
Ancient Bacterial Class Alphaproteobacteria Cytochrome P450 Monooxygenases Can Be Found in Other Bacterial Species

 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
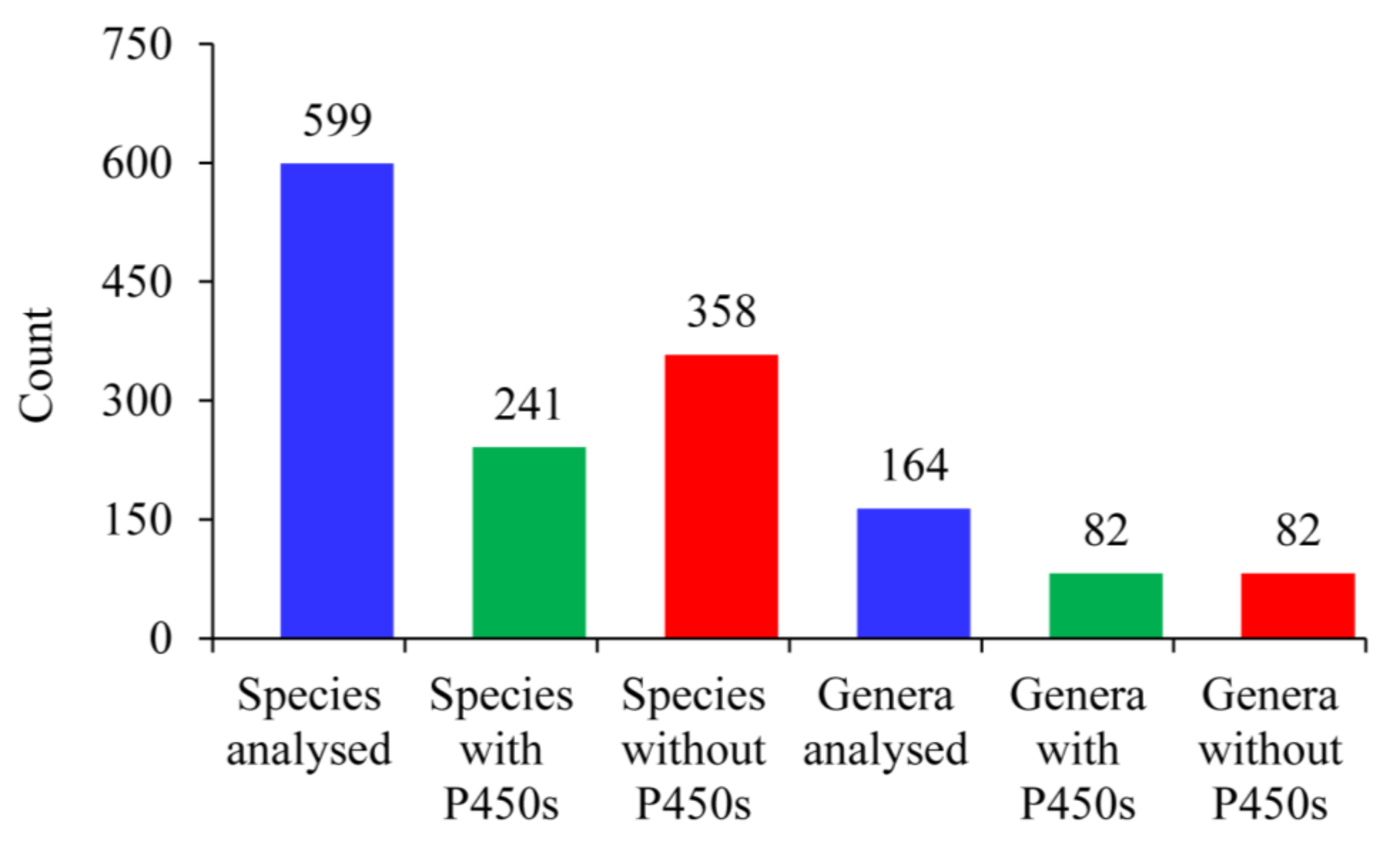
2.1. Only 40% of Alphaproteobacterial Species Have P450s
2.2. Alphaproteobacterial Species Have the Highest Number of P450 Families Next to Streptomyces
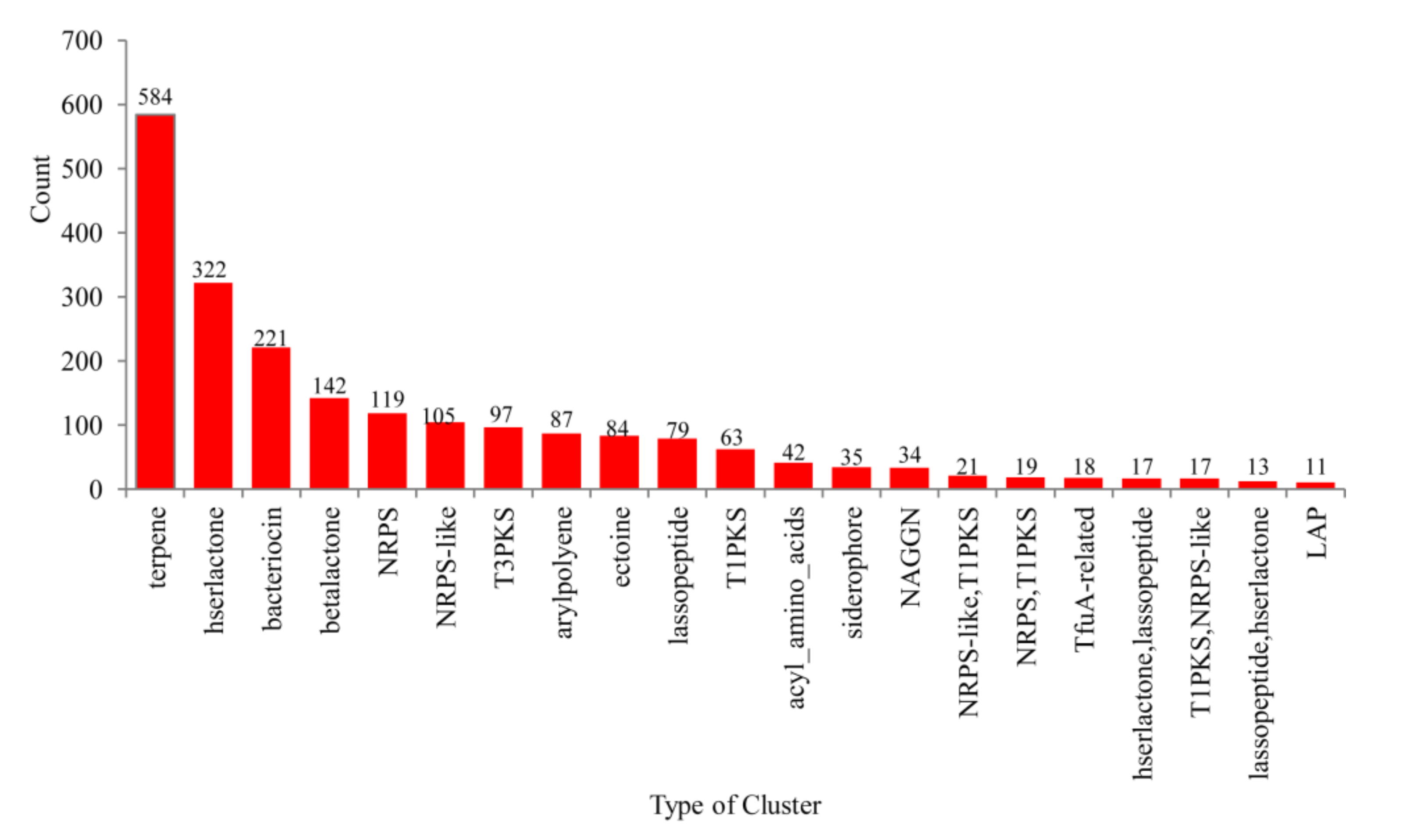
2.3. Only a Few P450s Are Involved in Secondary Metabolism in Alphaproteobacterial Species
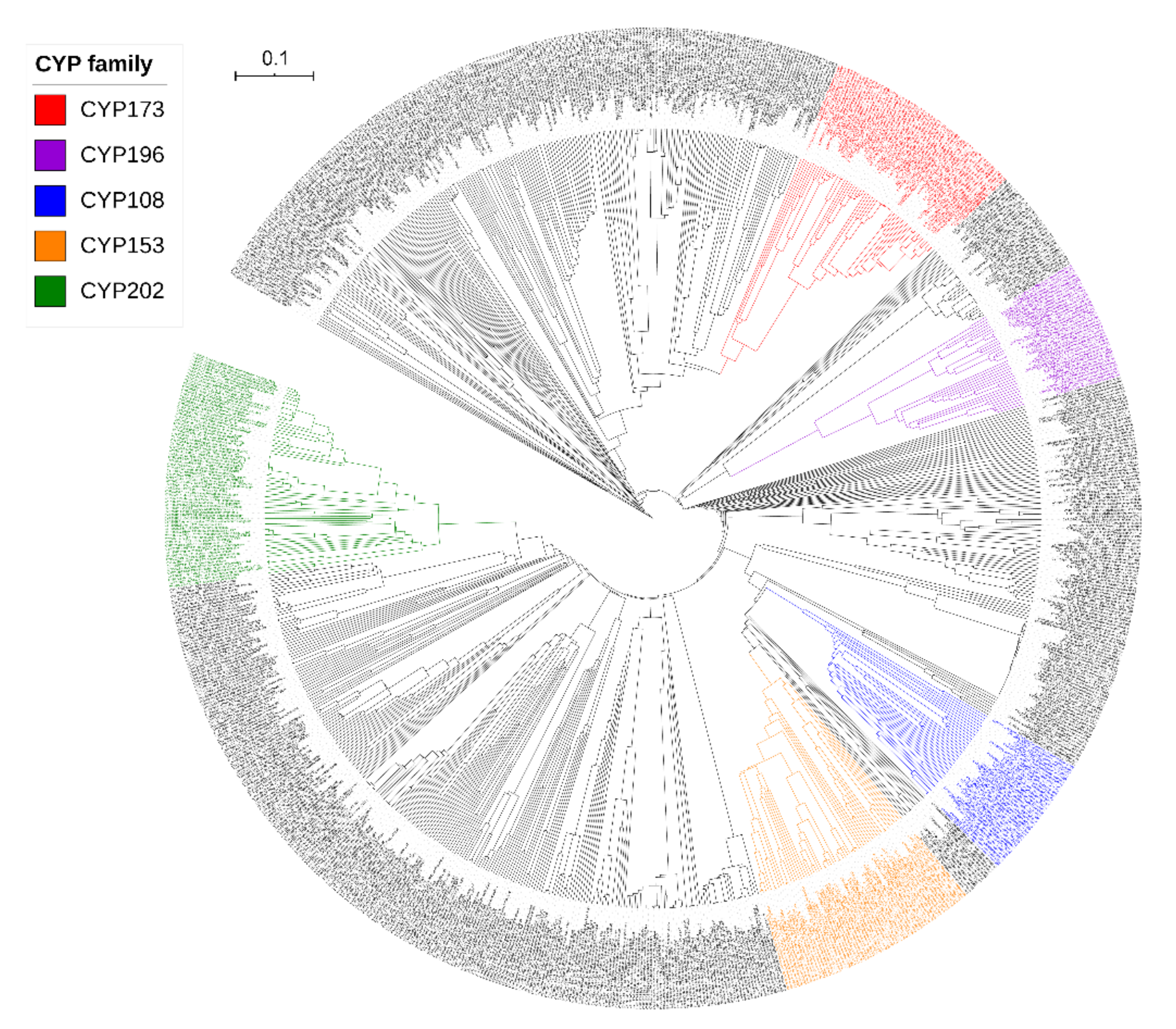
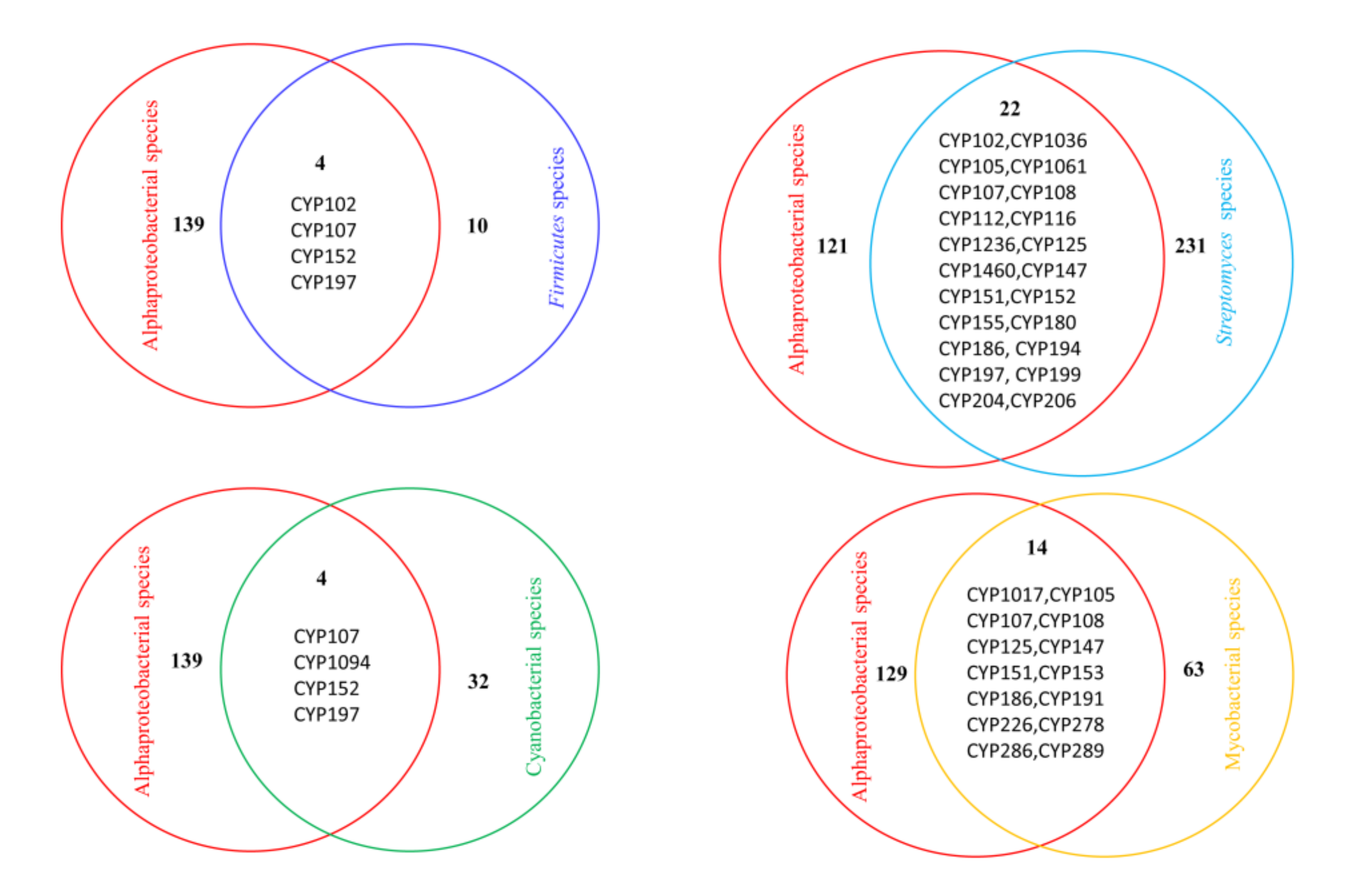
2.4. Alphaproteobacterial P450 Families Can Be Found in Other Bacterial Species
2.5. Functional Analysis of Alphaproteobacterial P450s
3. Materials and Methods
3.1. Species and Database
3.2. Genome Data Mining and Annotation of P450s
3.3. Phylogenetic Analysis of P450s
3.4. Generation of P450 Profile Heat Maps
3.5. Secondary Metabolite BGC Analysis and Identification of P450s That Are Part of BGCs
3.6. Identification of Cholesterol Side-Chain Oxidizing Genes/Proteins
3.7. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamazaki, H. Fifty Years of Cytochrome P450 Research; Springer Japan: Tokyo, Japan, 2014; pp. 1–409. [Google Scholar]
- Li, Z.; Jiang, Y.; Guengerich, F.P.; Ma, L.; Li, S.; Zhang, W. Engineering cytochrome P450 enzyme systems for biomedical and biotechnological applications. J. Biol. Chem. 2020, 295, 833–849. [Google Scholar] [CrossRef]
- Urlacher, V.B.; Girhard, M. Cytochrome P450 monooxygenases in biotechnology and synthetic biology. Trends Biotechnol. 2019, 37, 882–897. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R. Cytochrome P450 diversity in the tree of life. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2018, 1866, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Lamb, D.C.; Follmer, A.H.; Goldstone, J.V.; Nelson, D.R.; Warrilow, A.G.; Price, C.L.; True, M.Y.; Kelly, S.L.; Poulos, T.L.; Stegeman, J.J. On the occurrence of cytochrome P450 in viruses. Proc. Natl. Acad. Sci. USA 2019, 116, 12343–12352. [Google Scholar] [CrossRef] [Green Version]
- Guengerich, F.P. A history of the roles of cytochrome P450 enzymes in the toxicity of drugs. Toxicol. Res. 2021, 37, 1–23. [Google Scholar] [CrossRef]
- Lepesheva, G.I.; Friggeri, L.; Waterman, M.R. CYP51 as drug targets for fungi and protozoan parasites: Past, present and future. Parasitology 2018, 145, 1820–1836. [Google Scholar] [CrossRef]
- Jawallapersand, P.; Mashele, S.S.; Kovačič, L.; Stojan, J.; Komel, R.; Pakala, S.B.; Kraševec, N.; Syed, K. Cytochrome P450 monooxygenase CYP53 family in fungi: Comparative structural and evolutionary analysis and its role as a common alternative anti-fungal drug target. PLoS ONE 2014, 9, e107209. [Google Scholar] [CrossRef] [Green Version]
- Girvan, H.M.; Munro, A.W. Applications of microbial cytochrome P450 enzymes in biotechnology and synthetic biology. Curr. Opin. Chem. Biol. 2016, 31, 136–145. [Google Scholar] [CrossRef]
- McLean, K.J.; Leys, D.; Munro, A.W. Microbial cytochrome P450s In Cytochrome P450: Structure, Mechanism, and Biochemistry, 4th ed.; Oritz de Montellano, P.R., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 261–407. [Google Scholar]
- Kelly, S.L.; Kelly, D.E. Microbial cytochromes P450: Biodiversity and biotechnology. Where do cytochromes P450 come from, what do they do and what can they do for us? Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120476. [Google Scholar] [CrossRef] [Green Version]
- Girhard, M.; Bakkes, P.J.; Mahmoud, O.; Urlacher, V.B. P450 Biotechnology. In Cytochrome P450; Springer: Berlin/Heidelberg, Germany, 2015; pp. 451–520. [Google Scholar]
- Di Nardo, G.; Gilardi, G. Natural compounds as pharmaceuticals: The key role of cytochromes P450 reactivity. Trends Biochem. Sci. 2020, 45, 511–525. [Google Scholar] [CrossRef]
- Andersen, J.F.; Tatsuta, K.; Gunji, H.; Ishiyama, T.; Hutchinson, C.R. Substrate specificity of 6-deoxyerythronolide B hydroxylase, a bacterial cytochrome P450 of erythromycin A biosynthesis. Biochemistry 1993, 32, 1905–1913. [Google Scholar] [CrossRef]
