
The Protective Effects of EMF-LTE against DNA Double-Strand Break Damage In Vitro and In Vivo

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
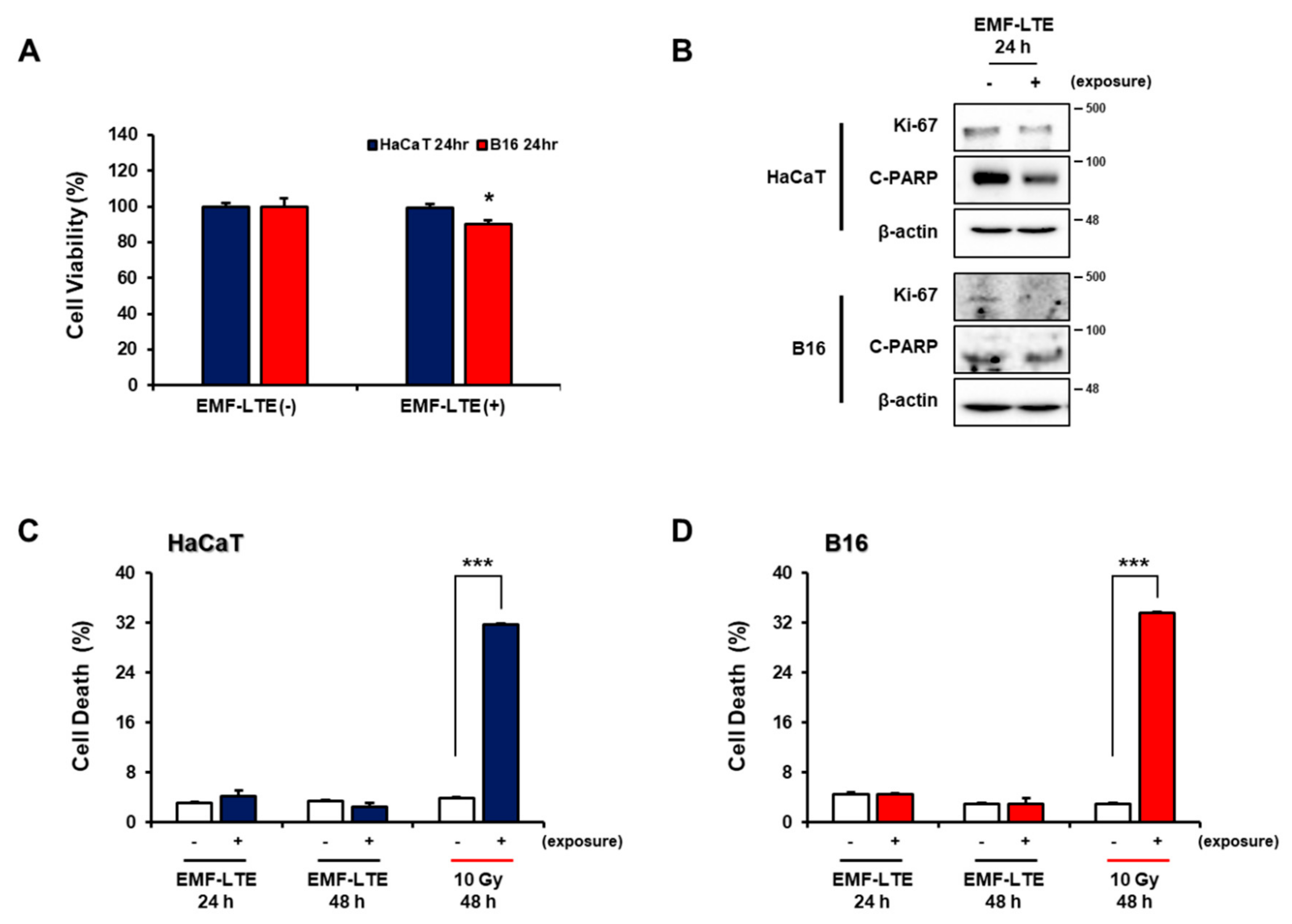
2.1. Effects of Electromagnetic Field-Long Term Evolution (EMF-LTE) on Cell Viability, Proliferation, Apoptosis, and Necrosis of Human Keratinocyte Cell Line (HaCaT) and Murine Melanoma Cell Line (B16) Cells
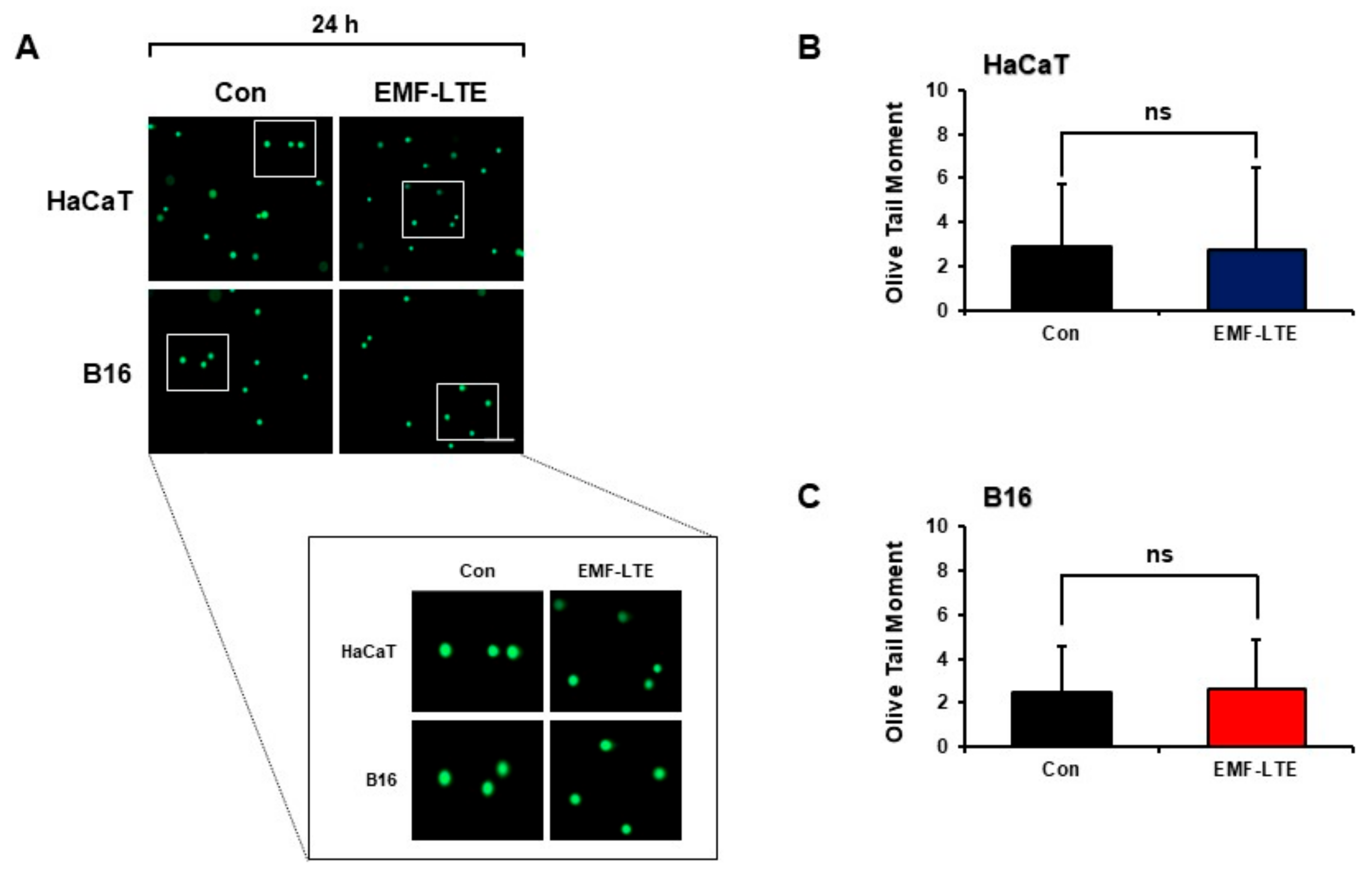
2.2. Effects of EMF-LTE on DNA Double-Strand Break Damage of HaCaT and B16 Cells
2.3. Effect of EMF-LTE on Ionizing Radiation (IR)- or Bleomycin (BLM)-Induced DNA Double-Strand Break (DSB) Damage
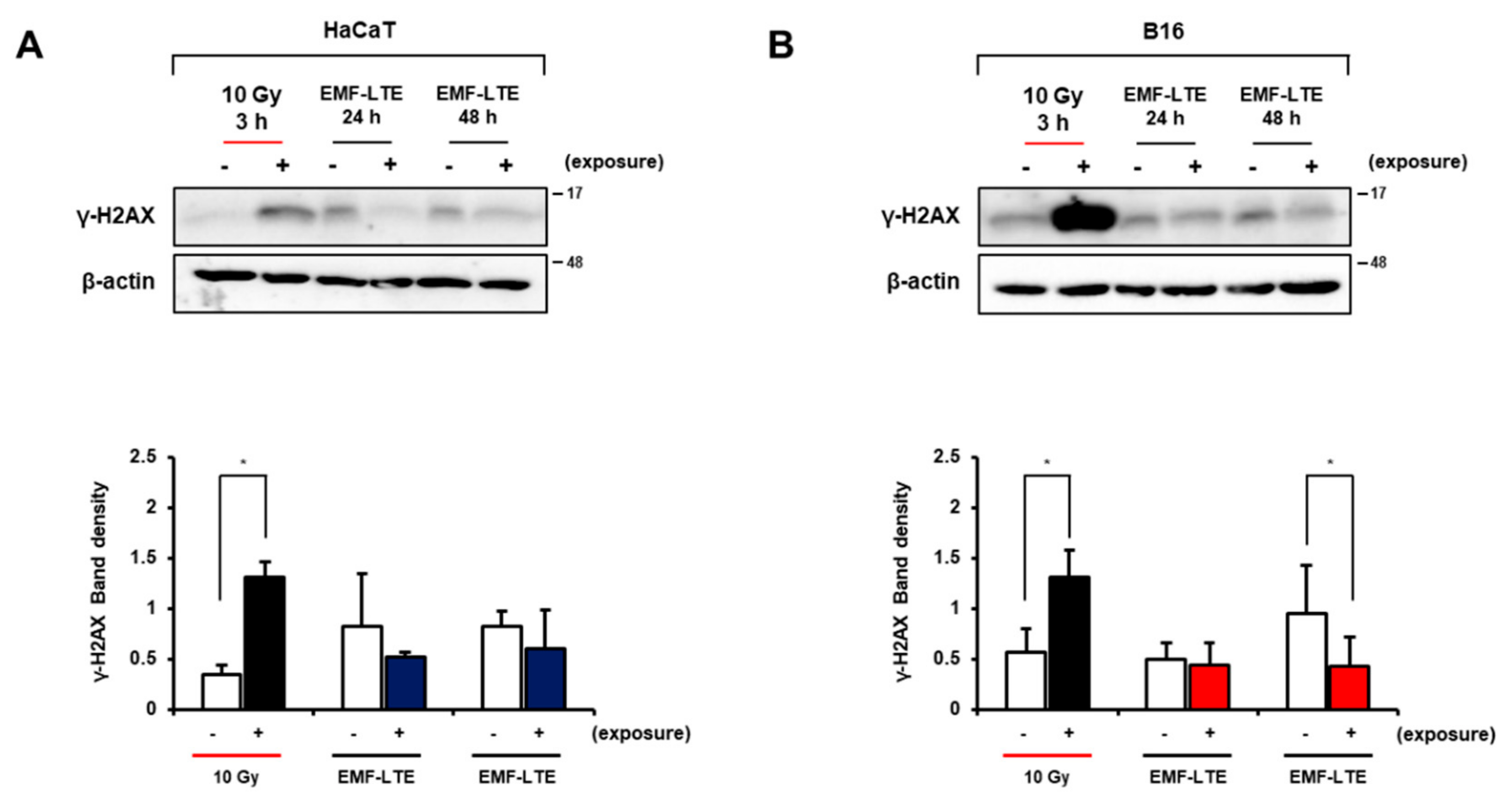
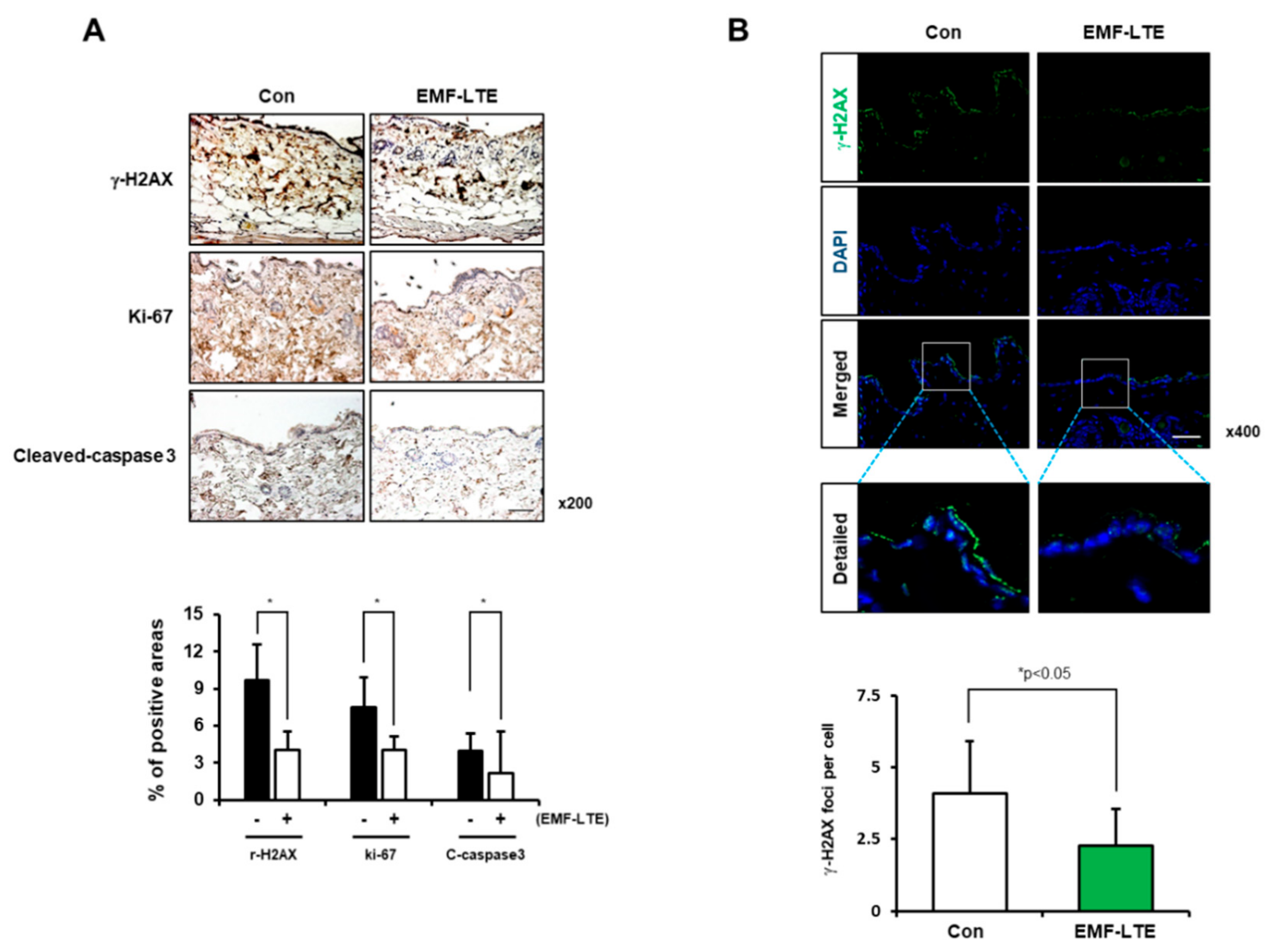
2.4. Effects of EMF-LTE on IR-Induced DNA DSB Damage Marker γ-H2AX in HaCaT and B16 Cells
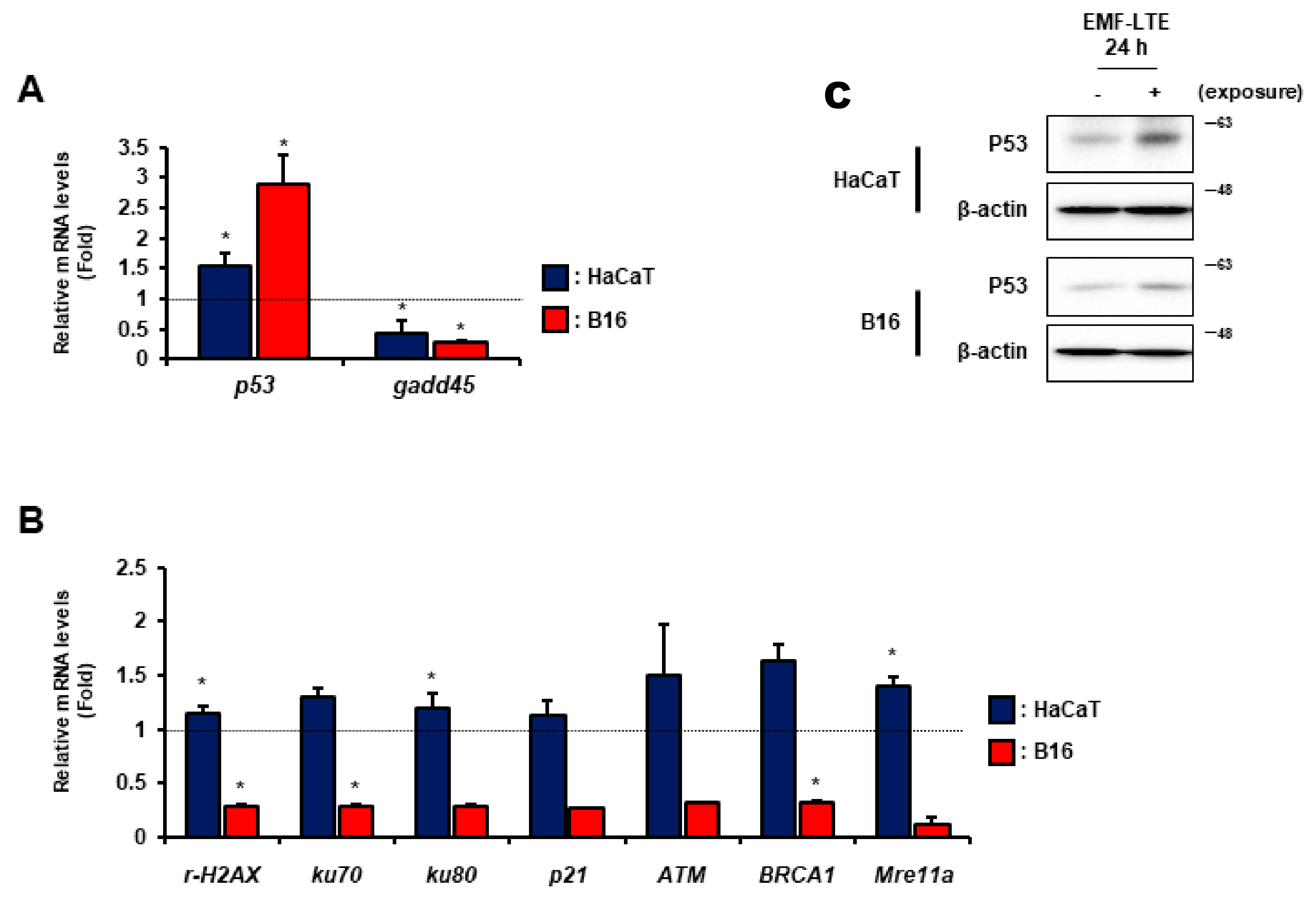
2.5. Effects of EMF-LTE on the Expression Levels of DNA Repair Genes
3. Discussion
4. Materials and Methods
4.1. In Vitro Exposure to EMF-LTE, IR, and Bleomycin
4.2. Cell Culture
4.3. MTT (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) Assay
4.4. Flow Cytometry Analysis
4.5. Neutral Comet Assay
4.6. Immunoblotting
4.7. Animals
4.8. Immunohistochemistry (IHC) and Immunofluorescence Staining (IF)
4.9. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| EMFs | Electromagnetic fields |
| LTE | Long term evolution |
| EMF-LTE | EMF-long term evolution |
| EMR | Electromagnetic radiation |
| IR | Ionizing radiation |
| NIR | Non-Ionizing radiation |
| ELF | Extremely low frequency |
| RF | Radio frequency |
| BER | Base excision repair |
| NER | Nucleotide excision repair |
| NHEJ | Non-homologous end-joining |
| MTT | [3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] |
References
