
Contribution of Massive Mitochondrial Fusion and Subsequent Fission in the Plant Life Cycle to the Integrity of the Mitochondrion and Its Genome

Abstract
:1. Introduction
2. Mitochondrial Division
2.1. Proteins of the Mitochondrial Fission Machinery
2.2. The Endoplasmic Reticulum and Mitochondrial Fission
3. Mitochondrial Fusion
3.1. Demonstration of Mitochondrial Fusion
3.2. The Mechanism of Mitochondrial Fusion
4. Significance of the Mitochondrial Fusion/Fission Cycle
4.1. Mitochondrial DNA Content per Mitochondrion Is Highly Variable
4.2. MtDNA Recombination
4.3. Cytoplasmic Male Sterility
4.4. Mitochondrial Fusion and Energetics
4.5. Mitochondrial Fusion and Evolution
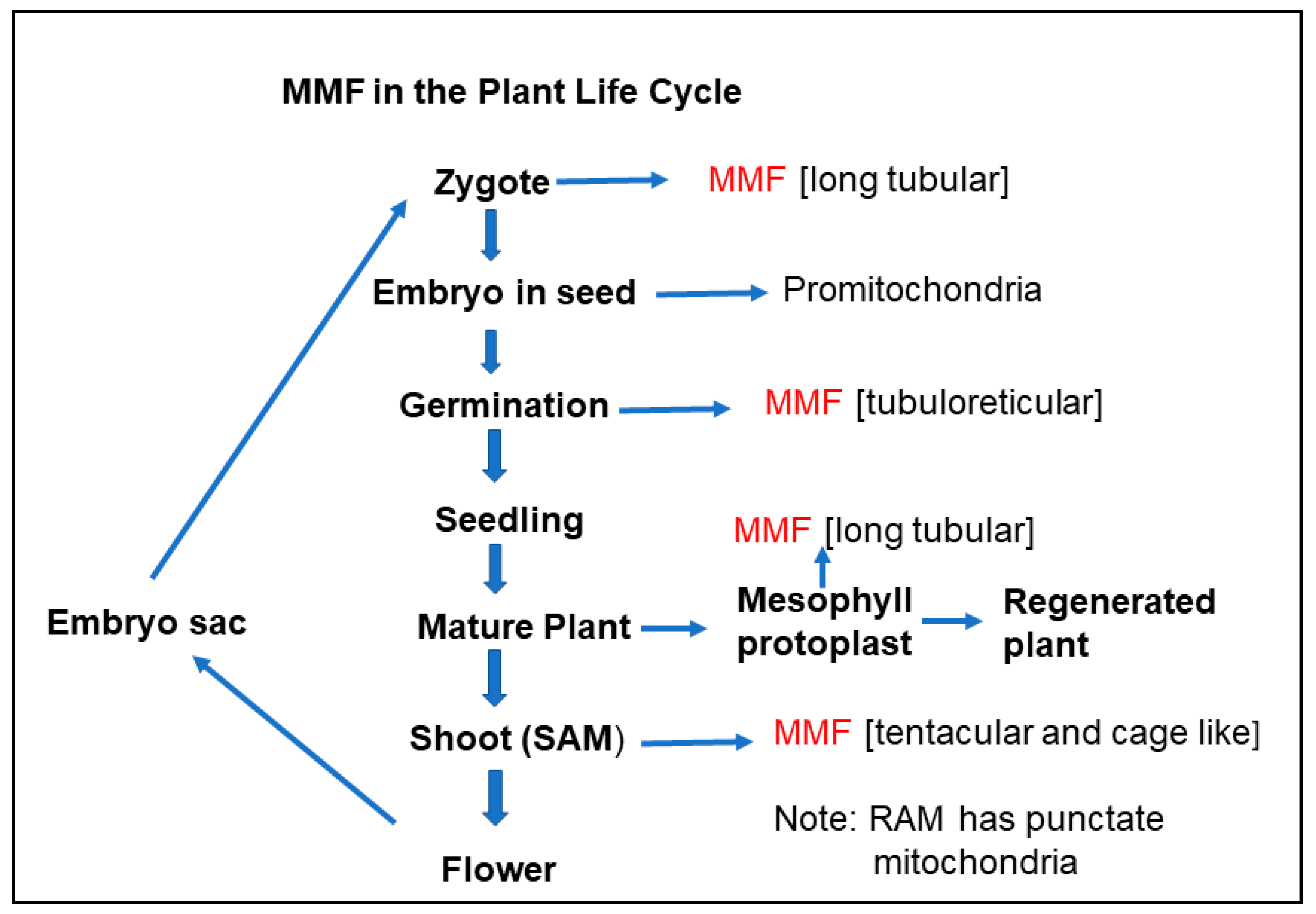
5. Massive Mitochondrial Fusion in the Plant Life Cycle
5.1. The Metabolising Non-Dividing Cell
5.2. Mitochondrial Fusion and the Cell Cycle
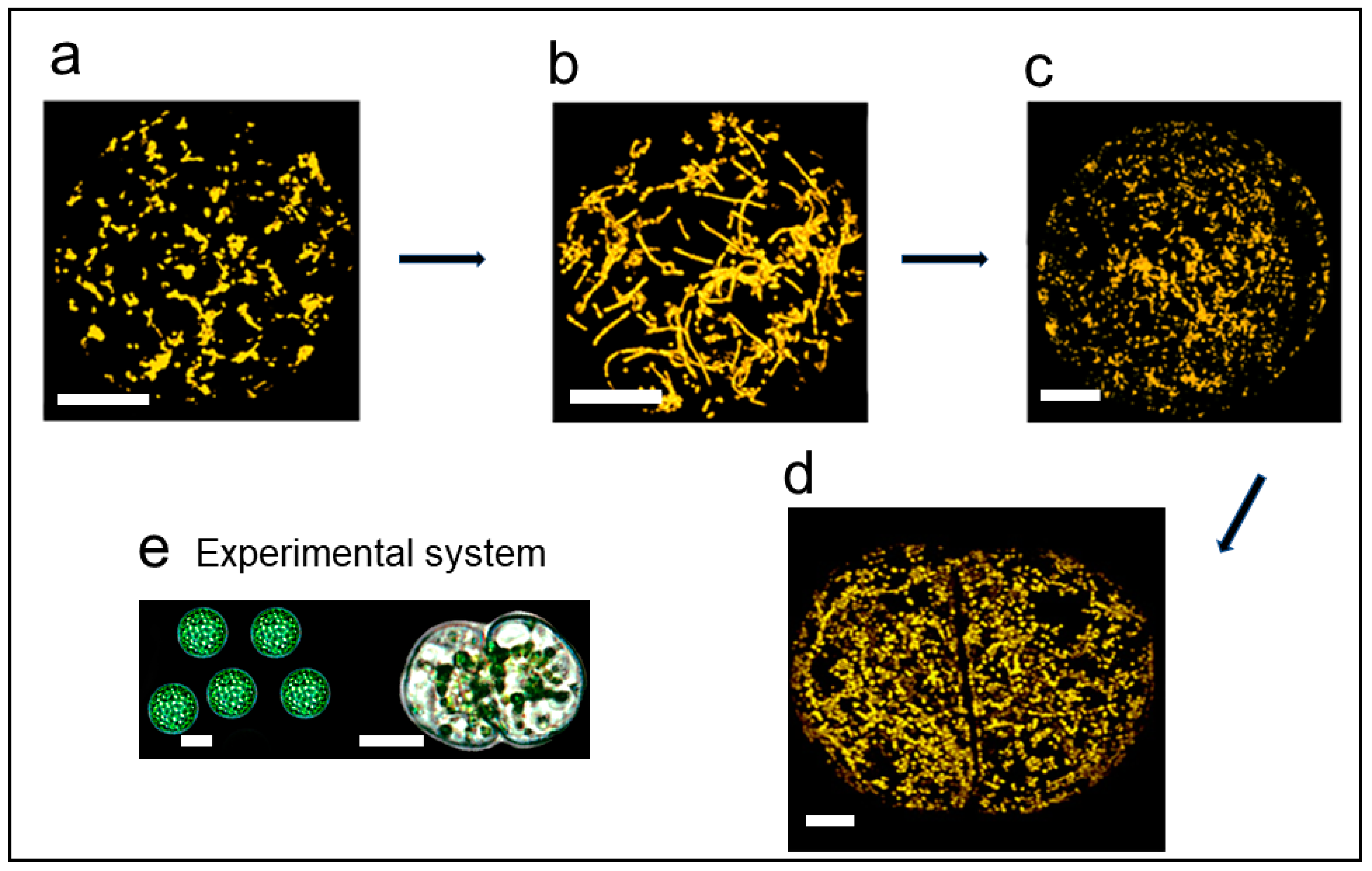
5.3. Massive Mitochondrial Fusion in Regenerating Protoplasts
5.4. Massive Mitochondrial Fusion in the Shoot Apical Meristem
5.5. Massive Mitochondrial Fusion in the Zygote
5.6. Massive Mitochondrial Fusion in Germination
5.7. Significance of Massive Mitochondrial Fusion
6. Conclusions
Funding
Conflicts of Interest
References
- Rose, R.J. Sustaining life: Maintaining chloroplasts and mitochondria and their genomes in plants. Yale J. Biol. Med. 2019, 92, 499–510. [Google Scholar]
- Mackenzie, S.; McIntosh, L. Higher plant mitochondria. Plant Cell 1999, 11, 571–585. [Google Scholar] [CrossRef]
- Smith, A.G.; Croft, M.T.; Moulin, M.; Webb, M.E. Plants need their vitamins too. Curr. Opin. Plant Biol. 2007, 10, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Sweetlove, L.J.; Fait, A.; Nunes-Nesi, A.; Williams, T.; Fernie, A.R. The mitochondrion: An integration point of cellular metabolism and signalling. Crit. Rev. Plant Sci. 2007, 26, 17–43. [Google Scholar] [CrossRef]
- Jaipargas, E.A.; Barton, K.A.; Mathur, N.; Mathur, J. Mitochondrial pleomorphy in plant cells is driven by contiguous ER dynamics. Front. Plant Sci. 2015, 6, 783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, P.; Rugen, N.; Carrie, C.; Elsässer, M.; Finkemeier, I.; Giese, J.; Hildebrandt, T.M.; Kühn, K.; Maurino, V.G.; Ruberti, C.; et al. Single organelle function and organization as estimated from Arabidopsis mitochondrial proteomics. Plant J. 2020, 101, 420–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheahan, M.B.; Rose, R.J.; McCurdy, D.W. Organelle inheritance in plant cell division: The actin cytoskeleton is required for unbiased inheritance of chloroplasts, mitochondria and endoplasmic reticulum in dividing protoplasts. Plant J. 2004, 37, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Preuten, T.; Cincu, E.; Fuchs, J.; Zoschke, R.; Liere, K.; Börner, T. Fewer genes than organelles: Extremely low and variable gene copy numbers in mitochondria of somatic plant cells. Plant J. 2010, 64, 948–959. [Google Scholar] [CrossRef]