- Bischoff, D.; Bister, B.; Bertazzo, M.; Pfeifer, V.; Stegmann, E.; Nicholson, G.J.; Keller, S.; Pelzer, S.; Wohlleben, W.; Süssmuth, R. The biosynthesis of vancomycin-type glycopeptide antibiotics-a model for oxidative side-chain cross-linking by oxygenases coupled to the action of peptide synthetases. ChemBioChem 2005, 6, 267–272. [Google Scholar] [CrossRef]
- Jennewein, S.; Park, H.; DeJong, J.M.; Long, R.M.; Bollon, A.P.; Croteau, R.B. Coexpression in yeast of Taxus cytochrome P450 reductase with cytochrome P450 oxygenases involved in Taxol biosynthesis. Biotechnol. Bioeng. 2005, 89, 588–598. [Google Scholar] [CrossRef]
- Van Beilen, J.B.; Holtackers, R.; Lüscher, D.; Bauer, U.; Witholt, B.; Duetz, W.A. Biocatalytic production of perillyl alcohol from limonene by using a novel Mycobacterium sp. cytochrome P450 alkane hydroxylase expressed in Pseudomonas putida. Appl. Environ. Microbiol. 2005, 71, 1737–1744. [Google Scholar] [CrossRef] [Green Version]
- Podust, L.M.; Sherman, D.H. Diversity of P450 enzymes in the biosynthesis of natural products. Nat. Prod. Rep. 2012, 29, 1251–1266. [Google Scholar] [CrossRef] [Green Version]
- Greule, A.; Stok, J.E.; De Voss, J.J.; Cryle, M.J. Unrivalled diversity: The many roles and reactions of bacterial cytochromes P450 in secondary metabolism. Nat. Prod. Rep. 2018, 35, 757–791. [Google Scholar] [CrossRef] [Green Version]
- Cimermancic, P.; Medema, M.H.; Claesen, J.; Kurita, K.; Brown, L.C.W.; Mavrommatis, K.; Pati, A.; Godfrey, P.A.; Koehrsen, M.; Clardy, J.; et al. Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 2014, 158, 412–421. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.R. Cytochrome P450 nomenclature. Methods Mol. Biol. 1998, 107, 15–24. [Google Scholar]
- Nelson, D.R. Cytochrome P450 nomenclature, 2004. Methods Mol. Biol. 2006, 320, 1–10. [Google Scholar]
- Nelson, D.R. The cytochrome p450 homepage. Hum. Genom. 2009, 4, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Padayachee, T.; Nzuza, N.; Chen, W.; Nelson, D.R.; Syed, K. Impact of lifestyle on cytochrome P450 monooxygenase repertoire is clearly evident in the bacterial phylum Firmicutes. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Gómez, S.; Hess, S.; Burger, G.; Lang, B.F.; Susko, E.; Slamovits, C.H.; Roger, A.J. An updated phylogeny of the Alphaproteobacteria reveals that the parasitic Rickettsiales and Holosporales have independent origins. eLife 2019, 8, 42535. [Google Scholar] [CrossRef] [PubMed]
- Ettema, T.J.; Andersson, S.G. The α-proteobacteria: The Darwin finches of the bacterial world. Biol. Lett. 2009, 5, 429–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhya, I.; Hansen, R.; El-Omar, E.M.; Hold, G.L. IBD—what role do Proteobacteria play? Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 219–230. [Google Scholar] [CrossRef]
- Kersters, K.; De Vos, P.; Gillis, M.; Swings, J.; Vandamme, P.; Stackebrandt, E. Introduction to the Proteobacteria. In The Prokaryotes; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2006; pp. 3–37. [Google Scholar]
- Gupta, R.S. Protein signatures distinctive of Alphaproteobacteria and its subgroups and a model for α –Proteobacterial evolution. Crit. Rev. Microbiol. 2005, 31, 101–135. [Google Scholar] [CrossRef]
- Phung, N.T.; Lee, J.; Kang, K.H.; Chang, I.S.; Gadd, G.M.; Kim, B.H. Analysis of microbial diversity in oligotrophic microbial fuel cells using 16S rDNA sequences. FEMS Microbiol. Lett. 2004, 233, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Timmermans, M.L.; Paudel, Y.P.; Ross, A.C. Investigating the biosynthesis of natural products from marine Proteobacteria: A survey of molecules and strategies. Mar. Drugs 2017, 15, 235. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Wright, G.D. An ecological perspective of microbial secondary metabolism. Curr. Opin. Biotechnol. 2011, 22, 552–558. [Google Scholar] [CrossRef]
- Tsukimoto, M.; Nagaoka, M.; Shishido, Y.; Fujimoto, J.; Nishisaka, F.; Matsumoto, S.; Harunari, E.; Imada, C.; Matsuzaki, T. Bacterial production of the tunicate-derived antitumor cyclic depsipeptide didemnin B. J. Nat. Prod. 2011, 74, 2329–2331. [Google Scholar] [CrossRef]
