
Current Perspectives on the Physiological Activities of Fermented Soybean-Derived Cheonggukjang


Abstract
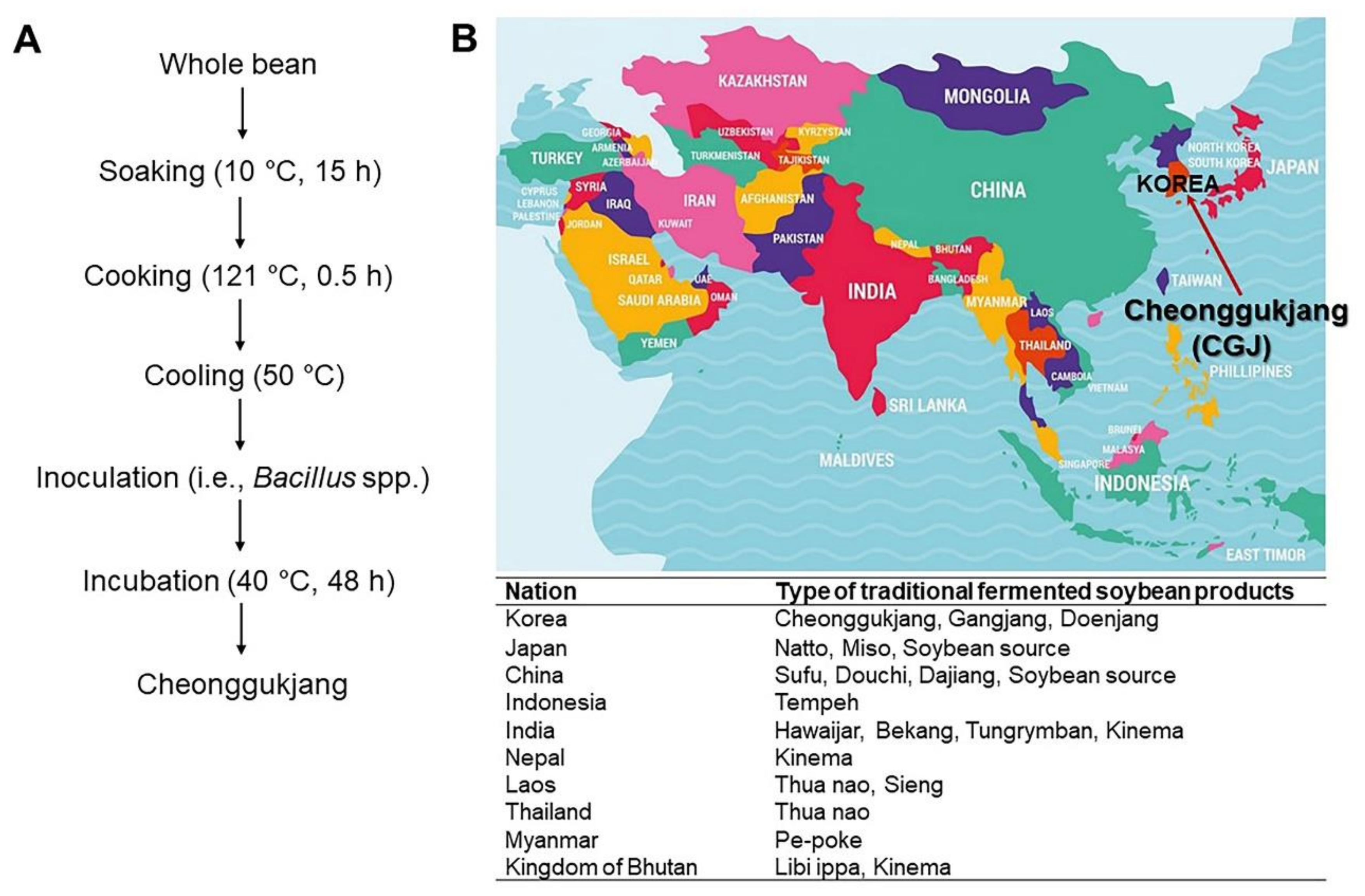
:1. Introduction
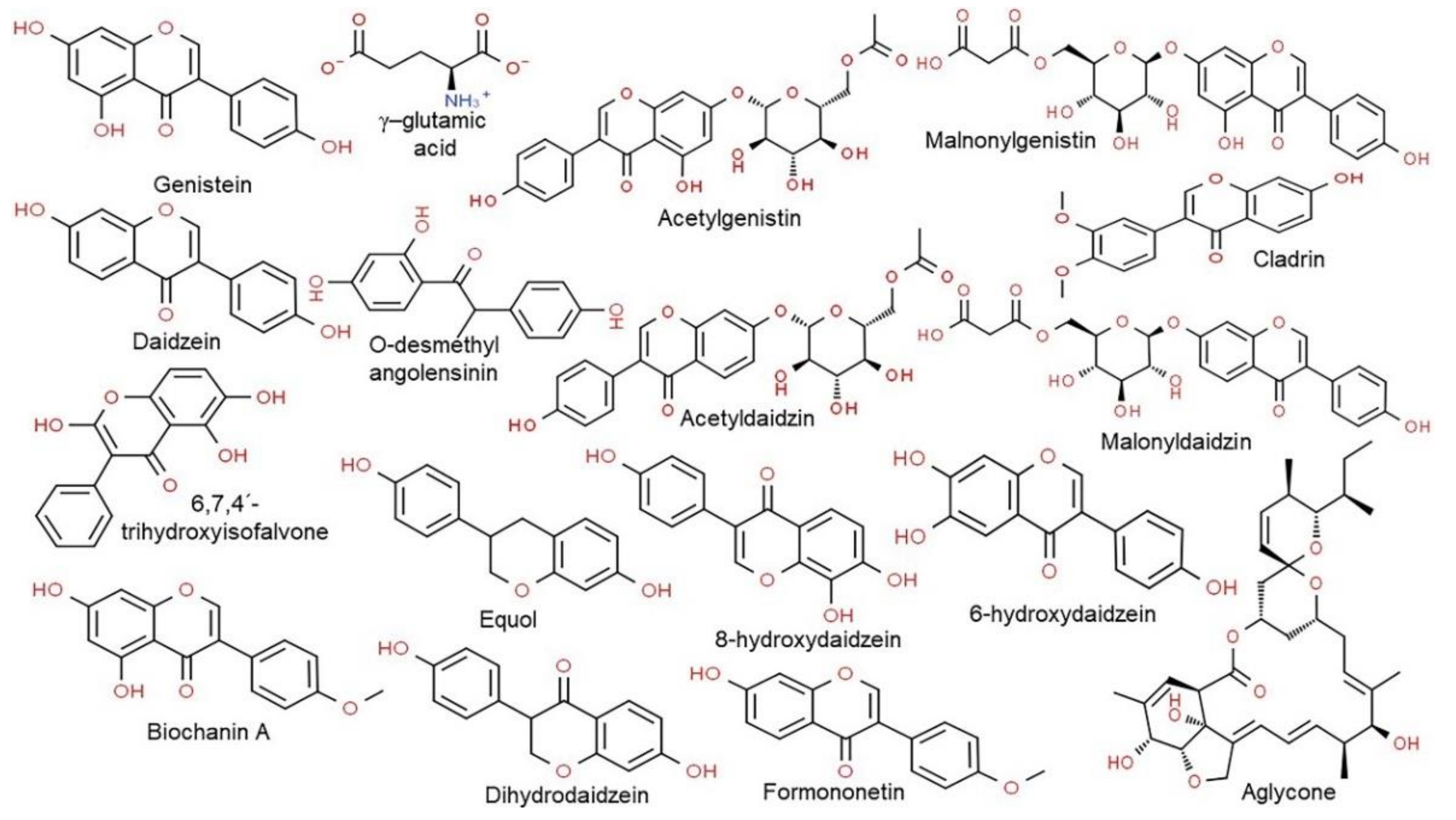
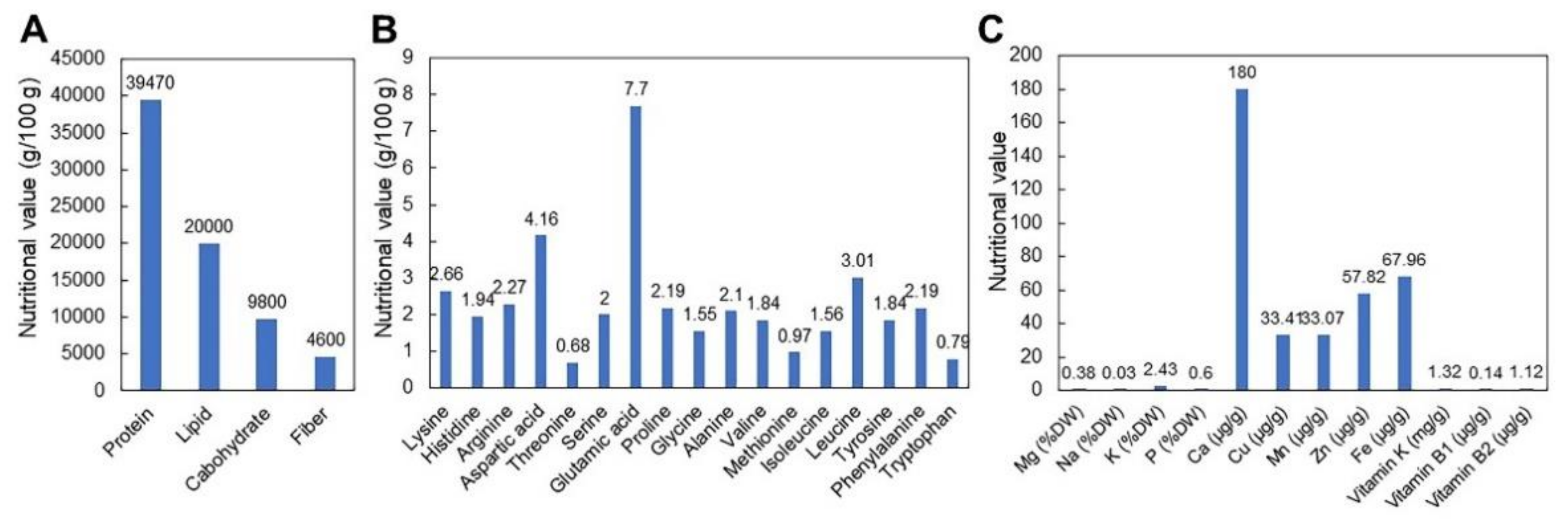
2. Nutritional Properties of CGJ
3. Bioactivities and Human Health Benefits of CGJ
3.1. Thrombotic Effects
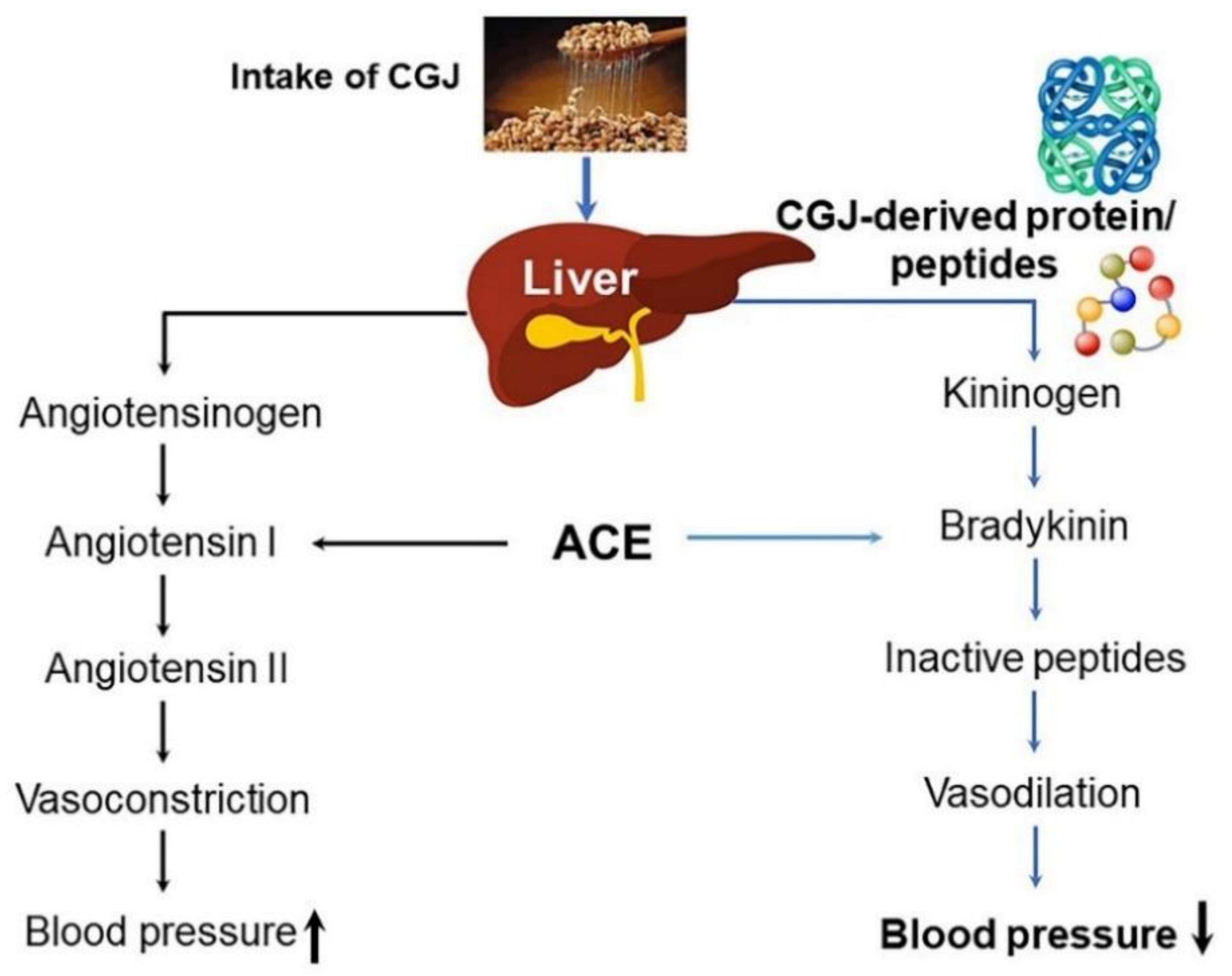
3.2. Blood Pressure-Lowering and Lipid-Lowering Effects
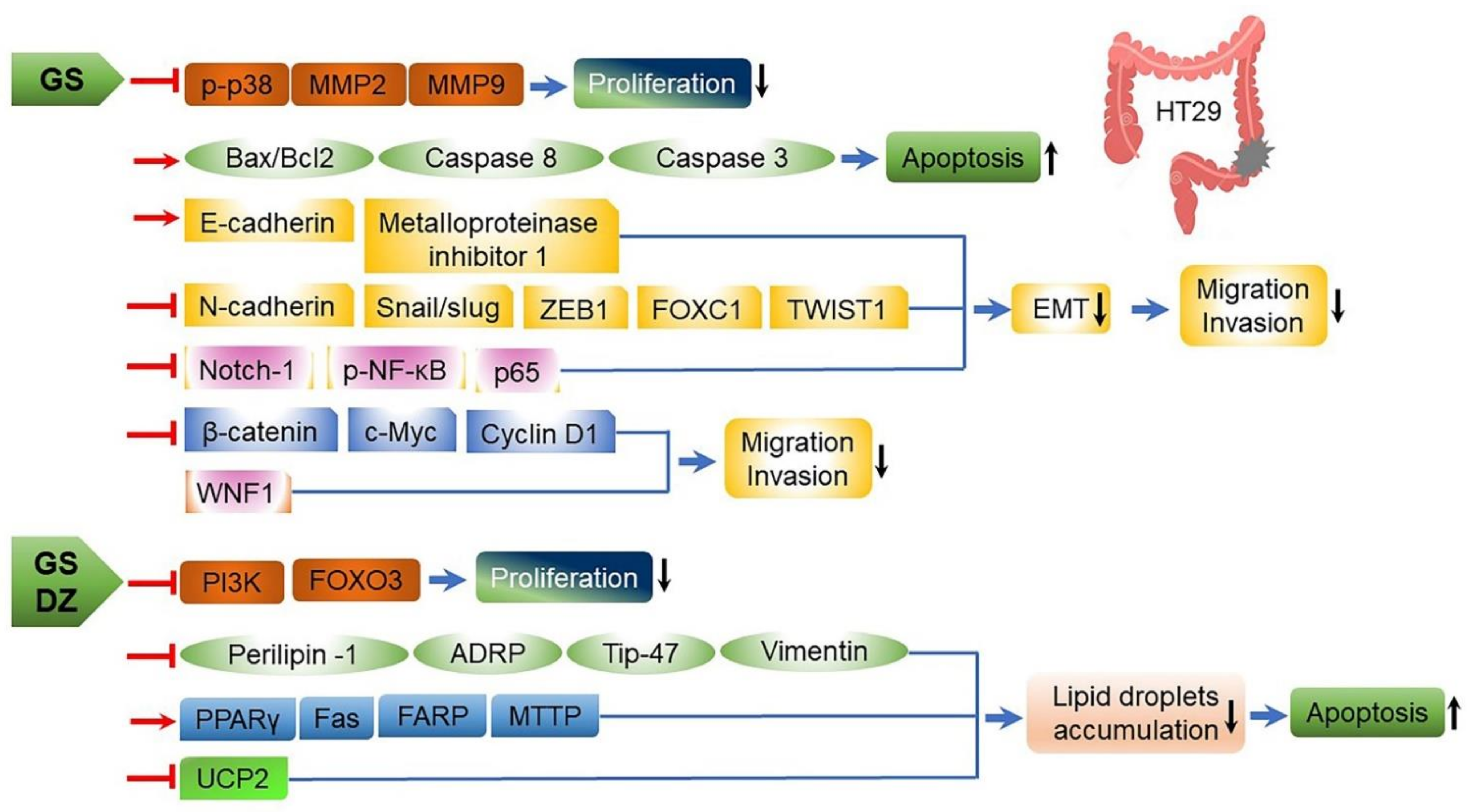
3.3. Anticancer Effects
3.4. Antioxidant Effects
3.5. Immunostimulatory Effects
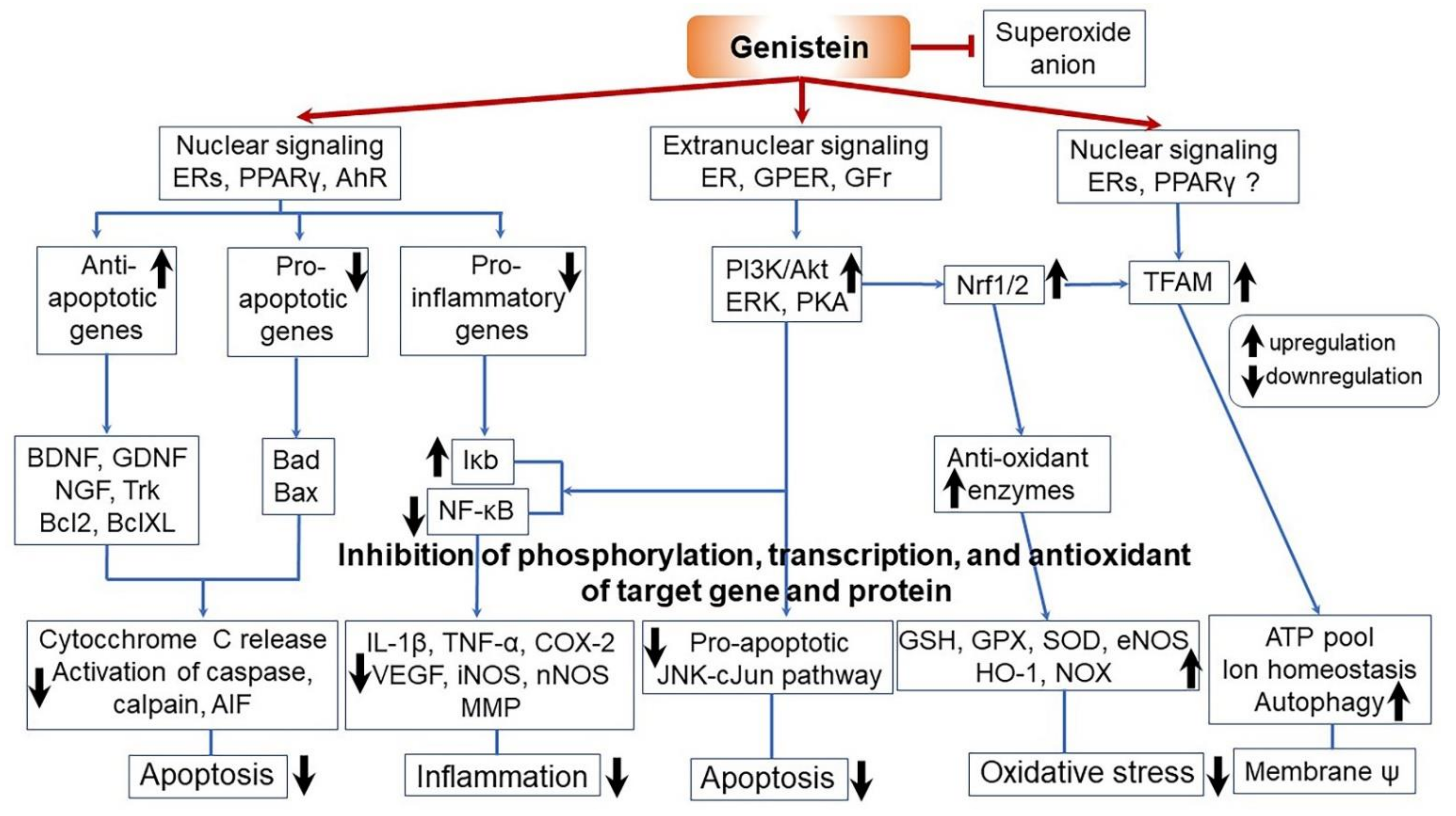
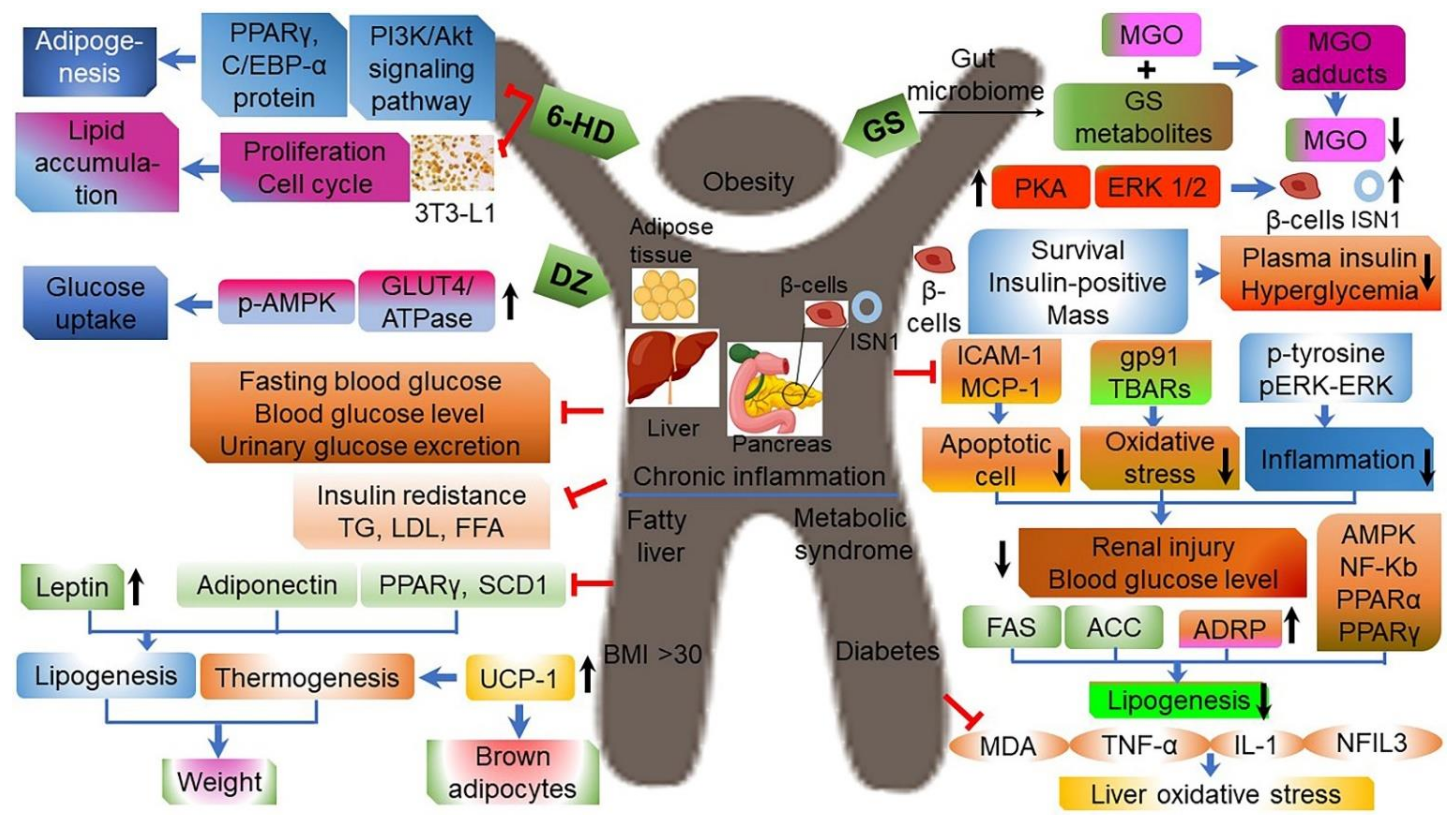
3.6. Anti-Obesity and Anti-Diabetic Effects
3.7. Anti-Osteoporotic Effect and Cognitive Functional Enhancement
3.8. Skin Improvement
3.9. Antimicrobial Activity and Improved Probiotic Efficacy
3.10. Other Effects and Functionality
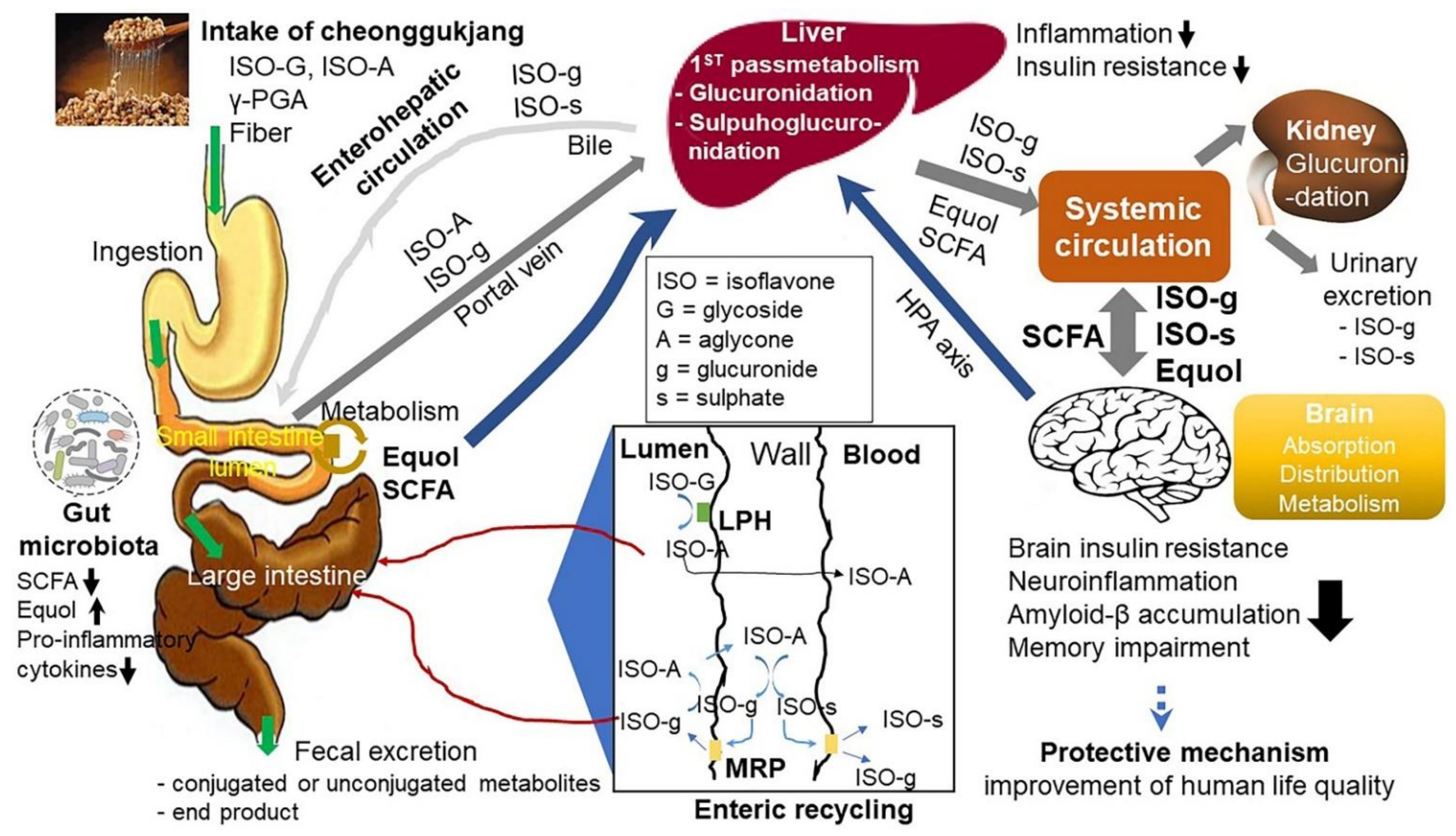
4. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Lee, B.Y.; Kim, D.N.; Kim, K.H. Physico-chemical properties of viscous substance extracted from chungkook-jang. Korean J. Food Sci. Technol. 1991, 23, 599–604. [Google Scholar]
- Lee, J.O.; Ha, S.D.; Kim, A.J.; Yuh, C.S.; Bang, I.S.; Park, S.H. Industrial application and physiological functions of chongkukjang. Food Sci. Ind. 2005, 38, 69–78. [Google Scholar]
- Jeong, D.Y.; Ryu, M.S.; Yang, H.J.; Park, S. Gamma-PGA-rich chungkookjang, short-term fermented soybeans: Prevents memory impairment by modulating brain insulin sensitivity, neuro-inflammation, and the gut-microbiome-brain axis. Foods 2021, 10, 221. [Google Scholar] [CrossRef] [PubMed]
- Jang, C.H.; Oh, J.; Lim, J.S.; Kim, H.J.; Kim, J.S. Fermented soy products: Beneficial potential in neurodegenerative diseases. Foods 2021, 10, 636. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, X.; Hao, L.; Zhang, G.; Jin, Z.; Li, C.; Yang, Y.; Rao, J.; Chen, B. Traditional fermented soybean products: Processing, flavor formation, nutritional and biological activities. Crit. Rev. Food Sci. Nutr. 2020, 1–19. [Google Scholar] [CrossRef]
- Tamang, J.P.; Shin, D.H.; Jung, S.J.; Chae, S.W. Functional properties of microorganisms in fermented foods. Front. Microbiol. 2016, 7, 578. [Google Scholar] [CrossRef] [Green Version]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligne, B.; Ganzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Sanlier, N.; Gokcen, B.B.; Sezgin, A.C. Health benefits of fermented foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef]
- Hsiao, Y.H.; Ho, C.T.; Pan, M.H. Bioavailability and health benefits of major isoflavone aglycones and their metabolites. J. Funct. Foods 2020, 74, 104164. [Google Scholar] [CrossRef]
- Mun, E.G.; Kim, B.; Kim, E.Y.; Lee, H.J.; Kim, Y.; Park, Y.; Cha, Y.S. Research trend in traditional fermented foods focused on health functional evaluation. J. Korean Soc. Food Sci. Nutr. 2018, 47, 373–386. [Google Scholar] [CrossRef]
- Nan, H.M.; Park, J.W.; Song, Y.J.; Yun, H.Y.; Park, J.S.; Hyun, T.; Youn, S.J.; Kim, Y.D.; Kang, J.W.; Kim, H. Kimchi and soybean pastes are risk factors of gastric cancer. World J. Gastroenterol. 2005, 11, 3175–3181. [Google Scholar] [CrossRef]
- Park, S.; Kim, M.J.; Hong, J.; Kim, H.J.; Yi, S.H.; Lee, M.K. Selection and characterization of Cheonggukjang (fast fermented soybean paste)-originated bacterial strains with a high level of S-adenosyl-L-methionine production and probiotics efficacy. J. Med. Food 2014, 17, 1170–1176. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.; Jeong, D. Korean traditional fermented soybean products: Jang. J. Ethn. Foods 2015, 2, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Takagi, A.; Kano, M.; Kaga, C. Possibility of breast cancer prevention: Use of soy isoflavones and fermented soy beverage produced using probiotics. Int. J. Mol. Sci. 2015, 16, 10907–10920. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhou, L.; Xie, N.; Nice, E.C.; Zhang, T.; Cui, Y.; Huang, C. Overcoming cancer therapeutic bottleneck by drug repurposing. Signal Transduct. Target. Ther. 2020, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in cancer therapy: Anti-cancer effects and mechanisms of action. Cell Biosci. 2017, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Wirngo, F.E.; Lambert, M.N.; Jeppesen, P.B. The physiological effects of dandelion (Taraxacum officinale) in type 2 diabetes. Rev. Diabet. Stud. 2016, 13, 113–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, E.R.; Liu, D. Anti-diabetic functions of soy isoflavone genistein: Mechanisms underlying its effects on pancreatic beta-cell function. Food Funct. 2013, 4, 200–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papachristoforou, E.; Lambadiari, V.; Maratou, E.; Makrilakis, K. Association of glycemic indices (hyperglycemia, glucose variability, and hypoglycemia) with oxidative stress and diabetic complications. J. Diabetes Res. 2020, 2020, 7489795. [Google Scholar] [CrossRef]
- Schreihofer, D.A.; Oppong-Gyebi, A. Genistein: Mechanisms of action for a pleiotropic neuroprotective agent in stroke. Nutr. Neurosci. 2019, 22, 375–391. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.L.; Zhang, H.S.; Zhao, X.S.; Xue, H.H.; Xue, J.; Sun, Y.H. Composition, distribution, and antioxidant activity of phenolic compounds in 18 soybean cultivars. J. AOAC Int. 2018, 101, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.W.; Shahzad, R.; Bilal, S.; Adhikari, B.; Kim, I.D.; Lee, J.D.; Lee, I.J.; Kim, B.O.; Shin, D.H. Comparison of antioxidants potential, metabolites, and nutritional profiles of Korean fermented soybean (Cheonggukjang) with Bacillus subtilis KCTC 13241. J. Food Sci. Technol. 2018, 55, 2871–2880. [Google Scholar] [CrossRef] [PubMed]
