
Simulated Microgravity Remodels Extracellular Matrix of Osteocommitted Mesenchymal Stromal Cells



Abstract
:1. Introduction
2. Results
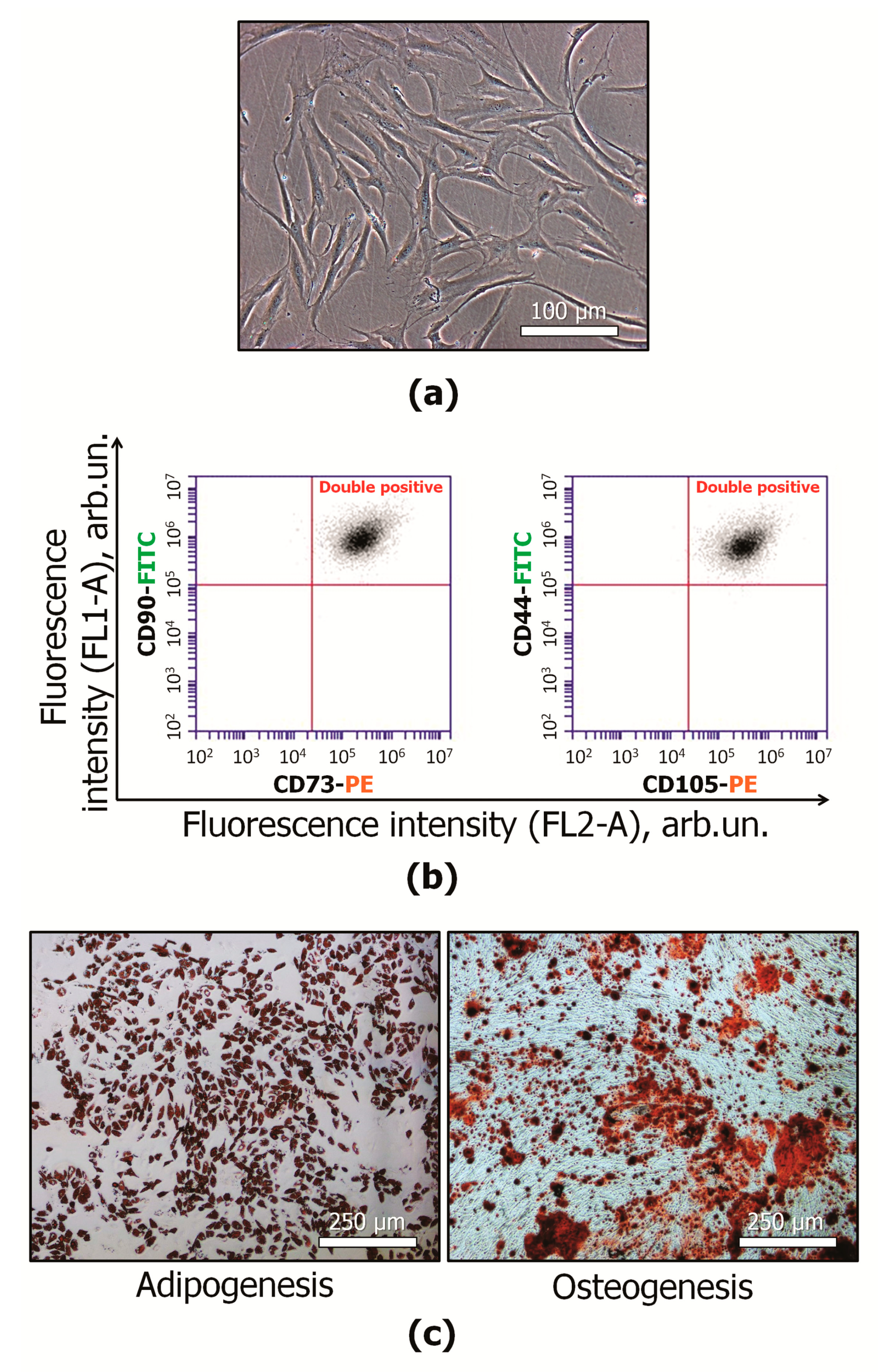
2.1. MSC Characterization
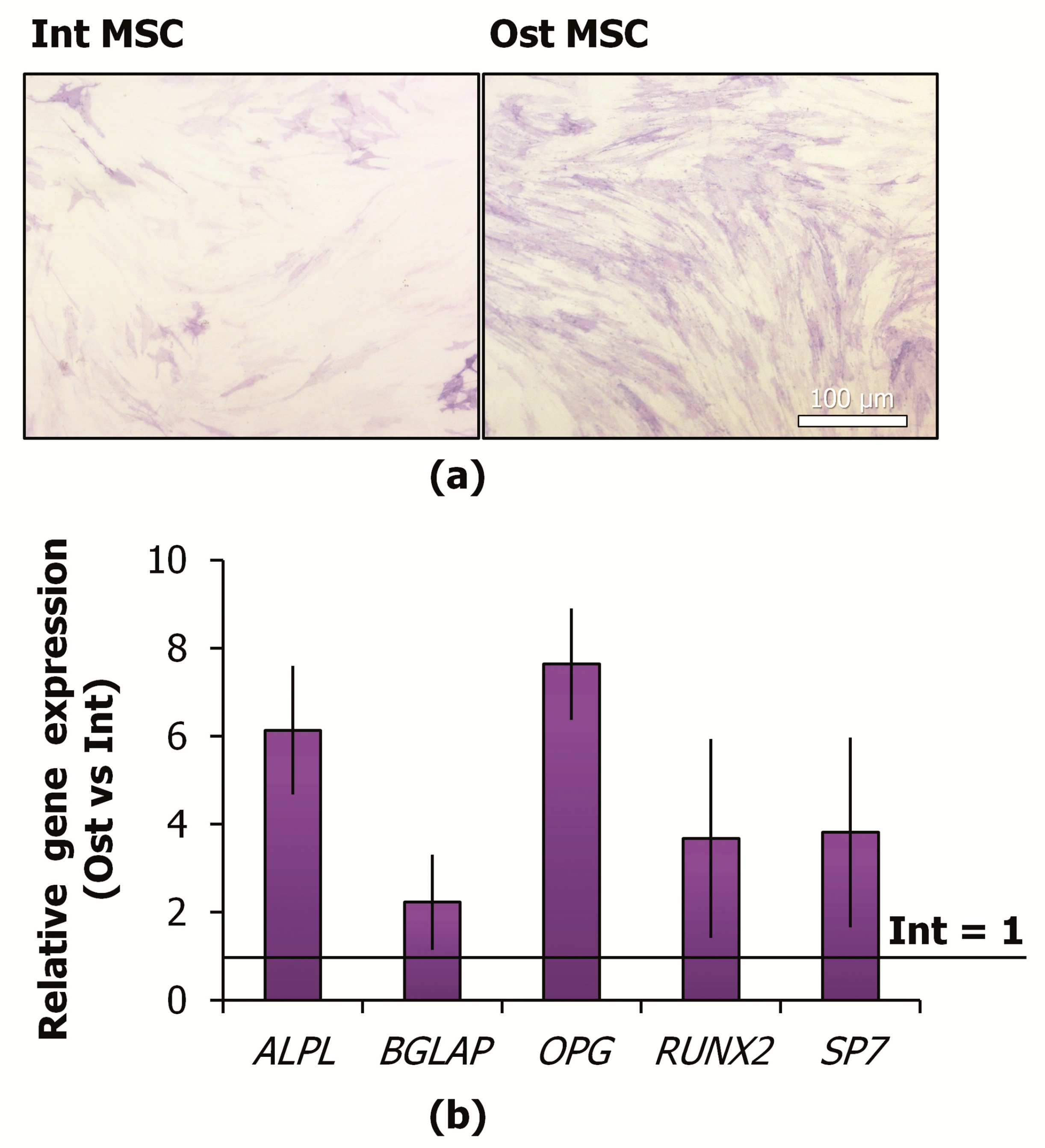
2.2. MSC Commitment to the Osteoblast Lineage
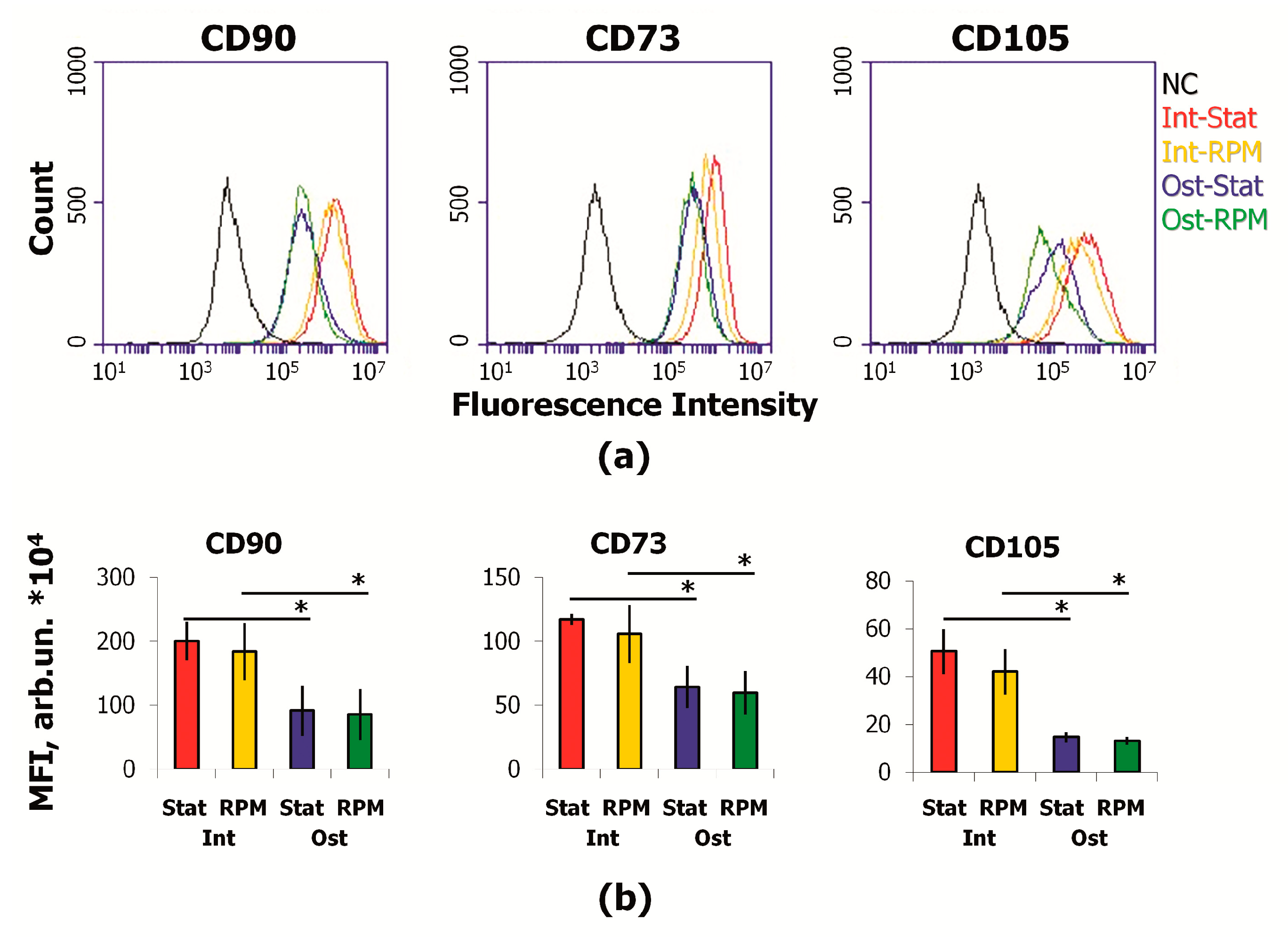
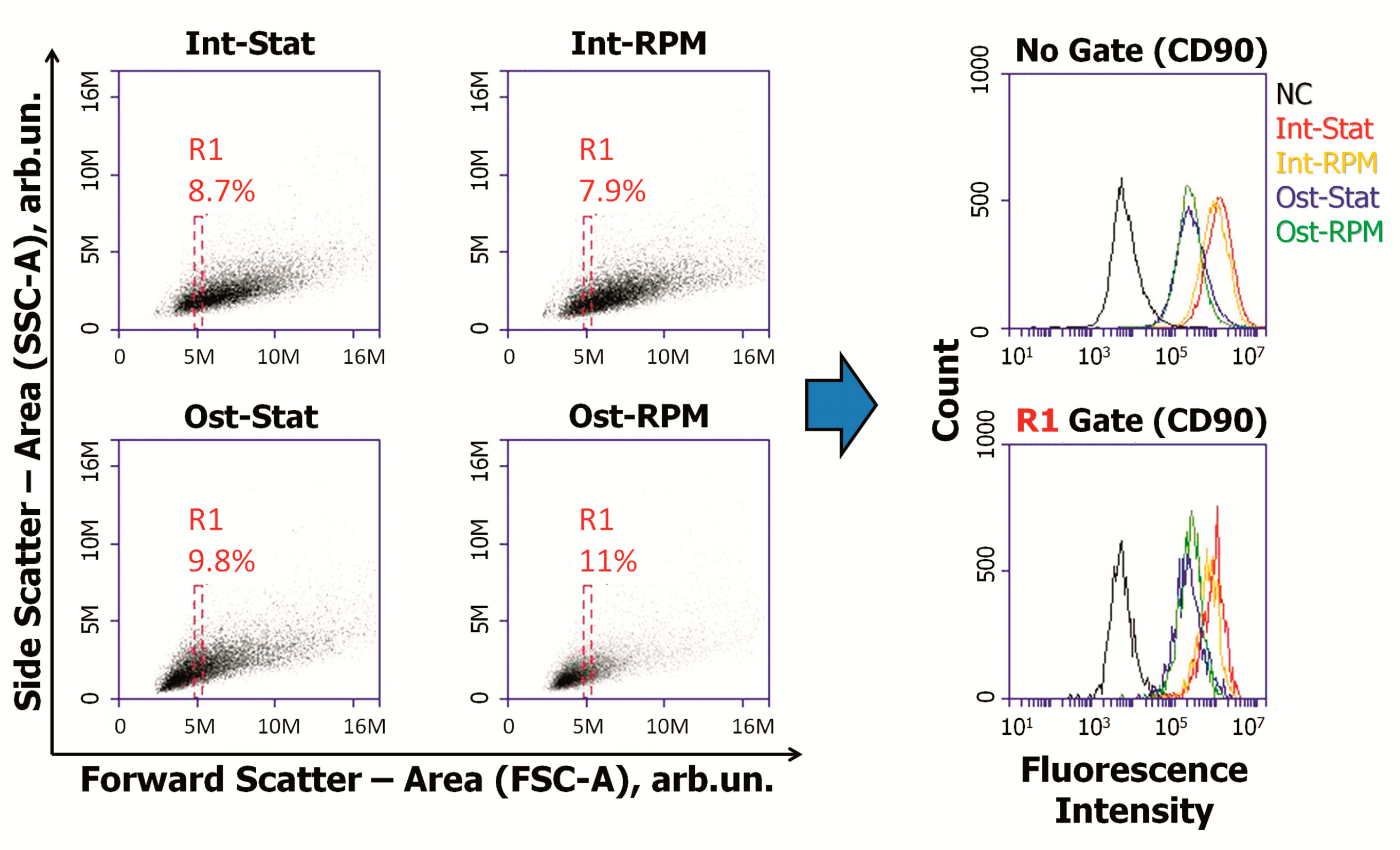
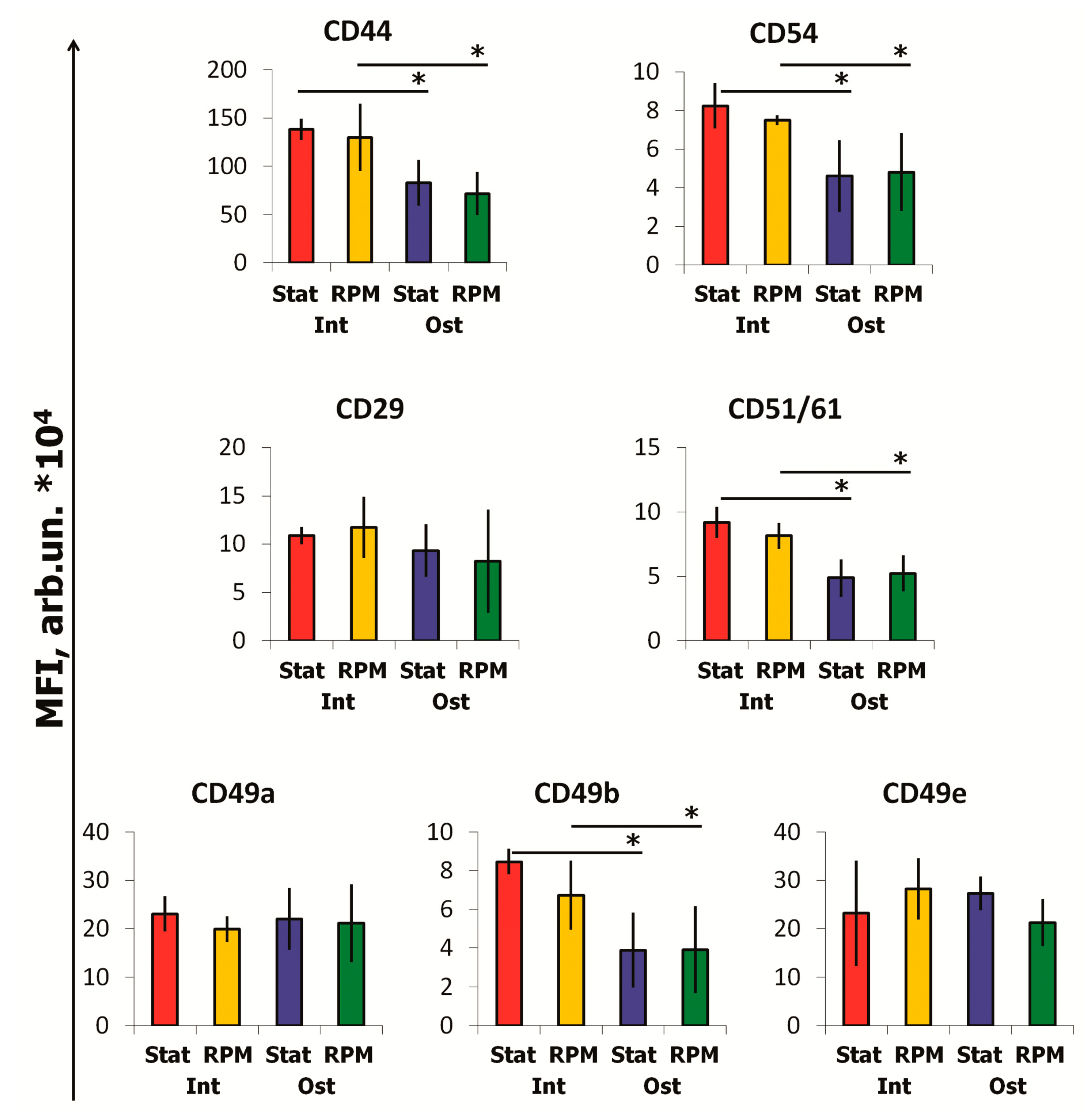
2.3. SMG Effects on Immunophenotype and Adhesion Molecules
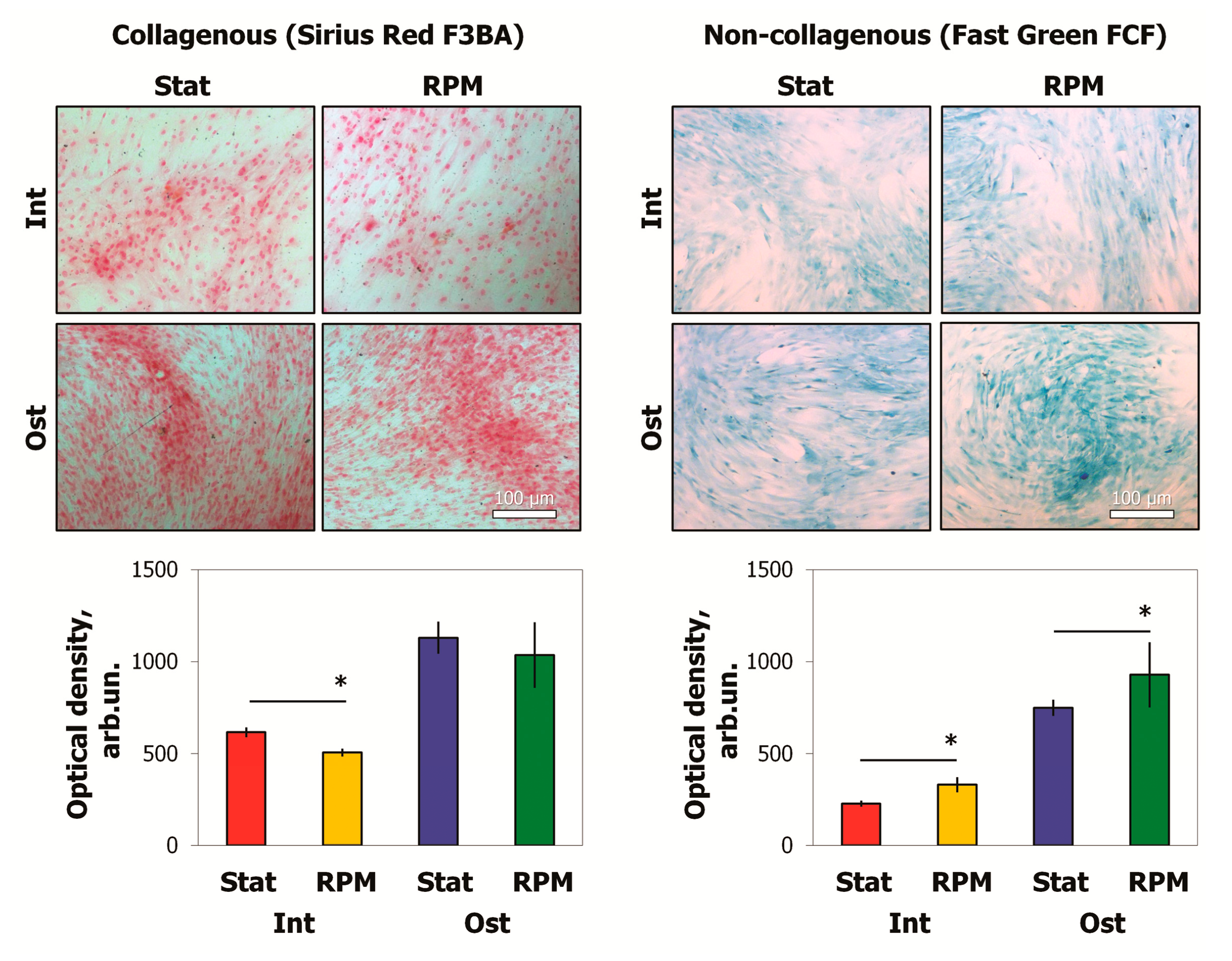
2.4. SMG Effects on ECM Proteins
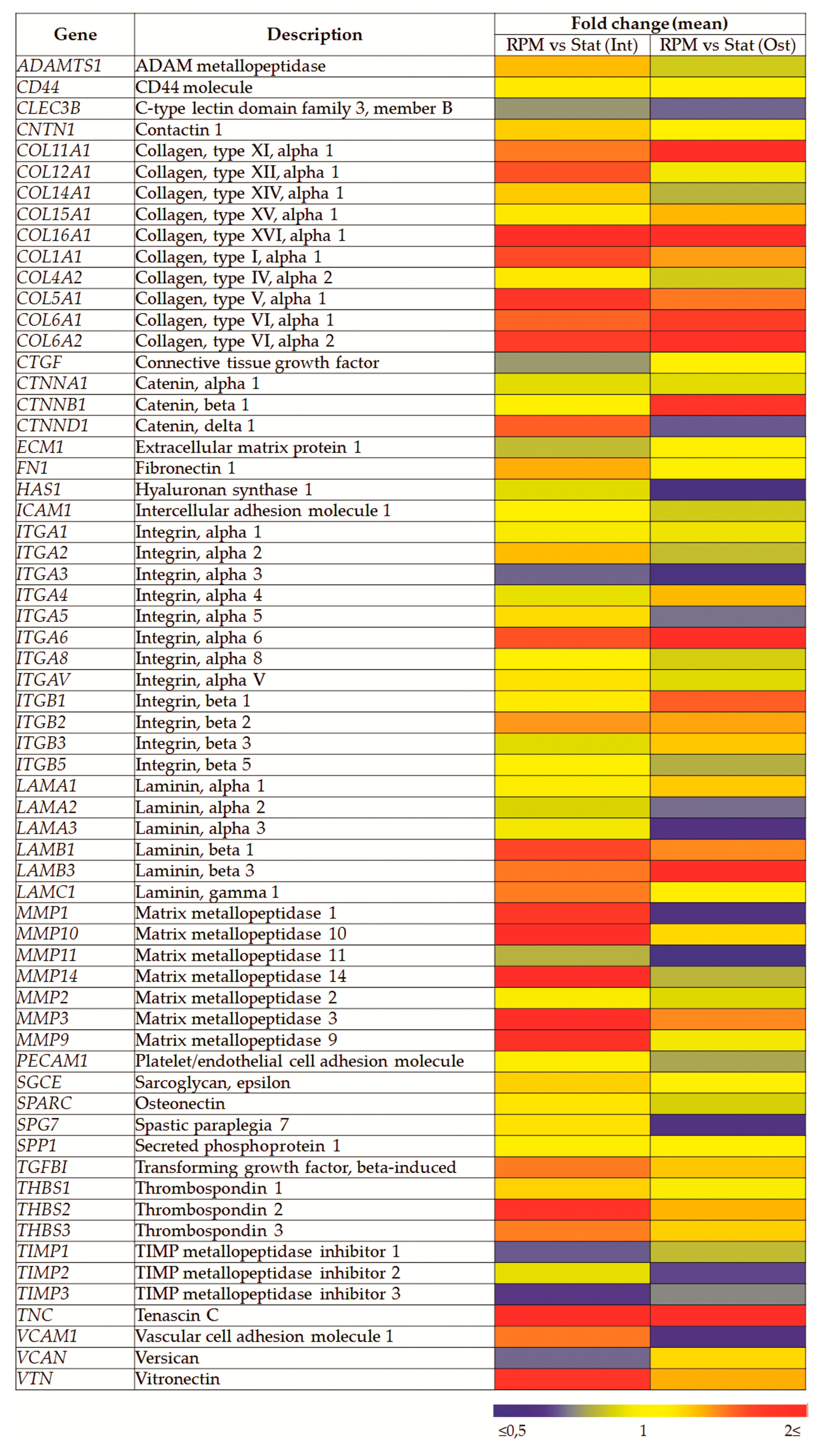
2.5. SMG Effects on Gene Expression
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kadler, K.E.; Baldock, C.; Bella, J.; Boot-Handford, R.P. Collagens at a glance. J. Cell Sci. 2007, 120, 1955–1958. [Google Scholar] [CrossRef] [Green Version]
- Bülow, H.E.; Hobert, O. The Molecular Diversity of Glycosaminoglycans Shapes Animal Development. Annu. Rev. Cell Dev. Biol. 2006, 22, 375–407. [Google Scholar] [CrossRef] [PubMed]
- Hyldig, K.; Riis, S.; Pennisi, C.P.; Zachar, V.; Fink, T. Implications of Extracellular Matrix Production by Adipose Tissue-Derived Stem Cells for Development of Wound Healing Therapies. Int. J. Mol. Sci. 2017, 18, 1167. [Google Scholar] [CrossRef] [Green Version]
- Sarathchandra, P.; Cassella, J.P.; Ali, S.Y. Ultrastructural localization of proteoglycans in bone in osteogenesis imperfecta as demonstrated by Cuprolinic Blue staining. J. Bone Miner. Metab. 2002, 20, 288–293. [Google Scholar] [CrossRef]
- Argentati, C.; Morena, F.; Tortorella, I.; Bazzucchi, M.; Porcellati, S.; Emiliani, C.; Martino, S. Insight into Mechanobiology: How stem cells feel mechanical forces and orchestrate biobical functions. Int. J. Mol. Sci. 2019, 20, 5337. [Google Scholar] [CrossRef] [Green Version]
- Mott, J.D.; Werb, Z. Regulation of matrix biology by matrix metalloproteinases. Curr. Opin. Cell Biol. 2004, 16, 558–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oganov, V.S.; Bakulin, A.V.; Novikov, V.E.; Kabitskaia, O.E.; Murashko, L.M. Characteristics and patterns of the human bone reactions to microgravity. Aerosp. Environ. Med. 2006, 40, 15–21. [Google Scholar]
- Grimm, D.; Grosse, J.; Wehland, M.; Mann, V.; Reseland, J.E.; Sundaresan, A.; Corydon, T.J. The impact of microgravity on bone in humans. Bone 2016, 87, 44–56. [Google Scholar] [CrossRef]
