
Double-Edged Sword of Vitamin D3 Effects on Primary Neuronal Cultures in Hypoxic States

 , and
, and
Abstract
:1. Introduction
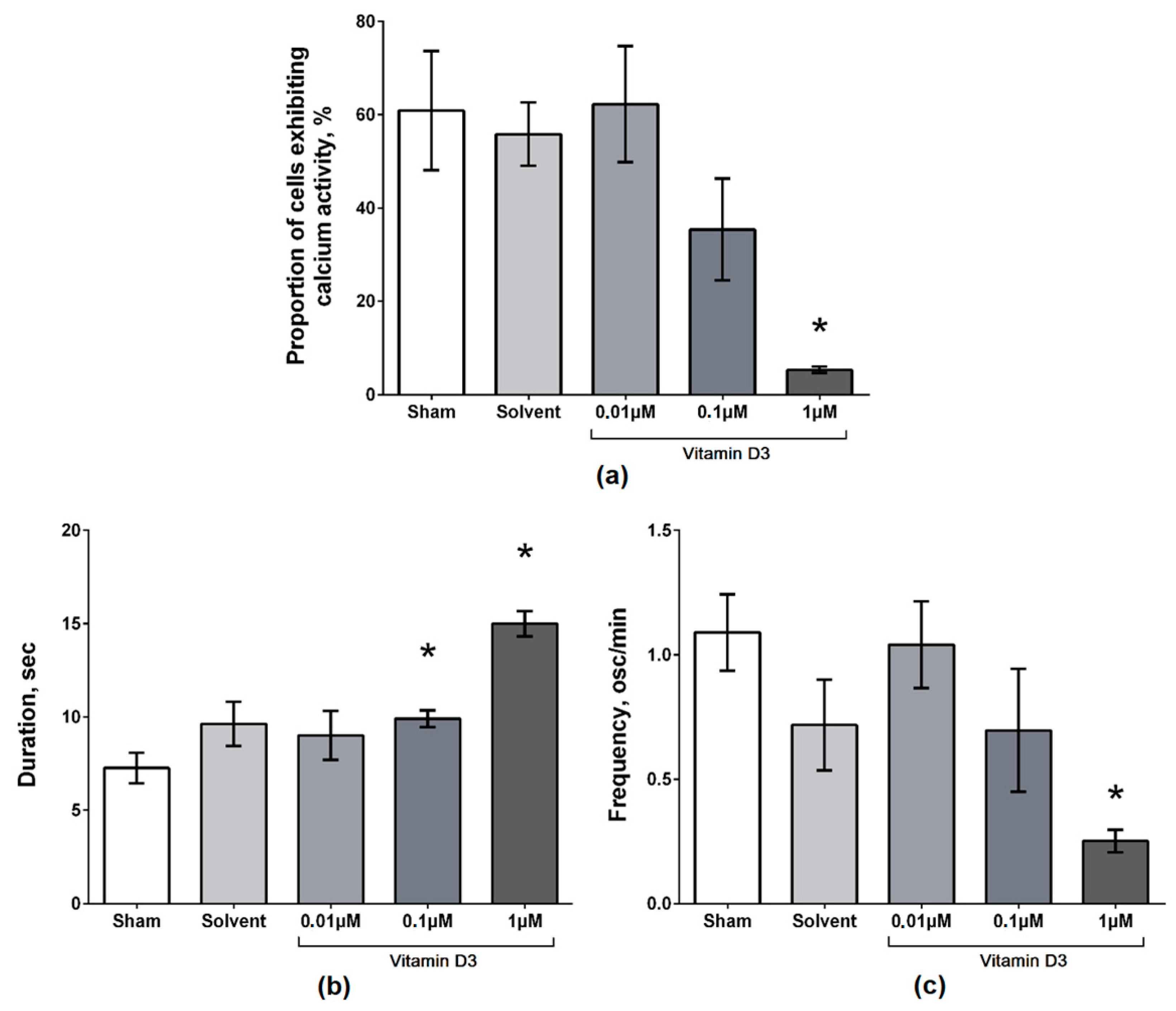
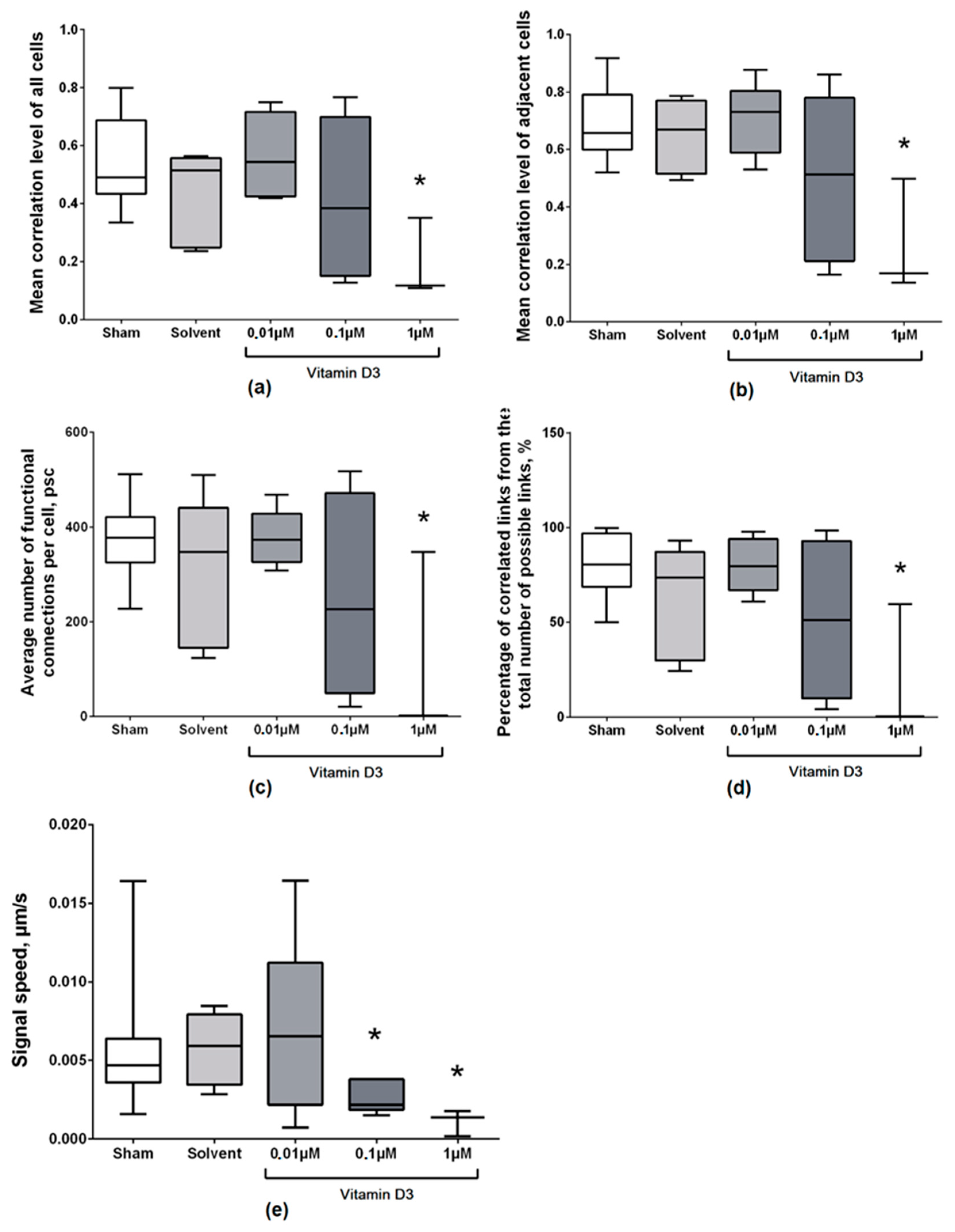
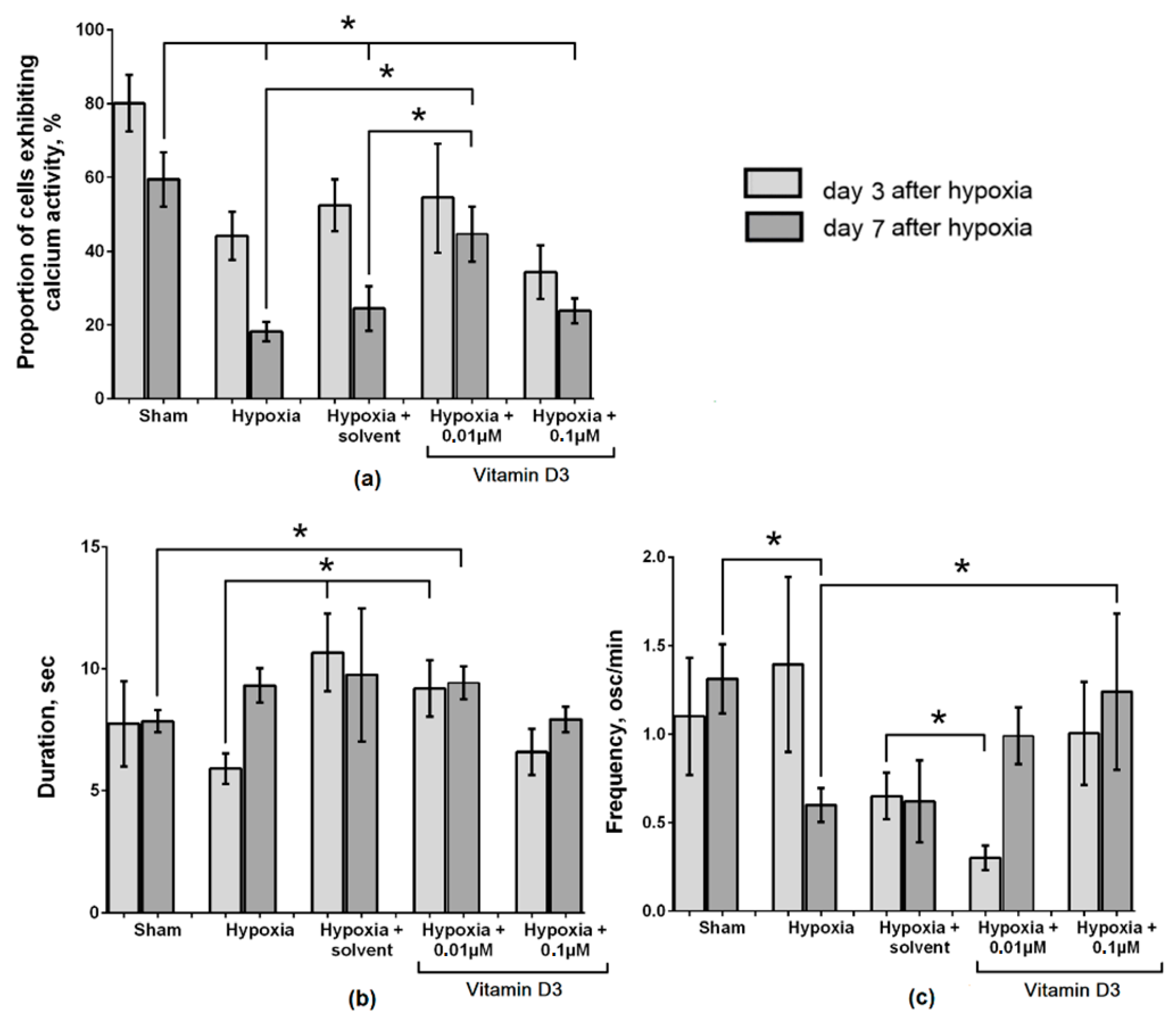
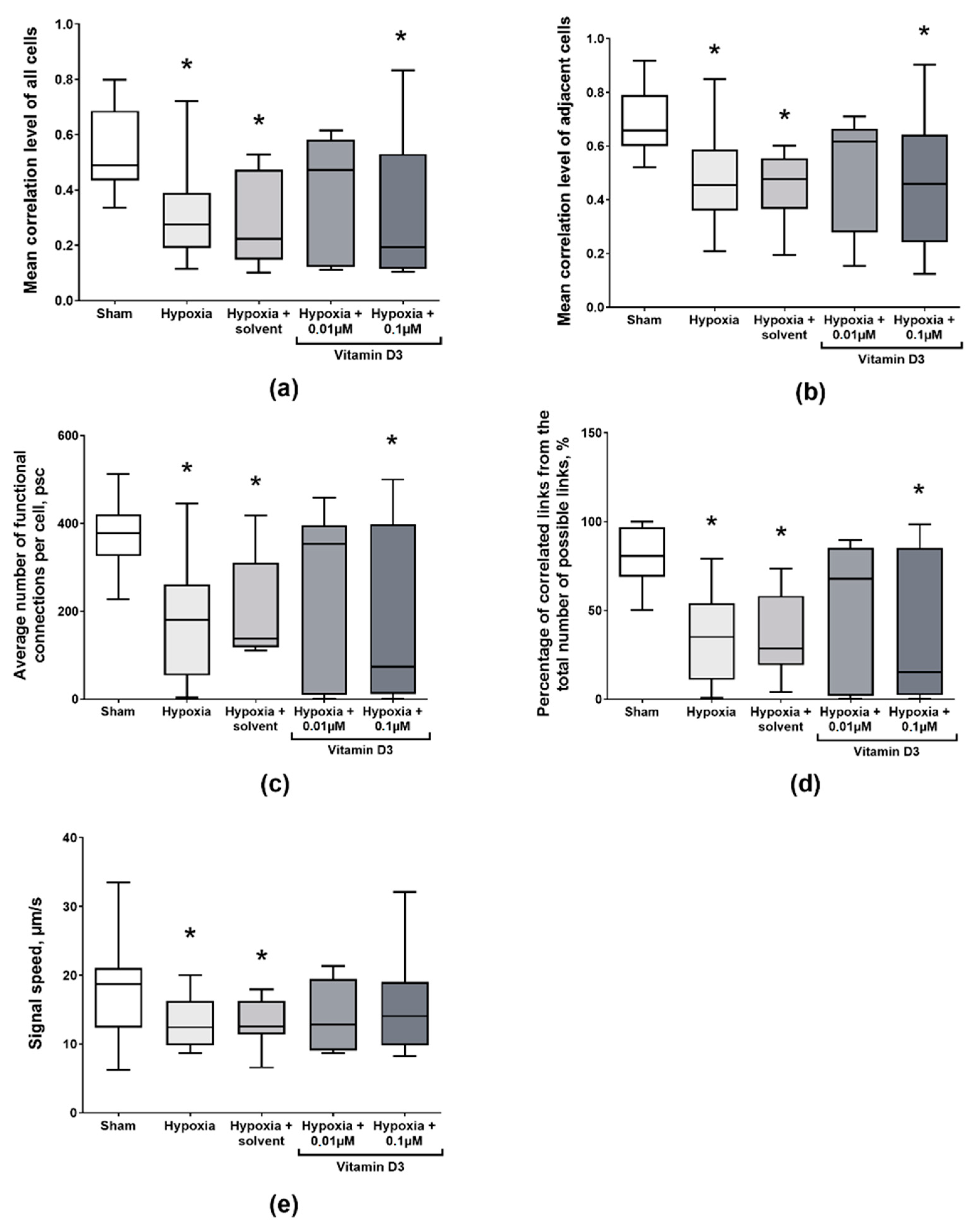
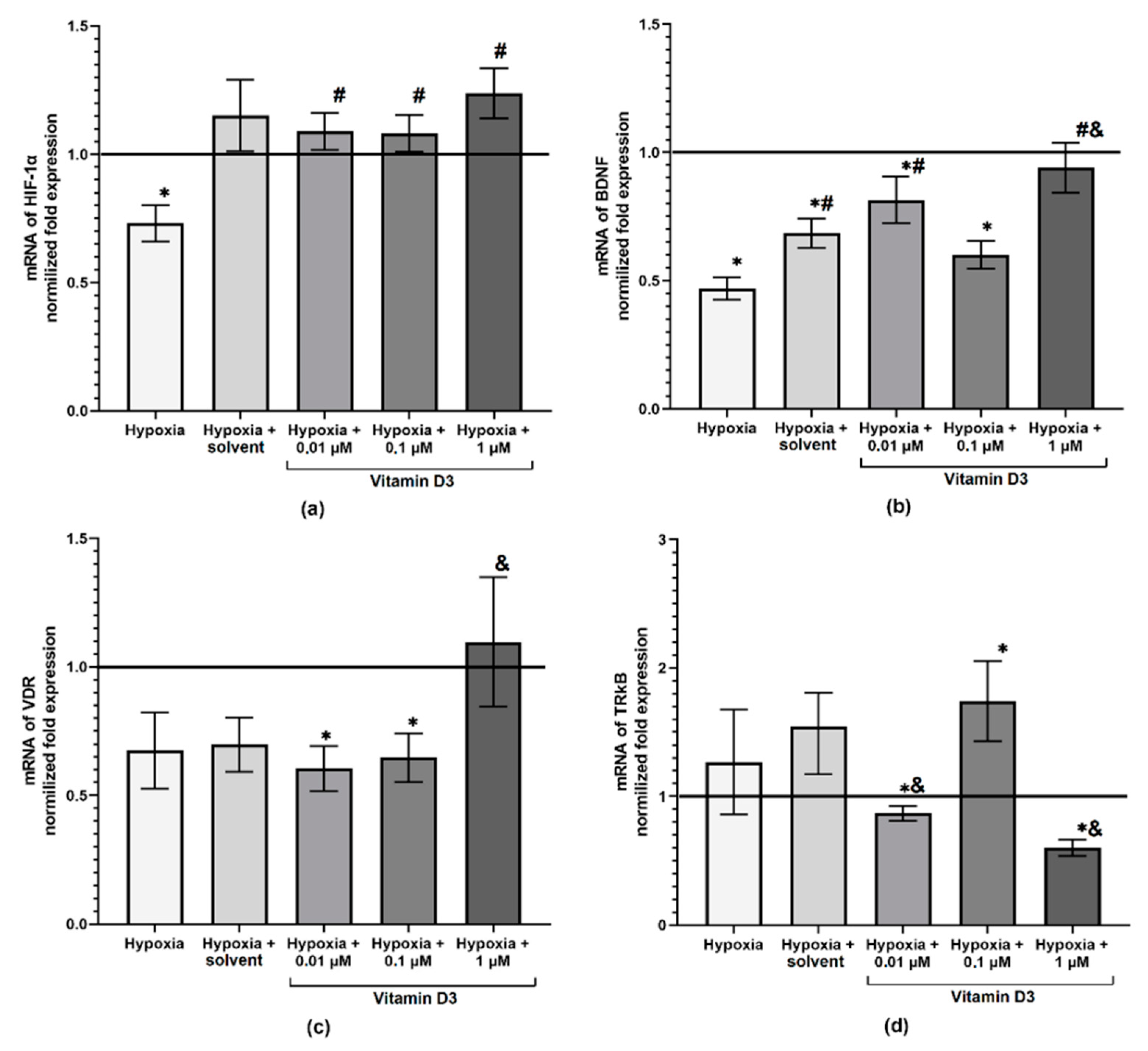
2. Results
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Experimental Design
4.3. Isolation of the Primary Neuronal Cultures
4.4. In Vitro Hypoxia Model
4.5. Cell Viability Analysis
4.6. Calcium Imaging
4.7. Analysis of Network Characteristics of Primary Neuronal Cultures Activity
4.8. RNA Extraction and RT-qPCR
- The following pairs of primers were used:
- Oaz1_fw5′-AAGGACAGTTTTGCAGCTCTCC-3′;
- Oaz1_rv5′-TCTGTCCTCACGGTTCTTGGG-3′;
- Hif1a_fw15′GCAATTCTCCAAGCCCTCCAAG-3′;
- Hif1a_rv15′-TTCATCAGTGGTGGCAGTTGTG-3′;
- mBDNF_fw 5′CCCAACGAAGAAAACCATAAGGA-3′;
- mBDNF_rv 5′-CCAGCAGAAAGAGTAGAGGAGGCT-3′;
- VDR_fw 5′-ATGCCCACCACAAGACCTACGAC-3′;
- VDR_rv 5′-AGT CTCCGGAGA AGCTGAGTGTGG-3′;
- TrkB_fw15′-TTTCCGCCACCTTGACTTGTCT-3′;
- TrkB_rv15′-GTCGGGGCTGGATTTAGTCTCC-3′;
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
- L-type channel _Cav 1.2 (1C) _fw 5′-GAGGAGTTGGACAAGGCTATGAAGGA-3′
- L-type channel _Cav 1.2 (1C) _rv 5′-TTGCCCCTGCTGTCACTCTGATAGTA-3′
- L-type channel _Cav 1.3 (1D) _rv 5′-CACCAGCACCAGAGAGTTCCACAG-3′
- L-type channel _Cav 1.3 (1D) _fw 5′-AGAAGACATCGATCCCGAGAACGA G-3′
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miyamoto, O.; Auer, R.N. Hypoxia, hyperoxia, ischemia, and brain necrosis. Neurology 2000, 54, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Radak, D.; Katsiki, N.; Resanovic, I.; Jovanovic, A.; Sudar-Milovanovic, E.; Zafirovic, S.; Mousad, S.A.; Isenovic, E.R. Apoptosis and Acute Brain Ischemia in Ischemic Stroke. Curr. Vasc. Pharmacol. 2017, 15, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Xue, H.; Xu, Y.; Niu, J.; Zhao, P. Sevoflurane Postconditioning Inhibits Autophagy through Activation of the Extracellular Signal-Regulated Kinase Cascade, Alleviating Hypoxic-Ischemic Brain Injury in Neonatal Rats. Neurochem. Res. 2019, 44, 347–356. [Google Scholar] [CrossRef]