- Mallikarjun Gouda, K.G.; Gowda, L.R.; Rao, A.G.; Prakash, V. Angiotensin I-converting enzyme inhibitory peptide derived from glycinin, the 11S globulin of soybean (Glycine max). J. Agric. Food Chem. 2006, 54, 4568–4573. [Google Scholar] [CrossRef] [PubMed]
- Serfozo, P.; Wysocki, J.; Gulua, G.; Schulze, A.; Ye, M.; Liu, P.; Jin, J.; Bader, M.; Myohanen, T.; Garcia-Horsman, J.A.; et al. Ang II (angiotensin II) conversion to angiotensin-(1-7) in the circulation is POP (prolyloligopeptidase)-dependent and ACE2 (angiotensin-converting enzyme 2)-independent. Hypertension 2020, 75, 173–182. [Google Scholar] [CrossRef]
- Mukherjee, R.; Chakraborty, R.; Dutta, A. Role of fermentation in improving nutritional quality of soybean meal—A review. Asian-Australas. J. Anim. Sci. 2016, 29, 1523–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashaolu, T.J.; Reale, A. A holistic review on Euro-Asian lactic acid bacteria fermented cereals and vegetables. Microorganisms 2020, 8, 1176. [Google Scholar] [CrossRef] [PubMed]
- Magnuson, B.; Munro, I.; Abbot, P.; Baldwin, N.; Lopez-Garcia, R.; Ly, K.; McGirr, L.; Roberts, A.; Socolovsky, S. Review of the regulation and safety assessment of food substances in various countries and jurisdictions. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess 2013, 30, 1147–1220. [Google Scholar] [CrossRef]
- Xiang, H.; Sun-Waterhouse, D.; Waterhouse, G.I.N.; Cui, C.; Ruan, Z. Fermentation-enabled wellness foods: A fresh perspective. Food Sci. Hum. Well. 2019, 8, 203–243. [Google Scholar] [CrossRef]
- Shahzad, R.; Shehzad, A.; Bilal, S.; Lee, I.J. Bacillus amyloliquefaciens RWL-1 as a new potential strain for augmenting biochemical and nutritional composition of fermented soybean. Molecules 2020, 25, 2346. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Garg, P.; Kumar, P.; Bhatia, S.K.; Kulshrestha, S. Microbial fermentation and its role in quality improvement of fermented foods. Fermentation 2020, 6, 106. [Google Scholar] [CrossRef]
- Chatterjee, C.; Gleddie, S.; Xiao, C.W. Soybean bioactive peptides and their functional properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceusters, N.; Frans, M.; Van den Ende, W.; Ceusters, J. Maltose processing and not beta-amylase activity curtails hydrolytic starch degradation in the CAM orchid phalaenopsis. Front. Plant Sci. 2019, 10, 1386. [Google Scholar] [CrossRef]
- Wu, W.J.; Lee, H.Y.; Lee, G.H.; Chae, H.J.; Ahn, B.Y. The antiosteoporotic effects of Cheonggukjang containing vitamin K2 (menaquinone-7) in ovariectomized rats. J. Med. Food 2014, 17, 1298–1305. [Google Scholar] [CrossRef]
- Wu, W.J.; Gao, H.; Jin, J.S.; Ahn, B.Y. A comparatively study of menaquinone-7 isolated from Cheonggukjang with vitamin K1 and menaquinone-4 on osteoblastic cells differentiation and mineralization. Food Chem. Toxicol. 2019, 131, 110540. [Google Scholar] [CrossRef]
- Pradhananga, M. Effect of processing and soybean cultivar on natto quality using response surface methodology. Food Sci. Nutr. 2019, 7, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Jia, F.; Huang, J. The variation of two extracellular enzymes and soybean meal bitterness during solid-state fermentation of Bacillus subtilis. Grain Oil Sci. Technol. 2019, 2, 39–43. [Google Scholar] [CrossRef]
- Zhao, X.; Song, J.L.; Wang, Q.; Qian, Y.; Li, G.J.; Pang, L. Comparisons of Shuidouchi, Natto, and Cheonggukjang in their physicochemical properties, and antimutagenic and anticancer effects. Food Sci. Biotechnol. 2013, 22, 1077–1084. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, H.; Nirmala, F.S.; Jung, C.H.; Jang, Y.J.; Ha, T.Y.; Ahn, J. Dry-fermented soybean food (Cheonggukjang) ameliorates senile osteoporosis in the senescence-accelerated mouse prone 6 model. J. Med. Food 2019, 22, 1047–1057. [Google Scholar] [CrossRef]
- Choi, J.H.; Pichiah, P.B.; Kim, M.J.; Cha, Y.S. Cheonggukjang, a soybean paste fermented with B. licheniformis-67 prevents weight gain and improves glycemic control in high fat diet induced obese mice. J. Clin. Biochem. Nutr. 2016, 59, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Soh, J.R.; Shin, D.H.; Kwon, D.Y.; Cha, Y.S. Effect of cheonggukjang supplementation upon hepatic acyl-CoA synthase, carnitine palmitoyltransferase I, acyl-CoA oxidase and uncoupling protein 2 mRNA levels in C57BL/6J mice fed with high fat diet. Genes Nutr. 2008, 2, 365–369. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.H.; Kim, S.Y.; Ko, J.M.; Jeong, D.Y.; Kim, Y.S. Biological activities of cheonggukjang prepared with several soybean cultivars. Food Sci. Biotechnol. 2012, 21, 475–483. [Google Scholar] [CrossRef]
- Na, H.G.; Park, Y.; Kim, M.A.; Lee, J.W.; So, G.; Kim, S.H.; Jang, K.H.; Kim, M.J.; Namkoong, S.; Koo, H.J.; et al. Secondary fermented extract of chaga-cheonggukjang attenuates the effects of obesity and suppresses inflammatory response in the liver and spleen of high-fat diet-induced obese mice. J. Microbiol. Biotechnol. 2019, 29, 739–748. [Google Scholar] [CrossRef]
- Lee, J.Y.; Shim, J.M.; Yao, Z.; Liu, X.; Lee, K.W.; Kim, H.J.; Ham, K.S.; Kim, J.H. Antimicrobial activity of Bacillus amyloliquefaciens EMD17 isolated from cheonggukjang and potential use as a starter for fermented soy foods. Food Sci. Biotechnol. 2016, 25, 525–532. [Google Scholar] [CrossRef]
