
Dynamics of Bacterial Community Structure in the Rhizosphere and Root Nodule of Soybean: Impacts of Growth Stages and Varieties

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Field Construction and Soil Sample Preparation
2.3. Root Nodule Preparation
2.4. Community-Level Physiological Profiling (CLPP) Analysis
2.5. PCR Amplification and Illumina Sequencing
2.6. Bioinformatics Analyses
2.7. Statistical Analysis
3. Results
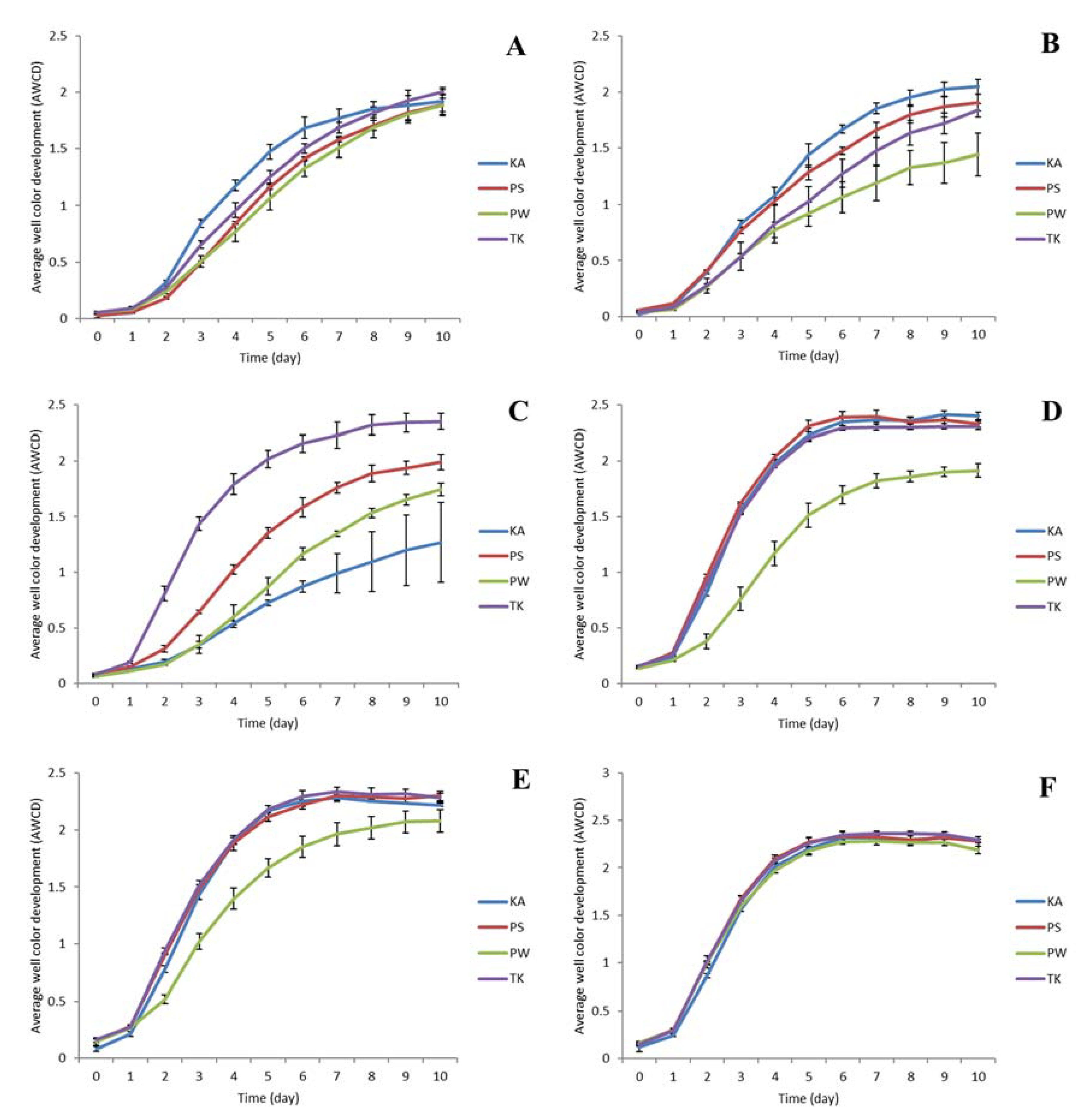
3.1. BioLog Substrate Utilization Assay
3.2. Alpha Diversity Analysis
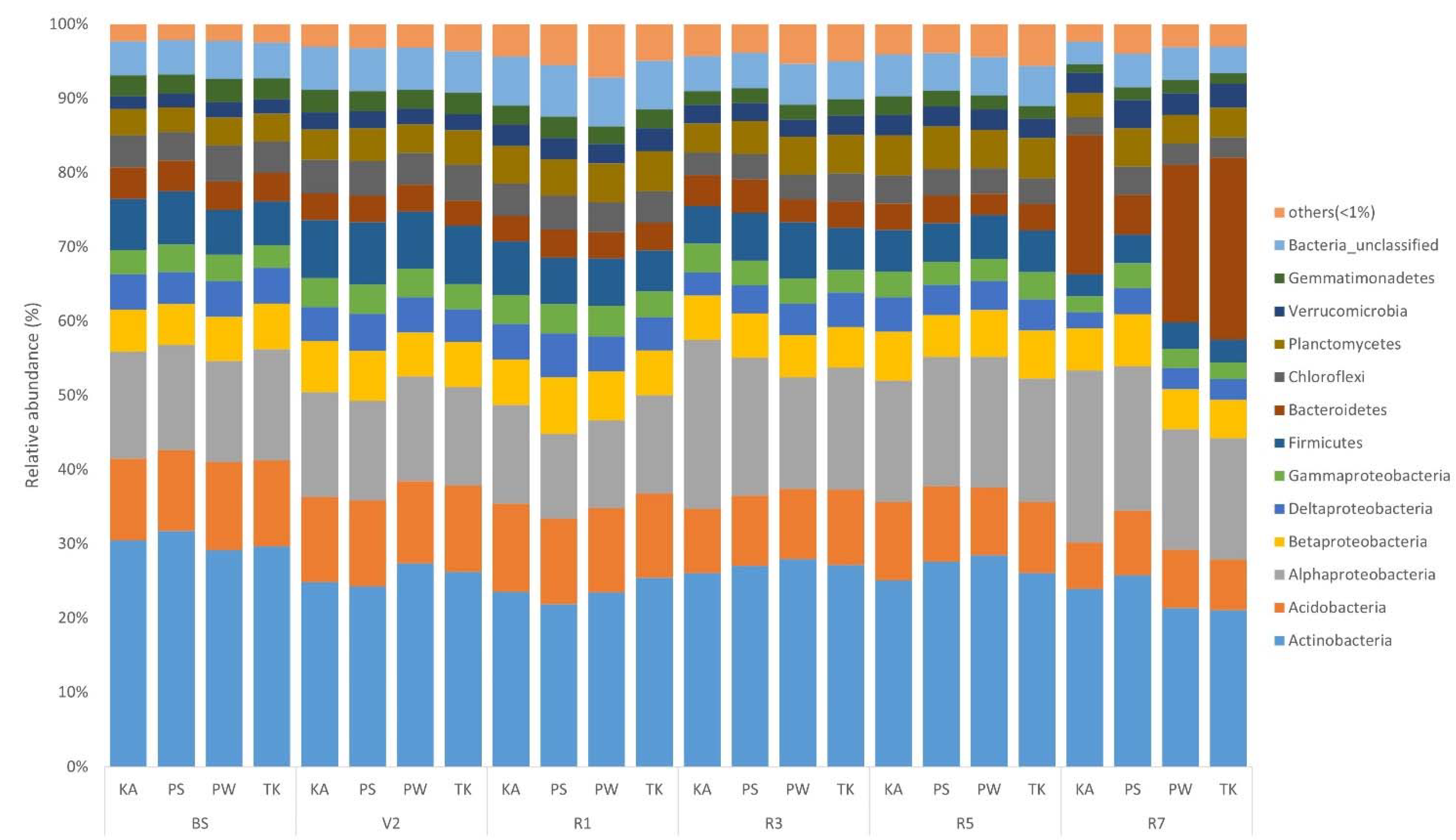
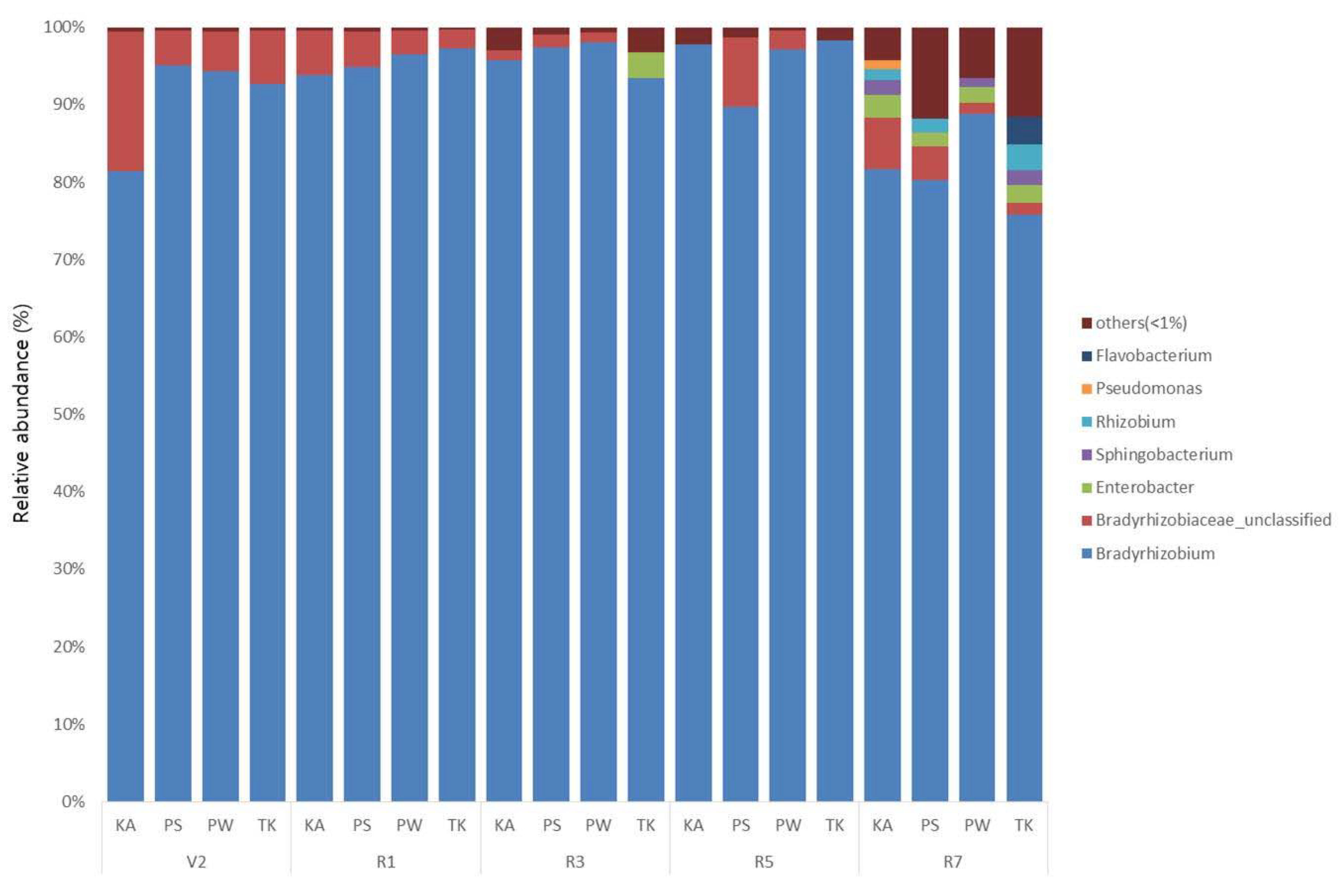
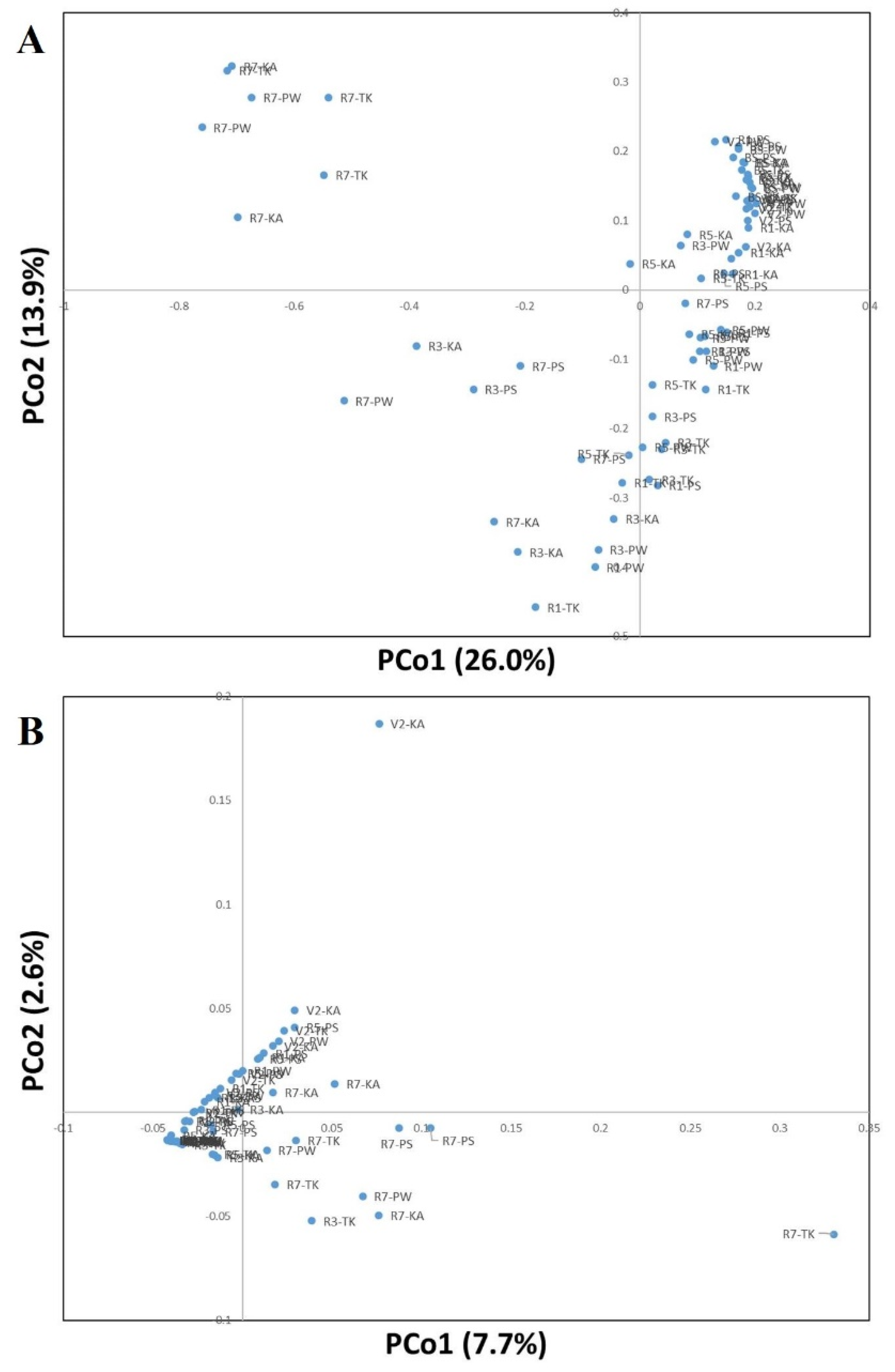
3.3. Phylogenetic Structure Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sugiyama, A. The soybean rhizosphere: Metabolites, microbes, and beyond—A review. J. Adv. Res. 2019, 19, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.-J.; Chen, P.-Y.; Liu, Z.-X.; Li, Y.-H.; Guan, R.-X.; Wang, L.-H.; Chang, R.-Z. The worldwide utilization of the Chinese soybean germplasm collection. Plant Genet. Resour. 2011, 9, 109–122. [Google Scholar] [CrossRef]
- USDA Foreign Agricultural Service. Available online: http://www.fas.usda.gov (accessed on 23 September 2020).
