
Delivery Systems of Retinoprotective Proteins in the Retina
by

Ivan T. Rebustini
*, 
Alexandra Bernardo-Colón
, 
Alejandra Isasi Nalvarte
and
S. Patricia Becerra
* Section of Protein Structure and Function, LRCMB-NEI-NIH, Bethesda, MD 20892-0608, USA
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(10), 5344; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105344
Submission received: 9 April 2021
/
Revised: 12 May 2021
/
Accepted: 17 May 2021
/
Published: 19 May 2021
(This article belongs to the Special Issue Retinal Degeneration: Molecular Mechanism, Pathogenesis and Treatment)
Abstract
:Retinoprotective proteins play important roles for retinal tissue integrity. They can directly affect the function and the survival of photoreceptors, and/or indirectly target the retinal pigment epithelium (RPE) and endothelial cells that support these tissues. Retinoprotective proteins are used in basic, translational and in clinical studies to prevent and treat human retinal degenerative disorders. In this review, we provide an overview of proteins that protect the retina and focus on pigment epithelium-derived factor (PEDF), and its effects on photoreceptors, RPE cells, and endothelial cells. We also discuss delivery systems such as pharmacologic and genetic administration of proteins to achieve photoreceptor survival and retinal tissue integrity.
1. Introduction
The neural retina, composed of layers of photoreceptors, horizontal cells, bipolar cells, Müller cells, amacrine cells and retinal ganglion cells, converts light stimulation into chemical signals sent through the optic nerve into the brain [1]. The crucial process of visual phototransduction occurs in the photoreceptors, which are highly specialized and terminally differentiated cells, and involves light-sensing opsins. In most species of vertebrates, adult photoreceptors do not proliferate and finish their differentiation at early postnatal stages [2]. Damage of photoreceptors and their supportive tissues, such as the retinal pigment epithelial (RPE) cells, causes multiple forms of retinal pathogenesis [3]. Retinoprotective proteins can prevent cell death of photoreceptors and RPE cells while maintaining cellular functions [4]. Another important role of retinoprotective proteins includes inhibiting angiogenesis that dysregulates blood vessels in the retina and generates pathological phenotypes observed in retinal degeneration [5].
In this review, we offer an overview of retinoprotective proteins that target retinopathies including their mechanisms of action (Section 2), and focus on pigment epithelium derived factor (PEDF, Section 3). We also describe systems of local delivery of PEDF into the retina (Section 4): (1) pharmacologic administration of PEDF (recombinantly produced proteins, extracted and purified from bacterial and mammalian cells) or their derived peptides (chemically synthesized peptide fragments from larger, modular proteins containing biologically active regions); (2) genetic administration of the Serpinf1 gene achieved by infections or transfections of expression vectors encoding for PEDF in the retinal or in the RPE tissues in vivo; and (3) Cell-based systems in which PEDF-expressing cells are encapsulated in vitro and transplanted into the retina in vivo for cell therapies are also discussed.
2. Overview of Retinoprotective Proteins
Retinoprotective proteins function either via autocrine signaling (when expressed and secreted by the same cells that they target), paracrine signaling (when expressed by supporting tissues such as the RPE to target photoreceptors) or endocrine signaling (when delivered from distal tissues to the photoreceptors by the choroidal and the inner retinal vasculature) [4]. Representative examples of retinoprotective proteins are briefly discussed and summarized in the Table 1.
2.1. Autocrine Retinoprotective Proteins
Naturally occurring proteins that act in an autocrine fashion during retinal development can be administered postnatally as retinoprotective agents. Among the most studied of these proteins include Insulin-like Growth Factor 1 (IGF1, [6]), Fibroblast Growth Factor 2 (FGF2, [7]), Ciliary Neurotrophic Factor (CNTF, [8]), Brain-Derived Neurotrophic Factor (BDNF, [9]), Nerve Growth Factor (NGF, [10]) and Vascular Endothelial Growth Factor (VEGF, [11]). The re-induction of IGF1 [18,19] and FGF2, which is retinoprotective to photoreceptors by regulating Ca2+ influx in a model of retinitis pigmentosa [7] are examples of retinoprotective proteins that are developmentally regulated in the retina, and can contribute to adult photoreceptor homeostasis when pharmacologically or genetically delivered in the retina. Studies with cntf KO mouse reveal that CNTF can protect cones and rods by suppressing excessive formation of visual pigments [8]. Recombinant human NGF promotes photoreceptor survival in a rat model of retinitis pigmentosa [10]. An example of the application of autocrine retinoprotective proteins for retinal protection is the use of 7,8-dihydroxyflavone hydrate (7,8-DHF), which is a drug that is a mimetic to BDNF [9], in combination with histone deacetylase inhibitors (HDACi) currently in clinical trial for the treatment of inherited retinal degeneration. The individual use or the combination of the BDNF mimetic and HDACi are capable to restore photoreceptors visual function in a zebrafish dye ucd6 blindness model [9]. An overview of representative autocrine retinoprotective proteins is summarized in the Table 1.
2.2. Paracrine Retinoprotective Proteins
Representative examples of paracrine retinoprotective proteins include the pituitary adenylate cyclase-activating polypeptide (PACAP) and vasoactive intestinal peptide (VIP, [12]), somatostatin [20], and Vascular Endothelial Growth Factor (VEGF, [11]). VIP is expressed in a population of amacrine cells [13], whereas PACAP and its receptors (PAC1, VPAC1, and VPAC2) occur in all retinal layers except the photoreceptors [14]. Topically administered PACAP and VIP derivatives in the form of eye drops attenuate ischemic retinal degeneration [21]. Of note, IGF1, previously mentioned above as possessing an autocrine signaling mechanism of action, can also act in a paracrine manner (Table 1), as it is expressed by the microglia and protects photoreceptors in the rd10 mouse model [6]. The paracrine retinoprotective peptide somatostatin or somatotropin release inhibiting factor (SRIF), is a neuropeptide with a broad inhibitory effect on protein secretion, proliferation, and angiogenesis [20]. The SRIF receptors (SST1-4 in mammals) are distributed in all retinal layers and RPE cells, but the main source of secreted SRIF is the RPE cells [14]. SRIF, via its receptor SSTR2, protects the retina by limiting vascular endothelial growth VEGF, which is normally secreted at the basolateral side of RPE cells, but light-induced damage changes the VEGF secretion to the apical side of RPE cells causing blood-retinal-barrier breakdown and irreversible photoreceptor damage [11]. This indicates that anti-VEGF compounds are of interest therapeutically for retina altered by stress.
2.3. Endocrine Retinoprotective Proteins
Finally, in addition to autocrine and paracrine mechanisms of action, retinoprotective proteins produced and secreted by distal tissues can be transported by the vasculature system to photoreceptors, which characterizes an endocrine signaling function. The deep retinal vessels and the choroidal vessels are the natural delivery ports of retinoprotective proteins from distal tissues to the photoreceptors and the RPE cells [22]. Retinoprotective proteins with endocrine signaling function include those in the proinsulin system, which is one of the firsts to be identified in the retina and protect photoreceptors [23]. All retinal cell layers are insulin-sensitive and express a variety of insulin receptors, however, insulin production within the retina has not been described. Proinsulin and insulin are attenuators of cell death in the developing and adult nervous system [16]. Transgenic expression of human proinsulin in skeletal muscle in the rd10 mouse model of Retinitis Pigmentosa [17] and in the P23H rat model of autosomal dominant Retinitis Pigmentosa [16] delay the death of photoreceptors, prolongs visual function, and attenuates retinal degeneration. Systemic administration of insulin in four mouse models harboring mutations in rod-specific genes (Pde6b−/−, Pde6g−/−, Rho−/− and P23H) prolong photoreceptor survival, whereas depletion of endogenous insulin in these mouse models has the opposite effect [17], exemplifying the importance of the insulin signaling in retinoprotection.
3. PEDF
3.1. Biogenesis
Among the retinoprotective proteins of interest in ocular research, pigment epithelium-derived factor (PEDF) is relevant due to its well-established protective properties in the retina, in particular the photoreceptors [24,25], RPE cells [26] and in preventing neovascularization of the choroid and retina [27]. A variety of tissues express the gene Serpinf1 for PEDF, [25] which is altered in retinopathies and in tumor tissues as well [28]. In the eye, the Serpinf1 gene is transcriptionally active in RPE cells but not detected in the photoreceptors [29]. Both RPE cells and photoreceptors express the gene Pnpla2 (patatin-like phospholipase domain-containing protein for the PEDF receptor (PEDF-R), a membrane linked enzyme that catalyzes the hydrolysis of phospholipids to release fatty acids and lysophospholipids in retinal cells [30,31].