- Cho, M.J.; Lee, J.Y.; Kim, J.H. Microbial and physiochemical properties of cheonggukjang fermented using Bacillus strains with antibacterial or antifungal activities. Food Sci. Biotechnol. 2014, 23, 1525–1532. [Google Scholar] [CrossRef]
- Mah, J.H.; Park, Y.K.; Jin, Y.H.; Lee, J.H.; Hwang, H.J. Bacterial production and control of biogenic amines in Asian fermented soybean foods. Foods 2019, 8, 85. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.J.; Kim, A.R.; Lee, H.; Kim, C.H.; Chang, H.C.; Lee, M.Y. Effects of soybean. cheonggukjang and doenjang on serum cholesterol level and weight reduction in rats fed a high-fat/high-cholesterol diet. Korean J. Food Preserv. 2011, 18, 226–235. [Google Scholar] [CrossRef]
- Kim, J.; Choi, J.N.; Choi, J.H.; Cha, Y.S.; Muthaiya, M.J.; Lee, C.H. Effect of fermented soybean product (cheonggukjang) intake on metabolic parameters in mice fed a high-fat diet. Mol. Nutr. Food Res. 2013, 57, 1886–1891. [Google Scholar] [CrossRef]
- Palta, S.; Saroa, R.; Palta, A. Overview of the coagulation system. Indian J. Anaesth. 2014, 58, 515–523. [Google Scholar] [CrossRef]
- Hayiroglu, M.I.; Cinar, T.; Tekkesin, A.I. Fibrinogen and D-dimer variances and anticoagulation recommendations in Covid-19: Current literature review. Rev. Assoc. Med. Bras. (1992) 2020, 66, 842–848. [Google Scholar] [CrossRef]
- Mosesson, M.W. Fibrinogen and fibrin structure and functions. J. Thromb. Haemost. 2005, 3, 1894–1904. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, Z.; Qiu, Y.; Zhang, J.; Wu, H.; Liang, R.; Chen, G.; Qin, G.; Li, Y.; Zou, D. Clinical significance of plasma D-dimer and fibrinogen in digestive cancer: A systematic review and meta-analysis. Eur. J. Surg. Oncol. 2018, 44, 1494–1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisel, J.W. Fibrinogen and fibrin. Adv. Protein Chem. 2005, 70, 247–299. [Google Scholar] [PubMed]
- Kattula, S.; Byrnes, J.R.; Wolberg, A.S. Fibrinogen and fibrin in hemostasis and thrombosis. Arterioscler. Thromb. Vasc. Biol. 2017, 37, e13–e21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rink, C.; Khanna, S. Significance of brain tissue oxygenation and the arachidonic acid cascade in stroke. Antioxid. Redox Signal. 2011, 14, 1889–1903. [Google Scholar] [CrossRef] [Green Version]
- Weiss, N.; Miller, F.; Cazaubon, S.; Couraud, P.O. The blood-brain barrier in brain homeostasis and neurological diseases. Biochim. Biophys. Acta 2009, 1788, 842–857. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Tian, X.; Wang, X. Diagnosis and treatment of cerebral venous thrombosis: A review. Front. Aging Neurosci. 2018, 10, 2. [Google Scholar] [CrossRef]
- Novitzke, J. The basics of brain aneurysms: A guide for patients. J. Vasc. Interv. Neurol. 2008, 1, 89–90. [Google Scholar]
- Chen, Z.; Venkat, P.; Seyfried, D.; Chopp, M.; Yan, T.; Chen, J. Brain-heart interaction: Cardiac complications after stroke. Circ. Res. 2017, 121, 451–468. [Google Scholar] [CrossRef]
- von Dadelszen, P.; Magee, L.A. Preventing deaths due to the hypertensive disorders of pregnancy. Best. Pract. Res. Clin. Obstet. Gynaecol. 2016, 36, 83–102. [Google Scholar] [CrossRef] [Green Version]
- Mayo, B.; Vazquez, L.; Florez, A.B. Equol: A bacterial metabolite from the daidzein isoflavone and its presumed beneficial health effects. Nutrients 2019, 11, 2231. [Google Scholar] [CrossRef] [Green Version]
- Larkin, T.; Price, W.E.; Astheimer, L. The key importance of soy isoflavone bioavailability to understanding health benefits. Crit. Rev. Food Sci. Nutr. 2008, 48, 538–552. [Google Scholar] [CrossRef] [Green Version]
- Ouriel, K.; Welch, E.L.; Shortell, C.K.; Geary, K.; Fiore, W.M.; Cimino, C. Comparison of streptokinase, urokinase, and recombinant tissue plasminogen activator in an in vitro model of venous thrombolysis. J. Vasc. Surg. 1995, 22, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Steinmetzer, T.; Pilgram, O.; Wenzel, B.M.; Wiedemeyer, S.J.A. Fibrinolysis inhibitors: Potential drugs for the treatment and prevention of bleeding. J. Med. Chem. 2020, 63, 1445–1472. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, J.; Chen, J.; Yu, H.; Zheng, Y.; Zhao, J.; Zhu, J. Recent advances in seafood bioactive peptides and their potential for managing osteoporosis. Crit. Rev. Food Sci. Nutr. 2020, 1–17. [Google Scholar] [CrossRef]
- Radnaabazar, C.; Park, C.M.; Kim, J.H.; Cha, J.; Song, Y.S. Fibrinolytic and antiplatelet aggregation properties of a recombinant Cheonggukjang kinase. J. Med. Food 2011, 14, 625–629. [Google Scholar] [CrossRef]
- Cao, Z.H.; Green-Johnson, J.M.; Buckley, N.D.; Lin, Q.Y. Bioactivity of soy-based fermented foods: A review. Biotechnol. Adv. 2019, 37, 223–238. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, Y.; Zhou, T.; Akkaya, M.S.; Wang, L.; Li, S.; Li, X. Biocontrol of Fusarium wilt disease in strawberries using bioorganic fertilizer fortified with Bacillus licheniformis X-1 and Bacillus methylotrophicus Z-1. 3 Biotech 2020, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.A.; Kim, J.H. Isolation of Bacillus amyloliquefaciens strains with antifungal activities from Meju. Prev. Nutr. Food Sci. 2012, 17, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.M.; Lee, A.R.; Lee, K.W.; Park, J.Y.; Chun, J.; Cha, J.; Song, Y.S.; Kim, J.H. Characterization of a 27 kDa fibrinolytic enzyme from Bacillus amyloliquefaciens CH51 isolated from cheonggukjang. J. Microbiol. Biotechnol. 2009, 19, 997–1004. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.J.; Park, J.Y.; Lee, J.Y.; Lee, K.W.; Cho, K.M.; Kim, G.M.; Shin, J.H.; Kim, J.S.; Kim, J.H. Improvement of fibrinolytic activity of Bacillus subtilis 168 by integration of a fibrinolytic gene into the chromosome. J. Microbiol. Biotechnol. 2015, 25, 1863–1870. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.J.; Heo, K.; Park, J.Y.; Lee, K.W.; Park, J.Y.; Joo, S.H.; Kim, J.H. Characterization of AprE176, a fibrinolytic enzyme from Bacillus subtilis HK176. J. Microbiol. Biotechnol. 2015, 25, 89–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S.J.; Kwon, G.H.; Chun, J.; Kim, J.S.; Park, C.S.; Kwon, D.Y.; Kim, J.H. Cloning of fibrinolytic enzyme gene from Bacillus subtilis isolated from Cheonggukjang and its expression in protease-deficient Bacillus subtilis strains. J. Microbiol. Biotechnol. 2007, 17, 1018–1023. [Google Scholar]