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Fact. 2014, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moe, L.A. Amino acids in the rhizosphere: From plants to microbes. Am. J. Bot. 2013, 100, 1692–1705. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, V.; Selvaraj, G.; Bais, H.P. Functional soil microbiome: Belowground solutions to an aboveground problem. Plant Physiol. 2014, 166, 689–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De-La-Peña, C.; Loyola-Vargas, V.M. Biotic interactions in the rhizosphere: A diverse cooperative enterprise for plant productivity. Plant Physiol. 2014, 166, 701–719. [Google Scholar] [CrossRef] [Green Version]
- Tkacz, A.; Poole, P. Role of root microbiota in plant productivity. J. Exp. Bot. 2015, 66, 2167–2175. [Google Scholar] [CrossRef] [Green Version]
- Pinton, R.; Varanini, Z.; Nannipieri, P. The Rhizosphere: Biochemistry and Organic Substances at the Soil-Plant Interface, 2nd ed.; Marcel Dekker: New York, NY, USA, 2001. [Google Scholar]
- Morgan, J.A.W.; Bending, G.D.; White, P.J. Biological costs and benefits to plant–microbe interactions in the rhizosphere. J. Exp. Bot. 2005, 56, 1729–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.-F.; Chaparro, J.M.; Reardon, K.; Zhang, R.; Shen, Q.; Vivanco, J.M. Rhizosphere interactions: Root exudates, microbes, and microbial communities. Botany 2014, 92, 267–275. [Google Scholar] [CrossRef]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S.; Tremblay, J.; Engelbrektson, A.; Kunin, V.; Del Rio, T.G.; et al. Defining the core Arabidopsis thaliana root microbiome. Nat. Cell Biol. 2012, 488, 86–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlaeppi, K.; Dombrowski, N.; Oter, R.G.; van Themaat, E.V.L.; Schulze-Lefert, P. Quantitative divergence of the bacterial root microbiota in Arabidopsis thaliana relatives. Proc. Natl. Acad. Sci. USA 2014, 111, 585–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakker, P.A.; Pieterse, C.M.; De Jonge, R.; Berendsen, R.L. The soil-borne legacy. Cell 2018, 172, 1178–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakker, P.A.; Doornbos, R.F.; Zamioudis, C.; Berendsen, R.L.; Pieterse, C.M. Induced systemic resistance and the rhizosphere microbiome. Plant Pathol. J. 2013, 29, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Mendes, L.W.; Braga, L.P.P.; Navarrete, A.A.; de Souza, D.G.; Silva, G.G.Z.; Tsai, S.M. Using metagenomics to connect microbial community biodiversity and functions. Curr. Issues Mol. Biol. 2017, 24, 103–118. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, G.; Teixeira, P.J.; Paredes, S.H.; Law, T.F.; De Lorenzo, L.; Feltcher, M.E.; Finkel, O.M.; Breakfield, N.W.; Mieczkowski, P.; Jones, P.M.C.D.; et al. Root microbiota drive direct integration of phosphate stress and immunity. Nat. Cell Biol. 2017, 543, 513–518. [Google Scholar] [CrossRef]
- Mendes, L.W.; Kuramae, E.E.; Navarrete, A.A.; Van Veen, J.A.; Tsai, S.M. Taxonomical and functional microbial community selection in soybean rhizosphere. ISME J. 2014, 8, 1577–1587. [Google Scholar] [CrossRef]
- Fitzpatrick, C.R.; Copeland, J.; Wang, P.W.; Guttman, D.S.; Kotanen, P.M.; Johnson, M.T.J. Assembly and ecological function of the root microbiome across angiosperm plant species. Proc. Natl. Acad. Sci. USA 2018, 115, E1157–E1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiyama, A.; Ueda, Y.; Zushi, T.; Takase, H.; Yazaki, K. Changes in the bacterial community of soybean rhizospheres during growth in the field. PLoS ONE 2014, 9, e100709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badri, D.V.; Weir, T.L.; van der Lelie, D.; Vivanco, J.M. Rhizosphere chemical dialogues: Plant–microbe interactions. Curr. Opin. Biotechnol. 2009, 20, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Badri, D.V.; Chaparro, J.M.; Zhang, R.; Shen, Q.; Vivanco, J.M. Application of natural blends of phytochemicals derived from the root exudates of Arabidopsis to the soil reveal that phenolic-related compounds predominantly modulate the soil microbiome. J. Biol. Chem. 2013, 288, 4502–4512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaparro, J.M.; Badri, D.V.; Bakker, M.G.; Sugiyama, A.; Manter, D.K.; Vivanco, J.M. Root exudation of phytochemicals in arabidopsis follows specific patterns that are developmentally programmed and correlate with soil microbial functions. PLoS ONE 2013, 8, e55731. [Google Scholar] [CrossRef]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Ke, M.; Lavoie, M.; Jin, Y.; Fan, X.; Zhang, Z.; Fu, Z.; Sun, L.; Gillings, M.; Peñuelas, J.; et al. Rhizosphere microorganisms can influence the timing of plant flowering. Microbiome 2018, 6, 1–12. [Google Scholar] [CrossRef]
