
PSA Depletion Induces the Differentiation of Immature Neurons in the Piriform Cortex of Adult Mice

 , and
, and
Abstract
:
1. Introduction
2. Results
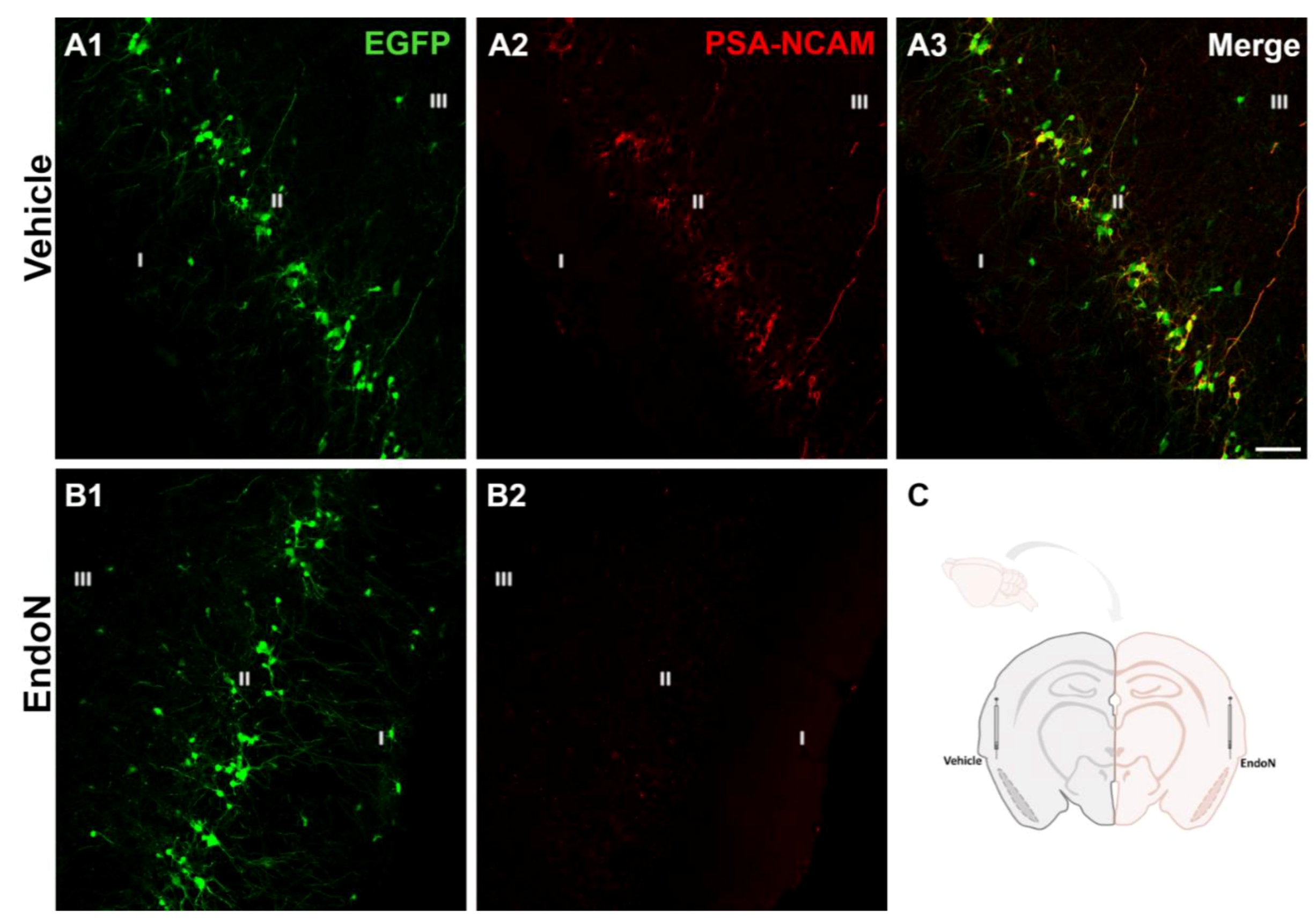
2.1. Endo-N Efficiently Depleted PSA from the PCX
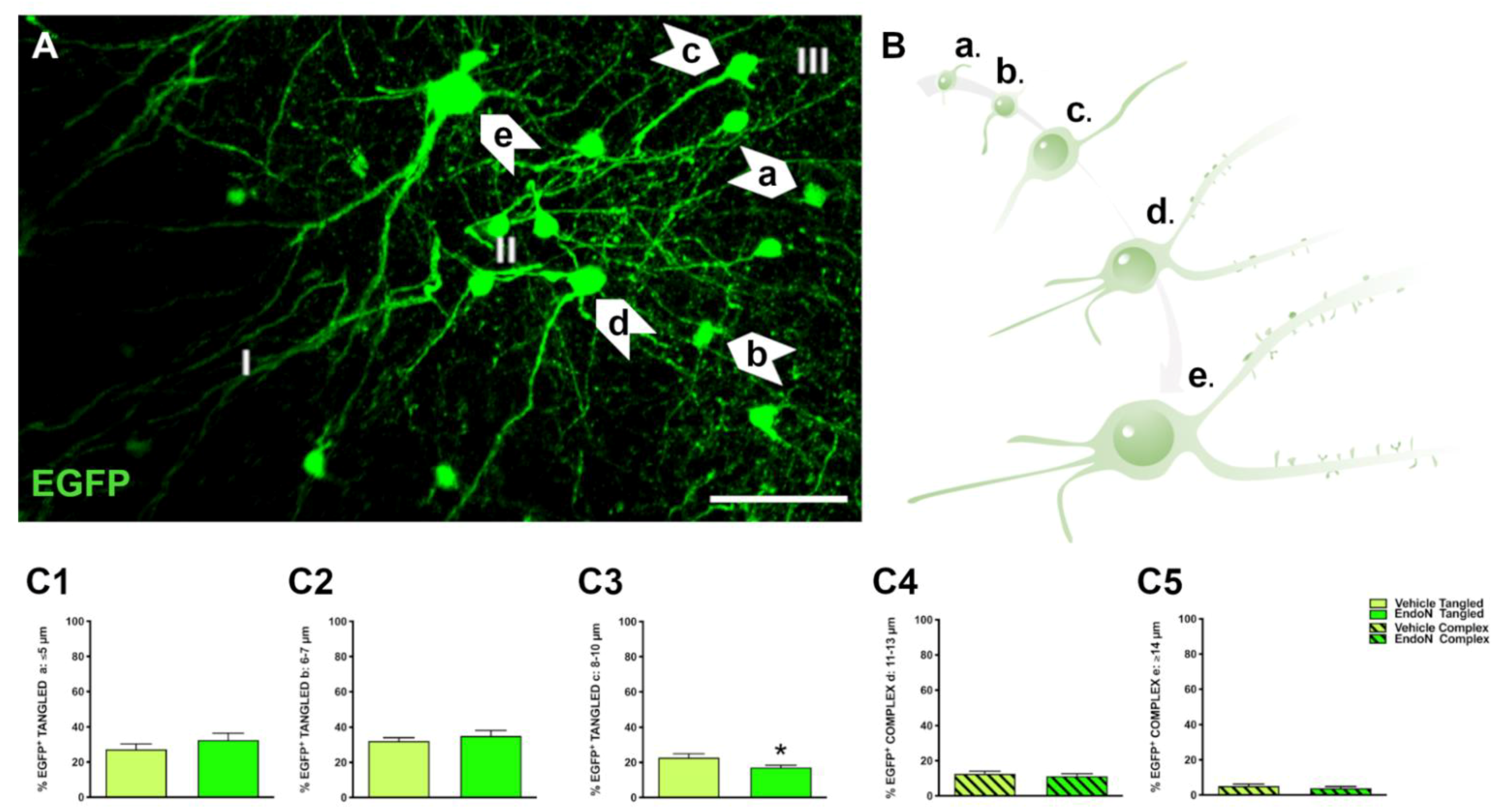
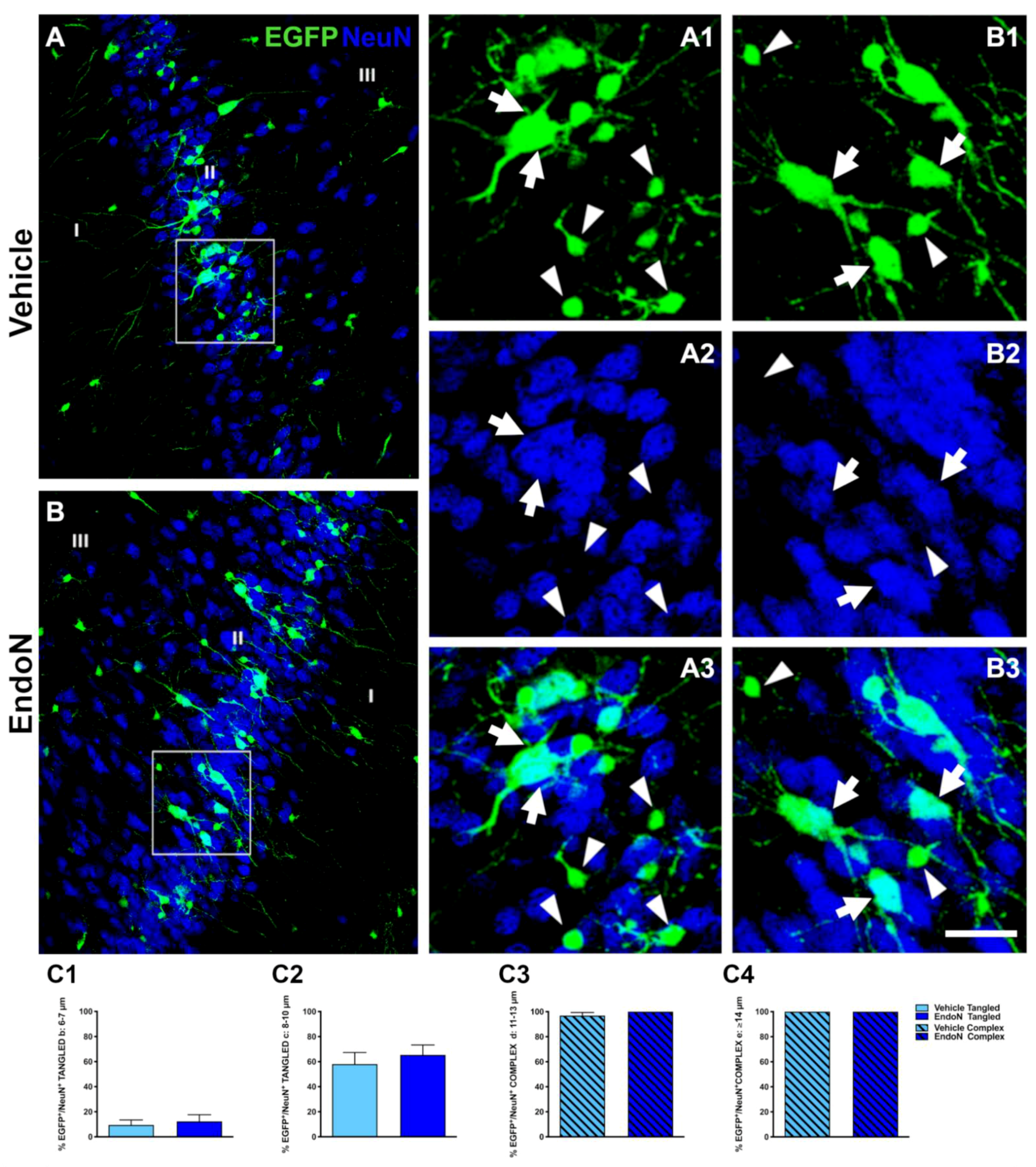
2.2. Classification of EGFP+ Neurons and Impact of PSA Depletion in the PCX Layer II
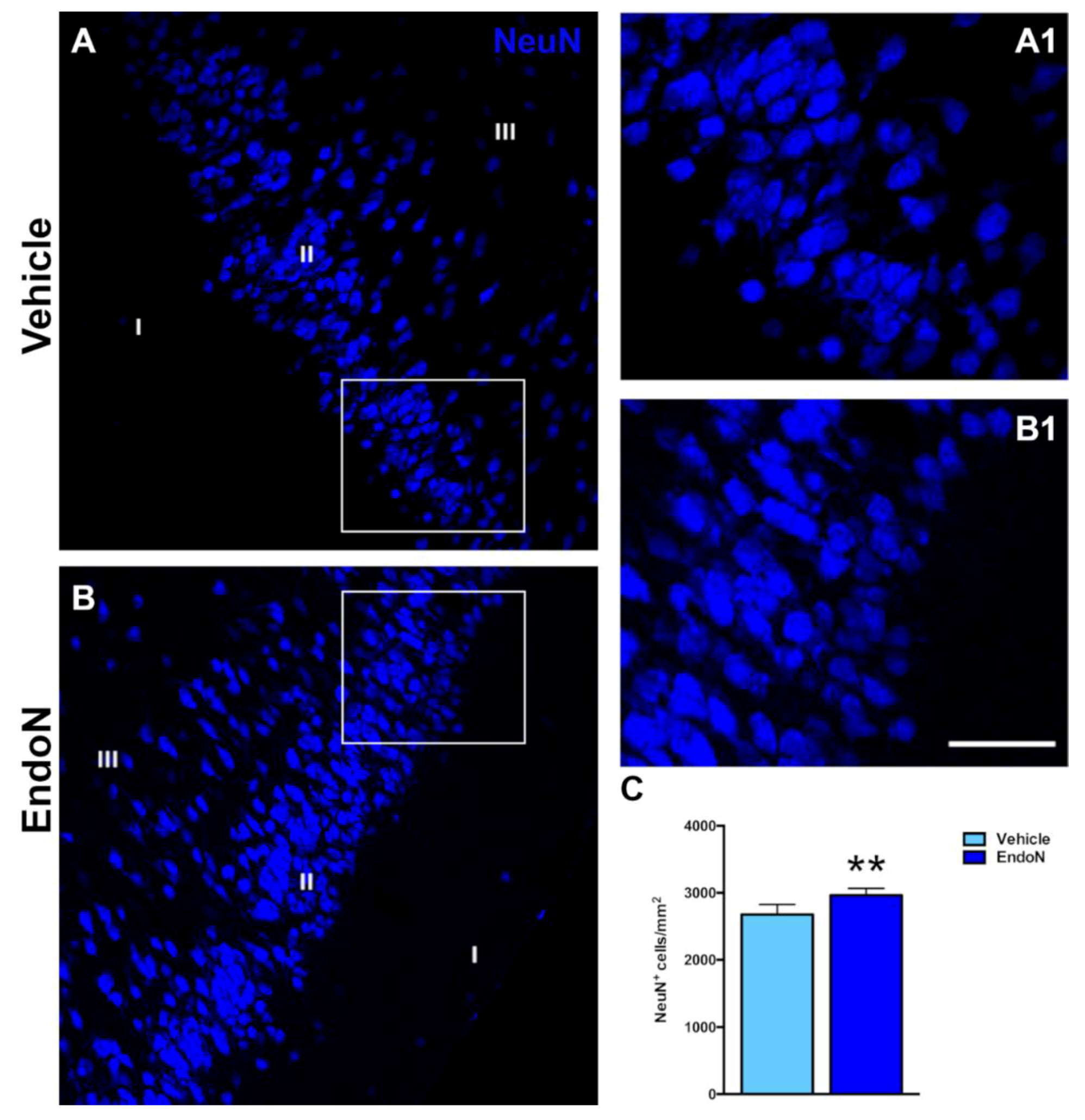
2.3. PSA Depletion Promoted Expression of Mature Neuronal Markers in EGFP+ Cells of the PCX Layer II
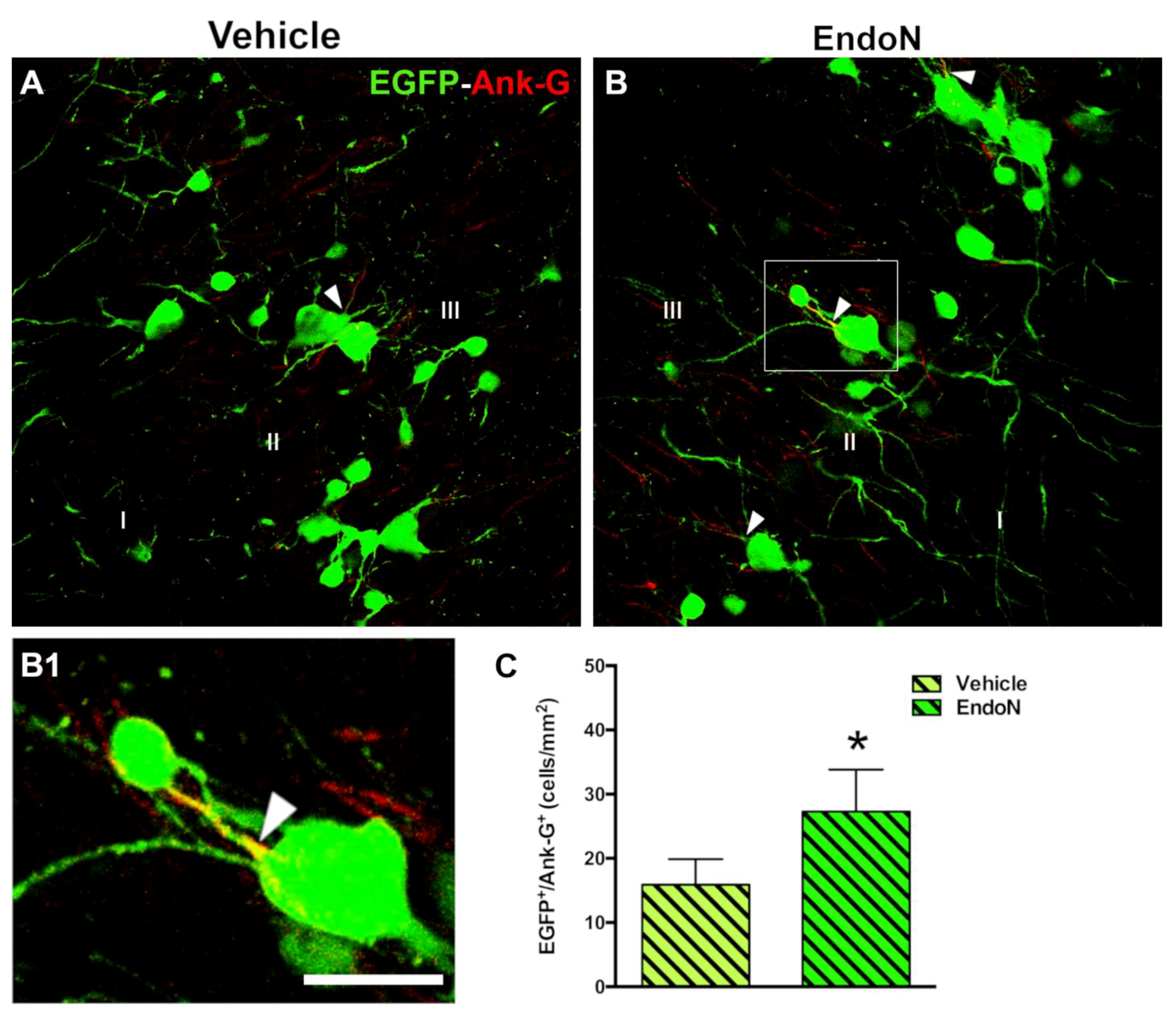
2.4. PSA Depletion Increased the Density of EGFP+ Cells Bearing an Axon Initial Segment in the PCX Layer II
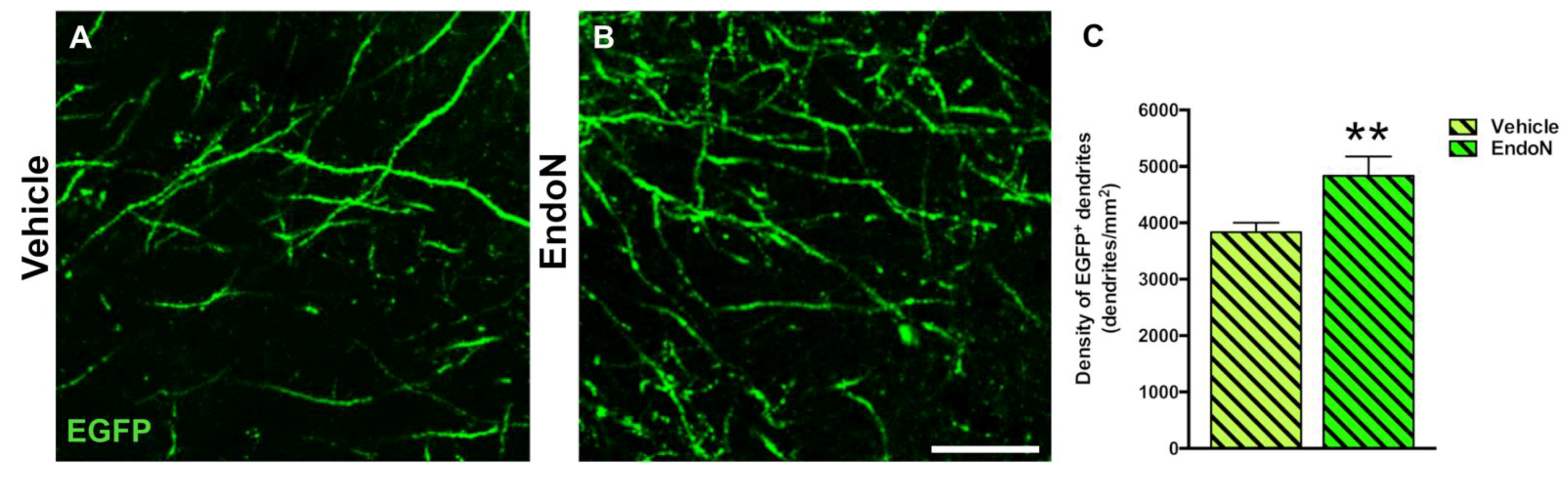
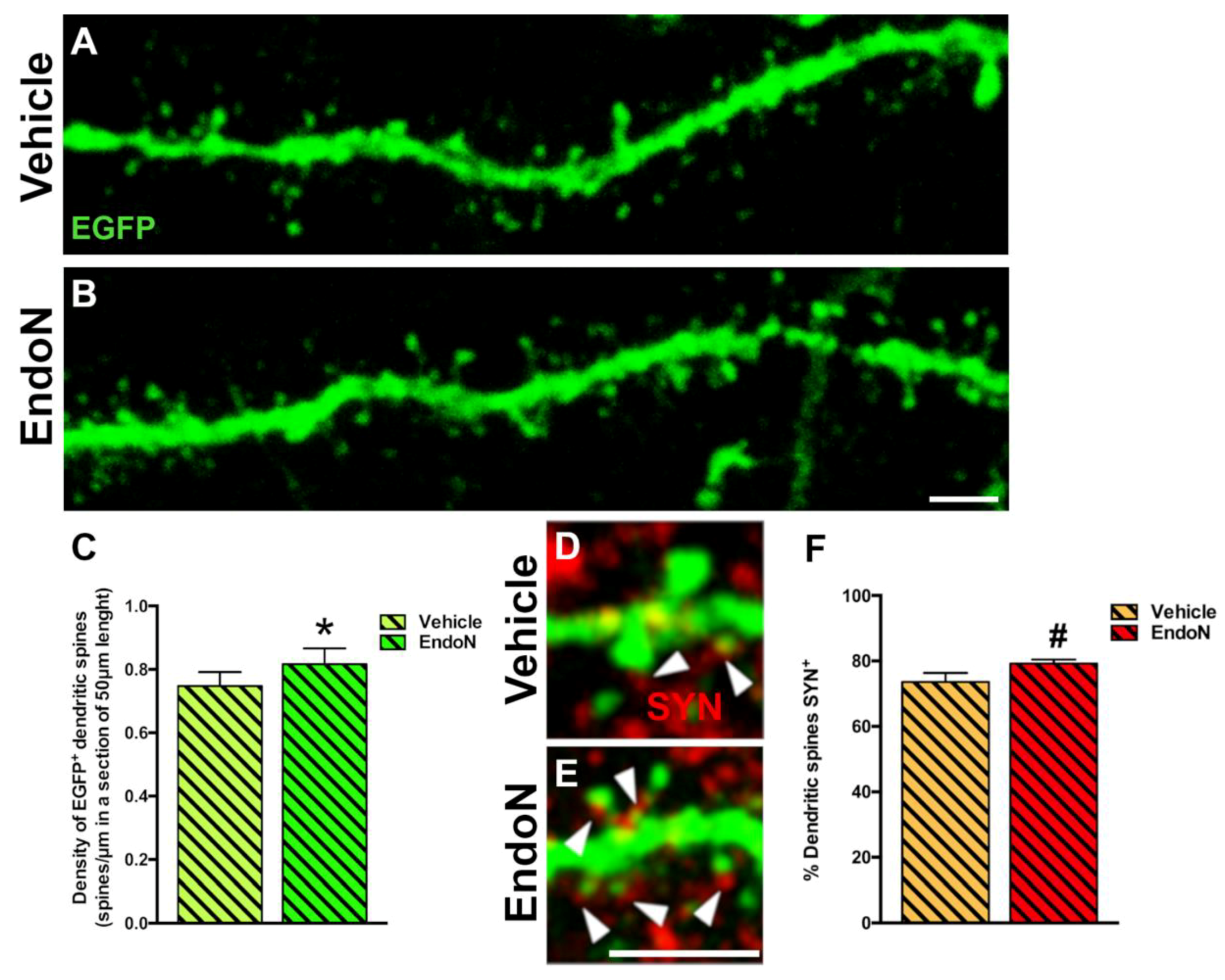
2.5. Effects of PSA Depletion on EGFP+ Dendrites and Dendritic Spines in the PCX Layer I
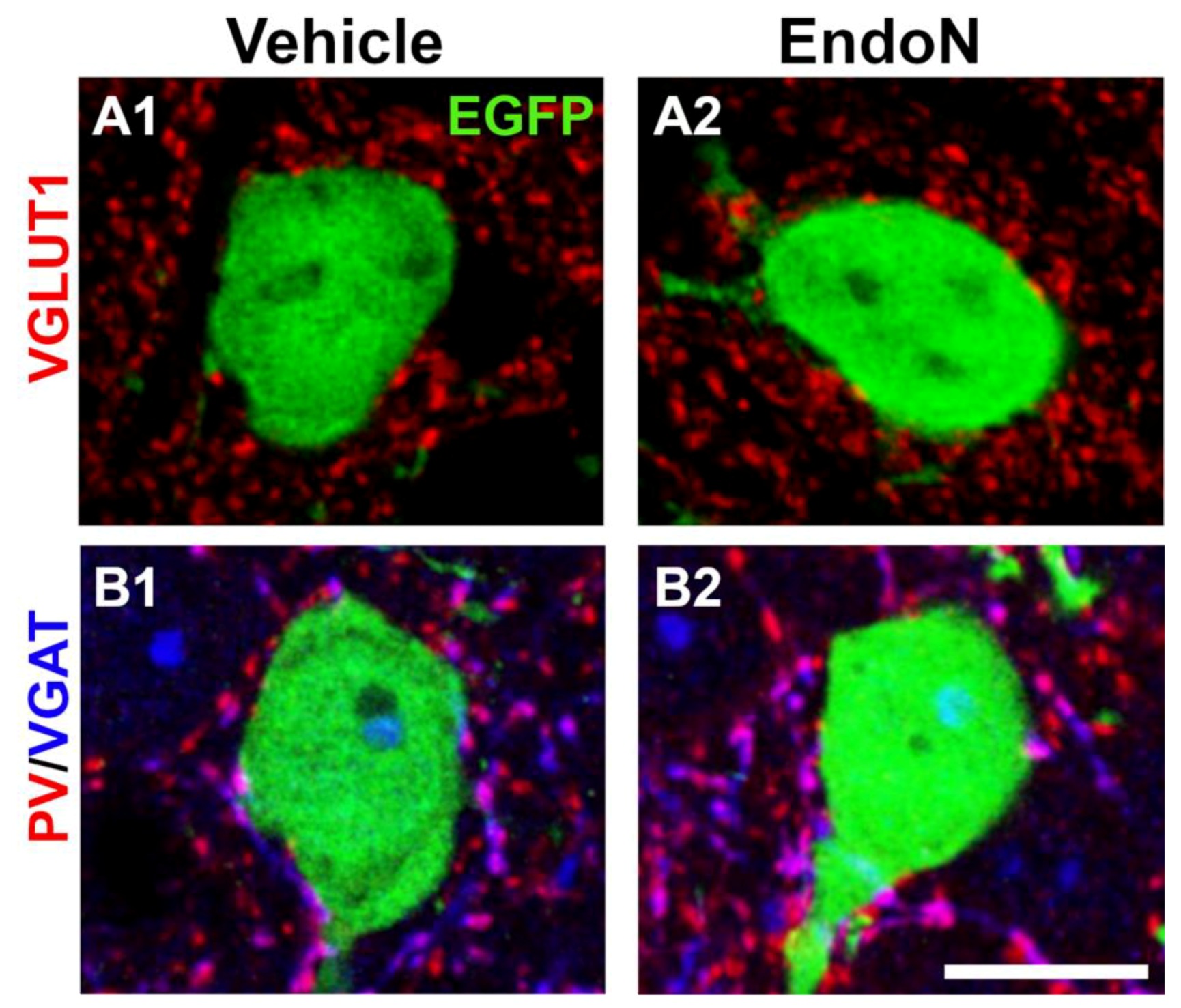
2.6. PSA Depletion Did Not Change the Density of Excitatory and Inhibitory Synaptic Puncta in the Perisomatic Region of EGFP+ Complex Cells
3. Discussion
4. Material and Methods
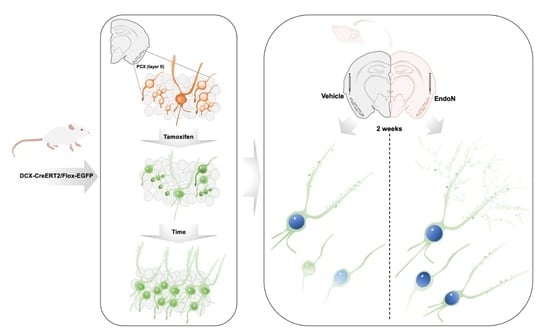
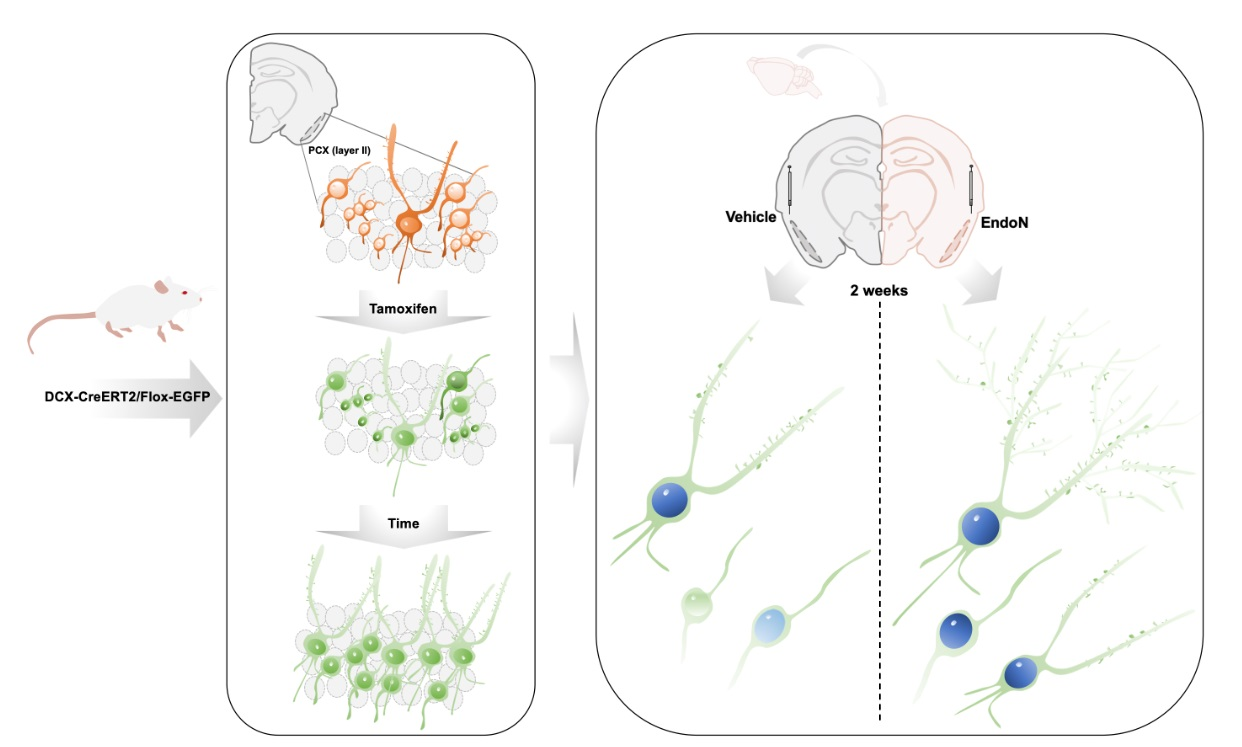
4.1. Transgenic Animals
4.2. Polysialic Acid Depletion
4.3. Histological Procedures
4.4. Immunohistochemistry
4.5. Confocal Analysis and Quantification
4.5.1. Classification and Phenotypic Characterization of EGFP Neurons
4.5.2. Quantification of Cellular Densities
4.5.3. Quantification of Axonal Initial Segment Densities
4.5.4. Analysis of Dendrites and Dendritic Spines
4.5.5. Analysis of Presynaptic Inputs on EGFP+ Spines
4.5.6. Analysis of Perisomatic Puncta Expressing Excitatory/Inhibitory Synaptic Markers
5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lois, C.; Alvarez-Buylla, A. Long-Distance Neuronal Migration in the Adult Mammalian Brain. Science 1994, 264, 1145–1148. [Google Scholar] [CrossRef]