- Arimura, S.I.; Yamamoto, J.; Aida, G.P.; Nakazono, M.; Tsutsumi, N. Frequent fusion and fission of plant mitochondria with unequal nucleoid distribution. Proc. Natl. Acad. Sci. USA 2004, 101, 7805–7808. [Google Scholar] [CrossRef] [Green Version]
- Seguí-Simarro, J.M.; Coronado, M.J.; Staehelin, L.A. The mitochondrial cycle of Arabidopsis shoot apical meristem and leaf primordium meristematic cells is defined by a perinuclear tentaculate/cage-like mitochondrion. Plant Physiol. 2008, 148, 1380–1393. [Google Scholar] [CrossRef] [Green Version]
- Unseld, M.; Marienfeld, J.R.; Brandt, P.; Brennicke, A. The mitochondrial genome of Arabidopsis thaliana contains 57 genes in 366,924 nucleotides. Nat. Genet. 1997, 15, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Gualberto, J.M.; Mileshina, D.; Wallet, C.; Niazi, A.K.; Weber-Lotfi, F.; Dietrich, A. The plant mitochondrial genome: Dynamics and maintenance. Biochimie 2014, 100, 107–120. [Google Scholar] [CrossRef]
- Gualberto, J.M.; Newton, K.J. Plant mitochondrial genomes: Dynamics and mechanisms of mutation. Annu. Rev. Plant Biol. 2017, 68, 225–252. [Google Scholar] [CrossRef]
- Arimura, S.I.; Tsutsumi, N. A dynamin-like protein (ADL2b), rather than FtsZ, is involved in Arabidopsis mitochondrial division. Proc. Natl. Acad. Sci. USA 2002, 99, 5727–5731. [Google Scholar] [CrossRef] [Green Version]
- Logan, D.C. Plant mitochondrial dynamics. Biochim. Biophys. Acta Mol. Cell Res. 2006, 1763, 430–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyall, S.D.; Brown, M.T.; Johnson, P.J. Ancient invasions: From endosymbionts to organelles. Science 2004, 304, 253–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takanashi, H.; Arimura, S.I.; Sakamoto, W.; Tsutsumi, N. Different amounts of DNA in each mitochondrion in rice root. Genes Genet. Syst. 2006, 81, 215–218. [Google Scholar] [CrossRef] [Green Version]
- Sheahan, M.B.; McCurdy, D.W.; Rose, R.J. Mitochondria as a connected population: Ensuring continuity of the mitochondrial genome during plant cell dedifferentiation through massive mitochondrial fusion. Plant J. 2005, 44, 744–755. [Google Scholar] [CrossRef] [PubMed]
- Logan, D.C. The mitochondrial compartment. J. Exp. Bot. 2006, 57, 1225–1243. [Google Scholar] [CrossRef] [PubMed]
