
Induced Pluripotent Stem Cell-Derived Conditioned Medium Promotes Endogenous Leukemia Inhibitory Factor to Attenuate Endotoxin-Induced Acute Lung Injury

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
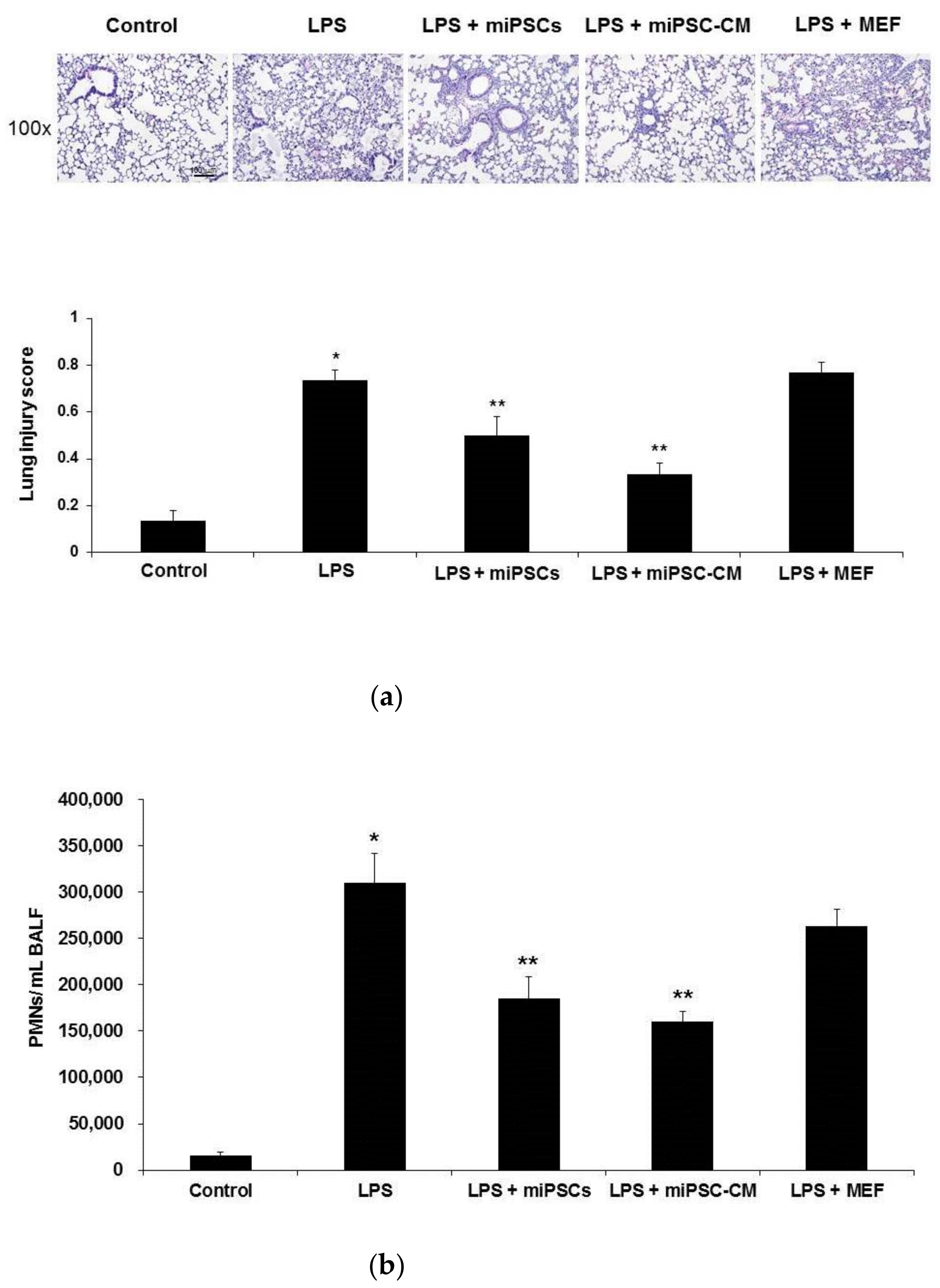
2.1. Effects of miPSC-CM on the Histopathology and Neutrophil Accumulation in the Lung in LPS-Induced ALI Mice
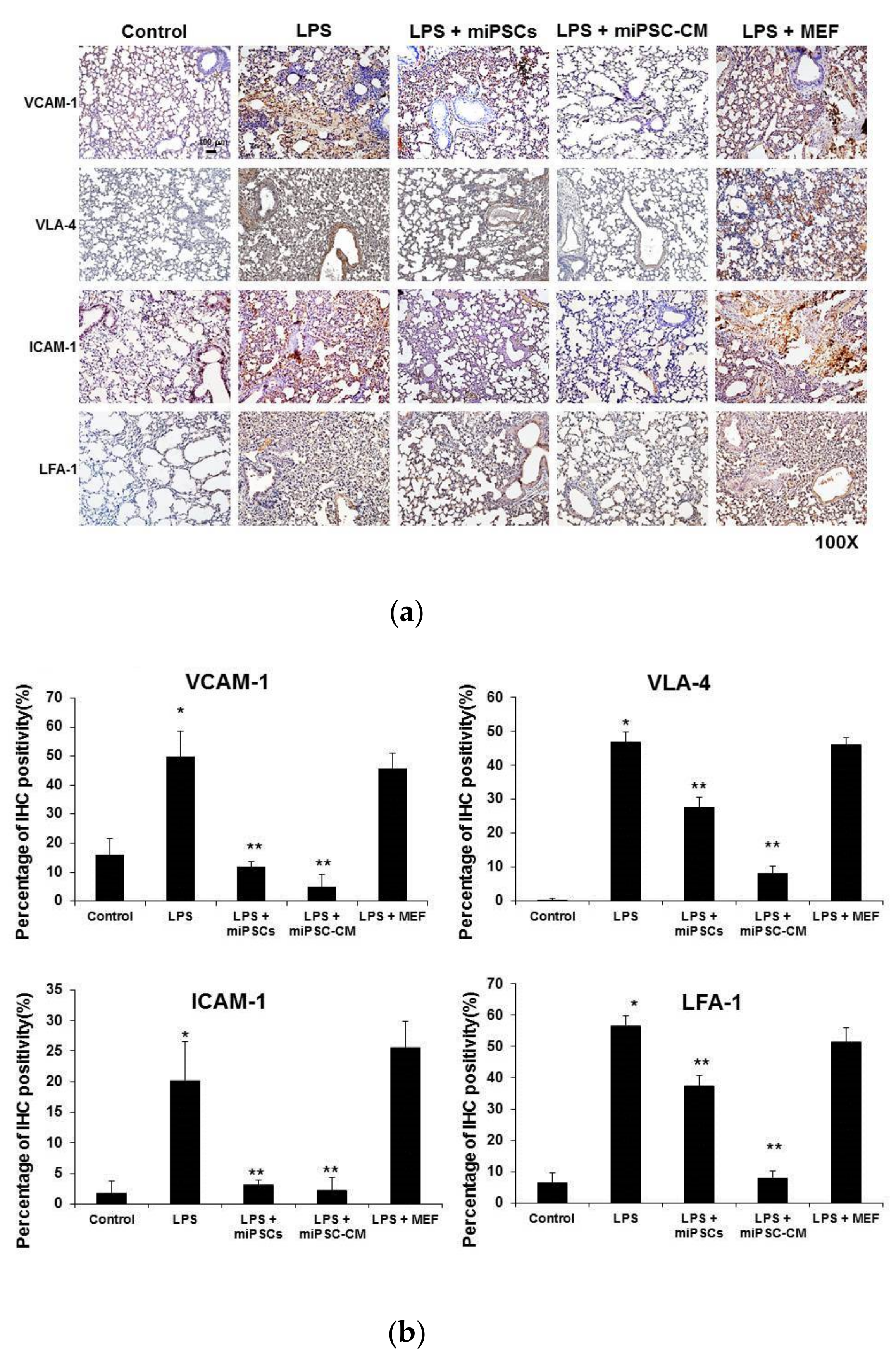
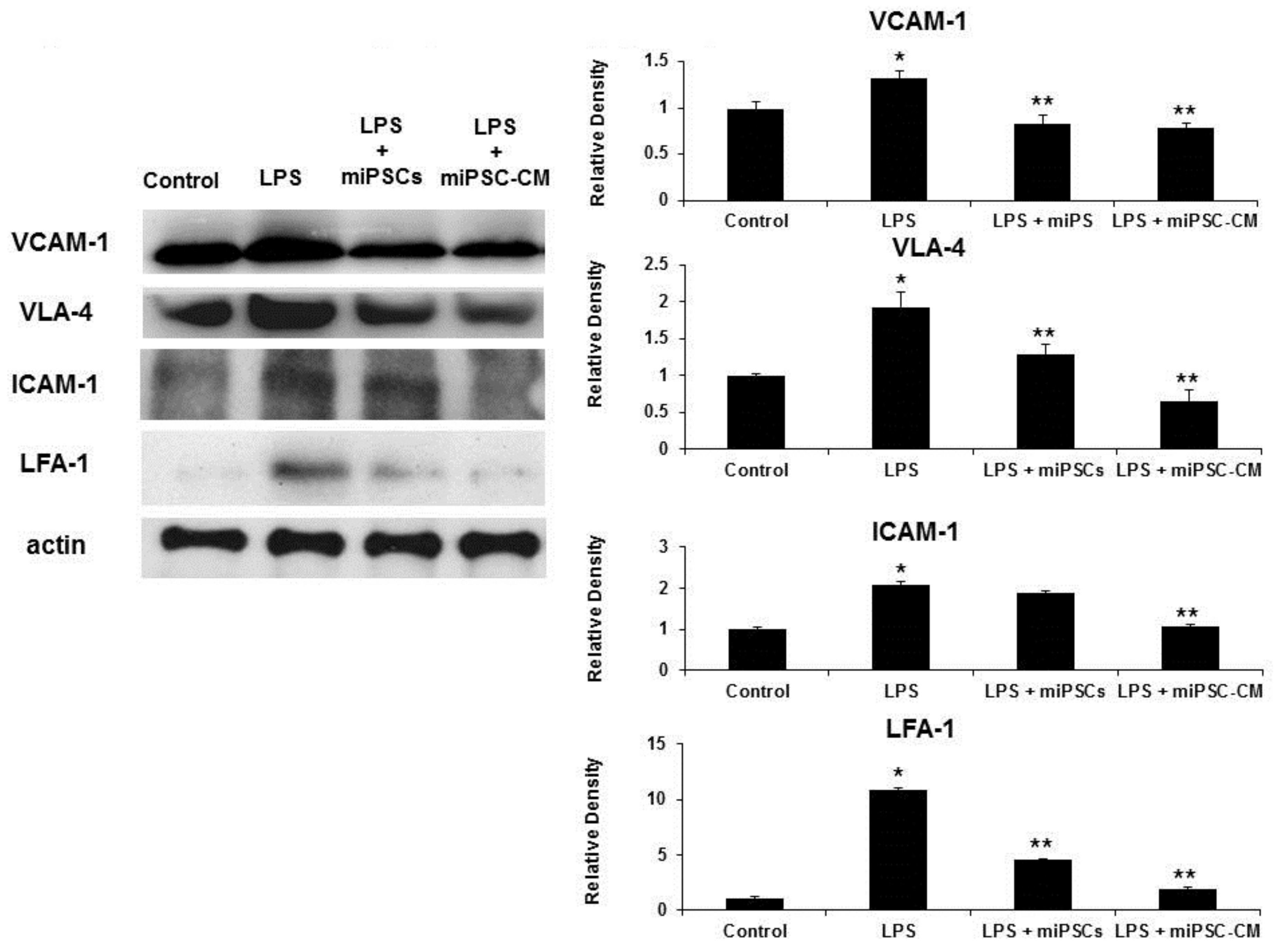
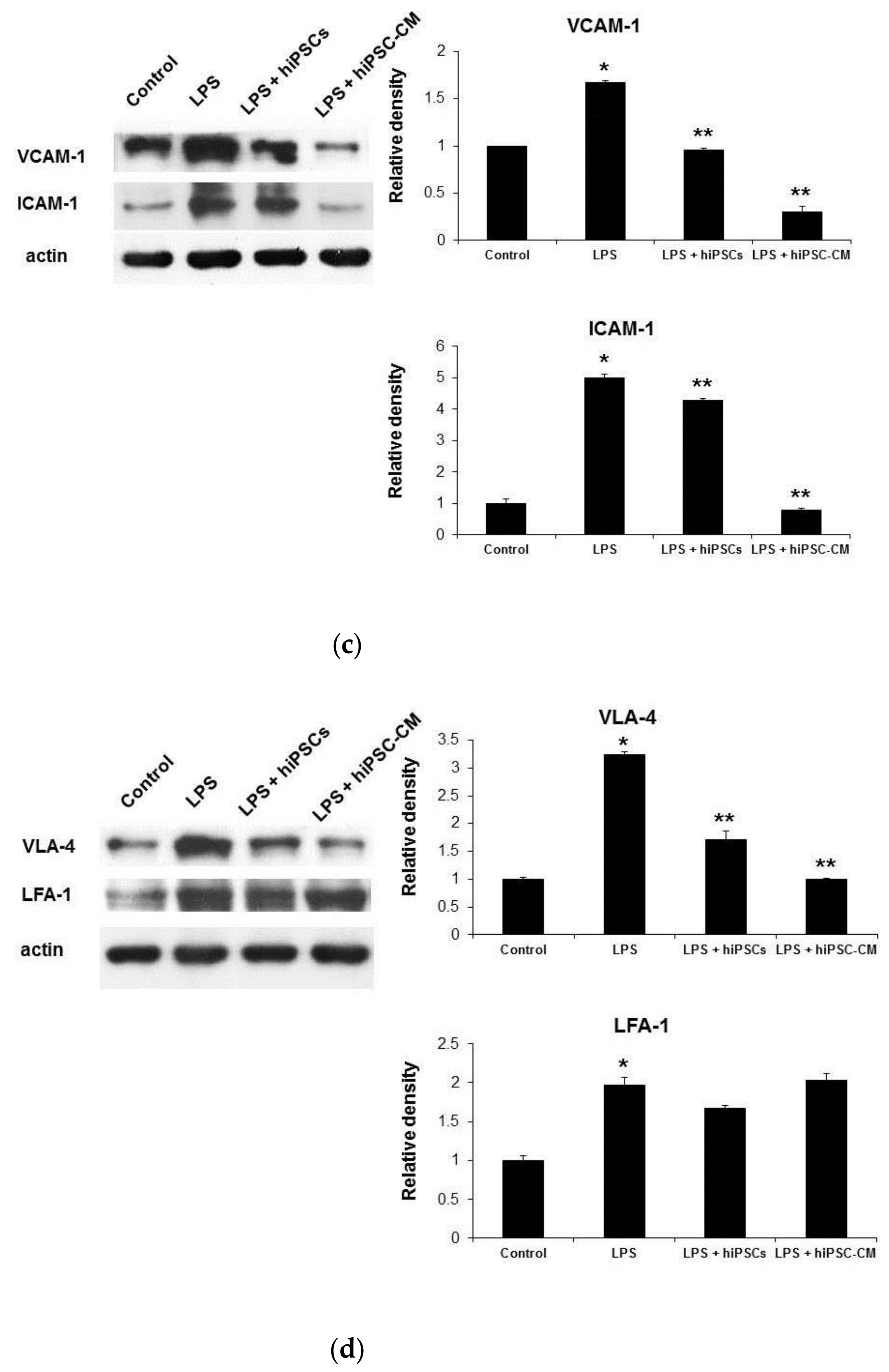
2.2. miPSC-CM Reduced ICAM-1/LFA-1 and VCAM-1/VLA-4 Expression in Lung Tissues Following LPS-Induced ALI
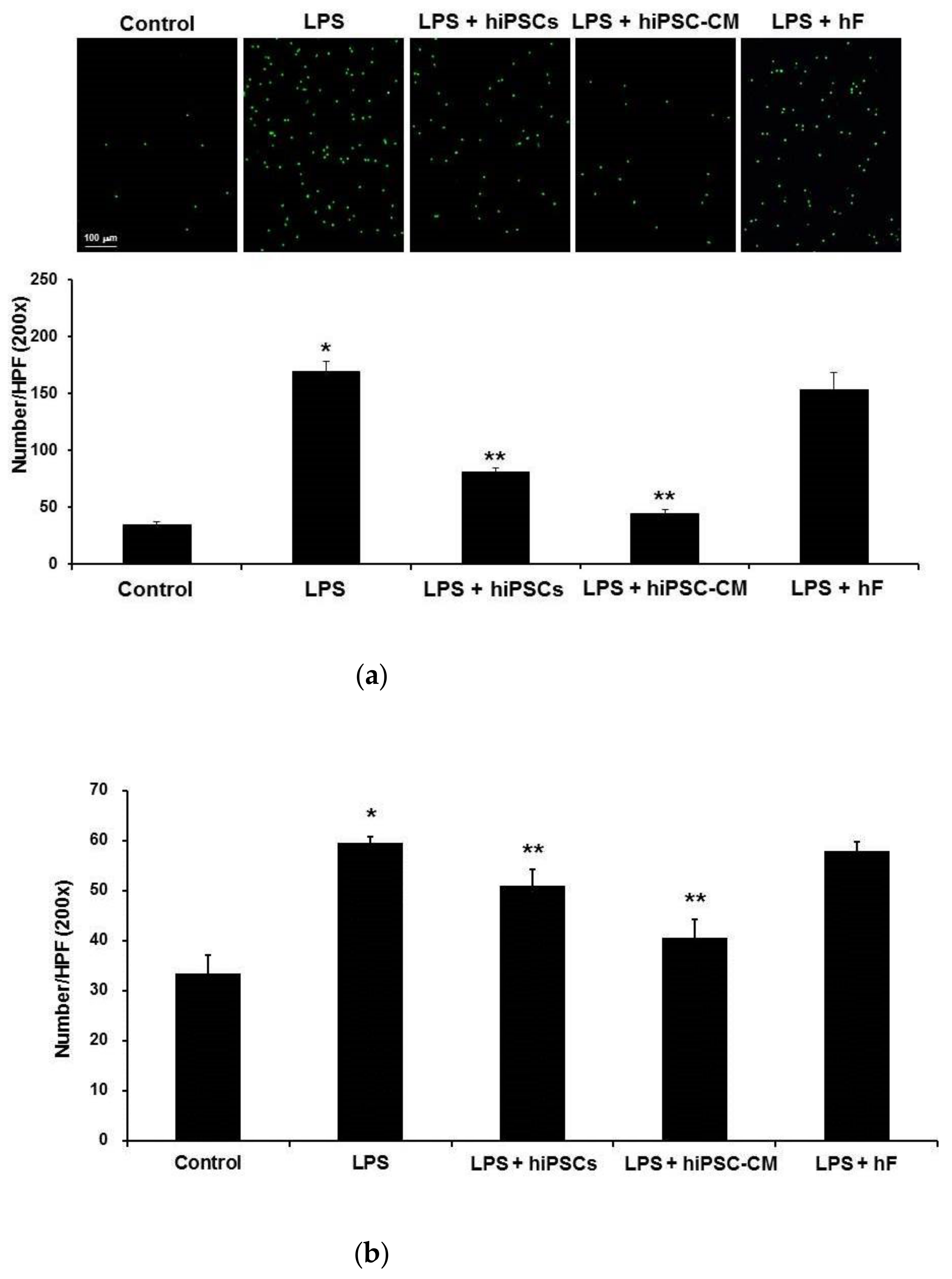
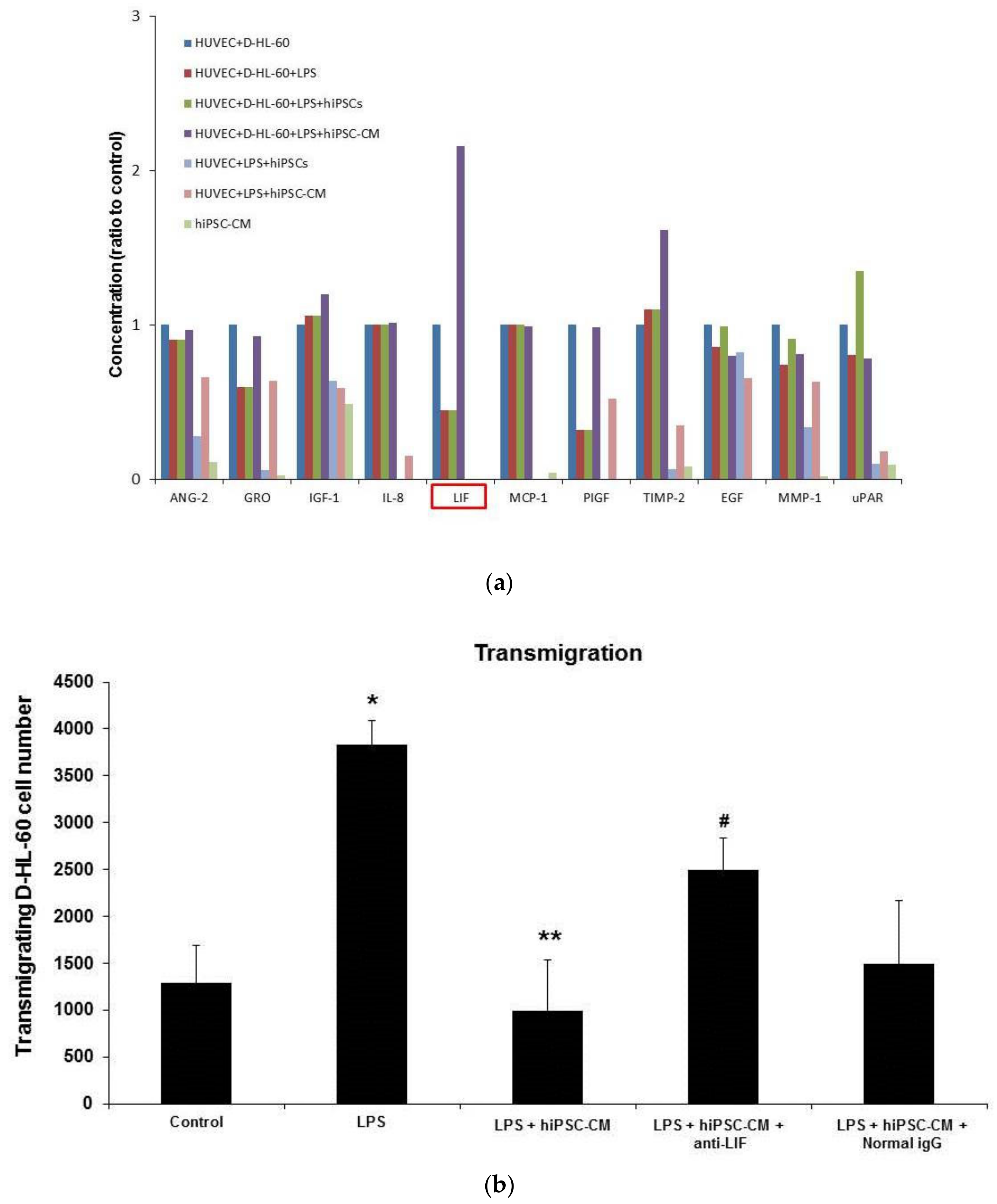
2.3. Effects of hiPSC-CM on Human Neutrophil–Endothelial Cell Interactions
2.4. hiPSC-CM Promoted Endogenous Leukemia Inhibitory Factor (LIF) Secretion to Decrease Neutrophils Transendothelial Migration (TEM) In Vitro
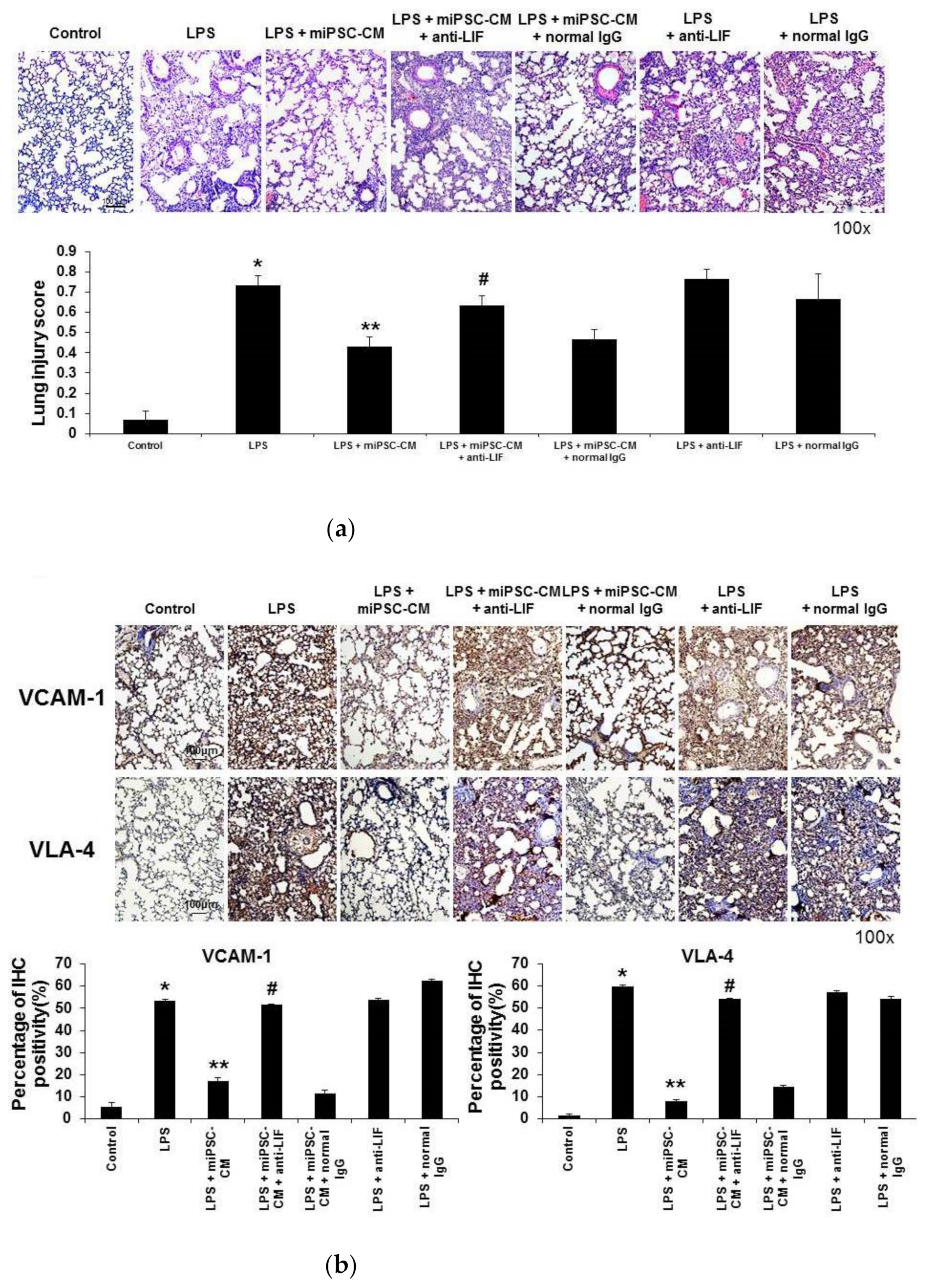
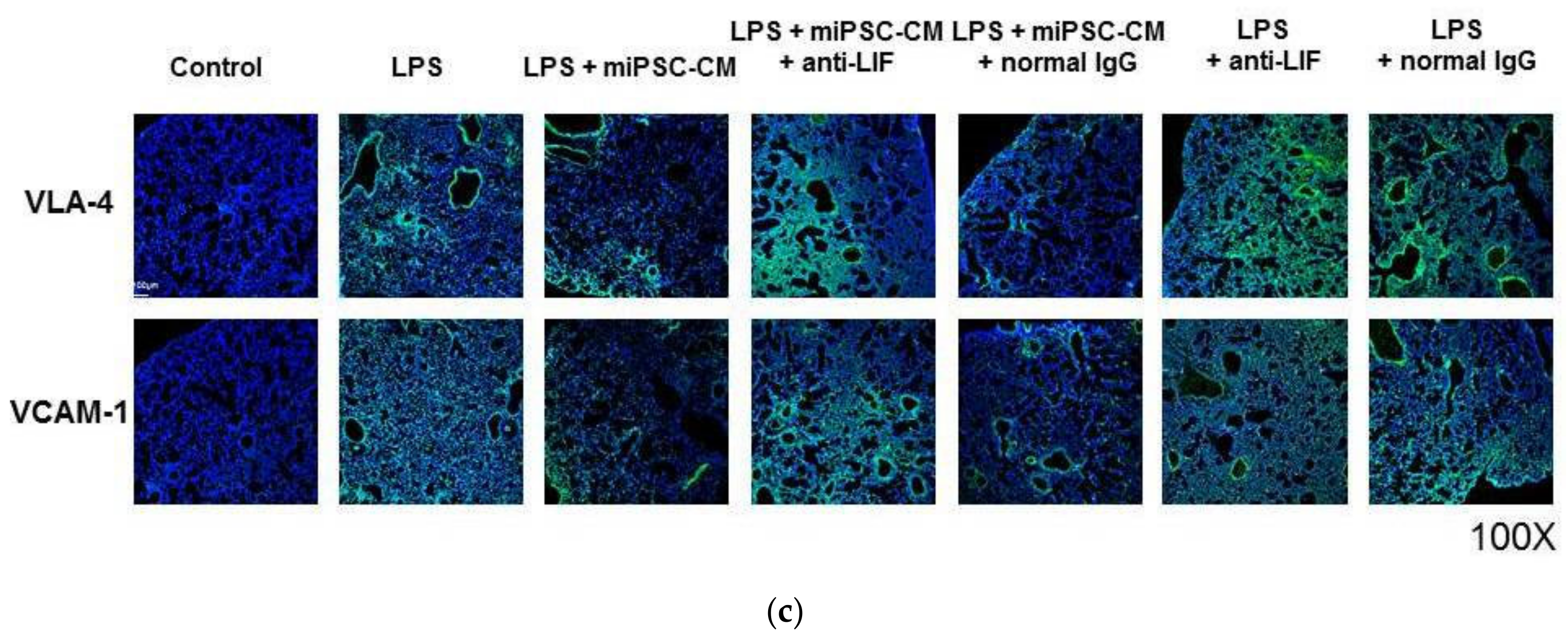
2.5. Anti-LIF Antibody Treatment Reversed the Effect of miPSC-CM on the Expression of VCAM-1 and VLA-4 and Pathological Changes in ALI Mice
3. Discussion
4. Methods
4.1. Experimental Animals
4.2. Generation and Culture of iPSCs
4.3. Experimental Design
4.4. Culture of Human Umbilical Vein Endothelial Cells (HUVECs)
4.5. Preparation of Dimethyl Sulfoxide (DMSO)-Induced HL-60 (D-HL-60) Cells
4.6. Flow Cytometry
4.7. Western Blotting