- Oh, D.-C.; Strangman, W.K.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Thalassospiramides A (I) and B (II), Immunosuppressive peptides from the marine bacterium Thalassospira sp. Org. Lett. 2007, 9, 1525–1528. [Google Scholar] [CrossRef]
- Meena, K.K.; Kumar, M.; Kalyuzhnaya, M.G.; Yandigeri, M.S.; Singh, D.P.; Saxena, A.K.; Arora, D.K. Epiphytic pink-pigmented methylotrophic bacteria enhance germination and seedling growth of wheat (Triticum aestivum) by producing phytohormone. Antonie van Leeuwenhoek 2011, 101, 777–786. [Google Scholar] [CrossRef]
- Siqueira, A.F.; Ormeño-Orrillo, E.; Souza, R.C.; Rodrigues, E.P.; Almeida, L.G.P.; Barcellos, F.G.; Batista, J.S.S.; Nakatani, A.S.; Martínez-Romero, E.; Vasconcelos, A.T.R.; et al. Comparative genomics of Bradyrhizobium japonicum CPAC 15 and Bradyrhizobium diazoefficiens CPAC 7: Elite model strains for understanding symbiotic performance with soybean. BMC Genom. 2014, 15, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.L.; Waqas, M.; Kang, S.-M.; Al-Harrasi, A.; Hussain, J.; Al-Rawahi, A.; Al-Khiziri, S.; Ullah, I.; Ali, L.; Jung, H.-Y.; et al. Bacterial endophyte Sphingomonas sp. LK11 produces gibberellins and IAA and promotes tomato plant growth. J. Microbiol. 2014, 52, 689–695. [Google Scholar] [CrossRef]
- Long, C.; Lu, X.-L.; Gao, Y.; Jiao, B.-H.; Liu, X.-Y. Description of a sulfitobacter strain and its extracellular cyclodipeptides. Evidence Based Complement. Altern. Med. 2011, 2011, 1–6. [Google Scholar]
- Senate, L.M.; Tjatji, M.P.; Pillay, K.; Chen, W.; Zondo, N.M.; Syed, P.R.; Mnguni, F.C.; Chiliza, Z.E.; Bamal, H.D.; Karpoormath, R.; et al. Similarities, variations, and evolution of cytochrome P450s in Streptomyces versus Mycobacterium. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mnguni, F.C.; Padayachee, T.; Chen, W.; Gront, D.; Yu, J.-H.; Nelson, D.R.; Syed, K. More P450s are involved in secondary metabolite biosynthesis in Streptomyces compared to Bacillus, Cyanobacteria and Mycobacterium. Int. J. Mol. Sci. 2020, 21, 4814. [Google Scholar] [CrossRef]
- Khumalo, M.J.; Nzuza, N.; Padayachee, T.; Chen, W.; Yu, J.-H.; Nelson, D.R.; Syed, K. Comprehensive analyses of cytochrome P450 monooxygenases and secondary metabolite biosynthetic gene clusters in Cyanobacteria. Int. J. Mol. Sci. 2020, 21, 656. [Google Scholar] [CrossRef] [Green Version]
- Parvez, M.; Qhanya, L.B.; Mthakathi, N.T.; Kgosiemang, I.K.R.; Bamal, H.D.; Pagadala, N.S.; Xie, T.; Yang, H.; Chen, H.; Theron, C.W.; et al. Molecular evolutionary dynamics of cytochrome P450 monooxygenases across kingdoms: Special focus on mycobacterial P450s. Sci. Rep. 2016, 6, 33099. [Google Scholar] [CrossRef] [Green Version]
- Mthethwa, B.C.; Chen, W.; Ngwenya, M.L.; Kappo, A.P.; Syed, P.R.; Karpoormath, R.; Yu, J.-H.; Nelson, D.R.; Syed, K. Comparative analyses of cytochrome P450s and those associated with secondary metabolism in Bacillus species. Int. J. Mol. Sci. 2018, 19, 3623. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.R.; Kamataki, T.; Waxman, D.J.; Guengerich, F.P.; Estabrook, R.W.; Feyereisen, R.; Gonzalez, F.J.; Coon, M.J.; Gunsalus, I.C.; Gotoh, O.; et al. The P450 Superfamily: Update on new sequences, gene mapping, accession numbers, early trivial names of enzymes, and nomenclature. DNA Cell Biol. 1993, 12, 1–51. [Google Scholar] [CrossRef]
- Nelson, D.R. Cytochrome P450 nomenclature. In Cytochrome P450 Protocols; Springer: Berlin/Heidelberg, Germany, 1998; pp. 15–24. [Google Scholar]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- McLean, K.J.; Lafite, P.; Levy, C.; Cheesman, M.R.; Mast, N.; Pikuleva, I.A.; Leys, D.; Munro, A.W. The structure of Mycobacterium tuberculosis CYP125: Molecular basis for cholesterol binding in a P450 needed for host infection. J Biol Chem 2009, 284, 35524–35533. [Google Scholar] [CrossRef] [Green Version]
- Ouellet, H.; Guan, S.; Johnston, J.B.; Chow, E.D.; Kells, P.M.; Burlingame, A.L.; Cox, J.S.; Podust, L.M.; De Montellano, P.R.O. Mycobacterium tuberculosis CYP125A1, a steroid C27 monooxygenase that detoxifies intracellularly generated cholest-4-en-3-one. Mol. Microbiol. 2010, 77, 730–742. [Google Scholar] [CrossRef] [Green Version]