- Chen, R.; Lai, U.H.; Zhu, L.; Singh, A.; Ahmed, M.; Forsyth, N.R. Reactive Oxygen Species Formation in the Brain at Different Oxygen Levels: The Role of Hypoxia Inducible Factors. Front. Cell Dev. Biol. 2018, 6, 132. [Google Scholar] [CrossRef] [Green Version]
- Stessman, L.E.; Peeples, E.S. Vitamin D and Its Role in Neonatal Hypoxic-Ischemic Brain Injury. Neonatology 2018, 113, 305–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Yuan, J.; Wang, J.; Cui, C.; Jiang, P. Calcitriol alleviates global cerebral ischemia-induced cognitive impairment by reducing apoptosis regulated by VDR/ERK signaling pathway in rat hippocampus. Brain Res. 2019, 1724, 146430. [Google Scholar] [CrossRef]
- Evans, M.A.; Kim, H.A.; Ling, Y.H.; Uong, S.; Vinh, A.; De Silva, T.M.; Arumugam, T.V.; Clarkson, A.N.; Zosky, G.R.; Drummond, G.R.; et al. Vitamin D3 Supplementation Reduces Subsequent Brain Injury and Inflammation Associated with Ischemic Stroke. NeuroMol. Med. 2018, 20, 147–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krisanova, N.; Pozdnyakova, N.; Pastukhov, A.; Dudarenko, M.; Maksymchuk, O.; Parkhomets, P.; Sivko, R.; Borisova, T. Vitamin D3 deficiency in puberty rats causes presynaptic malfunctioning through alterations in exocytotic release and uptake of glutamate/GABA and expression of EAAC-1/GAT-3 transporters. Food Chem. Toxicol. 2019, 123, 142–150. [Google Scholar] [CrossRef]
- Mutlu, M.; Sarıaydın, M.; Aslan, Y.; Şebnem, K.; Dereci, S.; Kart, C.; Yaman, S.Ö.; Kural, B. Status of vitamin D, antioxidant enzymes, and antioxidant substances in neonates with neonatal hypoxic-ischemic encephalopathy. J. Matern. Neonatal Med. 2016, 29, 2259–2263. [Google Scholar] [CrossRef]
- Lowe, D.W.; Fraser, J.L.; Rollins, L.G.; Bentzley, J.; Nie, X.; Martin, R.; Singh, I.; Jenkins, D. Vitamin D improves functional outcomes in neonatal hypoxic ischemic male rats treated with N -acetylcysteine and hypothermia. Neuropharmacology 2017, 123, 186–200. [Google Scholar] [CrossRef]