3.2. Structure
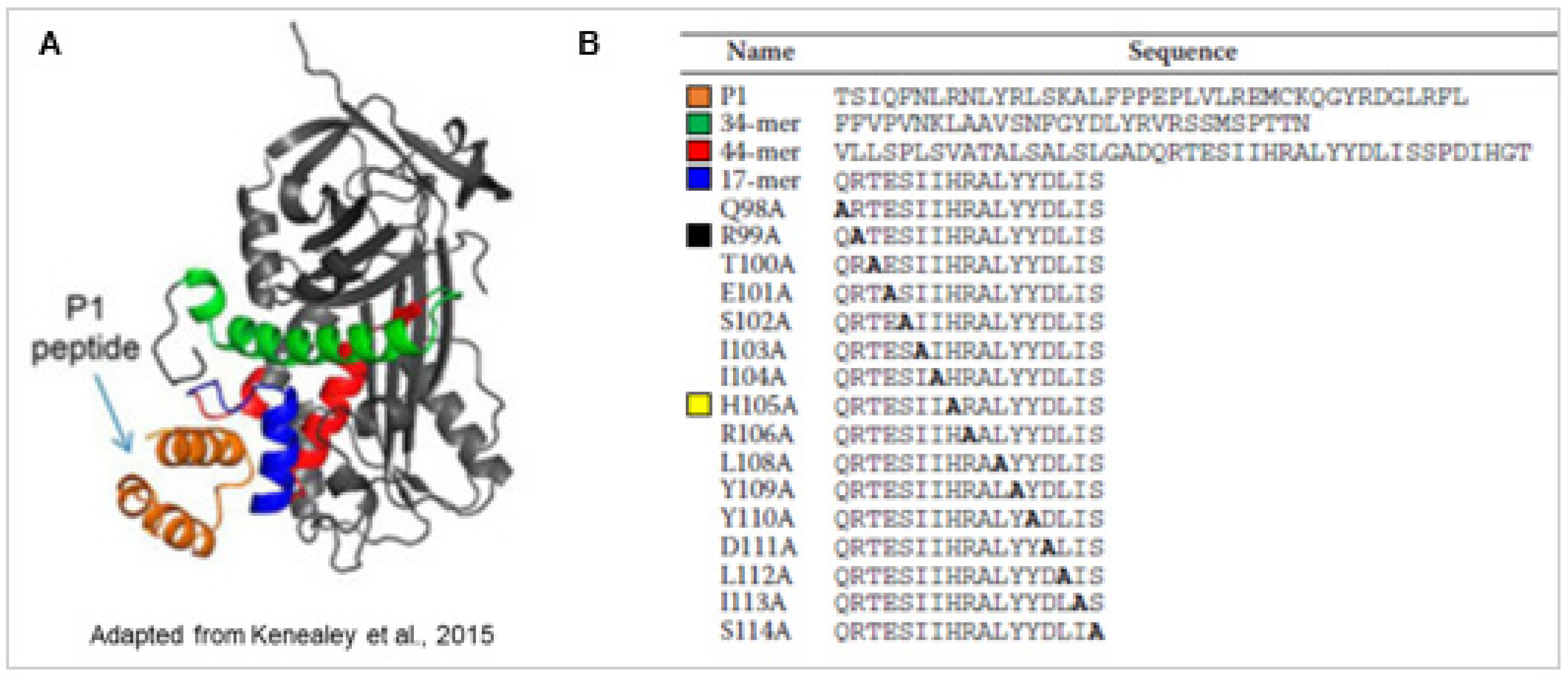
PEDF is a 50 kDa glycoprotein and a member of the serine protease inhibitor (serpin) superfamily. PEDF lacks the serine protease inhibitory activity and is secreted extracellularly, binds PEDF-R with high affinity (Kd = 3 nM) and has cytoprotective activity [30]. PEDF-R is a critical receptor for the survival activity of PEDF. PEDF binding to PEDF-R stimulates the PEDF-R enzymatic activity, and peptides derived from the ligand binding region of the receptor blocks the PEDF·PEDF-R interactions and their mediated retinal survival activities. We have mapped the ligand binding domain (LBD, residue positions T210-L232) in human PEDF-R and termed a blocking peptide derived from this region P1 peptide, T210-L249) [32]. The receptor-binding domain of human PEDF is also identified and termed 17-mer peptide (residue positions Q98–S114). P1 peptide docks to a cleft that contains a solvent-exposed region corresponding to α-helix C within the residues Q98–S114 (17-mer) of the three-dimensional structure of the human PEDF protein [33] (Figure 1). The chemically synthesized 17-mer peptide from the solvent-exposed region of PEDF binds to PEDF-R and exhibits retinoprotective activity [33] (Figure 1). An alanine scanning approach of the 17-mer peptide reveals key interacting residues for PEDF-R binding. The 17-mer peptide with an amino acid alteration at the position 105 from histidine to alanine, 17-mer[H105A], exhibits higher photoreceptor protection efficacy than the unaltered 17-mer, which a blocking peptide P1 and atglistatin, a PEDF-R inhibitor, abolishes. Both the 17-mer and the 17-mer[H105A] peptides protect rd1 photoreceptors from death in vivo [34]. The recombinant full length PEDF with the H105A amino acids substitution also exhibits affinity for PEDF-R and survival activities to R28 cells and photoreceptors similar and lower than those of their peptide counterparts and unmodified PEDF [33].
A comparative study using another serpin, protease nexin-1 (PN-1) shows that its homologous 17-mer region protects R28 retinal cells against serum-starvation-induced cells death in vitro, like full length PN-1, PEDF and the PEDF 17-mer peptides [35]. This observation implies that PEDF serves a model to search for small, pharmacologic peptide candidates in other proteins of the serpin family, such as PN-1. Given that the serpin PN-1 inhibits serine protease thrombin and that the region to inhibit this protease is not present in the 17-mer, it indicates that inhibition of proteases by PN-1 is dispensable for cytoprotective activity. In addition, the PN-1 variant that lacks inhibitory activity due to an alteration R346A in the inhibitory site does not lack cytoprotective activity. This further implies that PN-1 mechanism of cytoprotection is independent of serine protease inhibition like PEDF [35].
Important interactions of PEDF with the extracellular matrix (ECM) components such as collagens occur [36]. The crystal structure of PEDF in a complex with a disulfide cross-linked heterotrimeric type I collagen peptide shows that PEDF specifically interacts with an amphiphilic sequence of the type I collagen in the ECM. This indicates that collagen I in the ECM functions as an extracellular reservoir of PEDF that is slowly and temporally released by the ECM remodeling such as collagen turnover by metalloproteinases [37].
3.3. Mechanisms of Action
PEDF and the PEDF peptides 17-mer and 17-mer[H105A] protect photoreceptors from damage in the rd1 and rd10 mouse models of retinal degeneration [29,34], in the Mitf knockout mouse model of microphtalmia [38], and a focal light-emitting diode (LED)-induced phototoxicity mouse model [39]. These effects are mediated by binding of the extracellular PEDF to the cell surface PEDF-R and promoting intracellular signaling cascade that ends in photoreceptor cell survival. The direct intracellular targets of PEDF-R upon PEDF binding are beginning to be elucidated. The PEDF/PEDF-R axis protects photoreceptors of the rd1 mouse model by reducing intracellular calcium in the degenerating retina and engaging calpains, BAX and AIF proteins [40]. Other pathways described using models of retinoprotection in vitro have been reviewed elsewhere [25] and include PI3K/AKT, ERK1/2 and the docosahexaenoic acid (DHA)/neuroprotectin D1 (ND-1) signaling pathways that are also downstream effectors of the PEDF/PEDF-R complex.
The generation of the Serpinf1 knockout mouse is useful to address mechanisms of action for PEDF. The Serpinf1 knockout in a rd10 mouse background shows that the absence of PEDF affects the magnitude of photoreceptor degeneration [29]. These findings highlight the importance of PEDF as a natural signaling protein in the retina to mitigate an undergoing degenerative process in vivo.
Besides the retina, it is worth mentioning the neurotrophic properties of PEDF in other tissues. PEDF combined with DHA promotes corneal nerve regeneration and upregulates the expression of the retinoprotective genes Vip, Bdnf and Ngf (retinoprotective proteins discussed above) and axon growth protein semaphorin 7a (Sema7a) during the corneal nerve regeneration [41]. Administration of the 44-mer PEDF peptide to the cornea of rabbits also induces the corneal nerve regeneration following injury created by corneal epithelial debridement [42].
3.4. Regulation of PEDF Expression
In murine eyes with retinal degeneration, PEDF levels are lower than in their wild type counterparts [34,43], corroborating the fact that PEDF levels are also decreased in eyes affected by retinal degenerative processes in humans [43,44,45,46]. The levels of PEDF in eyes of patients with retinitis pigmentosa are significantly lower than those in eyes without this disease [46]. The vitreous of patients with choroidal neovascularization due to age-related macular degeneration contains lower PEDF levels and lacks the antiangiogenic activity of vitreous from age-matched controls [44]. These observations suggest that loss of PEDF creates a permissive environment for retinal degeneration and ocular neovascularization.