- Ortiz de Montellano, P.R. Potential drug targets in the Mycobacterium tuberculosis cytochrome P450 system. J. Inorg. Biochem. 2018, 180, 235–245. [Google Scholar] [CrossRef]
- Holert, J.; Cardenas, E.; Bergstrand, L.H.; Zaikova, E.; Hahn, A.S.; Hallam, S.J.; Mohn, W.W. Metagenomes reveal global distribution of bacterial steroid catabolism in natural, engineered, and host environments. mBio 2018, 9, e02345-17. [Google Scholar] [CrossRef] [Green Version]
- Van Wyk, R.; Van Wyk, M.; Mashele, S.S.; Nelson, D.R.; Syed, K. Comprehensive comparative analysis of cholesterol catabolic genes/proteins in mycobacterial species. Int. J. Mol. Sci. 2019, 20, 1032. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y.; Furumichi, M.; Morishima, K.; Tanabe, M. New approach for understanding genome variations in KEGG. Nucleic Acids Res. 2019, 47, D590–D595. [Google Scholar] [CrossRef] [Green Version]
- Hall, E.A.; Bell, S.G. The efficient and selective biocatalytic oxidation of norisoprenoid and aromatic substrates by CYP101B1 from Novosphingobium aromaticivorans DSM12444. RSC Adv. 2014, 5, 5762–5773. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Bell, S.G.; Yang, W.; Hao, Y.; Rees, N.H.; Bartlam, M.; Zhou, W.; Wong, L.-L.; Rao, Z. Structural Analysis of CYP101C1 from Novosphingobium aromaticivorans DSM12444. ChemBioChem 2010, 12, 88–99. [Google Scholar] [CrossRef]
- Yang, W.; Bell, S.G.; Wang, H.; Zhou, W.; Hoskins, N.; Dale, A.; Bartlam, M.; Wong, L.-L.; Rao, Z. Molecular characterization of a class I P450 electron transfer system from Novosphingobium aromaticivorans DSM12444. J. Biol. Chem. 2010, 285, 27372–27384. [Google Scholar] [CrossRef] [Green Version]
- Batabyal, D.; Poulos, T.L. Crystal structures and functional characterization of wild-type CYP101D1 and its active site mutants. Biochem. 2013, 52, 8898–8906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Bell, S.G.; Wang, H.; Zhou, W.; Bartlam, M.; Wong, L.-L.; Rao, Z. The structure of CYP101D2 unveils a potential path for substrate entry into the active site. Biochem. J. 2010, 433, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.G.; Yang, W.; Yorke, J.A.; Zhou, W.; Wang, H.; Harmer, J.; Copley, R.; Zhang, A.; Zhou, R.; Bartlam, M.; et al. Structure and function of CYP108D1 from Novosphingobium aromaticivorans DSM12444: An aromatic hydrocarbon-binding P450 enzyme. Acta Crystallogr. Sect. D Biol. Crystallogr. 2012, 68, 277–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.; Huang, C.; Zhang, A.; Bell, S.G.; Zhou, W.; Wong, L.-L. Crystallization and preliminary X-ray analysis of CYP153C1 from Novosphingobium aromaticivorans DSM12444. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2011, 67, 964–967. [Google Scholar] [CrossRef]
- Krings, U.; Berger, R.G. Terpene bioconversion–how does its future look? Nat. Prod. Commun. 2010, 5, 1934578X1000500927. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.-A.; Han, S.; Lim, Y.-R.; Kim, V.; Kim, H.; Kim, D. Streptomyces cytochrome P450 enzymes and their roles in the biosynthesis of macrolide therapeutic agents. Biomol. Ther. 2019, 27, 127–133. [Google Scholar] [CrossRef]
- Agematu, H.; Matsumoto, N.; Fujii, Y.; Kabumoto, H.; Doi, S.; Machida, K.; Ishikawa, J.; Arisawa, A. Hydroxylation of testosterone by bacterial cytochromes P450 using the Escherichia coli expression system. Biosci. Biotechnol. Biochem. 2006, 70, 307–311. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.; Xu, F.; Bell, S.G.; Pang, X.; Bartlam, M.; Wong, L.-L. Purification, crystallization and preliminary crystallographic analysis of CYP 195A2, a P450 enzyme from Rhodopseudomonas palustris. Protein Pept. Lett. 2008, 15, 423–426. [Google Scholar] [CrossRef]
- Furuya, T.; Kino, K. Discovery of 2-naphthoic acid monooxygenases by genome mining and their use as biocatalysts. ChemSusChem 2009, 2, 645–649. [Google Scholar] [CrossRef]