- Saleem, M.; Law, A.D.; Sahib, M.R.; Pervaiz, Z.H.; Zhang, Q. Impact of root system architecture on rhizosphere and root microbiome. Rhizosphere 2018, 6, 47–51. [Google Scholar] [CrossRef]
- Garbeva, P.; Van Elsas, J.D.; Van Veen, J.A. Rhizosphere microbial community and its response to plant species and soil history. Plant. Soil 2007, 302, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Schlemper, T.R.; Leite, M.F.A.; Lucheta, A.R.; Shimels, M.; Bouwmeester, H.J.; Van Veen, J.A.; Kuramae, E.E. Rhizobacterial community structure differences among sorghum cultivars in different growth stages and soils. FEMS Microbiol. Ecol. 2017, 93, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Q.; Wang, F.; Zhang, J.; Chen, Y.; Zhang, C.; Liu, G.; Zhang, H.; Ma, C.; Zhang, J. The variation in the rhizosphere microbiome of cotton with soil type, genotype and developmental stage. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; Van Themaat, E.V.L.; Ahmadinejad, N.; Assenza, F.; Rauf, P.; Huettel, B.; Reinhardt, R.; Schmelzer, E.; et al. Revealing structure and assembly cues for Arabidopsis root-inhabiting bacterial microbiota. Nat. Cell Biol. 2012, 488, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aira, M.; Gómez-Brandón, M.; Lazcano, C.; Bååth, E.; Domínguez, J. Plant genotype strongly modifies the structure and growth of maize rhizosphere microbial communities. Soil Biol. Biochem. 2010, 42, 2276–2281. [Google Scholar] [CrossRef]
- Liang, J.; Sun, S.; Ji, J.; Wu, H.; Meng, F.; Zhang, M.; Zheng, X.; Wu, C.; Zhang, Z. Comparison of the rhizosphere bacterial communities of zigongdongdou soybean and a high-methionine transgenic line of this cultivar. PLoS ONE 2014, 9, e103343. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.M.; Hoilett, N.; Lorenz, N.; Dick, R.P.; Liles, M.; Ramsier, C.; Kloepper, J.W. Glyphosate effects on soil rhizosphere-associated bacterial communities. Sci. Total. Environ. 2016, 543, 155–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okutani, F.; Hamamoto, S.; Aoki, Y.; Nakayasu, M.; Nihei, N.; Nishimura, T.; Yazaki, K.; Sugiyama, A. Rhizosphere modelling reveals spatiotemporal distribution of daidzein shaping soybean rhizosphere bacterial community. Plant. Cell Environ. 2020, 43, 1036–1046. [Google Scholar] [CrossRef]
- Albrecht, S.L.; Maier, R.J.; Hanus, F.J.; Russell, S.A.; Emerich, D.W.; Evans, H.J. Hydrogenase in Rhizobium japonicum increases nitrogen fixation by nodulated soybeans. Science 1979, 203, 1255–1257. [Google Scholar] [CrossRef]
- Williams, L.E.; Phillips, D.A. Increased soybean productivity with a Rhizobium japonicum mutant 1. Crop. Sci. 1983, 23, 246–250. [Google Scholar] [CrossRef]
- Xiao, X.; Chen, W.; Zong, L.; Yang, J.; Jiao, S.; Lin, Y.; Wang, E.; Wei, G. Two cultivated legume plants reveal the enrichment process of the microbiome in the rhizocompartments. Mol. Ecol. 2017, 26, 1641–1651. [Google Scholar] [CrossRef] [PubMed]
- Wigley, K.; Moot, D.; Wakelin, S.A.; Laugraud, A.; Blond, C.; Seth, K.; Ridgway, H. Diverse bacterial taxa inhabit root nodules of lucerne (Medicago sativa L.) in New Zealand pastoral soils. Plant. Soil 2017, 420, 253–262. [Google Scholar] [CrossRef]
- Martínez-Hidalgo, P.; Hirsch, A.M. The nodule microbiome: N2-fixing Rhizobia do not live alone. Phytobiomes J. 2017, 1, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Somasegaran, P.; Hoben, H.J. Collecting nodules and isolating rhizobia. In Handbook for Rhizobia; Springer: Yew York, NY, USA, 1994; pp. 7–23. [Google Scholar]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, M.; Tan, F.; Zhu, H.; Cheng, K.; Wu, X.; Wang, J.; Zhao, K.; Tang, X. Impact of Bt-transgenic rice (SHK601) on soil ecosystems in the rhizosphere during crop development. Plant. Soil Environ. 2012, 58, 217–223. [Google Scholar] [CrossRef] [Green Version]
- Uroz, S.; Buée, M.; Murat, C.; Frey-Klett, P.; Martin, F. Pyrosequencing reveals a contrasted bacterial diversity between oak rhizosphere and surrounding soil. Environ. Microbiol. Rep. 2010, 2, 281–288. [Google Scholar] [CrossRef]