- Gage, F.H. Mammalian Neural Stem Cells. Science 2000, 287, 1433–1438. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, D.M.; Bordey, A.; Bonfanti, L. Noncanonical Sites of Adult Neurogenesis in the Mammalian Brain. Cold Spring Harb. Perspect Biol. 2015, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo-Gómez, E.; Gómez-Climent, M.Á.; Varea, E.; Guirado, R.; Blasco-Ibáñez, J.M.; Crespo, C.; Martínez-Guijarro, F.J.; Nácher, J. Dopamine Acting through D2 Receptors Modulates the Expression of PSA-NCAM, a Molecule Related to Neuronal Structural Plasticity, in the Medial Prefrontal Cortex of Adult Rats. Exp. Neurol. 2008, 214, 97–111. [Google Scholar] [CrossRef]
- Bonfanti, L.; Nacher, J. New Scenarios for Neuronal Structural Plasticity in Non-Neurogenic Brain Parenchyma: The Case of Cortical Layer II Immature Neurons. Prog. Neurobiol. 2012, 98, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Rubio, A.; Bellés, M.; Belenguer, G.; Vidueira, S.; Fariñas, I.; Nacher, J. Characterization and Isolation of Immature Neurons of the Adult Mouse Piriform Cortex. Dev. Neurobiol. 2015, 76. [Google Scholar] [CrossRef]
- Xiong, K.; Luo, D.W.; Patrylo, P.R.; Luo, X.G.; Struble, R.G.; Clough, R.W.; Yan, X.X. Doublecortin-Expressing Cells Are Present in Layer II across the Adult Guinea Pig Cerebral Cortex: Partial Colocalization with Mature Interneuron Markers. Exp. Neurol. 2008, 211, 271–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Xiong, K.; Chu, Y.; Luo, D.-W.; Luo, X.-G.; Yuan, X.-Y.; Struble, R.G.; Clough, R.W.; Spencer, D.D.; Williamson, A.; et al. Doublecortin Expression in Adult Cat and Primate Cerebral Cortex Relates to Immature Neurons That Develop into GABAergic Subgroups. Exp. Neurol. 2009, 216, 342–356. [Google Scholar] [CrossRef] [Green Version]
- Varea, E.; Belles, M.; Vidueira, S.; Blasco-Ibáñez, J.M.; Crespo, C.; Pastor, Á.M.; Nacher, J. PSA-NCAM Is Expressed in Immature, but Not Recently Generated, Neurons in the Adult Cat Cerebral Cortex Layer II. Front. Neurosci. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- König, R.; Benedetti, B.; Rotheneichner, P.; O’ Sullivan, A.; Kreutzer, C.; Belles, M.; Nacher, J.; Weiger, T.M.; Aigner, L.; Couillard-Després, S. Distribution and Fate of DCX/PSA-NCAM Expressing Cells in the Adult Mammalian Cortex: A Local Reservoir for Adult Cortical Neuroplasticity? Front. Biol. 2016, 11, 193–213. [Google Scholar] [CrossRef]