- Stavnichuk, M.; Mikolajewicz, N.; Corlett, T.; Morris, M.; Komarova, S.V. A systematic review and meta-analysis of bone loss in space travelers. NPJ Microgravity 2020, 6, 1–9. [Google Scholar] [CrossRef]
- Blaber, E.A.; Dvorochkin, N.; Lee, C.; Alwood, J.S.; Yousuf, R.; Pianetta, P.; Globus, R.K.; Burns, B.P.; Almeida, E.A.C. Microgravity Induces Pelvic Bone Loss through Osteoclastic Activity, Osteocytic Osteolysis, and Osteoblastic Cell Cycle Inhibition by CDKN1a/p21. PLoS ONE 2013, 8, e61372. [Google Scholar] [CrossRef] [Green Version]
- Camirand, A.; Goltzman, D.; Gupta, A.; Kaouass, M.; Panda, D.; Karaplis, A. The Role of Parathyroid Hormone-Related Protein (PTHrP) in Osteoblast Response to Microgravity: Mechanistic Implications for Osteoporosis Development. PLoS ONE 2016, 11, e0160034. [Google Scholar] [CrossRef] [PubMed]
- Klein-Nulend, J.; Bacabac, R.; Veldhuijzen, J.; Van Loon, J. Microgravity and bone cell mechanosensitivity. Adv. Space Res. 2003, 32, 1551–1559. [Google Scholar] [CrossRef]
- Maier, J.A.M.; Cialdai, F.; Monici, M.; Morbidelli, L. The Impact of Microgravity and Hypergravity on Endothelial Cells. BioMed Res. Int. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ludtka, C.; Silberman, J.; Moore, E.; Allen, J.B. Macrophages in microgravity: The impact of space on immune cells. NPJ Microgravity 2021, 7, 1–10. [Google Scholar] [CrossRef]
- Wuest, S.L.; Richard, S.; Kopp, S.; Grimm, D.; Egli, M. Simulated Microgravity: Critical Review on the Use of Random Positioning Machines for Mammalian Cell Culture. BioMed Res. Int. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Luo, Q.; Yuan, L.; Song, G. Microgravity directs stem cell differentiation. Histol. Histopathol. 2016, 16, 99–106. [Google Scholar]
- Grimm, D.; Wise, P.; Lebert, M.; Richter, P.; Baatout, S. How and why does the proteome respond to microgravity? Expert Rev. Proteom. 2011, 8, 13–27. [Google Scholar] [CrossRef]
- Cristofaro, F.; Pani, G.; Pascucci, B.; Mariani, A.; Balsamo, M.; Donati, A.; Mascetti, G.; Rea, G.; Rizzo, A.M.; Visai, L. The NATO project: Nanoparticle-based countermeasures for microgravity-induced osteoporosis. Sci. Rep. 2019, 9, 17141-15. [Google Scholar] [CrossRef]
- Carmeliet, G.; Vico, L.; Bouillon, R. Space flight: A challenge for normal bone homeostasis. Crit. Rev. Eukaryot. Gene Expr. 2001, 11, 131–144. [Google Scholar] [CrossRef]