4.8. Histology and IHC
4.9. Immunofluorescence Stain
4.10. Scoring of Lung Injuries
4.11. In Vitro D-HL-60 Cells–Endothelial Cell Adhesion Assay
4.12. In Vitro D-HL-60 Cells Transendothelial Migration (TEM) Assay
4.13. Human Angiogenesis Factors Protein Array
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Butt, Y.; Kurdowska, A.; Allen, T.C. Acute Lung Injury: A Clinical and Molecular Review. Arch. Pathol. Lab. Med. 2016, 140, 345–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, V.Y.; Lin, C.S.; Hung, S.C.; Yang, K.Y. Mesenchymal Stem Cell-Conditioned Medium Induces Neutrophil Apoptosis Associated with Inhibition of the NF-kappaB Pathway in Endotoxin-Induced Acute Lung Injury. Int. J. Mol. Sci. 2019, 20, 2208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Okita, K.; Ichisaka, T.; Yamanaka, S. Generation of germline-competent induced pluripotent stem cells. Nature 2007, 448, 313–317. [Google Scholar] [CrossRef]
- Yang, K.Y.; Shih, H.C.; How, C.K.; Chen, C.Y.; Hsu, H.S.; Yang, C.W.; Lee, Y.C.; Perng, R.P.; Peng, C.H.; Li, H.Y.; et al. IV delivery of induced pluripotent stem cells attenuates endotoxin-induced acute lung injury in mice. Chest 2011, 140, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Su, V.Y.; Chiou, S.H.; Lin, C.S.; Chen, W.C.; Yu, W.K.; Chen, Y.W.; Chen, C.Y.; Yang, K.Y. Induced pluripotent stem cells reduce neutrophil chemotaxis via activating GRK2 in endotoxin-induced acute lung injury. Respirology 2017, 22, 1156–1164. [Google Scholar] [CrossRef]