- Rotheneichner, P.; Belles, M.; Benedetti, B.; König, R.; Dannehl, D.; Kreutzer, C.; Zaunmair, P.; Engelhardt, M.; Aigner, L.; Nacher, J.; et al. Cellular Plasticity in the Adult Murine Piriform Cortex: Continuous Maturation of Dormant Precursors into Excitatory Neurons. Cereb. Cortex 2018, 28, 2610–2621. [Google Scholar] [CrossRef] [Green Version]
- La Rosa, C.; Cavallo, F.; Pecora, A.; Chincarini, M.; Ala, U.; Faulkes, C.G.; Nacher, J.; Cozzi, B.; Sherwood, C.C.; Amrein, I.; et al. Phylogenetic Variation in Cortical Layer II Immature Neuron Reservoir of Mammals. eLife 2020, 9, e55456. [Google Scholar] [CrossRef]
- Gómez-Climent, M.A.; Castillo-Gómez, E.; Varea, E.; Guirado, R.; Blasco-Ibáñez, J.M.; Crespo, C.; Martínez-Guijarro, F.J.; Nácher, J. A Population of Prenatally Generated Cells in the Rat Paleocortex Maintains an Immature Neuronal Phenotype into Adulthood. Cereb. Cortex 2008, 18, 2229–2240. [Google Scholar] [CrossRef] [Green Version]
- Varea, E.; Castillo-Gómez, E.; Gómez-Climent, M.Á.; Guirado, R.; Blasco-Ibáñez, J.M.; Crespo, C.; Martínez-Guijarro, F.J.; Nácher, J. Differential Evolution of PSA-NCAM Expression during Aging of the Rat Telencephalon. Neurobiol. Aging 2009, 30, 808–818. [Google Scholar] [CrossRef]
- Benedetti, B.; Dannehl, D.; König, R.; Coviello, S.; Kreutzer, C.; Zaunmair, P.; Jakubecova, D.; Weiger, T.M.; Aigner, L.; Nacher, J.; et al. Functional Integration of Neuronal Precursors in the Adult Murine Piriform Cortex. Cereb. Cortex 2020, 30, 1499–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seki, T.; Arai, Y. Expression of Highly Polysialylated NCAM in the Neocortex and Piriform Cortex of the Developing and the Adult Rat. Anat. Embryol. 1991, 184, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Bonfanti, L.; Theodosis, D.T. Expression of Polysialylated Neural Cell Adhesion Molecule by Proliferating Cells in the Subependymal Layer of the Adult Rat, in Its Rostral Extension and in the Olfactory Bulb. Neuroscience 1994, 62, 291–305. [Google Scholar] [CrossRef]