- Gershovich, P.M.; Gershovich, I.G.; Buravkova, L.B. The effects of simulated microgravity on the pattern of gene expression in human bone marrow mesenchymal stem cells under osteogenic differentiation. Fiziologiia Cheloveka 2014, 39, 105–111. [Google Scholar]
- Buravkova, L.B.; Gershovich, Y.G.; Grigorev, A.I. Sensitivity of stromal precursor cells of different commitment to simulated microgravity. Dokl. Biol. Sci. 2010, 432, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Katsaras, G.N.; Lambrou, G.I. Gene Expression in Osteoblasts and Osteoclasts Under Microgravity Conditions: A Systematic Review. Curr. Genom. 2019, 20, 184–198. [Google Scholar] [CrossRef]
- Buravkova, L.; Gershovich, P.; Gershovich, J.; Grigor’Ev, A. Mechanisms of Gravitational Sensitivity of Osteogenic Precursor Cells. Acta Nat. 2010, 2, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhang, C.; Chen, J.-L.; Hong, F.-F.; Chen, P.; Wang, J.-F. Effects of simulated microgravity on the expression profiles of RNA during osteogenic differentiation of human bone marrow mesenchymal stem cells. Cell Prolif. 2018, 52, e12539. [Google Scholar] [CrossRef] [PubMed]
- Muncie, J.M.; Weaver, V.M. The Physical and Biochemical Properties of the Extracellular Matrix Regulate Cell Fate. Curr. Top. Dev. Biol. 2018, 130, 1–37. [Google Scholar] [CrossRef]
- Ozdil, B.; Güler, G.; Acikgoz, E.; Kocaturk, D.C.; Aktug, H. The effect of extracellular matrix on the differentiation of mouse embryonic stem cells. J. Cell. Biochem. 2020, 121, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Jha, A.K.; Jackson, W.M.; Healy, K.E. Controlling Osteogenic Stem Cell Differentiation via Soft Bioinspired Hydrogels. PLoS ONE 2014, 9, e98640. [Google Scholar] [CrossRef]
- Fernández-Pernas, P.; Barrachina, L.; Marquina, M.; Rodellar, C.; Arufe, M.; Costa, C. Mesenchymal stromal cells for articular cartilage repair: Preclinical studies. Eur. Cells Mater. 2020, 40, 88–114. [Google Scholar] [CrossRef] [PubMed]
- Barba, M.; Cicione, C.; Bernardini, C.; Michetti, F.; Lattanzi, W. Adipose-Derived Mesenchymal Cells for Bone Regereneration: State of the Art. BioMed Res. Int. 2013, 2013, 416391. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.B.; Moncivais, K.; Caplan, A.I. Mesenchymal stem cells: Environmentally responsive therapeutics for regenerative medicine. Exp. Mol. Med. 2013, 45, e54. [Google Scholar] [CrossRef] [Green Version]