- Kwon, G.H.; Park, J.Y.; Kim, J.S.; Lim, J.; Park, C.S.; Kwon, D.Y.; Kim, J.H. Cloning and expression of a bpr gene encoding Bacillopeptidase F from Bacillus amyloliquefaciens CH86-1. J. Microbiol. Biotechnol. 2011, 21, 515–518. [Google Scholar] [CrossRef]
- Heo, K.; Cho, K.M.; Lee, C.K.; Kim, G.M.; Shin, J.H.; Kim, J.S.; Kim, J.H. Characterization of a fibrinolytic enzyme secreted by Bacillus amyloliquefaciens CB1 and its gene cloning. J. Microbiol. Biotechnol. 2013, 23, 974–983. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Hwang, N.; Ko, G.; Tatsuya, U. Effects of digested Cheonggukjang on human microbiota assessed by in vitro fecal fermentation. J. Microbiol. 2021, 59, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Simonneau, G.; Montani, D.; Celermajer, D.S.; Denton, C.P.; Gatzoulis, M.A.; Krowka, M.; Williams, P.G.; Souza, R. Haemodynamic definitions and updated clinical classification of pulmonary hypertension. Eur. Respir. J. 2019, 53, 1801913. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Jaramillo, P.; Lopez-Lopez, J.; Lopez-Lopez, C.; Rodriguez-Alvarez, M.I. The goal of blood pressure in the hypertensive patient with diabetes is defined: Now the challenge is go from recommendations to practice. Diabetol. Metab. Syndr. 2014, 6, 31. [Google Scholar]
- Oparil, S.; Acelajado, M.C.; Bakris, G.L.; Berlowitz, D.R.; Cifkova, R.; Dominiczak, A.F.; Grassi, G.; Jordan, J.; Poulter, N.R.; Rodgers, A.; et al. Hypertension. Nat. Rev. Dis. Primers 2018, 4, 18014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munoz-Durango, N.; Fuentes, C.A.; Castillo, A.E.; Gonzalez-Gomez, L.M.; Vecchiola, A.; Fardella, C.E.; Kalergis, A.M. Role of the renin-angiotensin-aldosterone system beyond blood pressure regulation: Molecular and cellular mechanisms involved in end-organ damage during arterial hypertension. Int. J. Mol. Sci. 2016, 17, 797. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Aoki, S.; Xing, Y.; Sasano, H.; Rainey, W.E. Metastin stimulates aldosterone synthesis in human adrenal cells. Reprod. Sci. 2007, 14, 836–845. [Google Scholar] [CrossRef]
- Chartier, L.; Schiffrin, E.; Thibault, G.; Garcia, R. Atrial natriuretic factor inhibits the stimulation of aldosterone secretion by angiotensin II, ACTH and potassium in vitro and angiotensin II-induced steroidogenesis in vivo. Endocrinology 1984, 115, 2026–2028. [Google Scholar] [CrossRef] [PubMed]
- Wijesekara, I.; Kim, S.K. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: Prospects in the pharmaceutical industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shobako, N.; Ohinata, K. Anti-hypertensive effects of peptides derived from rice bran protein. Nutrients 2020, 12, 3060. [Google Scholar] [CrossRef]
- Zielinski, H.; Honke, J.; Topolska, J.; Baczek, N.; Piskula, M.K.; Wiczkowski, W.; Wronkowska, M. ACE inhibitory properties and phenolics profile of fermented flours and of baked and digested biscuits from buckwheat. Foods 2020, 9, 847. [Google Scholar] [CrossRef]
- Tanimoto, H.; Mori, M.; Motoki, M.; Torii, K.; Kadowaki, M.; Noguchi, T. Natto mucilage containing poly-gamma-glutamic acid increases soluble calcium in the rat small intestine. Biosci. Biotechnol. Biochem. 2001, 65, 516–521. [Google Scholar] [CrossRef]
- Singer, J.W.; De Vries, P.; Bhatt, R.; Tulinsky, J.; Klein, P.; Li, C.; Milas, L.; Lewis, R.A.; Wallace, S. Conjugation of camptothecins to poly-(L-glutamic acid). Ann. N. Y. Acad. Sci. 2000, 92, 136–150. [Google Scholar] [CrossRef]
- Daliri, E.B.; Ofosu, F.K.; Chelliah, R.; Park, M.H.; Kim, J.H.; Oh, D.H. Development of a soy protein hydrolysate with an antihypertensive effect. Int. J. Mol. Sci. 2019, 20, 1496. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, K.; Nakamaru, M.; Higashimori, K.; Inagami, T. Local generation and release of angiotensin II in peripheral vascular tissue. Hypertension 1988, 11, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Luc, G.; Bard, J.M.; Arveiler, D.; Ferrieres, J.; Evans, A.; Amouyel, P.; Fruchart, J.C.; Ducimetiere, P. Lipoprotein (a) as a predictor of coronary heart disease: The PRIME study. Atherosclerosis 2002, 163, 377–384. [Google Scholar] [CrossRef]
- Kim, B.N.; Kim, J.D.; Ham, S.S.; Choi, Y.S.; Lee, S.Y. Effects of spice added natto supplementation on the lipid metabolism in rats. J. Korean Soc. Food Nutr. 1995, 24, 121–126. [Google Scholar]
- Shin, S.K.; Kwon, J.H.; Jeong, Y.J.; Jeon, S.M.; Choi, J.Y.; Choi, M.S. Supplementation of cheonggukjang and red ginseng cheonggukjang can improve plasma lipid profile and fasting blood glucose concentration in subjects with impaired fasting glucose. J. Med. Food 2011, 14, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Iritani, N.; Hosomi, H.; Fukuda, H.; Tada, K.; Ikeda, H. Soybean protein suppresses hepatic lipogenic enzyme gene expression in Wistar fatty rats. J. Nutr. 1996, 126, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Sautier, C.; Doucet, C.; Flament, C.; Lemonnier, D. Effects of soy protein and saponins on serum, tissue and feces steroids in rat. Atherosclerosis 1979, 34, 233–241. [Google Scholar] [CrossRef]