- Arimura, S.I. Fission and fusion of plant mitochondria, and genome maintenance. Plant Physiol. 2018, 176, 152–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, R.J.; McCurdy, D.W. New beginnings: Mitochondrial renewal by massive mitochondrial fusion. Trends Plant Sci. 2017, 22, 641–643. [Google Scholar] [CrossRef] [PubMed]
- Arimura, S.I.; Aida, G.P.; Fujimoto, M.; Nakazono, M.; Tsutsumi, N. Arabidopsis dynamin-like protein 2a (ADL2a), like ADL2b, is involved in plant mitochondrial division. Plant Cell Physiol. 2004, 45, 236–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, D.C.; Scott, I.; Tobin, A.K. ADL2a, like ADL2b, is involved in the control of higher plant mitochondrial morphology. J. Exp. Bot. 2004, 55, 783–785. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.; Bednarek, S.Y.; Blumwald, E.; Hwang, I.; Jurgens, G.; Menzel, D.; Osteryoung, K.W.; Raikhel, N.V.; Shinozaki, K.; Tsutsumi, N.; et al. A unified nomenclature for Arabidopsis dynamin related large GTPases based on homology and possible functions. Plant Mol. Biol. 2003, 53, 261–265. [Google Scholar] [CrossRef]
- Logan, D.C. Mitochondrial fusion, division and positioning in plants. Biochem. Soc. Trans. 2010, 38, 789–795. [Google Scholar] [CrossRef]
- Aung, K.; Hu, J. Differential roles of Arabidopsis dynamin-related proteins DRP3A, DRP3B, and DRP5B in organelle division. J. Integr. Plant Biol. 2012, 54, 921–931. [Google Scholar] [PubMed]
- Jin, J.B.; Bae, H.; Kim, S.J.; Jin, Y.H.; Goh, C.H.; Kim, D.H.; Lee, Y.J.; Tse, Y.C.; Jiang, L.; Hwang, I. The Arabidopsis dynamin-like proteins ADL1C and ADL1E play a critical role in mitochondrial morphogenesis. Plant Cell 2003, 15, 2357–2369. [Google Scholar] [CrossRef] [Green Version]
- Bednarek, S.Y.; Backues, S.K. Plant dynamin-related protein families DRP1 and DRP2 in plant development. Biochem. Soc. Trans. 2010, 38, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; MacCready, J.S.; Ducat, D.C.; Osteryoung, K.W. The molecular machinery of chloroplast division. Plant Physiol. 2018, 176, 138–151. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Hu, J. The Arabidopsis chloroplast division protein DYNAMIN-RELATED PROTEIN5B also mediates peroxisome division. Plant Cell 2010, 22, 431–442. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, M.; Arimura, S.I.; Mano, S.; Kondo, M.; Saito, C.; Ueda, T.; Nakazono, M.; Nakano, A.; Nishimura, M.; Tsutsumi, N. Arabidopsis dynamin-related proteins DRP3A and DRP3B are functionally redundant in mitochondrial fission, but have distinct roles in peroxisomal fission. Plant J. 2009, 58, 388–400. [Google Scholar] [CrossRef]