- Ulbrich, C.; Wehland, M.; Pietsch, J.; Aleshcheva, G.; Wise, P.; Van Loon, J.; Magnusson, N.E.; Infanger, M.; Grosse, J.; Eilles, C.; et al. The Impact of Simulated and Real Microgravity on Bone Cells and Mesenchymal Stem Cells. BioMed Res. Int. 2014, 2014, 928507. [Google Scholar] [CrossRef] [PubMed]
- Ratushnyy, A.; Yakubets, D.; Andreeva, E.; Buravkova, L. Simulated microgravity modulates the mesenchymal stromal cell response to inflammatory stimulation. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Buravkova, L.B.; Gershovich, P.M.; Gershovich, J.G.; Grigoriev, A.I. Microgravity and Mesenchymal Stem Cell Response. Curr. Biotechnol. 2013, 999, 25–30. [Google Scholar] [CrossRef]
- Ratushnyy, A.Y.; Buravkova, L.B. Expression of focal adhesion genes in mesenchymal stem cells under simulated microgravity. Dokl. Biochem. Biophys. 2017, 477, 354–356. [Google Scholar] [CrossRef]
- Aleshcheva, G.; Sahana, J.; Ma, X.; Hauslage, J.; Hemmersbach, R.; Egli, M.; Infanger, M.; Bauer, J.; Grimm, D. Changes in Morphology, Gene Expression and Protein Content in Chondrocytes Cultured on a Random Positioning Machine. PLoS ONE 2013, 8, e79057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebnerasuly, F.; Hajebrahimi, Z.; Tabaie, S.M.; Darbouy, M. Simulated Microgravity Condition Alters the Gene Expression of some ECM and Adhesion Molecules in Adipose Derived Stem Cells. Int. J. Mol. Cell. Med. 2018, 7, 146–157. [Google Scholar]
- Buken, C.; Sahana, J.; Corydon, T.J.; Melnik, D.; Bauer, J.; Wehland, M.; Krüger, M.; Balk, S.; Abuagela, N.; Infanger, M.; et al. Morphological and Molecular Changes in Juvenile Normal Human Fibroblasts Exposed to Simulated Microgravity. Sci. Rep. 2019, 9, 11882. [Google Scholar] [CrossRef]
- Makihira, S.; Kawahara, Y.; Yuge, L.; Mine, Y.; Nikawa, H. Impact of the microgravity environment in a 3-dimensional clinostat on osteoblast- and osteoclast-like cells. Cell Biol. Int. 2008, 32, 1176–1181. [Google Scholar] [CrossRef] [Green Version]