- Bao, G.-Q.; Yu, J.-Y. Vitamin D3 promotes cerebral angiogenesis after cerebral infarction in rats by activating Shh signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7069–7077. [Google Scholar]
- Wajda, J.; Świat, M.; Owczarek, A.J.; Brzozowska, A.; Olszanecka-Glinianowicz, M.; Chudek, J. Severity of Vitamin D Deficiency Predicts Mortality in Ischemic Stroke Patients. Dis. Markers 2019, 2019, 3652894. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Guo, X.; Liu, Z.; Zhao, X.; Feng, Y.; Song, S.; Cui, C.; Jiang, P. Vitamin D receptor activation influences the ERK pathway and protects against neurological deficits and neuronal death. Int. J. Mol. Med. 2018, 41, 364–372. [Google Scholar] [CrossRef]
- Balamurugan, K. HIF-1 at the crossroads of hypoxia, inflammation, and cancer. Int. J. Cancer 2016, 138, 1058–1066. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Sharma, G.; Mishra, V.; Raghubir, R. Hypoxia Inducible Factor-1: Its Potential Role in Cerebral Ischemia. Cell. Mol. Neurobiol. 2012, 32, 491–507. [Google Scholar] [CrossRef]
- Benderro, G.F.; Sun, X.; Kuang, Y.; Lamanna, J.C. Decreased VEGF expression and microvascular density, but increased HIF-1 and 2α accumulation and EPO expression in chronic moderate hyperoxia in the mouse brain. Brain Res. 2012, 1471, 46–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Qu, Y.; Li, J.; Xiong, Y.; Mao, M.; Mu, D. Relationship between HIF-1α expression and neuronal apoptosis in neonatal rats with hypoxia–ischemia brain injury. Brain Res. 2007, 1180, 133–139. [Google Scholar] [CrossRef]
- Kulie, T.; Groff, A.; Redmer, J.; Hounshell, J.; Schrager, S. Vitamin D: An Evidence-Based Review. J. Am. Board Fam. Med. 2009, 22, 698–706. [Google Scholar] [CrossRef] [Green Version]
- Bikle, D.D. Vitamin D: An ancient hormone. Exp. Dermatol. 2011, 20, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Gil, Á.; Plaza-Diaz, J.; Mesa, M.D. Vitamin D: Classic and Novel Actions. Ann. Nutr. Metab. 2018, 72, 87–95. [Google Scholar] [CrossRef]