- Adair, R.K. Biophysical Limits on Athermal Effects of RF and Microwave Radiation. J. Bioelectromagn. Soc. 2003, 24, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Adair, R.K. Vibrational Resonances in Biological Systems at Microwave Frequencies. Biophys. J. 2002, 82, 1147–1152. [Google Scholar] [CrossRef] [Green Version]
- Baan, R.; Grosse, Y.; Lauby-Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Islami, F.; Galichet, L.; Straif, K. Carcinogenicity of Radiofrequency Electromagnetic Fields. Lancet Oncol. 2011, 12, 624–626. [Google Scholar] [CrossRef]
- Baek, S.; Quan, X.; Kim, S.; Lengner, C.; Park, J.-K.; Kim, J. Electromagnetic Fields Mediate Efficient Cell Reprogramming into a Pluripotent State. ACS. Nano. 2014, 8, 10125–10138. [Google Scholar] [CrossRef]
- Phillips, J.L.; Singh, N.P.; Lai, H. Electromagnetic Fields and DNA Damage. Pathophysiology 2009, 16, 79–88. [Google Scholar] [CrossRef]
- Ruediger, H.W. Genotoxic Effects of Radiofrequency Electromagnetic Fields. Pathophysiology 2009, 16, 89–102. [Google Scholar] [CrossRef]
- Zhao, T.-Y.; Zou, S.-P.; Knapp, P.E. Exposure to Cell Phone Radiation Up-Regulates Apoptosis Genes in Primary Cultures of Neurons and Astrocytes. Neurosci. Lett. 2007, 412, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Johnson, D.; Dunbar, K.; Dong, H.; Ge, X.; Kim, Y.C.; Wing, C.; Jayathilaka, N.; Emmanuel, N.; Zhou, C.Q. 2.45 GHz Radiofrequency Fields Alter Gene Expression in Cultured Human Cells. FEBS Lett. 2005, 579, 4829–4836. [Google Scholar] [CrossRef] [Green Version]
- Demsia, G.; Vlastos, D.; Matthopoulos, D.P. Effect of 910-MHz Electromagnetic Field on Rat Bone Marrow. Sci. World J. 2004, 4, 48–54. [Google Scholar] [CrossRef]
- Lai, H.; Singh, N.P. Magnetic-Field-Induced DNA Strand Breaks in Brain Cells of the Rat. Environ. Health Perspect. 2004, 112, 687–694. [Google Scholar] [CrossRef] [Green Version]