- Pardo, S.J.; Patel, M.J.; Sykes, M.C.; Platt, M.O.; Boyd, N.L.; Sorescu, G.P.; Xu, M.; Van Loon, J.J.W.A.; Wang, M.D.; Jo, H. Simulated microgravity using the Random Positioning Machine inhibits differentiation and alters gene expression profiles of 2T3 preosteoblasts. Am. J. Physiol. Physiol. 2005, 288, C1211–C1221. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Byers, B.A.; Pavlath, G.K.; Murphy, T.J.; Karsenty, G.; Garcia, A.J. Cell-Type-Dependent Up-Regulation of In Vitro Mineralization After Overexpression of the Osteoblast-Specific Transcription Factor Runx2/Cbfa1. J. Bone Miner. Res. 2002, 17, 1931–1944. [Google Scholar] [CrossRef] [PubMed]
- Ratushny, A.; Yakubets, D.; Zhivodernikov, I.; Buravkova, L. Adhesion molecules of multipotent mesenchymal stromal cells obtained from adipose tissue during simulation of the effects of microgravity. Aerosp. Environ. Med. 2017, 51, 38–43. [Google Scholar] [CrossRef]
- Kundu, A.K.; Khatiwala, C.B.; Putnam, A.J. Extracellular Matrix Remodeling, Integrin Expression, and Downstream Signaling Pathways Influence the Osteogenic Differentiation of Mesenchymal Stem Cells on Poly(Lactide-Co-Glycolide) Substrates. Tissue Eng. Part A 2009, 15, 273–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.-M.; Yang, L.; Wang, Z.; Liu, Y.-W.; Fan, J.-Z.; Fan, J.; Liu, J.; Luo, Z.-J. Overexpression of integrin a2 promotes osteogenic differentiation of hBMSCs from senile osteoporosis through the ERK pathway. Int. J. Clin. Exp. Pathol. 2013, 6, 841–852. [Google Scholar] [PubMed]
- Lee, H.M.; Seo, S.; Kim, J.; Kim, M.K.; Seo, H.; Kim, K.S.; Jang, Y.; Ryu, C.J. Expression dynamics of integrin α2, α3, and αV upon osteogenic differentiation of human mesenchymal stem cells. Stem Cell Res. Ther. 2020, 11, 210. [Google Scholar] [CrossRef] [PubMed]
- Picke, A.-K.; Campbell, G.M.; Blüher, M.; Krügel, U.; Schmidt, F.N.; Tsourdi, E.; Winzer, M.; Rauner, M.; Vukicevic, V.; Busse, B.; et al. Thy-1 (CD90) promotes bone formation and protects against obesity. Sci. Transl. Med. 2018, 10, eaao6806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradaschia-Correa, V.; Josephson, A.M.; Egol, A.J.; Mizrahi, M.M.; Leclerc, K.; Huo, J.; Cronstein, B.N.; Leucht, P. Ecto-5′-nucleotidase (CD73) regulates bone formation and remodeling during intramembranous bone repair in aging mice. Tissue Cell 2017, 49, 545–551. [Google Scholar] [CrossRef]