- AlJohri, R.; Alokail, M.; Haq, S.H. Neuroprotective role of vitamin D in primary neuronal cortical culture. eNeurologicalSci 2019, 14, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Shipton, E.A.; Shipton, E.E. Vitamin D and Pain: Vitamin D and Its Role in the Aetiology and Maintenance of Chronic Pain States and Associated Comorbidities. Pain Res. Treat. 2015, 19, 904967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veldurthy, V.; Wei, R.; Oz, L.; Dhawan, P.; Jeon, Y.H.; Christakos, S. Vitamin D, calcium homeostasis and aging. Bone Res. 2016, 4, 16041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bivona, G.; Gambino, C.M.; Iacolino, G.; Ciaccio, M. Vitamin D and the nervous system. Neurol. Res. 2019, 41, 827–835. [Google Scholar] [CrossRef]
- Holick, M.F. The vitamin D deficiency pandemic: Approaches for diagnosis, treatment and prevention. Rev. Endocr. Metab. Disord. 2017, 18, 153–165. [Google Scholar] [CrossRef] [PubMed]
- DeLuca, H.F. Overview of general physiologic features and functions of vitamin D. Am. J. Clin. Nutr. 2004, 80, 1689S–1696S. [Google Scholar] [CrossRef] [Green Version]
- Borel, P.; Desmarchelier, C. Bioavailability of Fat-Soluble Vitamins and Phytochemicals in Humans: Effects of Genetic Variation. Annu. Rev. Nutr. 2018, 38, 69–96. [Google Scholar] [CrossRef] [Green Version]
- Allgrove, J. Disorders of calcium metabolism. Curr. Paediatr. 2003, 13, 529–535. [Google Scholar] [CrossRef]
- Caballero-Villarraso, J.; Jiménez-Jiménez, M.J.; Escribano, B.M.; Agüera, E.; Santamaría, A.; Túnez, I. Implications of Vitamin D in Multiple Sclerosis and Other Neurodegenerative Processes: Bibliometric Analysis and Systematic Review. CNS Neurol. Disord. Drug Targets 2019, 18, 478–490. [Google Scholar] [CrossRef]
- Koduah, P.; Paul, F.; Dörr, J.-M. Vitamin D in the prevention, prediction and treatment of neurodegenerative and neuroinflammatory diseases. EPMA J. 2017, 8, 313–325. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, E.; Jørgensen, T.N. Immunological effects of vitamin D and their relations to autoimmunity. J. Autoimmun. 2019, 100, 7–16. [Google Scholar] [CrossRef]