Engineering PEDF-expressing RPE cells for cell therapies can enhance PEDF production in the retina. The apical secretion of PEDF by RPE cells [47] can serve to inhibit invasive choroidal neovascularization into the posterior retina and in turn maintain its avascularity and structural integrity. PEDF production and apical secretion by RPE cells are also important phenotypic markers for the differentiated and polarized status of RPE cells in vitro, as shown with ARPE-19 cells [48] and RPE cells derived from human pluripotent stem cells [49].
Furthermore, a recent study using a rat NaIO3-induced retinal degeneration model provides evidence of epigenetic regulatory mechanisms based on micro-RNA biology for PEDF expression in the RPE cells. The expression of the micro-RNA miR-25, which contains a seeding region that directly binds to PEDF mRNA and prevents its translation, is upregulated after induction of retinal degeneration [50]. PEDF mRNA is also a direct target for the micro-RNA miR-320c [51]. This micro-RNA is upregulated in nasopharyngeal carcinoma cells and is involved in decreasing PEDF expression and epithelial-to-mesenchymal transition that contributes to tumor metastasis [51]. This new regulatory level by miR-320c needs to be confirmed in the retina and exemplifies the increasing relevance of epigenetic events in regulating PEDF expression.
4. PEDF Delivery Systems
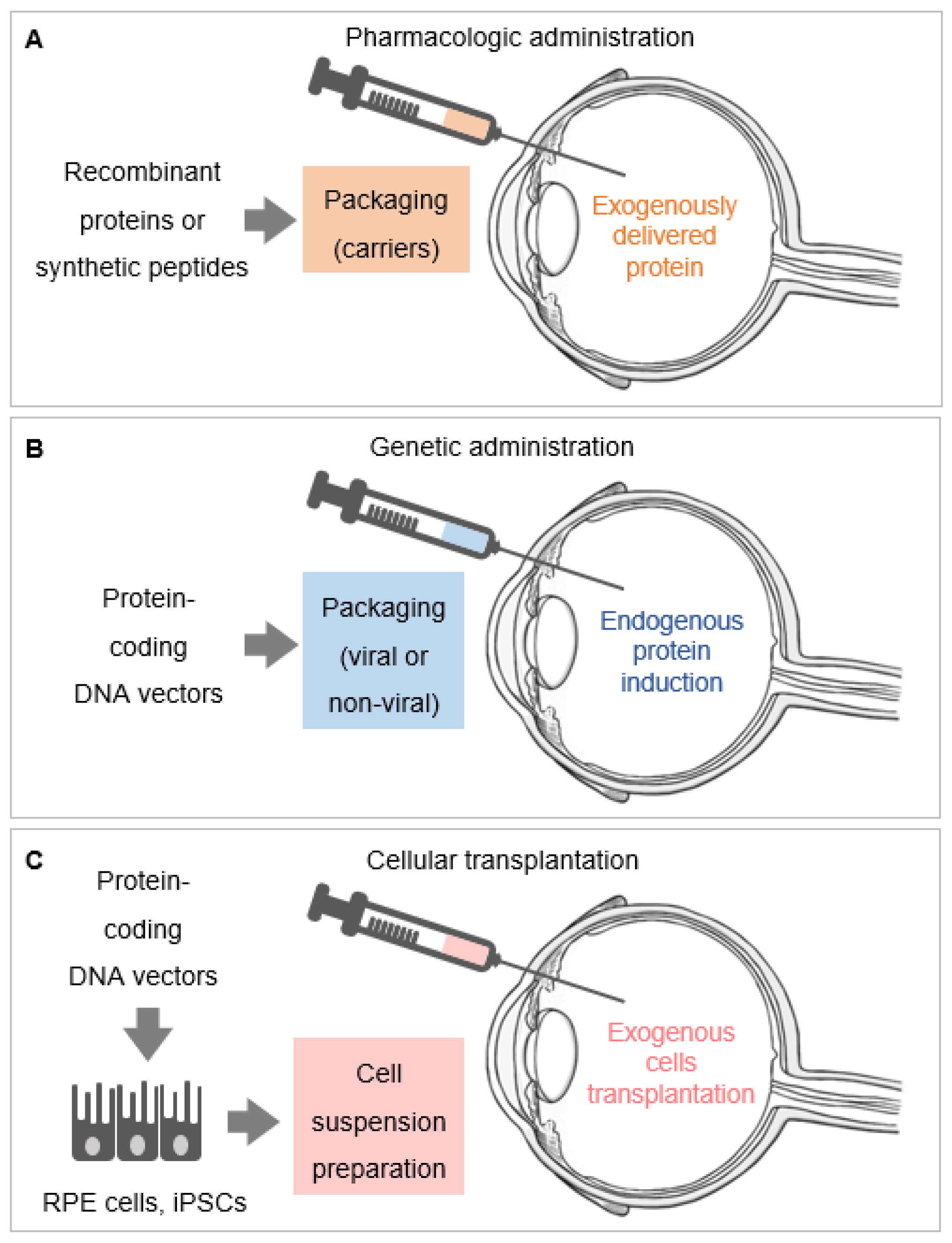
We focus on PEDF delivery systems reported during the last five years. Figure 2 summarizes three systems currently used to deliver PEDF in the retina in vivo.
4.1. Pharmacological Administration of PEDF
PEDF is obtained as a recombinant protein purified from bacterial and mammalian cells. Its derived peptides are chemically synthesized and purified. The routes of drug administration in vivo can be systemic or local such as suprachoroidal, intravitreal, subtenon or subretinal injections, and ectopic (corneal) application and have been extensively reviewed elsewhere [52]. The ectopic delivery of PEDF protein and peptides is performed in vivo with or without the assistance of molecular carriers (Table 1). Carriers can increase the stability of PEDF, which has a relatively short physiologic half-life [53] and prevent the need of repeated injections of PEDF. Table 2 shows a list of recent reports in which pharmacological administration of recombinant PEDF and its corresponding peptides was performed in vivo.
The Table 2 describes experimental models, molecules (PEDF and/or corresponding peptides), types of carrier (when used), routes of PEDF delivery and effect on target cells. Intravitreal injections of PEDF without the use of a carrier remains the most common approach for the pharmacologic administration of PEDF in vivo [34,38,40,54]. Ectopic (eye drops) administration of PEDF has also been used [38,52]. Photoreceptors, and endothelial cells (highlighted in green, and in pink, respectively) are the main targets of pharmacologic administration of PEDF in vivo. Retinal degeneration models such as rd1 [40] and rd10 mice [34] are useful to provide mechanistic insights into PEDF retinoprotection. Oxidative stress injury mouse models are used to investigate the effects of PEDF in preserving photoreceptors integrity [54]. Carriers such as polymer-conjugated ester prodrug [55] and Type I collagen [56] are used in rabbit and in mouse, respectively, to examine the inhibitory effects of PEDF against choroidal neovascularization.
4.2. Genetic Administration of PEDF and Cell-Based Therapies
The administration of genes for production of retinoprotective proteins provide a sustained endogenous expression and therefore avoid the need of multiple administrations. There are challenges associated with the genetic administration of vectors such as targeting specific cells in the retina, e.g., photoreceptors or RPE cells, and safety due to cellular toxicity associated with viral vectors. Table 3 summarizes studies of genetic administration of PEDF in vivo reported during the last five years. It includes organism model, DNA vectors, carriers/host cells, routes of delivery, and effect (including target cells) for genetic administration of PEDF in vivo.
Cell-based delivery systems are also used to administer PEDF to the retina, by first infecting placental-derived [60], human mesenchymal-derived (hMSC) [59] and neural [58] stem cells with viral vectors, or transfecting primary rat RPE cells [62] in vitro with expression vectors, and generating in vitro PEDF-secreting cells that can be transplanted in the vitreous or in the subretinal space in vivo. The cell-based delivery systems of PEDF reported during the last five years are summarized in the Table 3. Most of these studies employ viral vectors to pack PEDF-coding DNA [58,59,61,63], and in vivo infections using mouse and rat models of retinal degeneration and neovascularization. Approaches using cell-based genetic administration of PEDF from mesenchymal stem cells, and subsequently implanting these PEDF-producing cells in the subretinal space or in the vitreous are emerging in studies aiming at the regeneration of the RPE layer. Non-viral carriers for cellular transfections such as AMAXA Nucleofector technology [60] are alternatives to viral infections to transfect PEDF expression vectors. Different types of PEDF-coding, non-viral vectors such as the non-integrating episomal vector pEPito [57] and the non-viral, hyper-active sleeping beauty transposon [62] are used to engineer PEDF-expressing cells for potential cell-based therapies.