- Suleiman, M.K.; Dixon, K.; Commander, L.; Nevill, P.; Quoreshi, A.M.; Bhat, N.R.; Manuvel, A.J.; Sivadasan, M.T. Assessment of the diversity of fungal community composition associated with Vachellia pachyceras and its rhizosphere soil from Kuwait desert. Front. Microbiol. 2019, 10, 63. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wu, B.; Jiang, K.; Wei, M.; Wang, S. Effects of different concentrations and types of Cu and Pb on soil N-fixing bacterial communities in the wheat rhizosphere. Appl. Soil Ecol. 2019, 144, 51–59. [Google Scholar] [CrossRef]
- Kumar, A.; Maurya, B.; Raghuwanshi, R. Isolation and characterization of PGPR and their effect on growth, yield and nutrient content in wheat (Triticum aestivum L.). Biocatal. Agric. Biotechnol. 2014, 3, 121–128. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Cherdyntseva, T.A.; Botina, S.G.; Netrusov, A. Bacteria associated with orchid roots and microbial production of auxin. Microbiol. Res. 2007, 162, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Palaniyandi, S.A.; Yang, S.H.; Damodharan, K.; Suh, J.-W. Genetic and functional characterization of culturable plant-beneficial actinobacteria associated with yam rhizosphere. J. Basic Microbiol. 2013, 53, 985–995. [Google Scholar] [CrossRef]
- Miller, E.S.; Woese, C.R.; Brenner, S. Description of the erythromycin-producing bacterium Arthrobacter sp. strain NRRL B-3381 as Aeromicrobium erythreum gen. nov., sp. nov. Int. J. Syst. Bacteriol. 1991, 41, 363–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.-W.; Breakspear, A.; Guan, D.; Cerri, M.R.; Jackson, K.; Jiang, S.; Robson, F.; Radhakrishnan, G.V.; Roy, S.; Bone, C.; et al. NIN acts as a network hub controlling a growth module required for rhizobial infection. Plant. Physiol. 2019, 179, 1704–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupwayi, N.Z.; Clayton, G.W.; Hanson, K.G.; Rice, W.A.; Biederbeck, V.O. Endophytic rhizobia in barley, wheat and canola roots. Can. J. Plant. Sci. 2004, 84, 37–45. [Google Scholar] [CrossRef]
- Sharma, P.K.; Sarita, S.; Prell, J. Isolation and characterization of an endophytic bacterium related to Rhizobium/Agrobacterium from wheat (Triticum aestivum L.) roots. Curr. Sci. 2005, 89, 608–613. [Google Scholar]
- Soltani, A.-A.; Khavazi, K.; Asadi-Rahmani, H.; Omidvari, M.; Dahaji, P.A.; Mirhoseyni, H. Plant growth promoting characteristics in some Flavobacterium spp. isolated from soils of Iran. J. Agric. Sci. 2010, 2, p106. [Google Scholar] [CrossRef] [Green Version]
- Sharaf, H.; Rodrigues, R.R.; Moon, J.; Zhang, B.; Mills, K.; Williams, M.A. Unprecedented bacterial community richness in soybean nodules vary with cultivar and water status. Microbiome 2019, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, B.E.; Vest, G. Effects of Rhizobium japonicum strains on soybean yields. Crop. Sci. 1970, 10, 19–21. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant. Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patten, C.L.; Glick, B.R. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl. Environ. Microbiol. 2002, 68, 3795–3801. [Google Scholar] [CrossRef] [Green Version]
- Cornelis, P.; Matthijs, S. Diversity of siderophore-mediated iron uptake systems in fluorescent pseudomonads: Not only pyoverdines. Environ. Microbiol. 2002, 4, 787–798. [Google Scholar] [CrossRef] [Green Version]
- Compant, S.; Reiter, B.; Sessitsch, A.; Nowak, J.; Clément, C.; Barka, E.A. Endophytic colonization of Vitis vinifera L. by plant growth-promoting bacterium Burkholderia sp. strain PsJN. Appl. Environ. Microbiol. 2005, 71, 1685–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Kuklinsky-Sobral, J.; Araújo, W.L.; Mendes, R.; Geraldi, I.O.; Pizzirani-Kleiner, A.A.; Azevedo, J.L. Isolation and characterization of soybean-associated bacteria and their potential for plant growth promotion. Environ. Microbiol. 2004, 6, 1244–1251. [Google Scholar] [CrossRef]
- Polonenko, D.R.; Scher, F.M.; Kloepper, J.W.; Singleton, C.A.; Laliberte, M.; Zaleska, I. Effects of root colonizing bacteria on nodulation of soybean roots by Bradyrhizobium japonicum. Can. J. Microbiol. 1987, 33, 498–503. [Google Scholar] [CrossRef]
- Plessner, O.; Klapatch, T.; Guerinot, M.L. Siderophore Utilization by Bradyrhizobium japonicum. Appl. Environ. Microbiol. 1993, 59, 1688–1690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Dashti, N.; Hynes, R.K.; Smith, D.L. Plant growth promoting Rhizobacteria and soybean [Glycine max (L.) Merr.] nodulation and nitrogen fixation at suboptimal root zone temperatures. Ann. Bot. 1996, 77, 453–460. [Google Scholar] [CrossRef]