- Uddin, S.M.Z.; Qin, Y.-X. Enhancement of Osteogenic Differentiation and Proliferation in Human Mesenchymal Stem Cells by a Modified Low Intensity Ultrasound Stimulation under Simulated Microgravity. PLoS ONE 2013, 8, e73914. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, R.T.; Xiao, G. Regulation of the osteoblast-specific transcription factor, Runx2: Responsiveness to multiple signal transduction pathways. J. Cell. Biochem. 2003, 88, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, L.; Jiang, Y.; Wang, C.; Geng, B.; Wang, Y.; Chen, J.; Liu, F.; Qiu, P.; Zhai, G.; et al. Space microgravity drives transdifferentiation of human bone marrow-derived mesenchymal stem cells from osteogenesis to adipogenesis. FASEB J. 2018, 32, 4444–4458. [Google Scholar] [CrossRef] [Green Version]
- Meyers, V.E.; Zayzafoon, M.; Gonda, S.R.; Gathings, W.E.; McDonald, J.M. Modeled microgravity disrupts collagen I/integrin signaling during osteoblastic differentiation of human mesenchymal stem cells. J. Cell. Biochem. 2004, 93, 697–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colucci, S.; Colaianni, G.; Brunetti, G.; Ferranti, F.; Mascetti, G.; Mori, G.; Grano, M. Irisin prevents microgravity-induced impairment of osteoblast differentiation in vitro during the space flight CRS-14 mission. FASEB J. 2020, 34, 10096–10106. [Google Scholar] [CrossRef]
- Caillot-Augusseau, A.; Vico, L.; Heer, M.; Voroviev, D.; Souberbielle, J.C.; Zitterman, A.; Alexandre, C.; Lafage-Proust, M.H. Space flight is associated with rapid decreases of undercarboxylated osteocalcin and increases of markers of bone resorption without changes in their circadian variation: Observations in two cosmonauts. Clin. Chem. 2000, 46, 1136–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vico, L.; Hinsenkamp, M.; Jones, D.; Marie, P.J.; Zallone, A.; Cancedda, R. Osteobiology, strain, and microgravity. Part II: Studies at the tissue level. Calcif. Tissue Int. 2001, 68, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zandi, N.; Mostafavi, E.; Shokrgozar, M.A.; Tamjid, E.; Webster, T.J.; Annabi, N.; Simchi, A. Biomimetic proteoglycan nanoparticles for growth factor immobilization and delivery. Biomater. Sci. 2019, 8, 1127–1136. [Google Scholar] [CrossRef]