- Harth, Y. Painless, Safe, and Efficacious Noninvasive Skin Tightening, Body Contouring, and Cellulite Reduction Using Multisource 3DEEP Radiofrequency. J. Cosmet. Dermatol. 2015, 14, 70–75. [Google Scholar] [CrossRef]
- Kwan, K.R.; Kolansky, Z.; Abittan, B.J.; Farberg, A.S.; Goldenberg, G. Skin Tightening. Cutis 2020, 106, 134–137, 139, E1. [Google Scholar] [CrossRef]
- Yao, K.; Wu, W.; Wang, K.; Ni, S.; Ye, P.; Yu, Y.; Ye, J.; Sun, L. Electromagnetic Noise Inhibits Radiofrequency Radiation-Induced DNA Damage and Reactive Oxygen Species Increase in Human Lens Epithelial Cells. Mol. Vis. 2008, 14, 964. [Google Scholar]
- Çam, S.T.; Seyhan, N. Single-Strand DNA Breaks in Human Hair Root Cells Exposed to Mobile Phone Radiation. Int. J. Radiat. Biol. 2012, 88, 420–424. [Google Scholar] [CrossRef]
- Falzone, N.; Huyser, C.; Franken, D.R.; Leszczynski, D. Mobile Phone Radiation Does Not Induce Pro-Apoptosis Effects in Human Spermatozoa. Radiat. Res. 2010, 174, 169–176. [Google Scholar] [CrossRef]
- Maes, A.; Van Gorp, U.; Verschaeve, L. Cytogenetic Investigation of Subjects Professionally Exposed to Radiofrequency Radiation. Mutagenesis 2006, 21, 139–142. [Google Scholar] [CrossRef] [Green Version]
- Verschaeve, L.; Heikkinen, P.; Verheyen, G.; Van Gorp, U.; Boonen, F.; Vander Plaetse, F.; Maes, A.; Kumlin, T.; Mäki-Paakkanen, J.; Puranen, L. Investigation of Co-Genotoxic Effects of Radiofrequency Electromagnetic Fields in Vivo. Radiat. Res. 2006, 165, 598–607. [Google Scholar] [CrossRef]
- Mahmoudinasab, H.; Sanie-Jahromi, F.; Saadat, M. Effects of Extremely Low-Frequency Electromagnetic Field on Expression Levels of Some Antioxidant Genes in Human MCF-7 cells. Mol. Biol. Res. Commun. 2016, 5, 77. [Google Scholar]
- Sanie-Jahromi, F.; Saadat, M. Effects of Electromagnetic Field, Cisplatin and Morphine on Cytotoxicity and Expression Levels of DNA Repair Genes. Mol. Biol. Rep. 2018, 45, 807–814. [Google Scholar] [CrossRef]