- Rizzo, G.; Baroni, L. Soy, soy foods and their role in vegetarian diets. Nutrients 2018, 10, 43. [Google Scholar] [CrossRef] [Green Version]
- Cheryan, M. Phytic acid interactions in food systems. Crit. Rev. Food Sci. Nutr. 1980, 13, 297–335. [Google Scholar] [CrossRef]
- Pramitha, J.L.; Rana, S.; Aggarwal, P.R.; Ravikesavan, R.; Joel, A.J.; Muthamilarasan, M. Diverse role of phytic acid in plants and approaches to develop low-phytate grains to enhance bioavailability of micronutrients. Adv. Genet. 2021, 107, 89–120. [Google Scholar]
- Wang, X.; Yang, R.; Zhou, Y.; Gu, Z. A comparative transcriptome and proteomics analysis reveals the positive effect of supplementary Ca2+ on soybean sprout yield and nutritional qualities. J. Proteom. 2016, 143, 161–172. [Google Scholar] [CrossRef]
- Singh, R.P.; Agarwal, R. Prostate cancer and inositol hexaphosphate: Efficacy and mechanisms. Anticancer Res. 2005, 25, 2891–2903. [Google Scholar]
- Marolt, G.; Gricar, E.; Pihlar, B.; Kolar, M. Complex formation of phytic acid with selected monovalent and divalent metals. Front. Chem. 2020, 8, 582746. [Google Scholar] [CrossRef]
- Sanchez-Chino, X.; Jimenez-Martinez, C.; Davila-Ortiz, G.; Alvarez-Gonzalez, I.; Madrigal-Bujaidar, E. Nutrient and nonnutrient components of legumes, and its chemopreventive activity: A review. Nutr. Cancer 2015, 67, 401–410. [Google Scholar] [CrossRef]
- Xu, B.; Chang, S.K. Reduction of antiproliferative capacities, cell-based antioxidant capacities and phytochemical contents of common beans and soybeans upon thermal processing. Food Chem. 2011, 129, 974–981. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Jeong, M.K.; Kim, M.; Lee, J. Modification of isoflavone profiles in a fermented soy food with almond powder. J. Food Sci. 2012, 77, C128–C134. [Google Scholar] [CrossRef]
- Park, C.U.; Jeong, M.K.; Park, M.H.; Yeu, J.; Park, M.S.; Kim, M.J.; Ahn, S.M.; Chang, P.S.; Lee, J. Formation of succinyl genistin and succinyl daidzin by Bacillus species. J. Food Sci. 2010, 75, C128–C133. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, J.H.; Surh, J.; Kang, S.A.; Jang, K.H. Aglycone isoflavones and exopolysaccharides produced by Lactobacillus acidophilus in fermented soybean paste. Prev. Nutr. Food Sci. 2016, 21, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Jeong, P.H.; Shin, D.H.; Kim, Y.S. Effects of germination and osmopriming treatment on enhancement of isoflavone contents in various soybean cultivars and cheonggukjang (fermented unsalted soybean paste). J. Food Sci. 2008, 73, H187–H194. [Google Scholar] [CrossRef]
- Kim, M.J.; Lee, D.H.; Ahn, J.; Jang, Y.J.; Ha, T.Y.; Do, E.; Jung, C.H. Nutrikinetic study of fermented soybean paste (Cheonggukjang) isoflavones according to the Sasang typology. Nutr. Res. Pract. 2020, 14, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Song, D.Y.; Reddy, L.V.; Charalampopoulos, D.; Wee, Y.J. Poly-(gamma-glutamic acid) production and optimization from agro-industrial bioresources as renewable substrates by Bacillus sp. FBL-2 through response surface methodology. Biomolecules 2019, 9, 754. [Google Scholar] [CrossRef] [Green Version]
- Shin, E.J.; Sung, M.J.; Park, J.H.; Yang, H.J.; Kim, M.S.; Hur, H.J.; Hwang, J.T. Poly-gamma-glutamic acid induces apoptosis via reduction of COX-2 expression in TPA-induced HT-29 human colorectal cancer cells. Int. J. Mol. Sci. 2015, 16, 7577–7586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.J.; Park, K.B.; Kim, S.G.; Oh, S.H. Expression, purification, and biochemical properties of arginase from Bacillus subtilis 168. J. Microbiol. 2013, 51, 222–228. [Google Scholar] [CrossRef]
- Pan, L.; Chai, H.; Kinghorn, A.D. The continuing search for antitumor agents from higher plants. Phytochem. Lett. 2010, 3, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moy, Y.S.; Lai, Y.J.; Chou, C.C. Effects of ripening process on the mutagenicity and antimutagenicity of Sufu, a Chinese traditional fermented product of soybean. Food Bioproc. Technol. 2011, 5, 2972–2977. [Google Scholar] [CrossRef]
- Kim, S.Y.; Shon, Y.H.; Lee, J.S.; Kim, C.H.; Nam, K.S. Antimutagenic activity of soybeans fermented with basidiomycetes in Ames/Salmonella test. Biotechnol. Lett. 2000, 22, 1197–1202. [Google Scholar] [CrossRef]
- Piao, Y.Z.; Eun, J.B. Physicochemical characteristics and isoflavones content during manufacture of short-time fermented soybean product (cheonggukjang). J. Food Sci. Technol. 2020, 57, 2190–2197. [Google Scholar] [CrossRef]
- Kim, J.; Choi, J.N.; Kang, D.; Son, G.H.; Kim, Y.S.; Choi, H.K.; Kwon, D.Y.; Lee, C.H. Correlation between antioxidative activities and metabolite changes during Cheonggukjang fermentation. Biosci. Biotechnol. Biochem. 2011, 75, 732–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.J.; Lee, J.M.; Kim, S.; Kim, H.J. Simultaneous analysis and measurement of uncertainty estimation of six isoflavones in Cheonggukjang by liquid chromatography-electrospray tandem mass spectrometry. Food Chem. 2019, 289, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.S.; Lee, S.; Lee, S.H.; Kim, H.J.; Lee, C.H. Comparative evaluation of six traditional fermented soybean products in East Asia: A metabolomics approach. Metabolites 2019, 9, 183. [Google Scholar] [CrossRef] [Green Version]