- Bell, S.G.; Yang, W.; Tan, A.B.H.; Zhou, R.; Johnson, E.O.D.; Zhang, A.; Zhou, W.; Rao, Z.; Wong, L.-L. The crystal structures of 4-methoxybenzoate bound CYP199A2 and CYP199A4: Structural changes on substrate binding and the identification of an anion binding site. Dalton Trans. 2012, 41, 8703–8714. [Google Scholar] [CrossRef]
- Coleman, T.; Wong, S.H.; Podgorski, M.N.; Bruning, J.B.; De Voss, J.J.; Bell, S.G. Cytochrome P450 CYP199A4 from Rhodopseudomonas palustris catalyzes heteroatom dealkylations, sulfoxidation, and amide and cyclic hemiacetal formation. ACS Catal. 2018, 8, 5915–5927. [Google Scholar] [CrossRef] [Green Version]
- Berne, C.; Pignol, D.; Lavergne, J.; Garcia, D. CYP201A2, a cytochrome P450 from Rhodopseudomonas palustris, plays a key role in the biodegradation of tributyl phosphate. Appl. Microbiol. Biotechnol. 2007, 77, 135–144. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; I Hurwitz, D.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Kuma, K.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef]
- Boc, A.; Diallo, A.B.; Makarenkov, V. T-REX: A web server for inferring, validating and visualizing phylogenetic trees and networks. Nucleic Acids Res. 2012, 40, W573–W579. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Howe, E.A.; Sinha, R.; Schlauch, D.; Quackenbush, J. RNA-Seq analysis in MeV. Bioinformatics 2011, 27, 3209–3210. [Google Scholar] [CrossRef] [Green Version]
- Msomi, N.K.; Padayachee, R.; Nzuza, N.; Syed, P.R.; Krys, J.D.; Chen, W.; Gront, D.; Nelson, R.D.; Syed, K. In silico analysis of P450s and their role in secondary metabolism in the bacterial class Gammaproteobacteria. Int. J. Mol. Sci. 2021, 26, 1538. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Secondary Metabolite | Species (Source) | Biological Function | Reference |
|---|---|---|---|
| Didemnin B | Tistrella mobilis YIT 12409 | Anti-viral and anti-tumor properties | [33] |
| Thalassospiramide | Thalassospira sp. CNJ-328 | Immunosuppression | [34] |
| Tropodithietic acid | Phaeobacter gallaeciensis and P. porticola | Antibiotic, biocontrol and anti-cancer | [31] |
| Phytohormone | Methylobacterium sp. NC4 | Improve plant growth | [35] |
| 6-gingerol and curcumin | Bradyrhizobium japonicum CPAC 15 and Bradyrhizobium diazoefficiens CPAC 7 | Antitumor activity | [36] |
| Gibberellins, salicylic acid, indole-3-acetic acid, zeatin, and abscisic acid | Sphingomonas species | Promote plant germination and growth | [37] |
| Cyclodipeptides | Sulfitobacter species | Bioactive agents | [38] |
| Category | Alphaproteobacterial Species | Firmicutes Species | Streptomyces Species | Mycobacterial Species | Cyanobacterial Species |
|---|---|---|---|---|---|
| Total no of species analyzed | 599 | 972 | 203 | 60 | 114 |
| No of species that have P450s | 241 | 229 | 203 | 60 | 114 |
| No of P450s | 874 | 712 | 5460 | 1784 | 341 |
| No of families | 143 | 14 | 253 | 77 | 36 |
| No of subfamilies | 214 | 53 | 698 | 132 | 79 |
| Dominant P450 family | CYP202 | CYP107 | CYP107 | CYP125 | CYP110 |
| Average no. of P450s | 4 | 1 | 27 | 30 | 3 |
| P450 diversity percentage | 0.07 | 0.008 | 0.02 | 0.07 | 0.09 |
| No of P450s part of BGCs | 21 | 126 | 1231 | 204 | 27 |
| Percentage of P450s part of BGCs | 2.4 | 18 | 23 | 11 | 8 |
| Reference(s) | This work | [24,43] | [39,40] | [39,42] | [41] |
| P450 Family | P450 Count | Percentage Count | Subfamily | P450 Count |
|---|---|---|---|---|
| CYP1010 | 1 | 0.11% | A | 1 |
| CYP1017 | 1 | 0.11% | A | 1 |
| CYP101 | 10 | 1.14% | B | 1 |
| C | 3 | |||
| D | 2 | |||
| E | 2 | |||
| Q | 1 | |||
| R | 1 | |||
| CYP102 | 12 | 1.37% | A | 10 |
| K | 1 | |||
| AC | 1 | |||
| CYP1036 | 2 | 0.23% | B | 1 |
| C | 1 | |||
| CYP103 | 4 | 0.46% | A | 4 |
| CYP104 | 13 | 1.49% | A | 13 |
| CYP105 | 4 | 0.46% | BF | 3 |
| CX | 1 | |||
| CYP1061 | 2 | 0.23% | B | 2 |
| CYP1068 | 4 | 0.46% | A | 4 |
| CYP1075 | 1 | 0.11% | A | 1 |
| CYP1076 | 1 | 0.11% | A | 1 |
| CYP1077 | 1 | 0.11% | A | 1 |
| CYP1078 | 8 | 0.92% | A | 7 |
| C | 1 | |||
| CYP107 | 10 | 1.14% | AN | 8 |
| _ | 1 | |||
| JE | 1 | |||
| CYP1081 | 2 | 0.23% | B | 2 |
| CYP1082 | 1 | 0.11% | A | 1 |
| CYP1083 | 5 | 0.57% | A | 3 |
| C | 2 | |||