- Chun, S.Y.; Lim, J.O.; Lee, E.H.; Han, M.-H.; Ha, Y.-S.; Lee, J.N.; Kim, B.S.; Park, M.J.; Yeo, M.; Jung, B.; et al. Preparation and Characterization of Human Adipose Tissue-Derived Extracellular Matrix, Growth Factors, and Stem Cells: A Concise Review. Tissue Eng. Regen. Med. 2019, 16, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.-H.; Springer, T.A. Integrin structures and conformational signaling. Curr. Opin. Cell Biol. 2006, 18, 579–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremble, P.; Chiquet-Ehrismann, R.; Werb, Z. The extracellular matrix ligands fibronectin and tenascin collaborate in regulating collagenase gene expression in fibroblasts. Mol. Biol. Cell 1994, 5, 439–453. [Google Scholar] [CrossRef] [Green Version]
- Probstmeier, R.; Pesheva, P. Tenascin-C inhibits 1 integrin-dependent cell adhesion and neurite outgrowth on fibronectin by a disialoganglioside-mediated signaling mechanism. Glycobiology 1999, 9, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.K.; Du, W.; Shelton, S.J.; Oldham, M.C.; DiPersio, C.M.; Klein, O.D. A FAK-YAP-mTOR signaling axis regulates stem cell-based tissue renewal in mice. Cell Stem Cell 2017, 21, 91–106. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Li, Y.; Liu, S.; Chen, Q.; Liu, Y. ITGA3 serves as a diagnostic and prognostic biomarker for pancreatic cancer. OncoTargets Ther. 2019, 12, 4141–4152. [Google Scholar] [CrossRef] [Green Version]
- Docherty, A.J.; Murphy, G. The tissue metalloproteinase family and the inhibitor TIMP: A study using cDNAs and recombinant proteins. Ann. Rheum. Dis. 1990, 49, 469–479. [Google Scholar] [PubMed]
- Sun, S.; Bay-Jensen, A.-C.; Karsdal, M.A.; Siebuhr, A.S.; Zheng, Q.; Maksymowych, W.P.; Christiansen, T.G.; Henriksen, K. The active form of MMP-3 is a marker of synovial inflammation and cartilage turnover in inflammatory joint diseases. BMC Musculoskelet. Disord. 2014, 15, 93. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.; Homaei, A.; El-Seedi, H.R.; Akhtar, N. Cathepsins: Proteases that are vital for survival but can also be fatal. Biomed. Pharmacother. 2018, 105, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Kassiri, Z. Biology of Tissue Inhibitor of Metalloproteinase 3 (TIMP3), and Its Therapeutic Implications in Cardiovascular Pathology. Front. Physiol. 2020, 11, 661. [Google Scholar] [CrossRef]