- Su, V.Y.; Yang, K.Y.; Chiou, S.H.; Chen, N.J.; Mo, M.H.; Lin, C.S.; Wang, C.T. Induced Pluripotent Stem Cells Regulate Triggering Receptor Expressed on Myeloid Cell-1 Expression and the p38 Mitogen-Activated Protein Kinase Pathway in Endotoxin-Induced Acute Lung Injury. Stem Cells 2019, 37, 631–639. [Google Scholar] [CrossRef]
- Su, V.Y.; Chiou, S.H.; Lin, C.S.; Mo, M.H.; Yang, K.Y. iPS Cells Attenuate Endothelial Leakage in Acute Lung Injury via TIMP-1 to Reduce FAK Activity. Stem Cells 2019, 37, 1516–1527. [Google Scholar] [CrossRef]
- Reutershan, J.; Ley, K. Bench-to-bedside review: Acute respiratory distress syndrome—how neutrophils migrate into the lung. Crit. Care 2004, 8, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Nagase, T.; Ohga, E.; Sudo, E.; Katayama, H.; Uejima, Y.; Matsuse, T.; Fukuchi, Y. Intercellular adhesion molecule-1 mediates acid aspiration-induced lung injury. Am. J. Respir. Crit. Care Med. 1996, 154, 504–510. [Google Scholar] [CrossRef]
- Gando, S.; Kameue, T.; Matsuda, N.; Sawamura, A.; Hayakawa, M.; Kato, H. Systemic inflammation and disseminated intravascular coagulation in early stage of ALI and ARDS: Role of neutrophil and endothelial activation. Inflammation 2004, 28, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.A.; Issekutz, T.B.; Yagita, H.; Issekutz, A.C. The