- Mukund, V.; Mukund, D.; Sharma, V.; Mannarapu, M.; Alam, A. Genistein: Its role in metabolic diseases and cancer. Crit. Rev. Oncol. Hematol. 2017, 119, 13–22. [Google Scholar] [CrossRef]
- Choi, J.; Kwon, S.H.; Park, K.Y.; Yu, B.P.; Kim, N.D.; Jung, J.H.; Chung, H.Y. The anti-inflammatory action of fermented soybean products in kidney of high-fat-fed rats. J. Med. Food 2011, 14, 232–239. [Google Scholar] [CrossRef]
- Woo, J.E.; Seo, J.Y.; Kim, J.H.; Shin, J.H.; Cho, K.M.; Kim, J.S. Soy products fermented with sprouted garlic have increased neuroprotective activities and restore cognitive functions. Food Sci. Biotechnol. 2016, 25, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.W.; Kim, I.D.; Bilal, S.; Shahzad, R.; Saeed, M.T.; Adhikari, B.; Nabi, R.B.S.; Kyo, J.R.; Shin, D.H. Effects of bacterial fermentation on the biochemical constituents and antioxidant potential of fermented and unfermented soybeans using probiotic Bacillus subtilis (KCTC 13241). Molecules 2017, 22, 2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Hwang, C.E.; Lee, C.K.; Lee, J.H.; Kim, G.M.; Jeong, S.H.; Shin, J.H.; Kim, J.S.; Cho, K.M. Characteristics and antioxidant effect of garlic in the fermentation of Cheonggukjang by Bacillus amyloliquefaciens MJ1-4. J. Microbiol. Biotechnol. 2014, 24, 959–968. [Google Scholar] [CrossRef]
- Jin, Y.H.; Jeon, A.R.; Mah, J.H. Tyrosinase inhibitory activity of soybeans fermented with Bacillus subtilis capable of producing a phenolic glycoside, arbutin. Antioxidants 2020, 9, 1301. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Ji, J.; Baek, S.H.; Lee, J.H.; Ha, I.J.; Lim, S.S.; Yoon, H.J.; Nam, Y.J.; Ahn, K.S. Fermented dried Citrus unshiu peel extracts exert anti-inflammatory activities in LPS-induced RAW264.7 macrophages and improve skin moisturizing efficacy in immortalized human HaCaT keratinocytes. Pharm. Biol. 2019, 57, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Cho, C.W.; Han, C.J.; Rhee, Y.K.; Lee, Y.C.; Shin, K.S.; Shin, J.S.; Lee, K.T.; Hong, H.D. Cheonggukjang polysaccharides enhance immune activities and prevent cyclophosphamide-induced immunosuppression. Int. J. Biol. Macromol. 2015, 72, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Rim, H.K.; Jung, J.Y.; An, H.J.; Shin, J.S.; Cho, C.W.; Rhee, Y.K.; Hong, H.D.; Lee, K.T. Immunostimulatory activity of polysaccharides from Cheonggukjang. Food Chem. Toxicol. 2013, 59, 476–484. [Google Scholar] [CrossRef]
- Shin, H.S.; Shon, D.H. Food and natural materials target mechanisms to effectively regulate allergic responses. J. Nutr. Sci. Vitaminol. 2015, 61, S109–S111. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Paek, S.H.; Shin, H.W.; Lee, S.Y.; Moon, B.S.; Park, J.E.; Lim, G.D.; Kim, C.Y.; Heo, Y. Effect of fermented soybean products intake on the overall immune safety and function in mice. J. Vet. Sci. 2017, 18, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.H.; Chung, M.J.; Jeong, D.Y.; Oh, D.H. Immunostimulatory activity of isoflavone-glycosides and ethanol extract from a fermented soybean product in human primary immune cells. J. Med. Food 2014, 17, 1113–1121. [Google Scholar] [CrossRef]
- Hwang, J.S.; Yoo, H.J.; Song, H.J.; Kim, K.K.; Chun, Y.J.; Matsui, T.; Kim, H.B. Inflammation-related signaling pathways implicating TGFβ are revealed in the expression profiling of MCF7 cell treated with fermented soybean, chungkookjang. Nutr. Cancer 2011, 63, 645–652. [Google Scholar] [CrossRef]
- Lee, Y.J.; Kim, J.E.; Kwak, M.H.; Go, J.; Son, H.J.; Kim, D.S.; Kang, B.C.; Lee, H.S.; Hwang, D.Y. Toxicity of fermented soybean product (cheonggukjang) manufactured by mixed culture of Bacillus subtilis MC31 and Lactobacillus sakei 383 on liver and kidney of ICR mice. Lab. Anim. Res. 2014, 30, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.J.; Jeong, Y.J.; Kwon, J.H.; Moon, K.D.; Kim, H.J.; Jeon, S.M.; Lee, M.K.; Park, Y.B.; Choi, M.S. Beneficial effect of chungkukjang on regulating blood glucose and pancreatic beta-cell functions in C75BL/KsJ-db/db mice. J. Med. Food 2008, 11, 215–223. [Google Scholar] [CrossRef]
- Lee, S.Y.; Park, S.L.; Hwang, J.T.; Yi, S.H.; Nam, Y.D.; Lim, S.I. Antidiabetic effect of Morinda citrifolia (Noni) fermented by cheonggukjang in KK-A(y) diabetic mice. Evid. Based Complement. Alternat. Med. 2012, 2012, 163280. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Hwang, J.T.; Kim, M.J.; Yang, H.J.; Sung, M.J.; Kim, S.H.; Park, S.; Gu, E.J.; Park, Y.; Kwon, D.Y. The inhibitory effect of saponin derived from Cheonggukjang on adipocyte differentiation In vitro. Food Sci. Biotechnol. 2014, 23, 1273–1278. [Google Scholar] [CrossRef]
- Yang, H.J.; Kwon, D.Y.; Moon, N.R.; Kim, M.J.; Kang, H.J.; Jung, D.Y.; Park, S. Soybean fermentation with Bacillus licheniformis increases insulin sensitizing and insulinotropic activity. Food Funct. 2013, 4, 1675–1684. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.J.; Kim, H.J.; Kim, M.J.; Kang, S.; Kim, D.S.; Daily, J.W.; Jeong, D.Y.; Kwon, D.Y.; Park, S. Standardized chungkookjang, short-term fermented soybeans with Bacillus lichemiformis, improves glucose homeostasis as much as traditionally made chungkookjang in diabetic rats. J. Clin. Biochem. Nutr. 2013, 52, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, N.Y.; Song, E.J.; Kwon, D.Y.; Kim, H.P.; Heo, M.Y. Antioxidant and antigenotoxic activities of Korean fermented soybean. Food Chem. Toxicol. 2008, 46, 1184–1189. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.Y.; Hong, S.M.; Lee, J.E.; Sung, S.R.; Park, S. Long-term consumption of fermented soybean-derived Chungkookjang attenuates hepatic insulin resistance in 90% pancreatectomized diabetic rats. Horm. Metab. Res. 2007, 39, 752–757. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.W.; Cho, S.H.; Kang, S.H.; Park, Y.B.; Yoon, M.H.; Lee, J.B.; No, W.S.; Kim, J.B. Prevalence, genetic diversity, and antibiotic resistance of Bacillus cereus isolated from Korean fermented soybean products. J. Food Sci. 2015, 80, M123–M128. [Google Scholar] [CrossRef]