- Tian, H.-Q.; Cheng, L. The role of vitamin D in allergic rhinitis. Asia Pac. Allergy 2017, 7, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Lerchbaum, E.; Obermayer-Pietsch, B. MECHANISMS IN ENDOCRINOLOGY: Vitamin D and fertility: A systematic review. Eur. J. Endocrinol. 2012, 166, 765–778. [Google Scholar] [CrossRef]
- Altieri, B.; Muscogiuri, G.; Barrea, L.; Mathieu, C.; Vallone, C.V.; Mascitelli, L.; Bizzaro, G.; Altieri, V.M.; Tirabassi, G.; Balercia, G.; et al. Does vitamin D play a role in autoimmune endocrine disorders? A proof of concept. Rev. Endocr. Metab. Disord. 2017, 18, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Vuolo, L.; Di Somma, C.; Faggiano, A.; Colao, A.A. Vitamin D and Cancer. Front. Endocrinol. 2012, 3, 58. [Google Scholar] [CrossRef] [Green Version]
- Sacerdote, A.; Dave, P.; Lokshin, V.; Bahtiyar, G. Type 2 Diabetes Mellitus, Insulin Resistance, and Vitamin D. Curr. Diabetes Rep. 2019, 19, 101. [Google Scholar] [CrossRef] [PubMed]
- Rai, V.; Agrawal, D.K. Role of Vitamin D in Cardiovascular Diseases. Endocrinol. Metab. Clin. N. Am. 2017, 46, 1039–1059. [Google Scholar] [CrossRef] [PubMed]
- Gois, P.H.F.; Ferreira, D.; Olenski, S.; Seguro, A.C. Vitamin D and Infectious Diseases: Simple Bystander or Contributing Factor? Nutrients 2017, 9, 651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedotova, J.O. Vitamin D3 treatment differentially affects anxiety-like behavior in the old ovariectomized female rats and old ovariectomized female rats treated with low dose of 17β-estradiol. BMC Med. Genet. 2019, 20, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulkrane, M.S.; Fedotova, J.; Kolodyaznaya, V.; Micale, V.; Drago, F.; van den Tol, A.J.M.; Baranenko, D. Vitamin D and Depression in Women: A Mini-review. Curr. Neuropharmacol. 2020, 18, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Fedotova, J.; Pivina, S.; Sushko, A. Effects of Chronic Vitamin D3 Hormone Administration on Anxiety-Like Behavior in Adult Female Rats after Long-Term Ovariectomy. Nutrients 2017, 9, 28. [Google Scholar] [CrossRef]