- Brown, J.P.; Couillard-Després, S.; Cooper-Kuhn, C.M.; Winkler, J.; Aigner, L.; Kuhn, H.G. Transient Expression of Doublecortin during Adult Neurogenesis. J. Comp. Neurol. 2003, 467, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Couillard-Despres, S.; Winner, B.; Schaubeck, S.; Aigner, R.; Vroemen, M.; Weidner, N.; Bogdahn, U.; Winkler, J.; Kuhn, H.-G.; Aigner, L. Doublecortin Expression Levels in Adult Brain Reflect Neurogenesis. Eur. J. Neurosci. 2005, 21, 1–14. [Google Scholar] [CrossRef]
- Luzzati, F.; Bonfanti, L.; Fasolo, A.; Peretto, P. DCX and PSA-NCAM Expression Identifies a Population of Neurons Preferentially Distributed in Associative Areas of Different Pallial Derivatives and Vertebrate Species. Cereb. Cortex May 2009, 19, 1028–1041. [Google Scholar] [CrossRef] [Green Version]
- Seki, T.; Arai, Y. Distribution and Possible Roles of the Highly Polysialylated Neural Cell Adhesion Molecule (NCAM-H) in the Developing and Adult Central Nervous System. Neurosci. Res. 1993, 17, 265–290. [Google Scholar] [CrossRef]
- Bonfanti, L. PSA-NCAM in Mammalian Structural Plasticity and Neurogenesis. Prog. Neurobiol. 2006, 80, 129–164. [Google Scholar] [CrossRef] [PubMed]
- Gascon, E.; Vutskits, L.; Kiss, J.Z. Polysialic Acid–Neural Cell Adhesion Molecule in Brain Plasticity: From Synapses to Integration of New Neurons. Brain Res. Rev. 2007, 56, 101–118. [Google Scholar] [CrossRef]
- Rutishauser, U. Polysialic Acid in the Plasticity of the Developing and Adult Vertebrate Nervous System. Nat. Rev. Neurosci. 2008, 9, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, H.; Dityatev, A. Polysialic Acid in Brain Development and Synaptic Plasticity. In SialoGlyco Chemistry and Biology I: Biosynthesis, Structural Diversity and Sialoglycopathologies; Topics in Current Chemistry; Gerardy-Schahn, R., Delannoy, P., von Itzstein, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 55–96. ISBN 978-3-662-47940-7. [Google Scholar]