alpha 4 beta 1 (very late antigen (VLA)-4, CD49d/CD29) and alpha 5 beta 1 (VLA-5, CD49e/CD29) integrins mediate beta 2 (CD11/CD18) integrin-independent neutrophil recruitment to endotoxin-induced lung inflammation. J. Immunol. 2001, 166, 4644–4649. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Guo, Y.; Zhang, L.; More, S.; Weng, T.; Chintagari, N.R.; Huang, C.; Liang, Y.; Pushparaj, S.; Gou, D.; et al. A Critical Role for P2X7 Receptor-Induced VCAM-1 Shedding and Neutrophil Infiltration during Acute Lung Injury. J. Immunol. 2016, 197, 2828–2837. [Google Scholar] [CrossRef] [Green Version]
- Alon, R.; Kassner, P.D.; Carr, M.W.; Finger, E.B.; Hemler, M.E.; Springer, T.A. The integrin VLA-4 supports tethering and rolling in flow on VCAM-1. J. Cell Biol. 1995, 128, 1243–1253. [Google Scholar] [CrossRef]
- Berlin, C.; Bargatze, R.F.; Campbell, J.J.; von Andrian, U.H.; Szabo, M.C.; Hasslen, S.R.; Nelson, R.D.; Berg, E.L.; Erlandsen, S.L.; Butcher, E.C. Alpha 4 integrins mediate lymphocyte attachment and rolling under physiologic flow. Cell 1995, 80, 413–422. [Google Scholar] [CrossRef] [Green Version]
- Luscinskas, F.W.; Kansas, G.S.; Ding, H.; Pizcueta, P.; Schleiffenbaum, B.E.; Tedder, T.F.; Gimbrone, M.A., Jr. Monocyte rolling, arrest and spreading on IL-4-activated vascular endothelium under flow is mediated via sequential action of L-selectin, beta 1-integrins, and beta 2-integrins. J. Cell Biol. 1994, 125, 1417–1427. [Google Scholar] [CrossRef]