- He, L.; Pan, S.; Li, Y.; Zhang, L.; Zhang, W.; Yi, H.; Song, C.; Niu, Y. Increased proliferation and adhesion properties of human dental pulp stem cells in PLGA scaffolds via simulated microgravity. Int. Endod. J. 2015, 49, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Kraus, A.; Luetzenberg, R.; Abuagela, N.; Hollenberg, S.; Infanger, M. Spheroid formation and modulation of tenocyte-specific gene expression under simulated microgravity. Muscle Ligaments Tendons J. 2019, 7, 411. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human Adipose Tissue Is a Source of Multipotent Stem Cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef]
- Van Loon, J.J. Some history and use of the random positioning machine, RPM, in gravity related research. Adv. Space Res. 2007, 39, 1161–1165. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)). Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Description | Fold Change (Mean ± SD) | |
|---|---|---|---|
| RPM (Int) vs. Stat (Int) | RPM (Ost) vs. Stat (Ost) | ||
| COL11A1 | Collagen, type XI, alpha 1 | 1.48 ± 0.39 * | 1.92 ± 0.80 * |
| CTNNB1 | Catenin, beta 1 | 1.05 ± 0.59 | 1.82 ± 1.10 * |
| CTNND1 | Catenin, delta 1 | 1.56 ± 0.60 * | 0.70 ± 0.26 |
| HAS1 | Hyaluronan synthase 1 | 0.90 ± 0.13 | 0.55 ± 0.37 * |
| ITGA3 | Integrin, alpha 3 (antigen CD49C) | 0.72 ± 0.35 | 0.53 ± 0.23 * |
| ITGB1 | Integrin, beta 1 | 1.13 ± 0.26 | 1.56 ± 0.45 * |
| LAMA3 | Laminin, alpha 3 | 0.94 ± 0.69 | 0.63 ± 0.25 * |
| MMP1 | Matrix metallopeptidase 1 | 1.77 ± 1.53 | 0.63 ± 0.10 * |
| MMP11 | Matrix metallopeptidase 11 | 0.82 ± 0.13 | 0.51 ± 0.05 * |
| TIMP3 | TIMP metallopeptidase inhibitor 3 | 0.66 ± 0.13 * | 0.75 ± 0.24 |
| TNC | Tenascin C | 2.23 ± 0.99 * | 2.16 ± 1.55 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhivodernikov, I.; Ratushnyy, A.; Buravkova, L. Simulated Microgravity Remodels Extracellular Matrix of Osteocommitted Mesenchymal Stromal Cells. Int. J. Mol. Sci. 2021, 22, 5428. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115428
Zhivodernikov I, Ratushnyy A, Buravkova L. Simulated Microgravity Remodels Extracellular Matrix of Osteocommitted Mesenchymal Stromal Cells. International Journal of Molecular Sciences. 2021; 22(11):5428. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115428
Chicago/Turabian StyleZhivodernikov, Ivan, Andrey Ratushnyy, and Ludmila Buravkova. 2021. "Simulated Microgravity Remodels Extracellular Matrix of Osteocommitted Mesenchymal Stromal Cells" International Journal of Molecular Sciences 22, no. 11: 5428. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115428