5. Future Directions for Protein Delivery Systems in Ocular Research
The research on pharmacologic and genetic administration of retinoprotective proteins has had unprecedented success in generating different treatments toward the recovery of vision loss. Regarding the pharmacologic approach using exogenously prepared proteins and peptides as cargo molecules, a variety of neuron-targeted nanoparticles are available to deliver the cargo and to enhance neuronal protection [64]. Significant progress includes the understanding of the uptake mechanism and the physicochemical properties of nanoparticles and microparticles to encapsulate proteins [64]. The use of novel delivery methodologies, such as microneedles and intravitreal implants that allow the gradual intraocular release of proteins and carriers that are specifically designed for ocular research [65] are increasing the efficacy of the pharmacologic administration of retinoprotective proteins to target retinopathies.
Encapsulation of proteins with nano-formulation shows efficient delivery of retinoprotective proteins. For example, the intravitreal delivery of the neuroprotective peptide erythropoietin (EPO) is improved with the use of microparticles formulated with EPO and the hydrolytically degradable poly(lactic-co-glycolic acid) (PLGA) or the reactive oxygen species (ROS)-degradable poly(propylene sulfide) (PPS) in models of blast injury-induced retinal damage [66]. This research warrants the use of encapsulating PEDF in similar microparticles system to control and extend its release locally using in vivo retinopathy mouse models.
In addition to encapsulating retinoprotective proteins with nano-formulated microparticles, microemulsions (ME) [67,68] and hydrogels [68,69] are also available and known to behave as “smart carriers”. They change phase upon physiological conditions (water content, temperature) and undergo phase-transition from a water-in-oil state to a liquid crystalline state and then to coarse emulsion with different viscosities. This allows slower release of cargo molecules in the eye, which include druggable compounds and retinoprotective proteins. The various sizes of ME, from 5 to 200 nm [67], and their long-term stability make ME and hydrogels compelling delivery system employing the full length PEDF or its derived bioactive peptides for future research.
Regarding the genetic administration of PEDF, viral and non-viral vectors are successfully employed to introduce PEDF expression in retinal and RPE cells (Table 2). As precedent, an adeno-associated virus (AAV-2)-based gene therapy, Vortigern Neparvovec (Luxturnatm), is already approved by the FDA for the treatment of Leber Congenital Amaurosis (LCA) [70]. Despite the advantages of the AAV systems, such as the induction of sustained gene expression and recombinant protein production of a protein, there are safety concerns with toxicity and mutagenesis due to the integration of the virus into the genome, which could cause long term, unpredictable effects. Another limitation for the use of this AAV system is the maximal size of the DNA coding construct (approximately 5kB) that can be packaged into an AAV [71]. There is an increasing need to develop genetic delivery systems that overcome these limitations.
Moreover, it is possible to modify the local expression of naturally occurring proteins by editing their genes. The use of CRISPR-based editing approaches [72] and innovations in designing expression vectors containing both Cas9 and small guiding RNAs (sgRNAs), are now commonly used to infect retinas in vivo in order to edit mutated rhodopsin [73]. CRISPR-based approaches could be employed to edit the endogenous PEDF expressed in RPE cells in vivo, enhancing PEDF activity or increasing its production. Second generation PEDF with alterations that increase its neurotrophic activity are excellent choices for this approach, such as the single amino acid alteration in PEDF[H105A] [33]. Alterations of amino acids to increase the stability of PEDF without affecting its biological activity are also considered of interest for this approach.
Finally, methods of mRNA encapsulation in nanoparticles currently applied in mRNA vaccine technologies against several types of infectious diseases and cancer [74] offer an attractive alternative to viral systems for the genetic administration of retinoprotective genes in ocular research. Such mRNA-based approaches involve relatively novel concepts of RNA-based therapies that include methods in mRNA design and formulation of nanoparticles to encapsulate the mRNA of interest. An example of this methodology that could potentially be applied to the genetic administration of PEDF is the use of the cationic peptide protamine to transfect PEDF-encoding mRNA constructs in vivo [75].
6. Concluding Remarks
The use of retinoprotective proteins in ocular research is currently benefiting from the advances in research on specific carriers that can significantly increase the delivery of these proteins in the vitreous or in the retinal and RPE layers of the eye. Challenges associated with the release and the extracellular stability of these proteins are being addressed by designing packaging carriers that protect the retinoprotective proteins from proteolysis, and the development of slow release carriers that avoid the need of multiple injections of administration for a sustained release of these proteins. The advantage of pharmacologic delivery of retinoprotective proteins when compared with gene and cell therapies, includes the safety of delivering a purified drug without altering the genetic component of the endogenous cells, and/or avoiding rejection as a possible transplantation side effect, respectively.
Among retinoprotective proteins, PEDF is attractive for in vivo studies using pharmacologic or genetic delivery systems and cell therapy strategies. There are several advantages for the use of PEDF delivery systems: 1. PEDF is a retinoprotective protein that naturally occurs in the eye and is present in the aqueous humor of humans [76]; 2. PEDF is nontoxic [77]; 3. PEDF is multimodal and possesses neurotrophic [27], antiangiogenic [55], anti-inflammatory [78], antioxidant properties [79,80], all of which are beneficial for the treatment of several multifactorial retinal diseases; 4. PEDF is a soluble protein that can diffuse from the vitreous to the back of the eye and from the subconjuntiva to the retina [81,82,83,84]; 5. The modular PEDF can be fragmented to yield peptides with distinct individual activities [33]. However, some disadvantages are also associated with PEDF, such as: 1. Some of the mechanisms of action of PEDF remain to be elucidated; 2. PEDF clearance in the eye can occur within the first 24 h of administration and may require sustained release upon pharmacological delivery [82]; 3. Due to its multimodal properties [33], the use of PEDF as a full length protein could trigger undesirable side effects, which warrants the use of its relevant smaller peptides for an individual activity.
Author Contributions
I.T.R. conceptualized and wrote the original draft; A.B.-C., A.I.N. and S.P.B. reviewed and edited the drafted manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This review was supported by the Intramural Research Program of the National Eye Institute, National Institutes of Health (Project #EY000306, SPB).
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
Not Applicable.
Acknowledgments