- Pan, R.; Hu, J. Plant mitochondrial dynamics and the role of membrane lipids. Plant Signal. Behav. 2015, 10, e1050573. [Google Scholar] [CrossRef]
- Scott, I.; Tobin, A.K.; Logan, D.C. BIGYIN, an orthologue of human and yeast FIS1 genes functions in the control of mitochondrial size and number in Arabidopsis thaliana. J. Exp. Bot. 2006, 57, 1275–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.C.; Hu, J.P. FISSION1A and FISSION1B proteins mediate the fission of peroxisomes and mitochondria in Arabidopsis. Mol. Plant 2008, 1, 1036–1047. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, J. Two small protein families, DYNAMIN-RELATED PROTEIN3 and FISSION1, are required for peroxisome fission in Arabidopsis. Plant J. 2009, 57, 146–159. [Google Scholar] [CrossRef]
- Arimura, S.I.; Fujimoto, M.; Doniwa, Y.; Kadoya, N.; Nakazono, M.; Sakamoto, W.; Tsutsumi, N. Arabidopsis ELONGATED MITOCHONDRIA1 is required for localization of DYNAMIN-RELATED PROTEIN3A to mitochondrial fission sites. Plant Cell 2008, 20, 1555–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, R.; Jones, A.D.; Hu, J. Cardiolipin-mediated mitochondrial dynamics and stress response in Arabidopsis. Plant Cell 2014, 26, 391–409. [Google Scholar] [CrossRef] [Green Version]
- Aung, K.; Hu, J. The Arabidopsis tail-anchored protein PEROXISOMAL AND MITOCHONDRIAL DIVISION FACTOR1 is involved in the morphogenesis and proliferation of peroxisomes and mitochondria. Plant Cell 2011, 23, 4446–4461. [Google Scholar] [CrossRef] [Green Version]
- Friedman, J.R.; Lackner, L.L.; West, M.; DiBenedetto, J.R.; Nunnari, J.; Voeltz, G.K. ER tubules mark sites of mitochondrial division. Science 2011, 334, 358–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambold, A.S.; Lippincott-Schwartz, J. Severing mitochondria. Science 2011, 334, 186–187. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Yoon, Y. Mitochondrial fission and fusion. Biochem. Soc. Trans. 2016, 44, 1725–1735. [Google Scholar] [CrossRef]
- Mueller, S.J.; Reski, R. Mitochondrial dynamics and the ER: The plant perspective. Front. Cell Develop. Biol. 2015, 3, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawes, C.; Kiviniemi, P.; Kriechbaumer, V. The endoplasmic reticulum: A dynamic and well-connected organelle. J. Integr. Plant Biol. 2015, 57, 50–62. [Google Scholar] [CrossRef] [Green Version]