- Robison, J.G.; Pendleton, A.R.; Monson, K.O.; Murray, B.K.; O’Neill, K.L. Decreased DNA Repair Rates and Protection from Heat Induced Apoptosis Mediated by Electromagnetic Field Exposure. Bioelectromagn. Soc. Phys. Regul. Biol. Med. Eur. Bioelectromagn. Assoc. 2002, 23, 106–112. [Google Scholar] [CrossRef]
- Proksch, E.; Brandner, J.M.; Jensen, J.M. The skin: An Indispensable Barrier. Exp. Dermatol. 2008, 17, 1063–1072. [Google Scholar] [CrossRef]
- Marks, J.G.; Miller, J.J. Lookingbill and Marks’ Principles of Dermatology E-Book; Elsevier Health Sciences: New York, NY, USA, 2017. [Google Scholar]
- Shin, D.W. Various Biological Effects of Solar Radiation on Skin and Their Mechanisms: Implications for Phototherapy. Anim. Cells Syst. 2020, 24, 181–188. [Google Scholar] [CrossRef]
- Gupta, A.; Dai, T.; Hamblin, M.R. Effect of Red and Near-Infrared Wavelengths on Low-Level Laser (light) Therapy-Induced Healing of Partial-Thickness Dermal Abrasion in Mice. Lasers Med. Sci. 2014, 29, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J.; Kim, H.J.; Kim, H.L.; Kim, H.J.; Kim, H.S.; Lee, T.R.; Shin, D.W.; Seo, Y.R. A Protective Mechanism of Visible Red Light in Normal Human Dermal Fibroblasts: Enhancement of GADD45A-Mediated DNA Repair Activity. J. Investig. Dermatol. 2017, 137, 466–474. [Google Scholar] [CrossRef] [Green Version]
- Yoon, K.; Choi, S.; Choi, H.-D.; Kim, N.; Jeon, S.B.; Lim, K.-M.; Lee, H.-J.; Lee, Y.-S. Effects of Radiofrequency Electromagnetic Fields and Ionizing Radiation on Amyloid Precursor Protein Processing and Cell Death. J. Electromagn. Eng. Sci. 2020, 20, 307–319. [Google Scholar] [CrossRef]
- Tillmann, T.; Ernst, H.; Streckert, J.; Zhou, Y.; Taugner, F.; Hansen, V.; Dasenbrock, C. Indication of Cocarcinogenic Potential of Chronic UMTS-Modulated Radiofrequency Exposure in an Ethylnitrosourea Mouse Model. Int. J. Radiat. Biol. 2010, 86, 529–541. [Google Scholar] [CrossRef]