Authors would like to acknowledge the National Eye Institute at the National Institutes of Health to provide research support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Swaroop, A.; Kim, D.; Forrest, D. Transcriptional regulation of photoreceptor development and homeostasis in the mammalian retina. Nat. Rev. Neurosci. 2010, 11, 563–576. [Google Scholar] [CrossRef] [Green Version]
- Baden, T.; Euler, T.; Berens, P. Understanding the retinal basis of vision across species. Nat. Rev. Neurosci. 2019, 21, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Sengillo, J.D.; Justus, S.; Tsai, Y.-T.; Cabral, T.; Tsang, S.H. Gene and cell-based therapies for inherited retinal disorders: An update. Am. J. Med. Genet. Part C Semin. Med. Genet. 2016, 172, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Cervia, D.; Catalani, E.; Casini, G. Neuroprotective Peptides in Retinal Disease. J. Clin. Med. 2019, 8, 1146. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Sun, Y.; Cakir, B.; Tomita, Y.; Huang, S.; Wang, Z.; Liu, C.-H.; Cho, S.S.; Britton, W.; Kern, T.S.; et al. Targeting Neurovascular Interaction in Retinal Disorders. Int. J. Mol. Sci. 2020, 21, 1503. [Google Scholar] [CrossRef] [Green Version]
- Arroba, A.I.; Campos-Caro, A.; Aguilar-Diosdado, M.; Valverde, Á.M. IGF-1, Inflammation and Retinal Degeneration: A Close Network. Front. Aging Neurosci. 2018, 10, 203. [Google Scholar] [CrossRef]
- Wyse-Jackson, A.C.; Roche, S.L.; Ruiz-Lopez, A.M.; Moloney, J.N.; Byrne, A.M.; Cotter, T.G. Progesterone analogue protects stressed photoreceptors via bFGF-mediated calcium influx. Eur. J. Neurosci. 2016, 44, 3067–3079. [Google Scholar] [CrossRef]
- Li, S.; Sato, K.; Gordon, W.C.; Sendtner, M.; Bazan, N.G.; Jin, M. Ciliary neurotrophic factor (CNTF) protects retinal cone and rod photoreceptors by suppressing excessive formation of the visual pigments. J. Biol. Chem. 2018, 293, 15256–15268. [Google Scholar] [CrossRef] [Green Version]
- Daly, C.; Shine, L.; Heffernan, T.; Deeti, S.; Reynolds, A.L.; O’Connor, J.J.; Dillon, E.T.; Duffy, D.J.; Kolch, W.; Cagney, G.; et al. A Brain-Derived Neurotrophic Factor Mimetic Is Sufficient to Restore Cone Photoreceptor Visual Function in an Inherited Blindness Model. Sci. Rep. 2017, 7, 11320. [Google Scholar] [CrossRef] [Green Version]
- Sacchetti, M.; Mantelli, F.; Rocco, M.L.; Micera, A.; Brandolini, L.; Focareta, L.; Pisano, C.; Aloe, L.; Lambiase, A. Recombinant Human Nerve Growth Factor Treatment Promotes Photoreceptor Survival in the Retinas of Rats with Retinitis Pigmentosa. Curr. Eye Res. 2017, 42, 1064–1068. [Google Scholar] [CrossRef]
- Cachafeiro, M.; Bemelmans, A.-P.; Samardzija, M.; Afanasieva, T.; Pournaras, J.-A.; Grimm, C.; Kostic, C.; Philippe, S.; Wenzel, A.; Arsenijevic, Y. Hyperactivation of retina by light in mice leads to photoreceptor cell death mediated by VEGF and retinal pigment epithelium permeability. Cell Death Dis. 2013, 4, e781. [Google Scholar] [CrossRef]
- Shioda, S.; Takenoya, F.; Wada, N.; Hirabayashi, T.; Seki, T.; Nakamachi, T. Pleiotropic and retinoprotective functions of PACAP. Anat. Sci. Int. 2016, 91, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Akrouh, A.; Kerschensteiner, D. Morphology and function of three VIP-expressing amacrine cell types in the mouse retina. J. Neurophysiol. 2015, 114, 2431–2438. [Google Scholar] [CrossRef] [PubMed]
- Szabadfi, K.; Pintér, E.; Reglodi, R.; Gábriel, R. Neuropeptides, Trophic Factors, and Other Substances Providing Morphofunctional and Metabolic Protection in Experimental Models of Diabetic Retinopathy. Int. Rev. Cell Mol. Biol. 2014, 311, 1–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sall, J.W.; Klisovic, D.D.; O’Dorisio, M.; Katz, S.E. Somatostatin inhibits IGF-1 mediated induction of VEGF in human retinal pigment epithelial cells. Exp. Eye Res. 2004, 79, 465–476. [Google Scholar] [CrossRef]
- Fernández-Sánchez, L.; Lax, P.; Isiegas, C.; Ayuso, E.; Ruiz, J.M.; De La Villa, P.; Bosch, F.; De La Rosa, E.J.; Cuenca, N. Proinsulin Slows Retinal Degeneration and Vision Loss in the P23H Rat Model of Retinitis Pigmentosa. Hum. Gene Ther. 2012, 23, 1290–1300. [Google Scholar] [CrossRef] [Green Version]
- Corrochano, S.; Barhoum, R.; Boya, P.; Arroba, A.I.; Rodríguez-Muela, N.; Gómez-Vicente, V.; Bosch, F.; de Pablo, F.; de la Villa, P.; de la Rosa, E.J. Attenuation of Vision Loss and Delay in Apoptosis of Photoreceptors Induced by Proinsulin in a Mouse Model of Retinitis Pigmentosa. Investig. Opthalmology Vis. Sci. 2008, 49, 4188–4194. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Li, F.; Steinberg, R.H.; Lavail, M.M. Development of Normal and Injury-induced Gene Expression of aFGF, bFGF, CNTF, BDNF, GFAP and IGF-I in the Rat Retina. Exp. Eye Res. 2001, 72, 591–604. [Google Scholar] [CrossRef]
- West, E.L.; Pearson, R.A.; Duran, Y.; Cordero, A.G.; MacLaren, R.E.; Smith, A.J.; Sowden, J.C.; Ali, R.R. Manipulation of the Recipient Retinal Environment by Ectopic Expression of Neurotrophic Growth Factors Can Improve Transplanted Photoreceptor Integration and Survival. Cell Transplant. 2012, 21, 871–887. [Google Scholar] [CrossRef] [Green Version]
- Rai, U.; Thrimawithana, T.R.; Valery, C.; Young, S.A. Therapeutic uses of somatostatin and its analogues: Current view and potential applications. Pharmacol. Ther. 2015, 152, 98–110. [Google Scholar] [CrossRef]
- Atlasz, T.; Werling, D.; Song, S.; Szabo, E.; Vaczy, A.; Kovari, P.; Tamas, A.; Reglodi, D.; Yu, R. Retinoprotective Effects of TAT-Bound Vasoactive Intestinal Peptide and Pituitary Adenylate Cyclase Activating Polypeptide. J. Mol. Neurosci. 2018, 68, 397–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himawan, E.; Ekström, P.; Buzgo, M.; Gaillard, P.; Stefánsson, E.; Marigo, V.; Loftsson, T.; Paquet-Durand, F. Drug delivery to retinal photoreceptors. Drug Discov. Today 2019, 24, 1637–1643. [Google Scholar] [CrossRef]
- Punzo, C.; Kornacker, K.; Cepko, C.L. Stimulation of the insulin/mTOR pathway delays cone death in a mouse model of retinitis pigmentosa. Nat. Neurosci. 2008, 12, 44–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polato, F.; Becerra, S.P. Pigment Epithelium-Derived Factor, a Protective Factor for Photoreceptors in Vivo. Adv. Exp. Med. Biol. 2016, 854, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Pagan-Mercado, G.; Becerra, S.P. Signaling Mechanisms Involved in PEDF-Mediated Retinoprotection. Adv. Exp. Med. Biol. 2019, 1185, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Liu, J.; Pu, J.; Milne, G.; Chen, M.; Xu, H.; Shipley, A.; Forrester, J.V.; McCaig, C.D.; Lois, N. Polarized retinal pigment epithelium generates electrical signals that diminish with age and regulate retinal pathology. J. Cell. Mol. Med. 2018, 22, 5552–5564. [Google Scholar] [CrossRef]
- Fudalej, E.; Justyniarska, M.; Kasarełło, K.; Dziedziak, J.; Szaflik, J.P.; Cudnoch-Jędrzejewska, A. Neuroprotective factors of the retina and their role in promoting survival of retinal ganglion cells: A review. Ophthalmic Res. 2021. [Google Scholar] [CrossRef]
- Becerra, S.P.; Notario, V. The effects of PEDF on cancer biology: Mechanisms of action and therapeutic potential. Nat. Rev. Cancer 2013, 13, 258–271. [Google Scholar] [CrossRef] [Green Version]
- Dixit, S.; Polato, F.; Samardzija, M.; Abu-Asab, M.; Grimm, C.; Crawford, S.E.; Becerra, S.P. PEDF deficiency increases the susceptibility of rd10 mice to retinal degeneration. Exp. Eye Res. 2020, 198, 108121. [Google Scholar] [CrossRef]