- Korobova, F.; Ramabhadran, V.; Higgs, H.N. An actin-dependent step in mitochondrial fission mediated by the ER-associated formin INF2. Science 2013, 339, 464–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Liu, P.; Zhang, Q.; Zhu, J.; Chen, T.; Arimura, S.I.; Tsutsumi, N.; Lin, J. Phosphorylation and ubiquitination of dynamin-related proteins (AtDRP3A/3B) synergically regulate mitochondrial proliferation during mitosis. Plant J. 2012, 72, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Kraus, F.; Roy, K.; Pucadyil, T.J.; Ryan, M.T. Function and regulation of the divisome for mitochondrial fission. Nature 2021, 590, 57–66. [Google Scholar] [CrossRef]
- Belliard, G.; Vedel, F.; Pelletier, G. Mitochondrial recombination in cytoplasmic hybrids of Nicotiana tabacum by protoplast fusion. Nature 1979, 281, 401–403. [Google Scholar] [CrossRef]
- Rose, R.J.; Thomas, M.R.; Fitter, J.T. The transfer of cytoplasmic and nuclear genomes by somatic hybridisation. Funct. Plant Biol. 1990, 17, 303–321. [Google Scholar] [CrossRef]
- White, R.R.; Lin, C.; Leaves, I.; Castro, I.G.; Metz, J.; Bateman, B.C.; Botchway, S.W.; Ward, A.D.; Ashwin, P.; Sparkes, I. Miro2 tethers the ER to mitochondria to promote mitochondrial fusion in tobacco leaf epidermal cells. Commun. Biol. 2020, 3, 161. [Google Scholar] [CrossRef] [Green Version]
- Wakamatsu, K.; Fujimoto, M.; Nakazono, M.; Arimura, S.I.; Tsutsumi, N. Fusion of mitochondria in tobacco suspension cultured cells is dependent on the cellular ATP level but not on actin polymerization. Plant Cell Rep. 2010, 29, 1139–1145. [Google Scholar] [CrossRef]
- El Zawily, A.M.; Schwarzländer, M.; Finkemeier, I.; Johnston, I.G.; Benamar, A.; Cao, Y.; Gissot, C.; Meyer, A.J.; Wilson, K.; Datla, R.; et al. FRIENDLY regulates mitochondrial distribution, fusion, and quality control in Arabidopsis. Plant Physiol. 2014, 166, 808–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonsdale, D.M.; Brears, T.; Hodge, T.P.; Melville, S.E.; Rottmann, W.H. The plant mitochondrial genome: Homologous recombination as a mechanism for generating heterogeneity. Phil. Trans. Royal Soc. London B Biol. Sci. 1988, 319, 149–163. [Google Scholar]
- Bendich, A.J.; Gauriloff, L.P. Morphometric analysis of cucurbit mitochondria: The relationship between chondriome volume and DNA content. Protoplasma 1984, 119, 1–7. [Google Scholar] [CrossRef]
- Kuroiwa, T.; Fujie, M.; Kuroiwa, H. Studies on the behavior of mitochondrial DNA: Synthesis of mitochondrial DNA occurs actively in a specific region just above the quiescent center in the root meristem of Pelargonium zonale. J. Cell Sci. 1992, 101, 483–493. [Google Scholar] [CrossRef]
