainen, J.R.; Navia-Paldanius, D.; Cipriano, M.; Leppänen, J.; Koshevoy, I.O.; et al. Chiral 1,3,4-oxadiazol-2-ones as highly selective FAAH inhibitors. J. Med. Chem. 2013, 56, 8484–8496. [Google Scholar] [CrossRef] [PubMed]
- Panjkovich, A.; Daura, X. PARS: A web server for the prediction of Protein Allosteric and Regulatory Sites. Bioinformatics 2014, 30, 1314–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Guilloux, V.; Schmidtke, P.; Tuffery, P. Fpocket: An open source platform for ligand pocket detection. BMC Bioinform. 2009, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Edelsbrunner, H.; Woodward, C. Anatomy of protein pockets and cavities: Measurement of binding site geometry and implications for ligand design. Protein Sci. 1998, 7, 1884–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrödinger Release 2020–4: Glide; Schrödinger, LLC: New York, NY, USA, 2020.
- Available online: Http://www.Yasara.Org (accessed on 21 April 2021).
- Jomaa, B. Simple and rapid in vitro assay for detecting human thyroid peroxidase disruption. ALTEX 2015, 32, 191–200. [Google Scholar] [PubMed] [Green Version]
- Axelrod, B.; Cheesbrough, T.; Laakso, S. Lipoxygenase from Soybeans; Elsevier Inc.: Amsterdam, The Netherlands, 1981; Volume 71, pp. 441–451. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Km [mM] | Vmax [ΔAU/min] | AC50 [mM] |
|---|---|---|---|
| Guaiacol 1 | 0.06 ± 0.003 | 120.5 ± 5.38 d | - |
| trans–cinnamic acid | - | 128.2 ± 4.12 c | 0.10 ± 0.005 c |
| Syringic acid | - | 243.9 ± 9.54 b | 0.69 ± 0.034 a |
| Ferulic acid | - | 294.1 ± 9.12 a | 0.39 ± 0.019 b |
| Compound | Mode of Inhibition | Ki [mM] (Km without Inhibitor) | Vmax [ΔAU/min] | IC50 [mM] |
|---|---|---|---|---|
| Linoleic acid * | - | 0.007 ± 0.0003 a | 1111 ± 45.1 a | - |
| trans–cinnamic acid | uncompetitive | 0.009 ± 0.0004 c | 556 ± 10.13 b | 0.009 ± 0.0004 b |
| Syringic acid | uncompetitive | 0.005 ± 0.0003 d | 526.3 ± 11.02 c | 0.009 ± 0.0004 b |
| Ferulic acid | uncompetitive | 0.008 ± 0.0004 b | 455 ± 9.9 d | 0.027 ± 0.0013 a |
| Compound | IC50 [mM] |
|---|---|
| trans-cinnamic acid | 2.12 ± 0.106 a |
| ferulic acid | 0.41 ± 0.020 b |
| syringic acid | 0.22 ± 0.011 c |
| Compound | Ferulic Acid | trans–Cinnamic Acid | Syringic Acid |
|---|---|---|---|
| Ferulic acid | - | 0.98 ± 0.06 a Nearly additive | 0.24 ± 0.05 c Strong synergism |
| trans–cinnamic acid | 0.98 ± 0.06 a Nearly additive | - | 0.46 ± 0.04 b Synergism |
| Syringic acid | 0.24 ± 0.05 c Strong synergism | 0.46 ± 0.04 b Synergism | - |
| Compound | Ferulic Acid | trans–Cinnamic Acid | Syringic Acid |
|---|---|---|---|
| Ferulic acid | - | 0.53 ± 0.02 a Synergism | 0.48 ± 0.01 a Synergism |
| trans–cinnamic acid | 0.53 ± 0.02 a Synergism | - | 2.15 ± 0.08 b Antagonism |
| Syringic acid | 0.48 ± 0.01 a Synergism | 2.15 ± 0.08 b Antagonism | - |
| Compound | Ferulic Acid | trans–Cinnamic Acid | Syringic Acid |
|---|---|---|---|
| Ferulic acid | - | 0.63 ± 0.02 a Synergism | 2.21 ± 0.06 b Antagonism |
| trans–cinnamic acid | 0.63 ± 0.02 a Synergism | - | 0.69 ± 0.03 a Synergism |
| Syringic acid | 2.21 ± 0.06 b Antagonism | 0.69 ± 0.03 a Synergism | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habza-Kowalska, E.; Kaczor, A.A.; Bartuzi, D.; Piłat, J.; Gawlik-Dziki, U. Some Dietary Phenolic Compounds Can Activate Thyroid Peroxidase and Inhibit Lipoxygenase-Preliminary Study in the Model Systems. Int. J. Mol. Sci. 2021, 22, 5108. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105108
Habza-Kowalska E, Kaczor AA, Bartuzi D, Piłat J, Gawlik-Dziki U. Some Dietary Phenolic Compounds Can Activate Thyroid Peroxidase and Inhibit Lipoxygenase-Preliminary Study in the Model Systems. International Journal of Molecular Sciences. 2021; 22(10):5108. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105108
Chicago/Turabian StyleHabza-Kowalska, Ewa, Agnieszka A. Kaczor, Damian Bartuzi, Jacek Piłat, and Urszula Gawlik-Dziki. 2021. "Some Dietary Phenolic Compounds Can Activate Thyroid Peroxidase and Inhibit Lipoxygenase-Preliminary Study in the Model Systems" International Journal of Molecular Sciences 22, no. 10: 5108. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105108