- Notari, L.; Baladron, V.; Aroca-Aguilar, J.D.; Balko, N.; Heredia, R.; Meyer, C.; Notario, P.M.; Saravanamuthu, S.; Nueda, M.-L.; Sanchez-Sanchez, F.; et al. Identification of a Lipase-linked Cell Membrane Receptor for Pigment Epithelium-derived Factor. J. Biol. Chem. 2006, 281, 38022–38037. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, P.; Notario, P.M.; Becerra, S.P. Pigment epithelium-derived factor receptor (PEDF-R): A plasma membrane-linked phospholipase with PEDF binding affinity. Single Mol. Single Cell Seq. 2009, 664, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, P.; Locatelli-Hoops, S.; Kenealey, J.; DesJardin, J.; Notari, L.; Becerra, S.P. Pigment Epithelium-derived Factor (PEDF) Prevents Retinal Cell Death via PEDF Receptor (PEDF-R). J. Biol. Chem. 2013, 288, 23928–23942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenealey, J.; Subramanian, P.; Comitato, A.; Bullock, J.; Keehan, L.; Polato, F.; Hoover, D.; Marigo, V.; Becerra, S.P. Small Retinoprotective Peptides Reveal a Receptor-binding Region on Pigment Epithelium-derived Factor. J. Biol. Chem. 2015, 290, 25241–25253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Pinto, A.; Polato, F.; Subramanian, P.; de la Rocha-Muñoz, A.; Vitale, S.; de la Rosa, E.J.; Becerra, S.P. PEDF peptides promote photoreceptor survival in rd10 retina models. Exp. Eye Res. 2019, 184, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Winokur, P.N.; Subramanian, P.; Bullock, J.L.; Arocas, V.; Becerra, S.P. Comparison of two neurotrophic serpins reveals a small fragment with cell survival activity. Mol. Vis. 2017, 23, 372–384. [Google Scholar]
- Kawahara, K.; Yoshida, T.; Maruno, T.; Oki, H.; Ohkubo, T.; Koide, T.; Kobayashi, Y. Spatiotemporal regulation of PEDF signaling by type I collagen remodeling. Proc. Natl. Acad. Sci. USA 2020, 117, 11450–11458. [Google Scholar] [CrossRef]
- García-Onrubia, L.; Valentín-Bravo, F.; Coco-Martin, R.; González-Sarmiento, R.; Pastor, J.; Usategui-Martín, R.; Pastor-Idoate, S. Matrix Metalloproteinases in Age-Related Macular Degeneration (AMD). Int. J. Mol. Sci. 2020, 21, 5934. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, J.; Geng, H.; Li, L.; Li, J.; Cheng, B.; Ma, X.; Li, H.; Hou, L. Photoreceptor degeneration in microphthalmia (Mitf) mice: Partial rescue by pigment epithelium-derived factor. Dis. Model. Mech. 2019, 12. [Google Scholar] [CrossRef] [Green Version]
- Valiente-Soriano, F.J.; Di Pierdomenico, J.; García-Ayuso, D.; Ortín-Martínez, A.; De Imperial-Ollero, J.A.M.; Gallego-Ortega, A.; Jiménez-López, M.; Villegas-Pérez, M.P.; Becerra, S.P.; Vidal-Sanz, M. Pigment Epithelium-Derived Factor (PEDF) Fragments Prevent Mouse Cone Photoreceptor Cell Loss Induced by Focal Phototoxicity In Vivo. Int. J. Mol. Sci. 2020, 21, 7242. [Google Scholar] [CrossRef]
- Comitato, A.; Subramanian, P.; Turchiano, G.; Montanari, M.; Becerra, S.P.; Marigo, V. Pigment epithelium-derived factor hinders photoreceptor cell death by reducing intracellular calcium in the degenerating retina. Cell Death Dis. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.L.; He, J.; Kakazu, A.H.; Jun, B.; Bazan, N.G.; Bazan, H.E.P. Defining a mechanistic link between pigment epithelium–derived factor, docosahexaenoic acid, and corneal nerve regeneration. J. Biol. Chem. 2017, 292, 18486–18499. [Google Scholar] [CrossRef] [Green Version]
- Yeh, S.-I.; Yu, S.-H.; Chu, H.-S.; Huang, C.-T.; Tsao, Y.-P.; Cheng, C.-M.; Chen, W.-L. Pigment Epithelium-Derived Factor Peptide Promotes Corneal Nerve Regeneration: An In Vivo and In Vitro Study. Investig. Opthalmol. Vis. Sci. 2021, 62, 23. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Subramanian, P.; Shen, D.; Tuo, J.; Becerra, S.P.; Chan, C.-C. Pigment Epithelium-Derived Factor Reduces Apoptosis and Pro-Inflammatory Cytokine Gene Expression in a Murine Model of Focal Retinal Degeneration. ASN Neuro 2013, 5, e00126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holekamp, N.M.; Bouck, N.P.; Volpert, O.V. Pigment epithelium-derived factor is deficient in the vitreous of patients with choroidal neovascularization due to age-related macular degeneration. Am. J. Ophthalmol. 2002, 134, 220–227. [Google Scholar] [CrossRef]
- Ogata, N.; Nishikawa, M.; Nishimura, T.; Mitsuma, Y.; Matsumura, M. Unbalanced vitreous levels of pigment epithelium-derived factor and vascular endothelial growth factor in diabetic retinopathy. Am. J. Ophthalmol. 2002, 134, 348–353. [Google Scholar] [CrossRef]
- Ogata, N.; Matsuoka, M.; Imaizumi, M.; Arichi, M.; Matsumura, M. Decreased levels of pigment Epithelium–derived factor in eyes with neuroretinal dystrophic diseases. Am. J. Ophthalmol. 2004, 137, 1129–1130. [Google Scholar] [CrossRef] [PubMed]
- Becerra, S.P.; Fariss, R.N.; Wu, Y.Q.; Montuenga, L.M.; Wong, P.; Pfeffer, B.A. Pigment epithelium-derived factor in the monkey retinal pigment epithelium and interphotoreceptor matrix: Apical secretion and distribution. Exp. Eye Res. 2004, 78, 223–234. [Google Scholar] [CrossRef]
- Samuel, W.; Jaworski, C.; Postnikova, O.A.; Kutty, R.K.; Duncan, T.; Tan, L.X.; Poliakov, E.; Lakkaraju, A.; Redmond, T.M. Appropriately differentiated ARPE-19 cells regain phenotype and gene expression profiles similar to those of native RPE cells. Mol. Vis. 2017, 23, 60–89. [Google Scholar]
- Choudhary, P.; Booth, H.; Gutteridge, A.; Surmacz, B.; Louca, I.; Steer, J.; Kerby, J.; Whiting, P.J. Directing Differentiation of Pluripotent Stem Cells Toward Retinal Pigment Epithelium Lineage. STEM CELLS Transl. Med. 2016, 6, 490–501. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.W.J.; Wang, J.; Zheng, L.; Wang, M.; Lu, Y.; Li, Z.; Lian, C.; Mao, S.; Hou, X.; Li, S.; et al. miR-25 mediates retinal degeneration via inhibiting ITGAV and PEDF in rat. Curr. Mol. Med. 2018, 17, 359–374. [Google Scholar] [CrossRef]
- Zhang, T.; Yin, P.; Zhang, Z.; Xu, B.; Che, D.; Dai, Z.; Dong, C.; Jiang, P.; Hong, H.; Yang, Z.; et al. Deficiency of pigment epithelium-derived factor in nasopharyngeal carcinoma cells triggers the epithelial–mesenchymal transition and metastasis. Cell Death Dis. 2017, 8, e2838. [Google Scholar] [CrossRef]
- Castro-Balado, A.; Mondelo-García, C.; Zarra-Ferro, I.; Fernández-Ferreiro, A. New ophthalmic drug delivery systems. Farm Hosp. 2020, 44, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.-J.; Huang, L.-Z.; Xu, X.-L.; Du, W.; Zhou, A.-Y.; Yu, W.-Z.; Li, X.-X. Polyethylene Glycol-Modified Pigment Epithelial-Derived Factor: New Prospects for Treatment of Retinal Neovascularization. J. Pharmacol. Exp. Ther. 2012, 342, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liu, X.; Ren, Y.; Liu, Y.; Han, S.; Zhao, J.; Gou, X.; He, Y. PEDF protects human retinal pigment epithelial cells against oxidative stress via upregulation of UCP2 expression. Mol. Med. Rep. 2018, 19, 59–74. [Google Scholar] [CrossRef] [Green Version]
- Sheibani, N.; Wang, S.; Darjatmoko, S.R.; Fisk, D.L.; Shahi, P.K.; Pattnaik, B.R.; Sorenson, C.M.; Bhowmick, R.; Volpert, O.V.; Albert, D.M.; et al. Novel anti-angiogenic PEDF-derived small peptides mitigate choroidal neovascularization. Exp. Eye Res. 2019, 188, 107798. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Roh, K.-H.; Kim, S.W.; Park, S.J.; Lim, N.-Y.; Jung, H.; Choi, I.-W.; Park, S. Type I pig collagen enhances the efficacy of PEDF 34-mer peptide in a mouse model of laser-induced choroidal neovascularization. Graefe’s Arch. Clin. Exp. Ophthalmol. 2019, 257, 1709–1717. [Google Scholar] [CrossRef] [PubMed]
- Calado, S.M.; Diaz-Corrales, F.J.; Silva, G.A.; Calado, M.S.M. pEPito-driven PEDF Expression Ameliorates Diabetic Retinopathy Hallmarks. Hum. Gene Ther. Methods 2016, 27, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.M.; Zhang, Z.R.; Zhang, Y.G.; Gao, Y.S. Neural Stem Cell-based Intraocular Administration of Pigment Epithelium-derived Factor Promotes Retinal Ganglion Cell Survival and Axon Regeneration after Optic Nerve Crush Injury in Rat: An Experimental Study. Iran. J. Med. Sci. 2016, 41, 382. [Google Scholar]