- Oldenburg, D.J.; Bendich, A.J. DNA maintenance in plastids and mitochondria of plants. Front. Plant Sci. 2015, 6, 883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, M.; Nemoto, Y.; Kawano, S.; Nagata, T.; Hirokawa, H.; Kuroiwa, T. Organization of heterogeneous mitochondrial DNA molecules in mitochondrial nuclei of cultured tobacco cells. Protoplasma 1993, 175, 112–120. [Google Scholar] [CrossRef]
- Johnston, I.G. Tension and resolution: Dynamic, evolving populations of organelle genomes within plant cells. Mol. Plant 2019, 12, 7647–7683. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, Y.; Havey, M.J.; Shou, W. Copy numbers of mitochondrial genes change during melon leaf development and are lower than the numbers of mitochondria. Hortic. Res. 2019, 6, 95. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, K.H.; Li, W.H.; Sharp, P.M. Rates of nucleotide substitution vary greatly among plant mitochondrial, chloroplast, and nuclear DNAs. Proc. Natl. Acad. Sci. USA 1987, 84, 9054–9058. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.D.; Herbon, L.A. Plant mitochondrial DNA evolved rapidly in structure, but slowly in sequence. J. Mol. Evol. 1988, 28, 87–97. [Google Scholar] [CrossRef]
- Hu, J.; Huang, W.; Huang, Q.; Qin, X.; Yu, C.; Wang, L.; Li, S.; Zhu, R.; Zhu, Y. Mitochondria and cytoplasmic male sterility in plants. Mitochondrion 2014, 19, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Horn, R.; Gupta, K.J.; Colombo, N. Mitochondrion role in molecular basis of cytoplasmic male sterility. Mitochondrion 2014, 19, 198–205. [Google Scholar] [CrossRef]
- Fitter, J.T.; Thomas, M.R.; Niu, C.; Rose, R.J. Investigation of Nicotiana tabacum (+) N. suaveolens cybrids with carpelloid stamens. J. Plant Physiol. 2005, 162, 225–235. [Google Scholar] [PubMed]
- Miyamura, S.; Kuroiwa, T.; Nagata, T. Disappearance of plastid and mitochondrial nucleoids during the formation of generative cells of higher plants revealed by fluorescence microscopy. Protoplasma 1987, 141, 149–159. [Google Scholar] [CrossRef]
- Sato, M.; Sato, K. Maternal inheritance of mitochondrial DNA by diverse mechanisms to eliminate paternal mitochondrial DNA. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 1979–1984. [Google Scholar] [CrossRef] [Green Version]
- Rice, D.W.; Alverson, A.J.; Richardson, A.O.; Young, G.J.; Sanchez-Puerta, M.V.; Munzinger, J.; Barry, K.; Boore, J.L.; Zhang, Y.; DePamphilis, C.W.; et al. Horizontal transfer of entire genomes via mitochondrial fusion in the angiosperm Amborella. Science 2013, 342, 1468–1473. [Google Scholar] [CrossRef] [Green Version]