- Bochner, B.S.; Luscinskas, F.W.; Gimbrone, M.A., Jr.; Newman, W.; Sterbinsky, S.A.; Derse-Anthony, C.P.; Klunk, D.; Schleimer, R.P. Adhesion of human basophils, eosinophils, and neutrophils to interleukin 1-activated human vascular endothelial cells: Contributions of endothelial cell adhesion molecules. J. Exp. Med. 1991, 173, 1553–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhardt, P.H.; Elliott, J.F.; Kubes, P. Neutrophils can adhere via alpha4beta1-integrin under flow conditions. Blood 1997, 89, 3837–3846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhardt, P.H.; Kubes, P. Differential leukocyte recruitment from whole blood via endothelial adhesion molecules under shear conditions. Blood 1998, 92, 4691–4699. [Google Scholar] [CrossRef] [PubMed]
- Lomakina, E.B.; Waugh, R.E. Adhesion between human neutrophils and immobilized endothelial ligand vascular cell adhesion molecule 1: Divalent ion effects. Biophys. J. 2009, 96, 276–284. [Google Scholar] [CrossRef] [Green Version]
- Alexander, H.R.; Billingsley, K.G.; Block, M.I.; Fraker, D.L. D-factor/leukaemia inhibitory factor: Evidence for its role as a mediator in acute and chronic inflammatory disease. Cytokine 1994, 6, 589–596. [Google Scholar] [CrossRef]