| CYP1086 | 1 | 0.11% | B | 1 |
| CYP1088 | 2 | 0.23% | B | 2 |
| CYP1089 | 1 | 0.11% | A | 1 |
| CYP108 | 37 | 4.23% | A | 2 |
| D | 6 | |||
| G | 18 | |||
| L | 1 | |||
| P | 2 | |||
| U | 1 | |||
| V | 1 | |||
| W | 1 | |||
| X | 5 | |||
| CYP1090 | 2 | 0.23% | C | 2 |
| CYP1091 | 3 | 0.34% | A | 3 |
| CYP1094 | 4 | 0.46% | A | 3 |
| B | 1 | |||
| CYP1098 | 6 | 0.69% | A | 6 |
| CYP1101 | 29 | 3.32% | A | 29 |
| CYP1104 | 10 | 1.14% | A | 4 |
| C | 3 | |||
| E | 3 | |||
| CYP1107 | 16 | 1.83% | A | 12 |
| B | 3 | |||
| C | 1 | |||
| CYP1111 | 7 | 0.80% | A | 5 |
| B | 1 | |||
| C | 1 | |||
| CYP1116 | 3 | 0.34% | B | 3 |
| CYP1118 | 1 | 0.11% | B | 1 |
| CYP111 | 2 | 0.23% | A | 1 |
| B | 1 | |||
| CYP112 | 23 | 2.63% | A | 23 |
| CYP1135 | 1 | 0.11% | A | 1 |
| CYP1137 | 1 | 0.11% | A | 1 |
| CYP1138 | 2 | 0.23% | B | 2 |
| CYP1141 | 1 | 0.11% | A | 1 |
| CYP1145 | 1 | 0.11% | A | 1 |
| CYP114 | 22 | 2.52% | A | 22 |
| CYP1155 | 1 | 0.11% | B | 1 |
| CYP115 | 1 | 0.11% | A | 1 |
| CYP116 | 1 | 0.11% | B | 1 |
| CYP1170 | 7 | 0.80% | A | 7 |
| CYP1171 | 1 | 0.11% | A | 1 |
| CYP1173 | 1 | 0.11% | A | 1 |
| CYP1174 | 2 | 0.23% | A | 2 |
| CYP1175 | 1 | 0.11% | A | 1 |
| CYP117 | 22 | 2.52% | A | 22 |
| CYP1181 | 1 | 0.11% | A | 1 |
| CYP1182 | 1 | 0.11% | A | 1 |
| CYP1186 | 2 | 0.23% | A | 2 |
| CYP1187 | 1 | 0.11% | A | 1 |
| CYP1204 | 2 | 0.23% | A | 2 |
| CYP1206 | 1 | 0.11% | B | 1 |
| CYP1221 | 2 | 0.23% | B | 2 |
| CYP1229 | 4 | 0.46% | A | 1 |
| B | 2 | |||
| C | 1 | |||
| CYP1246 | 2 | 0.23% | A | 2 |
| CYP1250 | 1 | 0.11% | A | 1 |
| CYP1258 | 1 | 0.11% | A | 1 |
| CYP125 | 1 | 0.11% | P | 1 |
| CYP1275 | 1 | 0.11% | B | 1 |
| CYP127 | 19 | 2.17% | A | 18 |
| C | 1 | |||
| CYP1281 | 1 | 0.11% | A | 1 |
| CYP1291 | 5 | 0.57% | B | 3 |
| C | 2 | |||
| CYP1302 | 2 | 0.23% | A | 2 |
| CYP1311 | 2 | 0.23% | A | 2 |
| CYP1312 | 1 | 0.11% | A | 1 |
| CYP1326 | 2 | 0.23% | B | 2 |
| CYP1330 | 1 | 0.11% | A | 1 |
| CYP1337 | 1 | 0.11% | A | 1 |
| CYP133 | 1 | 0.11% | F | 1 |
| CYP1349 | 1 | 0.11% | A | 1 |
| CYP1350 | 1 | 0.11% | A | 1 |
| CYP1371 | 1 | 0.11% | B | 1 |
| CYP1376 | 2 | 0.23% | B | 1 |
| C | 1 | |||
| CYP1384 | 1 | 0.11% | A | 1 |
| CYP1396 | 1 | 0.11% | A | 1 |
| CYP1405 | 1 | 0.11% | B | 1 |
| CYP1406 | 3 | 0.34% | B | 2 |
| C | 1 | |||
| CYP1460 | 1 | 0.11% | C | 1 |
| CYP147 | 16 | 1.83% | D | 16 |
| CYP1515 | 2 | 0.23% | A | 2 |
| CYP151 | 2 | 0.23% | C | 2 |
| CYP152 | 13 | 1.49% | B | 4 |
| C | 3 | |||
| E | 2 | |||
| P | 1 | |||
| AA | 1 | |||
| AB | 2 | |||
| CYP153 | 60 | 6.86% | A | 44 |
| C | 2 | |||
| D | 14 | |||
| CYP155 | 4 | 0.46% | J | 3 |
| K | 1 | |||
| CYP1591 | 1 | 0.11% | A | 1 |
| CYP1597 | 1 | 0.11% | A | 1 |
| CYP1732 | 3 | 0.34% | A | 3 |
| CYP1733 | 2 | 0.23% | A | 1 |
| B | 1 | |||
| CYP1734 | 1 | 0.11% | A | 1 |
| CYP1735 | 1 | 0.11% | A | 1 |
| CYP1736 | 1 | 0.11% | A | 1 |
| CYP1737 | 2 | 0.23% | A | 2 |
| CYP1738 | 1 | 0.11% | A | 1 |
| CYP1739 | 2 | 0.23% | A | 2 |
| CYP173 | 60 | 6.86% | A | 45 |
| B | 8 | |||
| C | 3 | |||
| G | 1 | |||
| H | 1 | |||
| J | 2 | |||
| CYP1740 | 1 | 0.11% | A | 1 |
| CYP1741 | 1 | 0.11% | A | 1 |
| CYP1742 | 1 | 0.11% | A | 1 |
| CYP1743 | 1 | 0.11% | A | 1 |
| CYP1744 | 1 | 0.11% | A | 1 |
| CYP1745 | 1 | 0.11% | A | 1 |
| CYP1746 | 1 | 0.11% | A | 1 |
| CYP1747 | 1 | 0.11% | A | 1 |
| CYP1748 | 1 | 0.11% | A | 1 |
| CYP1749 | 2 | 0.23% | A | 1 |
| B | 1 | |||
| CYP1750 | 1 | 0.11% | A | 1 |
| CYP1751 | 1 | 0.11% | A | 1 |
| CYP1752 | 4 | A | 4 | |
| CYP1753 | 1 | 0.11% | A | 1 |
| CYP1754 | 1 | 0.11% | A | 1 |
| CYP1755 | 1 | 0.11% | A | 1 |
| CYP180 | 1 | 0.11% | D | 1 |
| CYP186 | 7 | 0.80% | K | 7 |
| CYP191 | 1 | 0.11% | B | 1 |
| CYP192 | 3 | 0.34% | A | 3 |
| CYP193 | 17 | 1.95% | A | 17 |
| CYP194 | 10 | 1.14% | A | 10 |
| CYP195 | 21 | 2.40% | A | 18 |
| D | 1 | |||
| E | 2 | |||
| CYP196 | 37 | 4.23% | A | 34 |
| B | 2 | |||
| C | 1 | |||
| CYP197 | 2 | 0.23% | R | 2 |
| CYP199 | 16 | 1.83% | A | 11 |
| B | 1 | |||
| J | 1 | |||
| K | 1 | |||
| L | 1 | |||
| M | 1 | |||
| CYP200 | 8 | 0.92% | A | 5 |
| B | 2 | |||
| C | 1 | |||
| CYP201 | 31 | 3.55% | A | 25 |