- Eckhardt, M.; Bukalo, O.; Chazal, G.; Wang, L.; Goridis, C.; Schachner, M.; Gerardy-Schahn, R.; Cremer, H.; Dityatev, A. Mice Deficient in the Polysialyltransferase ST8SiaIV/PST-1 Allow Discrimination of the Roles of Neural Cell Adhesion Molecule Protein and Polysialic Acid in Neural Development and Synaptic Plasticity. J. Neurosci. 2000, 20, 5234–5244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCall, T.; Weil, Z.M.; Nacher, J.; Bloss, E.B.; El Marouf, A.; Rutishauser, U.; McEwen, B.S. Depletion of Polysialic Acid from Neural Cell Adhesion Molecule (PSA-NCAM) Increases CA3 Dendritic Arborization and Increases Vulnerability to Excitotoxicity. Exp. Neurol. 2013, 241, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Guirado, R.; Perez-Rando, M.; Sanchez-Matarredona, D.; Castillo-Gómez, E.; Liberia, T.; Rovira-Esteban, L.; Varea, E.; Crespo, C.; Blasco-Ibáñez, J.M.; Nacher, J. The Dendritic Spines of Interneurons Are Dynamic Structures Influenced by PSA-NCAM Expression. Cereb. Cortex 2014, 24, 3014–3024. [Google Scholar] [CrossRef]
- Guirado, R.; La Terra, D.; Bourguignon, M.; Carceller, H.; Umemori, J.; Sipilä, P.; Nacher, J.; Castrén, E. Effects of PSA Removal from NCAM on the Critical Period Plasticity Triggered by the Antidepressant Fluoxetine in the Visual Cortex. Front Cell Neurosci. 2016, 10. [Google Scholar] [CrossRef] [Green Version]
- Vimr, E.R.; McCoy, R.D.; Vollger, H.F.; Wilkison, N.C.; Troy, F.A. Use of Prokaryotic-Derived Probes to Identify Poly(Sialic Acid) in Neonatal Neuronal Membranes. Proc. Natl. Acad. Sci. USA 1984, 81, 1971–1975. [Google Scholar] [CrossRef] [Green Version]
- Petridis, A.K.; Maarouf, A.E.; Rutishauser, U. Polysialic Acid Regulates Cell Contact-Dependent Neuronal Differentiation of Progenitor Cells from the Subventricular Zone. Dev. Dyn. 2004, 230, 675–684. [Google Scholar] [CrossRef]
- Burgess, A.; Wainwright, S.R.; Shihabuddin, L.S.; Rutishauser, U.; Seki, T.; Aubert, I. Polysialic Acid Regulates the Clustering, Migration, and Neuronal Differentiation of Progenitor Cells in the Adult Hippocampus. Dev. Neurobiol. 2008, 68, 1580–1590. [Google Scholar] [CrossRef]