- Alfieri, D.F.; Lehmann, M.F.; Oliveira, S.R.; Flauzino, T.; Delongui, F.; De Araújo, M.C.M.; Dichi, I.; Delfino, V.D.; Mezzaroba, L.; Simão, A.N.C.; et al. Vitamin D deficiency is associated with acute ischemic stroke, C-reactive protein, and short-term outcome. Metab. Brain Dis. 2017, 32, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, Y.; Huang, G.; Zhu, J.; Feng, W.; He, J. Association between vitamin D status and cognitive impairment in acute ischemic stroke patients: A prospective cohort study. Clin. Interv. Aging 2018, 13, 2503–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.-Y.; Chung, P.-W.; Kim, Y.B.; Moon, H.-S.; Suh, B.-C.; Won, Y.S.; Kim, J.-M.; Youn, Y.C.; Kwon, O.-S. Serum Vitamin D Status as a Predictor of Prognosis in Patients with Acute Ischemic Stroke. Cerebrovasc. Dis. 2015, 40, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Galior, K.; Grebe, S.; Singh, R. Development of Vitamin D Toxicity from Overcorrection of Vitamin D Deficiency: A Review of Case Reports. Nutrients 2018, 10, 953. [Google Scholar] [CrossRef] [Green Version]
- Mayne, P.E.; Burne, T.H. Vitamin D in Synaptic Plasticity, Cognitive Function, and Neuropsychiatric Illness. Trends Neurosci. 2019, 42, 293–306. [Google Scholar] [CrossRef]
- Sharapov, M.G.; Gudkov, S.V. Peroxiredoxin 1—Multifunctional antioxidant enzyme, protects from oxidative damages and increases the survival rate of mice exposed to total body irradiation. Arch. Biochem. Biophys. 2021, 697, 108671. [Google Scholar] [CrossRef]
- Lösel, R.; Wehling, M. Nongenomic actions of steroid hormones. Nat. Rev. Mol. Cell Biol. 2003, 4, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Brewer, L.D.; Thibault, V.; Chen, K.-C.; Langub, M.C.; Landfield, P.W.; Porter, N.M. Vitamin D Hormone Confers Neuroprotection in Parallel with Downregulation of L-Type Calcium Channel Expression in Hippocampal Neurons. J. Neurosci. 2001, 21, 98–108. [Google Scholar] [CrossRef]
- Ibi, M.; Sawada, H.; Nakanishi, M.; Kume, T.; Katsuki, H.; Kaneko, S.; Shimohama, S.; Akaike, A. Protective effects of 1α,25-(OH)2D3 against the neurotoxicity of glutamate and reactive oxygen species in mesencephalic culture. Neuropharmacology 2001, 40, 761–771. [Google Scholar] [CrossRef]