- Park, H.S.; Shin, S.R.; Hong, J.Y.; Yang, K.M. Comparison of the antioxidant activities of small-black-bean-Chungkukjang-added black food and soybean Chungkukjang extracts. Korean J. Food Preserv. 2013, 20, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Kim, D.S.; Kang, S.; Moon, B.R. Fermented soybeans, Chungkookjang, prevent hippocampal cell death and β-cell apoptosis by decreasing pro-inflammatory cytokines in gerbils with transient artery occlusion. Exp. Biol. Med. 2016, 241, 296–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Back, H.I.; Kim, S.R.; Yang, J.A.; Kim, M.G.; Chae, S.W.; Cha, Y.S. Effects of Chungkookjang supplementation on obesity and atherosclerotic indices in overweight/obese subjects: A 12-week, randomized, double-blind, placebo-controlled clinical trial. J. Med. Food 2011, 14, 532–537. [Google Scholar] [CrossRef]
- Byun, M.S.; Yu, O.K.; Cha, Y.S.; Park, T.S. Korean traditional Chungkookjang improves body composition, lipid profiles and atherogenic indices in overweight/obese subjects: A double-blind, randomized, crossover, placebo-controlled clinical trial. Eur. J. Clin. Nutr. 2016, 70, 1116–1122. [Google Scholar] [CrossRef]
- Kwon, D.Y.; Jang, J.S.; Hong, S.M.; Lee, J.E.; Sung, S.R.; Park, H.R.; Park, S. Long-term consumption of fermented soybean-derived Chungkookjang enhances insulinotropic action unlike soybeans in 90% pancreatectomized diabetic rats. Eur. J. Nutr. 2007, 46, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Go, J.; Kim, J.E.; Kwak, M.H.; Koh, E.K.; Song, S.H.; Sung, J.E.; Kim, D.S.; Hong, J.T.; Hwang, D.Y. Neuroprotective effects of fermented soybean products (Cheonggukjang) manufactured by mixed culture of Bacillus subtilis MC31 and Lactobacillus sakei 383 on trimethyltin-induced cognitive defects mice. Nutr. Neurosci. 2016, 19, 247–259. [Google Scholar] [CrossRef]
- Jung, Y.J.; Choi, M.J. Effects of Rubus coreanus-Cheonggukjang on bone mineral density and bone mineral content in growing rats. Prev. Nutr. Food Sci. 2015, 20, 260–265. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.S.; Kim, J.E.; Lee, Y.J.; Kwak, M.H.; Lee, H.G.; Kim, H.S.; Lee, H.S.; Hwang, D.Y. Growth sensitivity in the epiphyseal growth plate, liver and muscle of SD rats is significantly enhanced by treatment with a fermented soybean product (cheonggukjang) through stimulation of growth hormone secretion. Mol. Med. Rep. 2014, 9, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.S.; Kim, J.E.; Lee, Y.J.; Kwak, M.H.; Go, J.; Son, H.J.; Kim, D.S.; Hwang, D.Y. Fermented soybean product (Cheonggukjang) improved some attributes of protein and growth hormone measurements in Sprague-Dawley rats. Nutr. Res. 2014, 34, 355–367. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, H.; Nirmala, F.S.; Jung, C.H.; Kim, M.J.; Jang, Y.J.; Ha, T.Y.; Ahn, J. Dihydrodaidzein and 6-hydroxydaidzein mediate the fermentation-induced increase of antiosteoporotic effect of soybeans in ovariectomized mice. FASEB J. 2019, 33, 3252–3263. [Google Scholar] [CrossRef]
- Lee, D.H.; Kim, M.J.; Ahn, J.; Lee, S.H.; Lee, H.; Kim, J.H.; Park, S.H.; Jang, Y.J.; Ha, T.Y.; Jung, C.H. Nutrikinetics of isoflavone metabolites after fermented soybean product (cheonggukjang) ingestion in ovariectomized mice. Mol. Nutr. Food Res. 2017, 61, 1700322. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.J.; Ahn, B.Y. Improved menaquinone (Vitamin K2) production in cheonggukjang by optimization of the fermentation conditions. Food Sci. Biotechnol. 2011, 20, 1585–1591. [Google Scholar] [CrossRef]
- Choi, Y.E.; Kim, Y.J. Effects of soybean diet and back massage on 40’s∼50’s women’s facial skin. J. Korean Soc. Cosmet. 2017, 22, 1049–1060. [Google Scholar]
- Choi, Y.E.; Kim, Y.J. Effects of soybean diet and back massage on 40’s∼50’s women’s body. J. Korean Soc. Cosmet. 2017, 23, 115–125. [Google Scholar]
- Kim, S.C.; Cho, I.J.; Kang, S.M. Impact of ferment premilling Chung-guk diet and back meridian massage on the facial skin of middle-aged women. J. Korean Soc. Cosmet. 2017, 23, 168–178. [Google Scholar]
- Hanif, N.; Al-Shami, A.M.A.; Khalid, K.A.; Hadi, H.A. Plant-based skin lightening agents: A review. J. Phytopharm. 2020, 9, 54–60. [Google Scholar] [CrossRef]
- Qian, W.; Liu, W.; Zhu, D.; Cao, Y.; Tang, A.; Gong, G.; Su, H. Natural skin-whitening compounds for the treatment of melanogenesis (Review). Exp. Ther. Med. 2020, 20, 173–185. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.K.; Yeo, I.C.; Park, J.W.; Kang, B.S.; Hahm, Y.T. Isolation and characterization of a novel analyte from Bacillus subtilis SC-8 antagonistic to Bacillus cereus. J. Biosci. Bioeng. 2010, 110, 298–303. [Google Scholar] [CrossRef]
- Kindoli, S.; Lee, H.A.; Kim, J.H. Properties of Bac W42, a bacteriocin produced by Bacillus subtilis W42 isolated from Cheonggukjang. J. Microbiol. Biotechnol. 2012, 22, 1092–1100. [Google Scholar] [CrossRef]
- Bandara, N.; Jo, J.; Ryu, S.; Kim, K.P. Bacteriophages BCP1-1 and BCP8-2 require divalent cations for efficient control of Bacillus cereus in fermented foods. Food Microbiol. 2012, 31, 9–16. [Google Scholar] [CrossRef]