- Zhang, J.; Giesert, F.; Kloos, K.; Vogt Weisenhorn, D.M.; Aigner, L.; Wurst, W.; Couillard-Despres, S. A Powerful Transgenic Tool for Fate Mapping and Functional Analysis of Newly Generated Neurons. BMC Neurosci. 2010, 11, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, K.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates. The Coronal Plates and Diagrams; Franklin, K., Paxinos, G., Eds.; Academic Press: Cambridge, MA, USA, 2008; ISBN 9780123742445. [Google Scholar]
- Mullen, R.J.; Buck, C.R.; Smith, A.M. NeuN, a Neuronal Specific Nuclear Protein in Vertebrates. Development 1992, 116, 201–211. [Google Scholar] [CrossRef]
- Wayman, G.A.; Lee, Y.-S.; Tokumitsu, H.; Silva, A.; Soderling, T.R. Calmodulin-Kinases: Modulators of Neuronal Development and Plasticity. Neuron 2008, 59, 914–931. [Google Scholar] [CrossRef] [Green Version]
- Burgess, A.; Weng, Y.-Q.; Ypsilanti, A.R.; Cui, X.; Caines, G.; Aubert, I. Polysialic Acid Limits Septal Neurite Outgrowth on Laminin. Brain Res. 2007, 1144, 52–58. [Google Scholar] [CrossRef]
- Castillo-Gómez, E.; Pérez-Rando, M.; Vidueira, S.; Nacher, J. Polysialic Acid Acute Depletion Induces Structural Plasticity in Interneurons and Impairs the Excitation/Inhibition Balance in Medial Prefrontal Cortex Organotypic Cultures. Front. Cell. Neurosci. 2016, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toni, N.; Schinder, A.F. Maturation and Functional Integration of New Granule Cells into the Adult Hippocampus. Cold Spring Harb. Perspect Biol. 2016, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Praag, H.; Schinder, A.F.; Christie, B.R.; Toni, N.; Palmer, T.D.; Gage, F.H. Functional Neurogenesis in the Adult Hippocampus. Nature 2002, 415, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Spampanato, J.; Sullivan, R.K.; Turpin, F.R.; Bartlett, P.F.; Sah, P. Properties of Doublecortin Expressing Neurons in the Adult Mouse Dentate Gyrus. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- 2Evans, M.D.; Dumitrescu, A.S.; Kruijssen, D.L.H.; Taylor, S.E.; Grubb, M.S. Rapid Modulation of Axon Initial Segment Length Influences Repetitive Spike Firing. Cell Rep. 2015, 13, 1233–1245. [Google Scholar] [CrossRef] [Green Version]