- Gough, N.M.; Williams, R.L.; Hilton, D.J.; Pease, S.; Willson, T.A.; Stahl, J.; Gearing, D.P.; Nicola, N.A.; Metcalf, D. LIF: A molecule with divergent actions on myeloid leukaemic cells and embryonic stem cells. Reprod. Fertil. Dev. 1989, 1, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Ulich, T.R.; Fann, M.J.; Patterson, P.H.; Williams, J.H.; Samal, B.; Del Castillo, J.; Yin, S.; Guo, K.; Remick, D.G. Intratracheal injection of LPS and cytokines. V. LPS induces expression of LIF and LIF inhibits acute inflammation. Am. J. Physiol. 1994, 267, L442–L446. [Google Scholar] [CrossRef]
- Waring, P.; Wycherley, K.; Cary, D.; Nicola, N.; Metcalf, D. Leukemia inhibitory factor levels are elevated in septic shock and various inflammatory body fluids. J. Clin. Invest. 1992, 90, 2031–2037. [Google Scholar] [CrossRef] [Green Version]
- Waring, P.M.; Waring, L.J.; Metcalf, D. Circulating leukemia inhibitory factor levels correlate with disease severity in meningococcemia. J. Infect. Dis. 1994, 170, 1224–1228. [Google Scholar] [CrossRef]
- Waring, P.M.; Waring, L.J.; Billington, T.; Metcalf, D. Leukemia inhibitory factor protects against experimental lethal Escherichia coli septic shock in mice. Proc. Natl. Acad. Sci. USA 1995, 92, 1337–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, M.A.; Schnyder-Candrian, S.; Schnyder, B.; Quesniaux, V.; Poli, V.; Stewart, C.L.; Ryffel, B. Endogenous leukemia inhibitory factor attenuates endotoxin response. Lab. Investig. J. Tech. Methods Pathol. 2005, 85, 276–284. [Google Scholar] [CrossRef]