- Cui, P.; Wang, Y.; Li, Y.; Ge, L. Vitamin D Attenuates Hypoxia-Induced Injury in Rat Primary Neuron Cells through Downregulation of the Dual Oxidase 1 (DUOX1) Gene. Med. Sci. Monit. 2020, 26, e925350. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Xue, R.; Gu, J.; Xiao, Y.; Zhong, H.; Pan, X.; Ran, R. Neuroprotective effect of calcitriol on ischemic/reperfusion injury through the NR3A/CREB pathways in the rat hippocampus. Mol. Med. Rep. 2013, 8, 1708–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khairy, E.Y.; Attia, M.M. Protective effects of vitamin D on neurophysiologic alterations in brain aging: Role of brain-derived neurotrophic factor (BDNF). Nutr. Neurosci. 2019, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Koshkina, A.; Dudnichenko, T.; Baranenko, D.; Fedotova, J.; Drago, F. Effects of Vitamin D3 in Long-Term Ovariectomized Rats Subjected to Chronic Unpredictable Mild Stress: BDNF, NT-3, and NT-4 Implications. Nutrients 2019, 11, 1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gezen-Ak, D.; Dursun, E.; Yilmazer, S. The Effect of Vitamin D Treatment on Nerve Growth Factor (NGF) Release from Hippocampal Neurons. Noro Psikiyatr. Ars. 2014, 51, 157–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vedunova, M.; Sakharnova, T.; Mitroshina, E.; Perminova, M.; Pimashkin, A.; Zakharov, Y.; Dityatev, A.; Mukhina, I. Seizure-like activity in hyaluronidase-treated dissociated hippocampal cultures. Front. Cell. Neurosci. 2013, 7, 149. [Google Scholar] [CrossRef] [Green Version]
- Vedunova, M.V.; Mishchenko, T.A.; Mitroshina, E.V.; Mukhina, I.V. TrkB-Mediated Neuroprotective and Antihypoxic Properties of Brain-Derived Neurotrophic Factor. Oxid. Med. Cell Longev. 2015, 2015, 453901. [Google Scholar] [CrossRef] [PubMed]
- Zakharov, Y.N.; Mitroshina, E.V.; Shirokova, O.; Mukhina, I.V. Calcium Transient Imaging as Tool for Neuronal and Glial Network Interaction Study; Springer: New York, NY, USA, 2013; pp. 225–232. [Google Scholar]
- Kustikova, V.; Krivonosov, M.; Pimashkin, A.; Denisov, P.; Zaikin, A.; Ivanchenko, M.; Meyerov, I.; Semyanov, A. CalciumCV: Computer Vision Software for Calcium Signaling in Astrocytes; Springer International Publishing: Cham, Switzerland, 2018; pp. 168–179. [Google Scholar]