- Yamada, R.; Kuba, H. Structural and Functional Plasticity at the Axon Initial Segment. Front. Cell Neurosci. 2016, 10, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolós, M.; Terreros-Roncal, J.; Perea, J.R.; Pallas-Bazarra, N.; Ávila, J.; Llorens-Martín, M. Maturation Dynamics of the Axon Initial Segment (AIS) of Newborn Dentate Granule Cells in Young Adult C57BL/6J Mice. J. Neurosci. 2019, 1605–1620. [Google Scholar] [CrossRef] [PubMed]
- Kochlamazashvili, G.; Senkov, O.; Grebenyuk, S.; Robinson, C.; Xiao, M.-F.; Stummeyer, K.; Gerardy-Schahn, R.; Engel, A.K.; Feig, L.; Semyanov, A.; et al. Neural Cell Adhesion Molecule-Associated Polysialic Acid Regulates Synaptic Plasticity and Learning by Restraining the Signaling through GluN2B-Containing NMDA Receptors. J. Neurosci. 2010, 30, 4171–4183. [Google Scholar] [CrossRef]
- Kiss, J.Z.; Rougon, G. Cell Biology of Polysialic Acid. Curr. Opin. Neurobiol. 1997, 7, 640–646. [Google Scholar] [CrossRef]
- Maarouf, A.E.; Rutishauser, U. Removal of Polysialic Acid Induces Aberrant Pathways, Synaptic Vesicle Distribution, and Terminal Arborization of Retinotectal Axons. J. Comp. Neurol. 2003, 460, 203–211. [Google Scholar] [CrossRef]
- Troy, F.A.; Hallenbeck, P.C.; McCoy, R.D.; Vimr, E.R. Detection of Polysialosyl-Containing Glycoproteins in Brain Using P9rokaryotic-Derived Probes. Methods Enzymol. 1987, 138, 169–185. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Guirado, R.; Carceller, H.; Castillo-Gómez, E.; Castrén, E.; Nacher, J. Automated Analysis of Images for Molecular Quantification in Immunohistochemistry. Heliyon 2018, 4, e00669. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anti | Host | Isotype | Dilution | Cat Number | Company |
|---|---|---|---|---|---|
| Ank-G | Mouse | IgG1 | 1:200 | SC12719 | Santa Cruz |