- Lee, H.J.; Jin, Y.B.; Lee, J.S.; Choi, J.I.; Lee, J.W.; Myung, S.H.; Lee, Y.S. Combined Effects of 60 Hz Electromagnetic Field Exposure with Various Stress Factors on Cellular Transformation in NIH3T3 Cells. Bioelectromagnetics 2012, 33, 207–214. [Google Scholar] [CrossRef]
- Yoon, H.E.; Lee, J.S.; Myung, S.H.; Lee, Y.-S. Increased γ-H2AX by Exposure to a 60-Hz Magnetic Fields Combined with Ionizing Radiation, but Not Hydrogen Peroxide, in Non-Tumorigenic Human Cell Lines. Int. J. Radiat. Biol. 2014, 90, 291–298. [Google Scholar] [CrossRef]
- Cho, Y.H.; Jeon, H.K.; Chung, H.W. Effects of Extremely Low-Frequency Electromagnetic Fields on Delayed Chromosomal Instability Induced by Bleomycin in Normal Human Fibroblast Cells. J. Toxicol. Environ. Health 2007, 70, 1252–1258. [Google Scholar] [CrossRef]
- Dorr, R.T. Bleomycin Pharmacology: Mechanism of Action and Resistance, and Clinical Pharmacokinetics. Semin. Oncol. 1992, 19, 3–8. [Google Scholar]
- Lerchl, A.; Klose, M.; Drees, K. No Increased DNA Damage Observed in the Brain, Liver, and Lung of Fetal Mice Treated with Ethylnitrosourea and Exposed to UMTS Radiofrequency Electromagnetic Fields. Bioelectromagnetics 2020, 41, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Al-Serori, H.; Ferk, F.; Kundi, M.; Bileck, A.; Gerner, C.; Mišík, M.; Nersesyan, A.; Waldherr, M.; Murbach, M.; Lah, T.T. Mobile Phone Specific Electromagnetic Fields Induce Transient DNA Damage and Nucleotide Excision Repair in Serum-Deprived Human Glioblastoma Cells. PLoS ONE 2018, 13, e0193677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, C.; Wu, Z.; Eastman, A.; Bresnick, E. Irradiation-Induced Expression of O6-Methylguanine-DNA Methyltransferase in Mammalian Cells. Cancer Res. 1992, 52, 1804–1809. [Google Scholar] [PubMed]