- Savyuk, M.; Krivonosov, M.; Mishchenko, T.; Gazaryan, I.; Ivanchenko, M.; Khristichenko, A.; Poloznikov, A.; Hushpulian, D.; Nikulin, S.; Tonevitsky, E.; et al. Neuroprotective Effect of HIF Prolyl Hydroxylase Inhibition in an In Vitro Hypoxia Model. Antioxidants 2020, 9, 662. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Number of Viable Cells, % |
|---|---|
| Sham | 93.91 ± 1.24 |
| Solvent | 89.34 ± 2.21 |
| Vitamin D3 0.01 µM | 89.99 ± 1.98 |
| Vitamin D3 0.1 µM | 91.97 ± 1.86 |
| Vitamin D3 1 µM | 88.64 ± 3.09 |
| Group | Number of Viable Cells in the Post-Hypoxic Period, % | |
|---|---|---|
| Day 3 | Day 7 | |
| Sham | 96.88 ± 0.49 | 96.32 ± 0.53 |
| Hypoxia | 78.06 ± 2.82 * | 80.39 ± 2.17 * |
| Hypoxia + solvent | 81.52 ± 1.89 * | 80.91 ± 2.37 * |
| Hypoxia + vitamin D3 0.01 µM | 95.14 ± 0.7 * | 89.23 ± 0.99 *# |
| Hypoxia + vitamin D3 0.1 µM | 93.58 ± 1.1 *# | 86.89 ± 0.71 *# |
| Hypoxia + vitamin D3 1 µM | 8.89 ± 2.6 *# | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loginova, M.; Mishchenko, T.; Savyuk, M.; Guseva, S.; Gavrish, M.; Krivonosov, M.; Ivanchenko, M.; Fedotova, J.; Vedunova, M. Double-Edged Sword of Vitamin D3 Effects on Primary Neuronal Cultures in Hypoxic States. Int. J. Mol. Sci. 2021, 22, 5417. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115417
Loginova M, Mishchenko T, Savyuk M, Guseva S, Gavrish M, Krivonosov M, Ivanchenko M, Fedotova J, Vedunova M. Double-Edged Sword of Vitamin D3 Effects on Primary Neuronal Cultures in Hypoxic States. International Journal of Molecular Sciences. 2021; 22(11):5417. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115417
Chicago/Turabian StyleLoginova, Maria, Tatiana Mishchenko, Maria Savyuk, Svetlana Guseva, Maria Gavrish, Mikhail Krivonosov, Mikhail Ivanchenko, Julia Fedotova, and Maria Vedunova. 2021. "Double-Edged Sword of Vitamin D3 Effects on Primary Neuronal Cultures in Hypoxic States" International Journal of Molecular Sciences 22, no. 11: 5417. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115417