- Nascimento-Dos-Santos, G.; Teixeira-Pinheiro, L.C.; Da Silva-Júnior, A.J.; De Carvalho, L.R.P.; Mesentier-Louro, L.A.; Hauswirth, W.W.; Mendez-Otero, R.; Santiago, M.F.; Petrs-Silva, H. Effects of a combinatorial treatment with gene and cell therapy on retinal ganglion cell survival and axonal outgrowth after optic nerve injury. Gene Ther. 2019, 27, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, S.; Park, S.H.; Lee, D.; Kim, G.H.; Noh, J.E.; Lee, K.J.; Kim, G.J. Overexpression of pigment epithelium-derived factor in placenta-derived mesenchymal stem cells promotes mitochondrial biogenesis in retinal cells. Lab. Investig. 2021, 101, 51–69. [Google Scholar] [CrossRef] [PubMed]
- Askou, A.L.; Alsing, S.; Benckendorff, J.N.; Holmgaard, A.; Mikkelsen, J.G.; Aagaard, L.; Bek, T.; Corydon, T.J. Suppression of Choroidal Neovascularization by AAV-Based Dual-Acting Antiangiogenic Gene Therapy. Mol. Ther. Nucleic Acids 2019, 16, 38–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Garcia, L.; Recalde, S.; Hernandez, M.; Bezunartea, J.; Rodriguez-Madoz, J.R.; Johnen, S.; Diarra, S.; Marie, C.; Izsvák, Z.; Ivics, Z.; et al. Long-Term PEDF Release in Rat Iris and Retinal Epithelial Cells after Sleeping Beauty Transposon-Mediated Gene Delivery. Mol. Ther. Nucleic Acids 2017, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.-J.; Mo, B.; Liu, L.; Yue, Y.-K.; Yue, C.-L.; Liu, W. Inhibition of choroidal neovascularization by lentivirus-mediated PEDF gene transfer in rats. Int. J. Ophthalmol. 2016, 9, 1112–1120. [Google Scholar] [CrossRef]
- Sabel, B.A.; You, Q. Nanoparticles as a tool to deliver drugs to the retina and brain: An update. Neural Regen. Res. 2021, 16, 283–284. [Google Scholar] [CrossRef] [PubMed]
- Kang-Mieler, J.J.; Rudeen, K.M.; Liu, W.; Mieler, W.F. Advances in ocular drug delivery systems. Eye 2020, 34, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- DeJulius, C.; Bernardo-Colón, A.; Naguib, S.; Backstrom, J.; Kavanaugh, T.; Gupta, M.; Duvall, C.; Rex, T. Microsphere antioxidant and sustained erythropoietin-R76E release functions cooperate to reduce traumatic optic neuropathy. J. Control. Release 2021, 329, 762–773. [Google Scholar] [CrossRef]
- Üstündağ Okur, N.; Çağlar,, E.Ş.; Siafaka, P.I. Novel Ocular Drug Delivery Systems: An Update on Microemulsions. J. Ocul. Pharmacol. Ther. 2020, 36, 342–354. [Google Scholar] [CrossRef]
- Dubald, M.; Bourgeois, S.; Andrieu, V.; Fessi, H. Ophthalmic Drug Delivery Systems for Antibiotherapy—A Review. Pharmaceutics 2018, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Lynch, C.R.; Kondiah, P.P.D.; Choonara, Y.E.; Du Toit, L.C.; Ally, N.; Pillay, V. Hydrogel Biomaterials for Application in Ocular Drug Delivery. Front. Bioeng. Biotechnol. 2020, 8, 228. [Google Scholar] [CrossRef] [Green Version]
- Maguire, A.M.; Bennett, J.; Aleman, E.M.; Leroy, B.P.; Aleman, T.S. Clinical Perspective: Treating RPE65-Associated Retinal Dystrophy. Mol. Ther. 2021, 29, 442–463. [Google Scholar] [CrossRef]
- Chamberlain, K.; Riyad, J.M.; Weber, T. Expressing Transgenes That Exceed the Packaging Capacity of Adeno-Associated Virus Capsids. Hum. Gene Ther. Methods 2016, 27, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Backstrom, J.R.; Sheng, J.; Wang, M.C.; Bernardo-Colón, A.; Rex, T.S. Optimization of S. aureus dCas9 and CRISPRi Elements for a Single Adeno-Associated Virus that Targets an Endogenous Gene. Mol. Ther. Methods Clin. Dev. 2020, 19, 139–148. [Google Scholar] [CrossRef]
- Latella, M.C.; Di Salvo, M.T.; Cocchiarella, F.; Benati, D.; Grisendi, G.; Comitato, A.; Marigo, V.; Recchia, A. In vivo Editing of the Human Mutant Rhodopsin Gene by Electroporation of Plasmid-based CRISPR/Cas9 in the Mouse Retina. Mol. Ther. Nucleic Acids 2016, 5, e389. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarzebska, N.T.; Lauchli, S.; Iselin, C.; French, L.E.; Johansen, P.; Guenova, E.; Kündig, T.M.; Pascolo, S. Functional differences between protamine preparations for the transfection of mRNA. Drug Deliv. 2020, 27, 1231–1235. [Google Scholar] [CrossRef]
- Boehm, B.O.; Lang, G.; Volpert, O.; Jehle, P.M.; Kurkhaus, A.; Rosinger, S.; Bouck, N. Low content of the natural ocular anti-angiogenic agent pigment epithelium-derived factor (PEDF) in aqueous humor predicts progression of diabetic retinopathy. Diabetologia 2003, 46, 394–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Li, N.; Xu, B.; Wu, M.; Yan, X.; Zhong, L.; Cai, H.; Wang, T.; Wang, Q.; Long, F.; et al. Polymer-based nanoparticles for chemo/gene-therapy: Evaluation its therapeutic efficacy and toxicity against colorectal carcinoma. Biomed. Pharmacother. 2019, 118, 109257. [Google Scholar] [CrossRef]
- Ma, B.; Zhou, Y.; Liu, R.; Zhang, K.; Yang, T.; Hu, C.; Gao, Y.; Lan, Q.; Liu, Y.; Yang, X.; et al. Pigment epithelium-derived factor (PEDF) plays anti-inflammatory roles in the pathogenesis of dry eye disease. Ocul. Surf. 2021, 20, 70–85. [Google Scholar] [CrossRef]
- Liu, X.; Liu, H.; Lu, X.; Tombran-Tink, J.; Zhao, S. PEDF Attenuates Ocular Surface Damage in Diabetic Mice Model Through Its Antioxidant Properties. Curr. Eye Res. 2021, 46, 302–308. [Google Scholar] [CrossRef]
- Brook, N.; Brook, E.; Dharmarajan, A.; Chan, A.; Dass, C.R. The role of pigment epithelium-derived factor in protecting against cellular stress. Free. Radic. Res. 2019, 53, 1166–1180. [Google Scholar] [CrossRef]
- Patel, J.I.; Tombran-Tink, J.; Hykin, P.G.; Gregor, Z.J.; Cree, I.A. Vitreous and aqueous concentrations of proangiogenic, antiangiogenic factors and other cytokines in diabetic retinopathy patients with macular edema: Implications for structural differences in macular profiles. Exp. Eye Res. 2006, 82, 798–806. [Google Scholar] [CrossRef]
- Amaral, J.; Fariss, R.N.; Campos, M.M.; Robison, W.G.; Kim, H.; Lutz, R.; Becerra, S.P. Transscleral-RPE Permeability of PEDF and Ovalbumin Proteins: Implications for Subconjunctival Protein Delivery. Investig. Opthalmo. Vis. Sci. 2005, 46, 4383–4392. [Google Scholar] [CrossRef]
- Cayouette, M.; Smith, S.B.; Becerra, S.; Gravel, C. Pigment Epithelium-Derived Factor Delays the Death of Photoreceptors in Mouse Models of Inherited Retinal Degenerations. Neurobiol. Dis. 1999, 6, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Rieke, E.R.; Amaral, J.; Becerra, S.P.; Lutz, R.J. Sustained Subconjunctival Protein Delivery Using a Thermosetting Gel Delivery System. J. Ocul. Pharmacol. Ther. 2010, 26, 55–64. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Structure of PEDF and P1 peptide, and sequences of P1 and PEDF-derived peptides. (A) Modeling of binding interaction of LBD of human PEDF-R to human PEDF. The structure of the peptide P1 derived from the ab initio fragment assembly protocol of Rosetta is shown in orange. The resultant P1 peptide structure docked to the PEDF crystal structure (Protein Data Bank code 1IMV) using Rosetta program is shown. P1 peptide docked to a cleft that contained a solvent-exposed region corresponding to α-helix C within the residues 98–114 (17-mer; blue) of the neurotrophic 44 amino acid region (red). The antiangiogenic peptide region (34-mer; green) was not part of the docking region. (B) Peptides were chemically synthesized and purified. Sequences were from human PEDF-R peptide P1 (Thr210–Leu249) and human PEDF peptides 34-mer (Asp44–Asn77), 44-mer(Val78–Thr121), and 17-mer(Gln98–Ser115) as well as the set of 17-mer alanine scan peptides. The H105A alteration in the 17-mer peptide (yellow) exhibited higher PEDF-R P1 affinity and retinoprotective activity when compared with the unmodified 17-mer, while the R99A alteration (black) abolished the P1 affinity and retinoprotective activity. (Adapted from [32]).
Figure 1.