| CAMK-II | Mouse | IgG1 | 1:1000 | ab22609 | Abcam |
| DCX | Rabbit | IgG | 1:2000 | ab207175 | Abcam |
| GFP | Chicken | IgY | 1:1000 | A10262 | Thermo Fisher |
| NeuN | Mouse | IgG1 | 1:1000 | MAB377 | Sigma-Aldrich |
| PSA-NCAM | Mouse | IgM | 1:1400 | MAB5324 | Sigma-Aldrich |
| PV | Guinea pig | IgG | 1:2000 | 195004 | SySy |
| SYN | Mouse | IgG1 | 1:1000 | S5768 | Sigma-Aldrich |
| VGAT | Rabbit | IgG | 1:1000 | 131002 | SySy |
| VGLUT1 | Guinea pig | IgG | 1:1000 | AB5905 | Sigma-Aldrich |
| Anti | Host | Conjugate | Dilution | Cat number | Company |
|---|---|---|---|---|---|
| Chicken IgY | Donkey | CF488 | 1:400 | 20020 | Biotium |
| Guinea pig IgG | Goat | AF647 | 1:400 | 106605003 | Jackson ImmunoResearch |
| Mouse IgG1 | Goat | CF647 | 1:400 | 20252 | Biotium |
| Mouse IgG1 | Goat | AF555 | 1:400 | A21127 | Thermo Fisher |
| Mouse IgM | Goat | CF555 | 1:400 | 20485 | Biotium |
| Rabbit IgG | Donkey | CF555 | 1:400 | 20038 | Biotium |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coviello, S.; Benedetti, B.; Jakubecova, D.; Belles, M.; Klimczak, P.; Gramuntell, Y.; Couillard-Despres, S.; Nacher, J. PSA Depletion Induces the Differentiation of Immature Neurons in the Piriform Cortex of Adult Mice. Int. J. Mol. Sci. 2021, 22, 5733. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115733
Coviello S, Benedetti B, Jakubecova D, Belles M, Klimczak P, Gramuntell Y, Couillard-Despres S, Nacher J. PSA Depletion Induces the Differentiation of Immature Neurons in the Piriform Cortex of Adult Mice. International Journal of Molecular Sciences. 2021; 22(11):5733. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115733
Chicago/Turabian StyleCoviello, Simona, Bruno Benedetti, Dominika Jakubecova, Maria Belles, Patrycja Klimczak, Yaiza Gramuntell, Sebastien Couillard-Despres, and Juan Nacher. 2021. "PSA Depletion Induces the Differentiation of Immature Neurons in the Piriform Cortex of Adult Mice" International Journal of Molecular Sciences 22, no. 11: 5733. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115733