- Lerchl, A.; Krüger, H.; Niehaus, M.; Streckert, J.R.; Bitz, A.K.; Hansen, V. Effects of Mobile Phone Electromagnetic Fields at Nonthermal SAR Values on Melatonin and Body Weight of Djungarian Hamsters (Phodopus sungorus). J. Pineal Res. 2008, 44, 267–272. [Google Scholar] [CrossRef]
- Szudziński, A.; Pietraszek, A.; Janiak, M.; Wrembel, J.; Kałczak, M.; Szmigielski, S. Acceleration of the Development of Benzopyrene-Induced Skin Cancer in Mice by Microwave Radiation. Arch. Dermatol. Res. 1982, 274, 303–312. [Google Scholar] [CrossRef]
- Chagnaud, J.L.; Moreau, J.M.; Veyret, B. No Effect of Short-Term Exposure to GSM-Modulated Low-Power Microwaves on Benzo(a)Pyrene-Induced Tumours in Rat. Int. J. Radiat. Biol. 1999, 75, 1251–1256. [Google Scholar] [CrossRef]
- Choi, J.; Min, K.; Jeon, S.; Kim, N.; Pack, J.K.; Song, K. Continuous Exposure to 1.7 GHz LTE Electromagnetic Fields Increases Intracellular Reactive Oxygen Species to Decrease Human Cell Proliferation and Induce Senescence. Sci. Rep. 2020, 10, 9238. [Google Scholar] [CrossRef]
- Nikolova, T.; Czyz, J.; Rolletschek, A.; Blyszczuk, P.; Fuchs, J.; Jovtchev, G.; Schulderer, J.; Kuster, N.; Wobus, A.M. Electromagnetic Fields Affect Transcript Levels of Apoptosis-Related Genes in Embryonic Stem Cell-Derived Neural Progenitor Cells. FASEB J. 2005, 19, 1686–1688. [Google Scholar] [CrossRef] [Green Version]
- Baharara, J.; Hosseini, N.; Farzin, T.R. Extremely low Frequency Electromagnetic Field Sensitizes Cisplatin-Resistant Human Ovarian Adenocarcinoma Cells Via P53 Activation. Cytotechnology 2016, 68, 1403–1413. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.J.; Kim, E.; Mun, J.; Park, S.; Smith, M.; Han, S.S.; Seo, Y. Base Excision DNA Repair Defect in Gadd45a-Deficient Cells. Oncogene 2007, 26, 7517–7525. [Google Scholar] [CrossRef] [Green Version]