- Jha, P.; Kumar, A. Characterization of novel plant growth promoting endophytic bacterium Achromobacter xylosoxidans from wheat plant. Microb. Ecol. 2009, 58, 179–188. [Google Scholar] [CrossRef]
- Dawwam, G.E.; Elbeltagy, A.; Emara, H.M.; Abbas, I.H.; Hassan, M.M. Beneficial effect of plant growth promoting bacteria isolated from the roots of potato plant. Ann. Agric. Sci. 2013, 58, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Rahman, H.M.; Salem, A.A.; Moustafa, M.M.A.; El-Garhy, H.A.S. A novice Achromobacter sp. EMCC1936 strain acts as a plant-growth-promoting agent. Acta Physiol. Plant. 2017, 39, 61. [Google Scholar] [CrossRef]
- De Campos, S.B.; Youn, J.-W.; Farina, R.; Jaenicke, S.; Jünemann, S.; Szczepanowski, R.; Beneduzi, A.; Vargas, L.; Goesmann, A.; Wendisch, V.F.; et al. Changes in root bacterial communities associated to two different development stages of canola (Brassica napus L. var oleifera) evaluated through next-generation sequencing technology. Microb. Ecol. 2013, 65, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Croes, S.; Weyens, N.; Janssen, J.; Vercampt, H.; Colpaert, J.; Carleer, R.; Vangronsveld, J. Bacterial communities associated with Brassica napus L. grown on trace element-contaminated and non-contaminated fields: A genotypic and phenotypic comparison. Microb. Biotechnol. 2013, 6, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Agler, M.T.; Ruhe, J.; Kroll, S.; Morhenn, C.; Kim, S.-T.; Weigel, D.; Kemen, E.M. Microbial hub taxa link host and abiotic factors to plant microbiome variation. PLoS Biol. 2016, 14, e1002352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, T.; Singh, D.K. Rhizospheric Microbacterium sp. P27 showing potential of lindane degradation and plant growth promoting Traits. Curr. Microbiol. 2019, 76, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Jaramillo, J.E.; Mendes, R.; Raaijmakers, J.M. Impact of plant domestication on rhizosphere microbiome assembly and functions. Plant. Mol. Biol. 2016, 90, 635–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueiredo, M.V.; Burity, H.A.; Martinez, C.; Chanway, C.P. Alleviation of drought stress in the common bean (Phaseolus vulgaris L.) by co-inoculation with Paenibacillus polymyxa and Rhizobium tropici. Appl. Soil Ecol. 2008, 40, 182–188. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | GS | Sobs | Coverage(%) | Chao | Ace | Shannon | Invsimpson | Npshannon |
|---|---|---|---|---|---|---|---|---|
| KA | BS | 2110.0 ± 65.4a | 0.9 ± 0.0 | 2875.1 ± 177.6ab | 3037.2 ± 169.4a | 6.1 ± 0.0ab | 84.1 ± 4.3ab | 6.3 ± 0.0a |
| V2 | 2255.0 ± 55.5a | 0.9 ± 0.0 | 3026.2 ± 223.6b | 3181.8 ± 222.3a | 6.3 ± 0.1b | 114.8 ± 32.7bc | 6.5 ± 0.1a | |
| R1 | 2230.7 ± 183.9a | 1.0 ± 0.0 | 2826.2 ± 475.8ab | 2962.2 ± 524.9a | 6.4 ± 0.1b | 140.4 ± 17.5bd | 6.6 ± 0.1a | |
| R3 | 2004.7 ± 152.7ab | 1.0 ± 0.0 | 2721.0 ± 333.3ab | 3039.6 ± 539.2a | 5.8 ± 0.4ab | 59.4 ± 30.5ac | 6.0 ± 0.4ab | |
| R5 | 2044.7 ± 117.9ab | 1.0 ± 0.0 | 2621.8 ± 123.2ab | 2755.7 ± 133.5a | 6.1 ± 0.3ab | 79.1 ± 46.1ab | 6.2 ± 0.3ab | |
| R7 | 1612.3 ± 274.3b | 1.0 ± 0.0 | 2185.0 ± 287.8a | 2308.4 ± 261.1a | 5.0 ± 0.9ac | 30.2 ± 25.7a | 5.1 ± 0.9b | |
| PS | BS | 2024.0 ± 128.3a | 1.0 ± 0.0 | 2689.3 ± 239.2a | 2815.0 ± 252.2a | 6.0 ± 0.1a | 73.5 ± 7.2a | 6.2 ± 0.1a |
| V2 | 2335.0 ± 183.4a | 0.9 ± 0.0 | 3156.0 ± 546.0a | 3512.8 ± 843.2a | 6.4 ± 0.1a | 110.2 ± 7.2a | 6.5 ± 0.1a | |
| R1 | 2397.7 ± 191.6a | 0.9 ± 0.0 | 3224.7 ± 480.2a | 3387.5 ± 514.2a | 6.5 ± 0.1a | 147.2 ± 64.3a | 6.7 ± 0.1a | |
| R3 | 2076.3 ± 113.9a | 1.0 ± 0.0 | 2728.8 ± 151.7a | 2852.6 ± 145.1a | 6.0 ± 0.3a | 75.7 ± 36.7a | 6.2 ± 0.3a | |
| R5 | 2252.0 ± 108.1a | 1.0 ± 0.0 | 2835.8 ± 200.2a | 2979.6 ± 190.0a | 6.4 ± 0.1a | 127.1 ± 15.1a | 6.5 ± 0.1a | |
| R7 | 2055.3 ± 257.5a | 0.9 ± 0.0 | 2789.6 ± 398.7a | 2942.4 ± 391.5a | 6.1 ± 0.4a | 105.6 ± 51.2a | 6.3 ± 0.3a | |
| PW | BS | 2113.3 ± 63.3a | 0.9 ± 0.0 | 2836.3 ± 88.0a | 2983.7 ± 130.1a | 6.2 ± 0.1ab | 98.3 ± 23.4ab | 6.3 ± 0.1ab |
| V2 | 2182.3 ± 446.2a | 0.9 ± 0.0 | 2952.6 ± 926.4a | 3284.5 ± 1210.4a | 6.2 ± 0.4ab | 103.2 ± 43.2ab | 6.4 ± 0.4b | |
| R1 | 2380.7 ± 309.2a | 0.9 ± 0.0 | 3321.0 ± 697.2a | 3474.1 ± 693.9a | 6.4 ± 0.2a | 149.5 ± 46.1a | 6.6 ± 0.2b | |