Structure of PEDF and P1 peptide, and sequences of P1 and PEDF-derived peptides. (A) Modeling of binding interaction of LBD of human PEDF-R to human PEDF. The structure of the peptide P1 derived from the ab initio fragment assembly protocol of Rosetta is shown in orange. The resultant P1 peptide structure docked to the PEDF crystal structure (Protein Data Bank code 1IMV) using Rosetta program is shown. P1 peptide docked to a cleft that contained a solvent-exposed region corresponding to α-helix C within the residues 98–114 (17-mer; blue) of the neurotrophic 44 amino acid region (red). The antiangiogenic peptide region (34-mer; green) was not part of the docking region. (B) Peptides were chemically synthesized and purified. Sequences were from human PEDF-R peptide P1 (Thr210–Leu249) and human PEDF peptides 34-mer (Asp44–Asn77), 44-mer(Val78–Thr121), and 17-mer(Gln98–Ser115) as well as the set of 17-mer alanine scan peptides. The H105A alteration in the 17-mer peptide (yellow) exhibited higher PEDF-R P1 affinity and retinoprotective activity when compared with the unmodified 17-mer, while the R99A alteration (black) abolished the P1 affinity and retinoprotective activity. (Adapted from [32]).

Figure 2.
Systems of local delivery of retinoprotective proteins. (A) Pharmacologic administration of retinoprotective proteins, which consists of delivering purified recombinant proteins and synthetic peptides. These can be packaged into carriers (nanoparticles, microparticles, microemulsions) before administration and delivered into the vitreous (as exemplified in the figure). (B) Genetic administration of retinoprotective proteins is accomplished by using DNA expression vectors packaged into viral and non-viral carriers to induce the endogenous expression of these proteins. (C) The genetic administration of retinoprotective proteins can be performed in exogenous cells in vitro, via DNA expression vectors packaged into viral and non-viral carriers, in order to induce the production of recombinant retinoprotective proteins. These exogenously transfected or infected cells, exemplified by retinal pigment epithelium (RPE) and induced pluripotent stem cells (iPSCs), are then transplanted into the retina. Only one route of delivery (intravitreal injection) is shown in the Figure 2A–C; other routes of delivery of retinoprotective proteins include injections in the subretinal space or in the subconjunctiva, and the use of topical corneal drops.
Figure 2.
Systems of local delivery of retinoprotective proteins. (A) Pharmacologic administration of retinoprotective proteins, which consists of delivering purified recombinant proteins and synthetic peptides. These can be packaged into carriers (nanoparticles, microparticles, microemulsions) before administration and delivered into the vitreous (as exemplified in the figure). (B) Genetic administration of retinoprotective proteins is accomplished by using DNA expression vectors packaged into viral and non-viral carriers to induce the endogenous expression of these proteins. (C) The genetic administration of retinoprotective proteins can be performed in exogenous cells in vitro, via DNA expression vectors packaged into viral and non-viral carriers, in order to induce the production of recombinant retinoprotective proteins. These exogenously transfected or infected cells, exemplified by retinal pigment epithelium (RPE) and induced pluripotent stem cells (iPSCs), are then transplanted into the retina. Only one route of delivery (intravitreal injection) is shown in the Figure 2A–C; other routes of delivery of retinoprotective proteins include injections in the subretinal space or in the subconjunctiva, and the use of topical corneal drops.


{kind=link}
{kind=link}
Table 1.
Representative examples of mechanisms of action for retinoprotective proteins *.
| Mechanism of Action | Retinoprotective Protein | Abbreviation | Expressing | Target | Ref. |
|---|---|---|---|---|---|
| Cells | Cells | ||||
| Autocrine | insulin-like growth factor 1 | IGF-1 | photoreceptors | photoreceptors | [6] |
| fibroblast growth factor 2 | FGF-2 | photoreceptors | photoreceptors | [7] | |
| ciliary neurotrophic factor | CNTF | photoreceptors | photoreceptors | [8] | |
| brain-derived neurotrophic factor | BDNF | photoreceptors | photoreceptors | [9] | |
| nerve Growth factor | NGF | photoreceptors | photoreceptors | [10] | |
| vascular endothelial growth factor | VEGF | photoreceptors | photoreceptors | [11] | |
| Paracrine | pituitary adenylate cyclase-activating polypeptide | PACAP | retinal cells | photoreceptors | [12] |
| vasoactive intestinal peptide | VIP | (except photoreceptors) | photoreceptors | [13] | |
| insulin-like growth factor 1 | IGF-1 | amacrine cells | photoreceptors | [6] | |
| somatostatin, somatotropin release inhibiting factor | SRIF | microglia | photoreceptors | [14] | |
| vascular endothelial growth factor | VEGF | RPE | choroidal endothelium | [15] | |
| Endocrine | proinsulin | pro-INS | skeletal muscle | photoreceptors | [16,17] |
| insulin | INS | none (systemic administration) | photoreceptors | [17] |
* Color-code shows the mechanism of action: autocrine (green), paracrine (purple) and endocrine (pink).
Table 2.
Pharmacologic administration of PEDF and derived peptides in vivo *.
| Organism Model | Molecules | Carrier | Route of | Effect | Ref. |
|---|---|---|---|---|---|
| Delivery | |||||
| mouse LED-induced | rhuPEDF | none | intravitreal | photoreceptor survival | [39] |
| Phototoxicity | 17-mer 17-mer[H105A] | injection | |||
| mouse Mitf KO-induced microphtalmia | rhuPEDF | none | eye drops | photoreceptor survival | [38] |
| 17-mer | |||||
| mouse retinal | rhuPEDF44-mer | none | intravitreal | photoreceptor survival | [34] |
| degeneration (rd10) | 17-mer | injection | |||
| 17-mer[H105A] | |||||
| mouse retinal degeneration (rd1) | rhuPEDF | none | intravitreal | photoreceptor survival | [40] |
| 17-mer | injection | ||||
| 17-mer[H105A] | |||||
| mouse oxidative stress | rhuPEDF | none | intravitreal | photoreceptor survival | [54] |
| injury (H2O2) | injection | ||||
| rabbit laser-induced | 8-mer (peptide 335, modified from 34-mer) | polymeric | intravitreal | choroidal neovascularization inhibition | [55] |
| choroidal | ester | injection | |||
| neovascularization | prodrug | ||||
| mouse laser-induced | 34-mer | type I | intravitreal | choroidal neovascularization inhibition | [56] |
| choroidal | collagen | injection | |||
| neovascularization | eye drops |
* The color code indicates the target cells for the pharmacologic administration of PEDF in vivo (green: retinal cells; pink: endothelial cells); rhu: recombinant, human.
Table 3.
Genetic administration of PEDF in vivo *.
| Organism Model | DNA Vector | Carrier/Host Cells | Route of Delivery | Effect | Ref. |
|---|---|---|---|---|---|
| Ins2Akita diabetic mouse | pEPito-hCMV-PEDF episomal vector | none | subretinal injection | photoreceptors survival | [57] |
| (electroporation) | |||||
| rat optic nerve crush | PEDF-secreting neural stem cell (NSC)-based system, lentivirus | none | subretinal cell transplantation | photoreceptors survival | [58] |
| host cells: human neural stem cells | |||||
| rat optic nerve crush | AAV2.PEDF combined with human mesenchymal stem cell | none | Intravitreal cell transplantation | RGC survival and ON injury | [59] |
| host cells: human mesenchymal stem cells | |||||
| rat H2O2-induced retinal degeneration | Placenta-derived-mesenchymal stem cells overexpressing PEDF plasmid | none | Intravitreal cell transplantation | Mitochondrial biogenesis in RPE cells | [60] |
| host cells: human placental-derived stem cells | |||||
| mouse laser-induced neovascularization | multigenic AAV5.PEDF + multiple miRNAs targeting the VEGF-A gene, p/miR(5,B,7/Irr)-AsR/PEDF-PE | none | Intravitreal injection | Choroidal cells inhibition | [61] |
| rat choroidal neovascularization | IPE or RPE cells with pFAR4-ITRs CMV PEDF BGH plasmid, sleeping beauty transposon | none | Subretinal cell transplantation | choroidal cells inhibition | [62] |
| (electroporation) | |||||
| host cells: primary rat RPE cells | |||||
| rat choroidal neovascularization | Lentivirus-PEDF-green fluorescent protein (GFP) | none | Intravitreal injection | choroidal cells inhibition | [63] |
* The color code indicates the target cells for these studies (green: retinal photoreceptor cells; purple: RPE cells; pink: endothelial cells). When cell therapies are associated with the genetic delivery of PEDF, the host cells to be transplanted are indicated.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rebustini, I.T.; Bernardo-Colón, A.; Nalvarte, A.I.; Becerra, S.P. Delivery Systems of Retinoprotective Proteins in the Retina. Int. J. Mol. Sci. 2021, 22, 5344. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105344
AMA Style
Rebustini IT, Bernardo-Colón A, Nalvarte AI, Becerra SP. Delivery Systems of Retinoprotective Proteins in the Retina. International Journal of Molecular Sciences. 2021; 22(10):5344. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105344
Chicago/Turabian StyleRebustini, Ivan T., Alexandra Bernardo-Colón, Alejandra Isasi Nalvarte, and S. Patricia Becerra. 2021. "Delivery Systems of Retinoprotective Proteins in the Retina" International Journal of Molecular Sciences 22, no. 10: 5344. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105344
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.