| B | 3 | |||
| C | 3 | |||
| CYP202 | 70 | 8.01% | A | 46 |
| B | 24 | |||
| CYP203 | 13 | 1.49% | A | 12 |
| B | 1 | |||
| CYP204 | 2 | 0.23% | A | 2 |
| CYP206 | 19 | 2.17% | A | 9 |
| B | 1 | |||
| C | 3 | |||
| D | 6 | |||
| CYP219 | 2 | 0.23% | A | 2 |
| CYP223 | 3 | 0.34% | A | 2 |
| E | 1 | |||
| CYP224 | 1 | 0.11% | A | 1 |
| CYP225 | 7 | 0.80% | A | 7 |
| CYP226 | 2 | 0.23% | C | 1 |
| D | 1 | |||
| CYP278 | 2 | 0.23% | A | 1 |
| C | 1 | |||
| CYP286 | 1 | 0.11% | C | 1 |
| CYP288 | 2 | 0.23% | B | 2 |
| CYP289 | 33 | 3.78% | A | 33 |
| CYP290 | 6 | 0.69% | A | 5 |
| B | 1 | |||
| CYP2140 | 1 | 0.11% | A | 1 |
| CYP1236 | 1 | 0.11% | A | 1 |
| CYP2334 | 2 | 0.23% | A | 2 |
| Species Name | P450 | Reference Cluster Information | ||
|---|---|---|---|---|
| Cluster Type | Most Similar Known Cluster | Similarity | ||
| Novosphingobium aromaticivorans | CYP153C1 | Terpene | Astaxanthin dideoxyglycoside | 75% |
| Mesorhizobium japonicum MAFF 303099 | CYP127A3v1 | Hserlactone | - | - |
| Agrobacterium fabrum | CYP206A1 | Terpene | - | - |
| Agrobacterium sp. H13-3 | CYP206A4 | Terpene | - | - |
| Agrobacterium tumefaciens | CYP206A4 | Terpene | - | - |
| Agrobacterium rhizogenes | CYP206A2 | Terpene | - | - |
| Rhizobium sp. IRBG74 | CYP206A3 | Terpene | - | - |
| Rhizobium sp. NT-26 | CYP107JE1 | NAGGN | - | - |
| Bradyrhizobium sp. S23321 | CYP199A26 | Hserlactone, t2pks | Colabomycin | 4% |
| Bradyrhizobium oligotrophicum | CYP108L2 | NRPS | - | - |
| Bosea vaviloviae | CYP1101A30 | T1PKS | S56-p1, NRPS | 3% |
| Beijerinckia indica | CYP173J1 | NRPS, T1PKS | - | - |
| Pseudorhodoplanes sinuspersici | CYP1104E2 | T1PKS | Sphingan polysaccharide, saccharide | 13% |
| Rhodobacter sphaeroides ATCC 17025 | CYP152C2 | hserlactone | Conglobatin, NRPS | 10% |
| Celeribacter indicus | CYP1101A27 | Ectoine | Ectoine, other | 100% |
| Hyphomonadaceae bacterium UKL13-1 | CYP1246A4 | Terpene | - | - |
| Sphingopyxis macrogoltabida 203 | CYP1302A1 | NRPS | - | - |
| Xanthobacter autotrophicus | CYP1138B1 | T1PKS | - | - |
| Sulfitobacter sp.AM1-D1 | CYP1326A2 | acyl_amino_acids | - | - |
| CYP195A21 | bacteriocin | - | - | |
| CYP2334A1 | NRPS-like | - | - | |
| Proteins Involved in Cholesterol Side-Chain Oxidation in Mycobacterium tuberculosis H37Rv | Homolog Proteins in Phenylobacterium zucineum | |||||
|---|---|---|---|---|---|---|
| Gene Name | Gene ID | Enzyme | Protein ID | % Identity | % Homology | Enzyme |
| fadD36 | Rv1193 | Acyl-CoA synthetase | PHZ_c1345 (fadD) | 29 | 41 | Long-chain acyl-CoA synthetase |
| pzu:PHZ_c1155 | 38 | 52 | Malonyl-CoA/methylmalonyl-CoA synthetase | |||
| fadD19 | Rv3515c | Probable fatty-acid-CoA ligase | pzu:PHZ_c2065 | 39 | 57 | Fatty-acid-CoA ligase |
| fadD3 | Rv3561 | Acyl-CoA synthetase (AMP forming) | pzu:PHZ_c1909 | 35 | 47 | Long-chain-fatty-acid—CoA ligase |
| fadD17 | Rv3506 | Possible fatty-acid-CoA ligase | pzu:PHZ_c0597 | 30 | 46 | Fatty-acyl-CoA synthase |
| fadD19 | Rv3515c | Probable fatty-acid-CoA ligase | pzu:PHZ_c2065 | 39 | 57 | Long-chain fatty acid:CoA ligase |
| fadD10 | Rv0099 | Fatty acid-CoA synthase | pzu:PHZ_c2512 | 29 | 43 | Long-chain-fatty-acid--CoA ligase |
| fadD9 | Rv2590 | Fatty acid-CoA synthase | pzu:PHZ_c2123 | 25 | 38 | Long-chain acyl-CoA synthetase |
| fadD18 | Rv3513c | Possible fatty-acid-CoA ligase | pzu:PHZ_c2065 | 55 | 73 | CoA-synthetase, long-chain fatty acid:CoA ligase |
| fadE22 | Rv3061c | Acyl-CoA dehydrogenase | pzu:PHZ_c2678 | 40 | 54 | Dehydrogenase family protein |
| fadE23 | Rv3140 | Acyl-CoA dehydrogenase | pzu:PHZ_c0890 | 33 | 51 | Acyl-CoA dehydrogenase |
| fadE24 | Rv3139 | Acyl-CoA dehydrogenase | pzu:PHZ_c2365 | 32 | 48 | Acyl-CoA dehydrogenase |
| fadE25 | Rv3274c | Acyl-CoA dehydrogenase FADE25 | pzu:PHZ_c1680 | 40 | 60 | Acyl-CoA dehydrogenase |
| fadE26 | Rv3504 | Probable acyl-CoA dehydrogenase | pzu:PHZ_c2336 | 33 | 50 | Acyl-CoA dehydrogenase |
| fadE27 | Rv3505 | Probable acyl-CoA dehydrogenase | pzu:PHZ_c2406 | 28 | 45 | Isovaleryl CoA dehydrogenase |
| fadE28 | Rv3544c | Probable acyl-CoA dehydrogenase | pzu:PHZ_c2498 | 26 | 43 | Acyl-CoA dehydrogenase |