| R3 | 2062.7 ± 68.5a | 1.0 ± 0.0 | 2658.1 ± 5.0a | 2783.4 ± 8.1a | 6.1 ± 0.2ab | 83.5 ± 26.8ab | 6.2 ± 0.2ab | |
| R5 | 2066.7 ± 185.0a | 1.0 ± 0.0 | 2661.0 ± 324.6a | 2792.4 ± 256.3a | 6.2 ± 0.1ab | 108.1 ± 10.8ab | 6.3 ± 0.1ab | |
| R7 | 1862.3 ± 101.7a | 1.0 ± 0.0 | 2617.1 ± 82.4a | 2881.8 ± 347.0a | 5.4 ± 0.6bc | 31.3 ± 23.6bc | 5.5 ± 0.6ac | |
| TK | BS | 2063.3 ± 123.2ab | 1.0 ± 0.0 | 2730.5 ± 346.2a | 2844.0 ± 336.1a | 6.1 ± 0.1ab | 89.2 ± 13.1ab | 6.3 ± 0.1a |
| V2 | 2246.7 ± 115.9a | 0.9 ± 0.0 | 3132.7 ± 432.3a | 3464.0 ± 732.1a | 6.3 ± 0.0b | 102.7 ± 2.0b | 6.4 ± 0.0a | |
| R1 | 2212.3 ± 186.2a | 0.9 ± 0.0 | 3073.3 ± 513.0a | 3375.9 ± 756.7a | 6.3 ± 0.2b | 104.6 ± 51.3b | 6.4 ± 0.2a | |
| R3 | 2105.0 ± 52.8ab | 1.0 ± 0.0 | 2784.4 ± 168.5a | 2909.4 ± 174.7a | 6.2 ± 0.1ab | 106.2 ± 13.5b | 6.4 ± 0.1a | |
| R5 | 2141.3 ± 98.2ab | 1.0 ± 0.0 | 2714.2 ± 202.7a | 2874.4 ± 203.6a | 6.2 ± 0.1ab | 100.9 ± 13.2b | 6.4 ± 0.1a | |
| R7 | 1792.7 ± 189.5b | 1.0 ± 0.0 | 2447.5 ± 270.0a | 2590.1 ± 253.3a | 5.3 ± 0.8ac | 29.2 ± 20.3ac | 5.4 ± 0.7b |
| Variety | GS | Sobs | Coverage (%) | Chao | Ace | Shannon | Invsimpson | Npshannon |
|---|---|---|---|---|---|---|---|---|
| KA | V2 | 23 ± 1.4a | 1.0 ± 0.0 | 33 ± 3.6a | 46.8 ± 15.1a | 0.5 ± 0.1ab | 1.4 ± 0.1ab | 0.5 ± 0.1ab |
| R1 | 22 ± 2.2a | 1.0 ± 0.0 | 37.7 ± 3.8a | 55.8 ± 8.0a | 0.2 ± 0.1a | 1.1 ± 0.1bc | 0.2 ± 0.1b | |
| R3 | 67.7 ± 27.4b | 1.0 ± 0.0 | 100.3 ± 30.9a | 139.2 ± 32.5a | 0.3 ± 0.1a | 1.1 ± 0.1bc | 0.3 ± 0.1b | |
| R5 | 38.0 ± 17.1ab | 1.0 ± 0.0 | 60.8 ± 40.3a | 88.0 ± 68.1a | 0.2 ± 0.1a | 1.0 ± 0.0c | 0.2 ± 0.1b | |
| R7 | 71.3 ± 14.1bc | 1.0 ± 0.0 | 94.8 ± 36.0a | 108.6 ± 49.2a | 0.8 ± 0.3b | 1.5 ± 0.2a | 0.9 ± 0.3ac | |
| PS | V2 | 32 ± 9.2a | 1.0 ± 0.0 | 60.3 ± 16.5a | 73.9 ± 13.2a | 0.2 ± 0.1a | 1.1 ± 0.0a | 0.2 ± 0.1a |
| R1 | 31.3 ± 11.0a | 1.0 ± 0.0 | 43.4 ± 9.6a | 56.3 ± 4.2a | 0.2 ± 0.1a | 1.1 ± 0.1a | 0.2 ± 0.1a | |
| R3 | 53.0 ± 6.2a | 1.0 ± 0.0 | 82.0 ± 8.0a | 85.1 ± 12.6a | 0.2 ± 0.1a | 1.1 ± 0.0a | 0.2 ± 0.1a | |
| R5 | 28.0 ± 14.2a | 1.0 ± 0.0 | 44.9 ± 22.2a | 74.0 ± 34.6a | 0.4 ± 0.1ab | 1.2 ± 0.1a | 0.4 ± 0.1ab | |
| R7 | 118.7 ± 75.0a | 1.0 ± 0.0 | 138.5 ± 91.4a | 148.7 ± 95.1a | 1.1 ± 0.6b | 1.6 ± 0.4a | 1.1 ± 0.6bc | |
| PW | V2 | 31.5 ± 0.5a | 1.0 ± 0.0 | 46 ± 7.7a | 65 ± 17.6a | 0.2 ± 0.2a | 1.1 ± 0.1a | 0.2 ± 0.2a |
| R1 | 25.7 ± 1.2a | 1.0 ± 0.0 | 34.5 ± 6.7a | 43.5 ± 10.6a | 0.1 ± 0.1a | 1.1 ± 0.1a | 0.2 ± 0.1a | |
| R3 | 41.3 ± 8.7a | 1.0 ± 0.0 | 74.1 ± 31.9ab | 83.9 ± 31.5a | 0.1 ± 0.1a | 1.0 ± 0.0a | 0.1 ± 0.1a | |
| R5 | 21.0 ± 8.6a | 1.0 ± 0.0 | 24.3 ± 9.7a | 34.8 ± 14.8a | 0.1 ± 0.1a | 1.1 ± 0.1a | 0.1 ± 0.1a | |
| R7 | 83.0 ± 25.2b | 1.0 ± 0.0 | 121.3 ± 37.0b | 156.0 ± 31.2b | 0.6 ± 0.5a | 1.3 ± 0.3a | 0.6 ± 0.5a | |
| TK | V2 | 23.7 ± 0.9a | 10. ± 0.0 | 48.4 ± 12.6ab | 84.8 ± 45.8a | 0.3 ± 0.2ab | 1.2 ± 0.1a | 0.3 ± 0.2ab |
| R1 | 22.3 ± 3.9a | 1.0 ± 0.0 | 36.0 ± 8.3a | 46.1 ± 17.9a | 0.1 ± 0.1a | 1.1 ± 0.0a | 0.1 ± 0.1a | |
| R3 | 47.3 ± 24.0a | 1.0 ± 0.0 | 70.9 ± 23.0ab | 92.2 ± 32.7a | 0.3 ± 0.3ab | 1.2 ± 0.2a | 0.3 ± 0.3ab | |
| R5 | 29.3 ± 18.2a | 1.0 ± 0.0 | 37.7 ± 24.3a | 40.2 ± 24.9a | 0.1 ± 0.1a | 1.0 ± 0.0a | 0.1 ± 0.1a | |
| R7 | 103.3 ± 61.4a | 1.0 ± 0.0 | 133.0 ± 64.2b | 132.5 ± 68.2a | 1.2 ± 0.8bc | 2.0 ± 1.0a | 1.2 ± 0.8bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sohn, S.-I.; Ahn, J.-H.; Pandian, S.; Oh, Y.-J.; Shin, E.-K.; Kang, H.-J.; Cho, W.-S.; Cho, Y.-S.; Shin, K.-S. Dynamics of Bacterial Community Structure in the Rhizosphere and Root Nodule of Soybean: Impacts of Growth Stages and Varieties. Int. J. Mol. Sci. 2021, 22, 5577. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115577
Sohn S-I, Ahn J-H, Pandian S, Oh Y-J, Shin E-K, Kang H-J, Cho W-S, Cho Y-S, Shin K-S. Dynamics of Bacterial Community Structure in the Rhizosphere and Root Nodule of Soybean: Impacts of Growth Stages and Varieties. International Journal of Molecular Sciences. 2021; 22(11):5577. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115577
Chicago/Turabian StyleSohn, Soo-In, Jae-Hyung Ahn, Subramani Pandian, Young-Ju Oh, Eun-Kyoung Shin, Hyeon-Jung Kang, Woo-Suk Cho, Youn-Sung Cho, and Kong-Sik Shin. 2021. "Dynamics of Bacterial Community Structure in the Rhizosphere and Root Nodule of Soybean: Impacts of Growth Stages and Varieties" International Journal of Molecular Sciences 22, no. 11: 5577. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115577