- Arakaki, N.; Nishihama, T.; Owaki, H.; Kuramoto, Y.; Suenaga, M.; Miyoshi, E.; Emoto, Y.; Shibata, H.; Shono, M.; Higuti, T. Dynamics of mitochondria during the cell cycle. Biol. Pharm. Bull. 2006, 29, 1962–1965. [Google Scholar] [CrossRef] [Green Version]
- Margineantu, D.H.; Cox, W.G.; Sundell, L.; Sherwood, S.W.; Beechem, J.M.; Capaldi, R.A. Cell cycle dependent morphology changes and associated mitochondrial DNA redistribution in mitochondria of human cell lines. Mitochondrion 2002, 1, 425–435. [Google Scholar] [CrossRef]
- Mitra, K.; Wunder, C.; Roysam, B.; Lin, G.; Lippincott-Schwartz, J. A hyperfused mitochondrial state achieved at G1–S regulates cyclin E buildup and entry into S phase. Proc. Nat. Acad. Sci. USA 2009, 106, 11960–11965. [Google Scholar] [CrossRef] [Green Version]
- Finkel, T.; Hwang, P.M. The Krebs cycle meets the cell cycle: Mitochondria and the G1–S transition. Proc. Nat. Acad. Sci. USA 2009, 106, 11825–11826. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Guo, X.; Liu, X.Q.; Zhang, L.; Huang, J.; Tan, L.; Lin, Z.; Nagawa, S.; Wang, D.Y. Changes in mitochondrial DNA levels during early embryogenesis in Torenia fournieri and Arabidopsis thaliana. Plant J. 2018, 95, 785–795. [Google Scholar] [CrossRef] [Green Version]
- Cupp, J.D.; Nielsen, B.L. Minireview: DNA replication in plant mitochondria. Mitochondrion 2014, 19, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, H.; Lo, Y.S.; Litvinchuk, A.; Wang, Y.T.; Jane, W.N.; Hsiao, L.J.; Chiang, K.S. Structural and functional characterizations of mung bean mitochondrial nucleoids. Nucleic Acids Res. 2005, 33, 4725–4739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, S.; Hagihara, S.; Izumi, M. Mitophagy in plants. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129916. [Google Scholar] [CrossRef]
- Ren, K.; Feng, L.; Sun, S.; Zhuang, X. Plant mitophagy in comparison to mammals: What is still missing? Int. J. Mol. Sci. 2021, 22, 1236. [Google Scholar] [CrossRef] [PubMed]
- Broda, M.; Millar, A.H.; Van Aken, O. Mitophagy: A mechanism for plant growth and survival. Trends Plant Sci. 2018, 23, 434–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto-Burgos, J.; Zhuang, X.; Jiang, L.; Bassham, D.C. Dynamics of autophagosome formation. Plant Physiol. 2018, 176, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minibayeva, F.; Dmitrieva, S.; Ponomareva, A.; Ryabovol, V. Oxidative stress-induced autophagy in plants: The role of mitochondria. Plant Physiol. Biochem. 2012, 59, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Chung, T.; Vierstra, R.D. AUTOPHAGY-RELATED11 plays a critical role in general autophagy-and senescence-induced mitophagy in Arabidopsis. Plant Cell 2014, 26, 788–807. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Hagihara, S.; Otomo, K.; Ishida, H.; Hidema, J.; Nemoto, T.; Izumi, M. Autophagy contributes to the quality control of leaf mitochondria. Plant Cell Physiol. 2021, 62, 229–247. [Google Scholar] [CrossRef]