- Aghajanian, H.; Cho, Y.K.; Manderfield, L.J.; Herling, M.R.; Gupta, M.; Ho, V.C.; Li, L.; Degenhardt, K.; Aharonov, A.; Eldad Tzahor, E.; et al. Coronary vasculature patterning requires a novel endothelial ErbB2 holoreceptor. Nat. Commun. 2016, 7, 12038. [Google Scholar] [CrossRef] [Green Version]
- Aghajanian, H.; Choi, C.; Ho, V.C.; Gupta, M.; Singh, M.K.; Epstein, J.A. Semaphorin 3d and Semaphorin 3e Direct Endothelial Motility through Distinct Molecular Signaling Pathways. J. Biol. Chem. 2014, 289, 17971–17979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gee, D.J.; Wright, L.K.; Zimmermann, J.; Cole, K.; Soule, K.; Ubowski, M. Dimethylsulfoxide exposure modulates HL-60 cell rolling interactions. Biosci. Rep. 2012, 32, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Trayner, I.D.; Bustorff, T.; Etches, A.E.; Mufti, G.J.; Foss, Y.; Farzaneh, F. Changes in antigen expression on differentiating HL60 cells treated with dimethylsulphoxide, all-trans retinoic acid, alpha1,25-dihydroxyvitamin D3 or 12-O-tetradecanoyl phorbol-13-acetate. Leuk. Res. 1998, 22, 537–547. [Google Scholar] [CrossRef]
- Joseph, C.; David, G.; Iris, G.; Sandra, J.; Ronald, V.M. Endotoxin-Induced Endothelial Cell Proinflammatory Phenotypic Differentiation Requires Stress Fiber Polymerization. Shock 2003, 19, 433–439. [Google Scholar]