| fadE29 | Rv3543c | Probable acyl-CoA dehydrogenase | pzu:PHZ_c2541 | 32 | 51 | Acyl-CoA dehydrogenase |
| fadE30 | Rv3560c | Probable acyl-CoA dehydrogenase | pzu:PHZ_c2541 | 41 | 57 | Acyl-CoA dehydrogenase |
| fadE31 | Rv3562 | Probable acyl-CoA dehydrogenase | pzu:PHZ_c2678 | 36 | 53 | Dehydrogenase family protein |
| fadE32 | Rv3563 | Probable acyl-CoA dehydrogenase | pzu:PHZ_c2498 | 32 | 45 | Acyl-CoA dehydrogenase |
| fadE33 | Rv3564 | Probable acyl-CoA dehydrogenase | pzu:PHZ_c2679 | 33 | 48 | Acyl-CoA dehydrogenase family protein |
| fadE34 | Rv3573c | Probable acyl-CoA dehydrogenase | pzu:PHZ_c2678 | 38 | 56 | Dehydrogenase family protein |
| fadE5 | Rv0244c | Acyl-CoA dehydrogenase | pzu:PHZ_c3354 | 36 | 51 | Acyl-CoA dehydrogenase |
| mbtN (fadE14) | Rv1346 | Acyl-CoA dehydrogenase | pzu:PHZ_c0388 | 30 | 48 | Acyl-CoA dehydrogenase |
| echA9 | Rv1071c | 3-Hydroxyisobutyryl-CoA hydrolase | pzu:PHZ_c1679 | 39 | 57 | Enoyl-CoA hydratase/isomerase family protein |
| echA19 | Rv3516 | Possible enoyl-CoA hydratase | pzu:PHZ_c2535 | 38 | 54 | Enoyl-CoA hydratase/carnithine racemase |
| echA20 | Rv3550 | Possible enoyl-CoA hydratase | pzu:PHZ_c3429 | 33 | 51 | Enoyl-CoA hydratase/isomerase family protein |
| fadB2 | Rv0468 | hydroxybutyryl-CoA dehydrogenase | pzu:PHZ_c3152 | 43 | 60 | 3-Hydroxyacyl-CoA dehydrogenase |
| fadB3 | Rv1715 | Hydroxybutyryl-CoA dehydrogenase | pzu:PHZ_c3152 | 35 | 49 | 3-Hydroxyacyl-CoA dehydrogenase |
| fadA5 | Rv3546 | Acetoacetyl-CoA thiolase | pzu:PHZ_c2504 | 40 | 60 | Acetyl-CoA C-acetyltransferase |
| hsd4A | Rv3502c | 17β-Hydroxysteroid dehydrogenase (17β-HSD) | pzu:PHZ_c2008 | 36 | 49 | Short-chain dehydrogenase/reductase SDR |
| ltp2 | Rv3540c | Probable ketoacyl-CoA thiolase | pzu:PHZ_c3245 | 36 | 49 | Acetyl-CoA C-acetyltransferase |
| P450 | Function | Reference |
|---|---|---|
| CYP101B1 | α-ionone, β-damascone, phenylcyclohexane and p-cymene hydroxylation | [53] |
| CYP101C1 | Ionone derivative hydroxylation | [54] |
| CYP101D1 | Terpenoid (camphor) hydroxylase | [55,56] |
| CYP101D2 | Camphor 5-exo hydroxylase | [57] |
| CYP108D1 | Aromatic hydrocarbon hydroxylase | [55,58] |
| CYP153C1, CYP153D1 | Alkane hydroxylase | [59] |
| CYP111A2 | Oxidizes linalool to 8-hydroxylinalool | [55] |
| CYP111B1 | β-Ionone hydroxylation | [60] |
| CYP112A2 | Oxidation of Rapamycin | [61] |
| CYP127A3 | Hydroxylation of testosterone | [62] |
| CYP195A2 | Degradation of 4-fluoro-, 4-chloro- and 4-methylsalicylic acid, and 3-chloro- and 3-methylsalicylic acid | [63] |
| CYP199A1 | Hydroxylation of 2-naphthoic acid | [64] |
| CYP199A2 | Hydroxylation of 2-naphthoic acid, para-substituted benzoic acids and involved in the degradation of ligninolic compounds | [64,65] |
| CYP199A4 | Catalyzes heteroatom dealkylations, sulfoxidation, and amide and cyclic hemiacetal formation | [66] |
| CYP200A1 | Hydroxylation of testosterone | [62] |
| CYP201A2 | Involved in the biodegradation of tributyl phosphate | [67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nzuza, N.; Padayachee, T.; Syed, P.R.; Kryś, J.D.; Chen, W.; Gront, D.; Nelson, D.R.; Syed, K. Ancient Bacterial Class Alphaproteobacteria Cytochrome P450 Monooxygenases Can Be Found in Other Bacterial Species. Int. J. Mol. Sci. 2021, 22, 5542. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115542
Nzuza N, Padayachee T, Syed PR, Kryś JD, Chen W, Gront D, Nelson DR, Syed K. Ancient Bacterial Class Alphaproteobacteria Cytochrome P450 Monooxygenases Can Be Found in Other Bacterial Species. International Journal of Molecular Sciences. 2021; 22(11):5542. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115542
Chicago/Turabian StyleNzuza, Nomfundo, Tiara Padayachee, Puleng Rosinah Syed, Justyna Dorota Kryś, Wanping Chen, Dominik Gront, David R. Nelson, and Khajamohiddin Syed. 2021. "Ancient Bacterial Class Alphaproteobacteria Cytochrome P450 Monooxygenases Can Be Found in Other Bacterial Species" International Journal of Molecular Sciences 22, no. 11: 5542. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115542