- Ma, J.; Liang, Z.; Zhao, J.; Wang, P.; Ma, W.; Mai, K.K.; Andrade, J.A.F.; Zeng, Y.; Grujic, N.; Jiang, L.; et al. Friendly mediates membrane depolarization-induced mitophagy in Arabidopsis. Curr. Biol. 2021, 31, 1931–1934. [Google Scholar] [CrossRef]
- Seguí-Simarro, J.M.; Staehelin, L.A. Mitochondrial reticulation in shoot apical meristem cells of Arabidopsis provides a mechanism for homogenization of mtDNA prior to gamete formation. Plant Signal. Behav. 2009, 4, 168–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimata, Y.; Higaki, T.; Kurihara, D.; Ando, N.; Matsumoto, H.; Higashiyama, T.; Ueda, M. Mitochondrial dynamics and segregation during the asymmetric division of Arabidopsis zygotes. Quant. Plant Biol. 2020, 1, e3. [Google Scholar] [CrossRef]
- Mitra, K. Mitochondrial fission-fusion as an emerging key regulator of cell proliferation and differentiation. Bioessays 2013, 35, 955–964. [Google Scholar] [CrossRef]
- Iwata, R.; Casimir, P.; Vanderhaeghen, P. Mitochondrial dynamics in postmitotic cells regulate neurogenesis. Science 2020, 369, 858–862. [Google Scholar] [CrossRef] [PubMed]
- Paszkiewicz, G.; Gualberto, J.M.; Benamar, A.; Macherel, D.; Logan, D.C. Arabidopsis seed mitochondria are bioenergetically active immediately upon imbibition and specialize via biogenesis in preparation for autotrophic growth. Plant Cell 2017, 29, 109–128. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rose, R.J. Contribution of Massive Mitochondrial Fusion and Subsequent Fission in the Plant Life Cycle to the Integrity of the Mitochondrion and Its Genome. Int. J. Mol. Sci. 2021, 22, 5429. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115429
Rose RJ. Contribution of Massive Mitochondrial Fusion and Subsequent Fission in the Plant Life Cycle to the Integrity of the Mitochondrion and Its Genome. International Journal of Molecular Sciences. 2021; 22(11):5429. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115429
Chicago/Turabian StyleRose, Ray J. 2021. "Contribution of Massive Mitochondrial Fusion and Subsequent Fission in the Plant Life Cycle to the Integrity of the Mitochondrion and Its Genome" International Journal of Molecular Sciences 22, no. 11: 5429. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115429