- Galdiero, M.; Folgore, A.; Molitierno, M.; Greco, R. Porins and ipopolysaccharide (LPS) from Salmonella typhimurium induce leucocyte transmigration through human endothelial cells in vitro. Clin. Exp. Immunol. 1999, 116, 453–461. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, V.Y.-F.; Chiou, S.-H.; Chen, W.-C.; Yu, W.-K.; Wu, H.-H.; Chen, H.; Yang, K.-Y. Induced Pluripotent Stem Cell-Derived Conditioned Medium Promotes Endogenous Leukemia Inhibitory Factor to Attenuate Endotoxin-Induced Acute Lung Injury. Int. J. Mol. Sci. 2021, 22, 5554. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115554
Su VY-F, Chiou S-H, Chen W-C, Yu W-K, Wu H-H, Chen H, Yang K-Y. Induced Pluripotent Stem Cell-Derived Conditioned Medium Promotes Endogenous Leukemia Inhibitory Factor to Attenuate Endotoxin-Induced Acute Lung Injury. International Journal of Molecular Sciences. 2021; 22(11):5554. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115554
Chicago/Turabian StyleSu, Vincent Yi-Fong, Shih-Hwa Chiou, Wei-Chih Chen, Wen-Kuang Yu, Huai-Hsuan Wu, Hao Chen, and Kuang-Yao Yang. 2021. "Induced Pluripotent Stem Cell-Derived Conditioned Medium Promotes Endogenous Leukemia Inhibitory Factor to Attenuate Endotoxin-Induced Acute Lung Injury" International Journal of Molecular Sciences 22, no. 11: 5554. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115554