- Banath, P.; Fushiki, M.; Olive, P. Rejoining of DNA single-And Double-Strand Breaks in Human White Blood Cells Exposed to Ionizing Radiation. Int. J. Radiat. Biol. 1998, 73, 649–660. [Google Scholar] [CrossRef]
- Jeong, Y.J.; Son, Y.; Choi, H.D.; Kim, N.; Lee, Y.S.; Ko, Y.G.; Lee, H.J. Behavioral Changes and Gene Profile Alterations after Chronic 1,950-MHz Radiofrequency Exposure: An Observation in C57BL/6 Mice. Brain Behav. 2020, 10, e01815. [Google Scholar] [CrossRef]
- Son, Y.; Kim, J.S.; Jeong, Y.J.; Jeong, Y.K.; Kwon, J.H.; Choi, H.D.; Pack, J.K.; Kim, N.; Lee, Y.S.; Lee, H.J. Long-Term RF Exposure on Behavior and Cerebral Glucose Metabolism in 5xFAD mice. Neurosci. Lett. 2018, 666, 64–69. [Google Scholar] [CrossRef]
- IEC 61000-4-21. Electromagnetic Compatibility (EMC)–Part 4–21: Testing and Measurement Techniques–Reverberation Chamber Test Methods; IEEE: New York, NY, USA, 2011. [Google Scholar]
- Jung, K.B.; Kim, T.H.; Kim, J.L.; Doh, H.J.; Chung, Y.C.; Choi, J.H.; Pack, J.K. Development and Validation of Reverberation-Chamber Type Whole-Body Exposure System for Mobile-Phone Frequency. Electromagn. Biol. Med. 2008, 27, 73–82. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence (Human) | Sequence (Mouse) | |
|---|---|---|---|
| 1 | p53 | Forward: AGGCCTTGGAACTCAAGGAT | Forward: CACGTACTCTCCTCCCCTCAAT |
| Reverse: TGAGTCAGGCCCTTCTGTCT | Reverse: AACTGCACAGGGCACGTCTT | ||
| 2 | r-H2AX | Forward: TGGAAAGGGTCAGGGAACG | Forward: AACGACGAGGAGCTCAACAAGC |
| Reverse: GACTTGTGCTGGTATCTGGGTG | Reverse: TGGCGCTGCTCTTCTTGGGCA | ||
| 3 | ku80 | Forward: GTTCTAAAGGTCTTTGCAGCAAGA | Forward: AACATGGTCGCCATCGTCCGAT |
| Reverse: AAAAGCCACGCCGACTTGAGGA | Reverse: CCGCAAGTCTTCCATGAAAGGC | ||
| 5 | ku70 | Forward: GGTTTCAAGCCGTTGGTACTGC | Forward: GCAGTCTACTCCTGCCTAGTGA |
| Reverse: CTCCAGACACTTGATGAGCAGAG | Reverse: ACCTGGCTCATCAAACCGCTTC | ||
| 7 | gadd45 | Forward: CGTTTTGCTGCGAGAACGAC | Forward: CCTGGAGGAAGTGCTCAGCAAG |
| Reverse: GAACCCATTGATCCATGTAG | Reverse: GTCGTCTTCGTCAGCAGCCAG | ||
| 8 | p21 | Forward: GCTTCATGCCAGCTACTTCC | Forward: TCGCTGTCTTGCACTCTGGTGT |
| Reverse: CCCTTCAAAGTGCCATCTGT | Reverse: CCAATCTGCGCTTGGAGTGATAG | ||
| 9 | ATM | Forward: TGTTCCAGGACACGAAGGGAGA | Forward: CCAAGATGGCAGTGAACCAGAC |
| Reverse: CAGGGTTCTCAGCACTATGGGA | Reverse: ATGCTGGACAGCTATGGTGGAG | ||
| 10 | BRCA1 | Forward: CTGAAGACTGCTCAGGGCTATC | Forward: CGAGGAAATGGCAACTTGCCTAG |
| Reverse: AGGGTAGCTGTTAGAAGGCTGG | Reverse: TCACTCTGCGAGCAGTCTTCAG | ||
| 13 | Mre11a | Forward: CAGCAACCAACAAAGGAAGAGGC | Forward: CCCTGACAATCCTAAGGTGACC |
| Reverse: GAGTTCCTGCTACGGGTAGAAG | Reverse: CGTAGTCGGATAAGAGGCTTCC | ||
| 14 | gapdh | Forward: TTCGACAGTCAGCCGCATCTTCTT | Forward: ACTGTGGTCATGAGCCCTTC |
| Reverse: GCCCAATACGACCAAATCCGTTGA | Reverse: GGGTGTGAACCACGAGAAAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, H.; Kim, K.; Park, G.-Y.; Kim, M.; Lee, H.-J.; Jeon, S.; Kim, J.H.; Kim, H.R.; Lim, K.-M.; Lee, Y.-S. The Protective Effects of EMF-LTE against DNA Double-Strand Break Damage In Vitro and In Vivo. Int. J. Mol. Sci. 2021, 22, 5134. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105134
Jin H, Kim K, Park G-Y, Kim M, Lee H-J, Jeon S, Kim JH, Kim HR, Lim K-M, Lee Y-S. The Protective Effects of EMF-LTE against DNA Double-Strand Break Damage In Vitro and In Vivo. International Journal of Molecular Sciences. 2021; 22(10):5134. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105134
Chicago/Turabian StyleJin, Hee, Kyuri Kim, Ga-Young Park, Minjeong Kim, Hae-June Lee, Sangbong Jeon, Ju Hwan Kim, Hak Rim Kim, Kyung-Min Lim, and Yun-Sil Lee. 2021. "The Protective Effects of EMF-LTE against DNA Double-Strand Break Damage In Vitro and In Vivo" International Journal of Molecular Sciences 22, no. 10: 5134. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105134