- Son, G.H.; Kim, J.; Muthaiya, M.J.; Lee, S.; Kim, H.Y.; Lee, C.H. Antimicrobial compounds profile during Cheonggukjang fermentation against Xanthomonas oryzae pv. oryzae (Xoo). J. Microbiol. Biotechnol. 2011, 21, 1147–1150. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.J.; Mah, J.H.; Kim, J.H.; Kim, Y.W.; Hwang, H.J. Reduction of tyramine by addition of Schizandra chinensis Baillon in Cheonggukjang. J. Med. Food 2012, 15, 1109–1115. [Google Scholar] [CrossRef]
- Hong, S.W.; Kim, J.H.; Bae, H.J.; Ham, J.S.; Yoo, J.G.; Chung, K.S.; Oh, M.H. Selection and characterization of broad-spectrum antibacterial substance-producing Lactobacillus curvatus PA40 as a potential probiotic for feed additives. Anim. Sci. J. 2018, 89, 1459–1467. [Google Scholar] [CrossRef]
- Ghosh, K.; Kang, H.S.; Hyun, W.B.; Kim, K.P. High prevalence of Bacillus subtilis-infecting bacteriophages in soybean-based fermented foods and its detrimental effects on the process and quality of Cheonggukjang. Food Microbiol. 2018, 76, 196–203. [Google Scholar] [CrossRef]
- Ghosh, K.; Senevirathne, A.; Kang, H.S.; Hyun, W.B.; Kim, J.E.; Kim, K.P. Complete nucleotide sequence analysis of a novel Bacillus subtilis-infecting bacteriophage BSP10 and its effect on poly-gamma-glutamic acid degradation. Viruses 2018, 10, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, C.; Ju, H.K.; Kim, S.C.; Park, J.H.; Lim, J.; Kwon, S.W.; Lee, J. Determination of bioactive compounds in fermented soybean products using GC/MS and further investigation of correlation of their bioactivities. J. Chromatogr. B 2012, 880, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Cho, B.O.; Shin, J.Y.; Kim, J.S.; Che, D.N.; Kang, H.J.; Jeong, D.Y.; Jang, S.I. Soybean fermented with Bacillus amyloliquefaciens (Cheonggukjang) ameliorates atopic dermatitis-like skin lesion in mice by suppressing infiltration of mast cells and production of IL-31 cytokine. J. Microbiol. Biotechnol. 2019, 29, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Kim, J.E.; Kwak, M.H.; Go, J.; Kim, D.S.; Son, H.J.; Hwang, D.Y. Quantitative evaluation of the therapeutic effect of fermented soybean products containing a high concentration of GABA on phthalic anhydride-induced atopic dermatitis in IL-4/Luc/CNS-1 Tg mice. Int. J. Mol. Med. 2014, 33, 1185–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, D.W.; Lee, H.S.; Shim, M.S.; Yum, K.J.; Seo, J.T. Do kimchi and cheonggukjang probiotics as a functional food improve androgenetic alopecia? A clinical pilot study. World J. Mens Health 2020, 38, 95–102. [Google Scholar] [CrossRef]
- Bae, M.J.; Shin, H.S.; See, H.J.; Chai, O.H.; Shon, D.H. Cheonggukjang ethanol extracts inhibit a murine allergic asthma via suppression of mast cell-dependent anaphylactic reactions. J. Med. Food 2014, 17, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Ratha, P.; Jhon, D.Y. Factors increasing poly-γ-glutamic acid content of cheongguk-jang fermented by Bacillus subtilis 168. Food Sci. Biotechnol. 2019, 28, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.M.; Lim, H.J.; Kim, M.S.; Kim, D.S.; Hwang, C.E.; Nam, S.H.; Joo, O.S.; Lee, B.W.; Kim, J.K.; Shin, E.C. Time course effects of fermentation on fatty acid and volatile compound profiles of Cheonggukjang using new soybean cultivars. J. Food Drug Anal. 2017, 25, 637–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, C.U.; Lee, S.G.; Chung, Y.R.; Cho, I.; Kim, J.H. Cloning of a family 11 xylanase gene from Bacillus amyloliquefaciens CH51 isolated from cheonggukjang. Indian J. Microbiol. 2012, 52, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Na, H.G.; Kim, M.A.; Shin, J.W.; Cha, Y.J.; Park, B.S.; Kim, M.J.; Lee, J.C.; You, S.; Jang, K.H. Improvement of viscous substance production during Cheonggukjang fermentation added with glycine. Food Sci. Biotechnol. 2020, 29, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.G.; Shim, S.M.; Kwon, D.Y.; Choi, H.K.; Lee, C.H.; Kim, Y.S. Metabolite profiling of Cheonggukjang, a fermented soybean paste, inoculated with various Bacillus strains during fermentation. Biosci. Biotechnol. Biochem. 2010, 74, 1860–1868. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Choi, J.N.; John, K.M.; Kusano, M.; Oikawa, A.; Saito, K.; Lee, C.H. GC-TOF-MS- and CE-TOF-MS-based metabolic profiling of cheonggukjang (fast-fermented bean paste) during fermentation and its correlation with metabolic pathways. J. Agric. Food Chem. 2012, 60, 9746–9753. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.K.; Jin, Y.H.; Lee, J.H.; Byun, B.Y.; Lee, J.; Jeong, K.C.; Mah, J.H. The role of Enterococcus faecium as a key producer and fermentation condition as an influencing factor in tyramine accumulation in cheonggukjang. Foods 2020, 9, 915. [Google Scholar] [CrossRef]
- Kang, H.R.; Kim, H.S.; Mah, J.H.; Kim, Y.W.; Hwang, H.J. Tyramine reduction by tyrosine decarboxylase inhibitor in Enterococcus faecium for tyramine controlled cheonggukjang. Food Sci. Biotechnol. 2018, 27, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.; Kim, J.S.; Hong, S.B.; Park, B.Y.; Kim, S.J.; Kwon, S.W. Genetic marker gene, recQ, differentiating Bacillus subtilis and the closely related Bacillus species. FEMS Microbiol. Lett. 2019, 366, fnz172. [Google Scholar] [CrossRef]
- Kim, S.J.; Dunlap, C.A.; Kwon, S.W.; Rooney, A.P. Bacillus glycinifermentans sp. nov., isolated from fermented soybean paste. Int. J. Syst. Evol. Microbiol. 2015, 65, 3586–3590. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B. amyloliquefaciens | B. subtilis | Soybeans | |||
|---|---|---|---|---|---|
| SCGB1 | SRCM 100730 | SRCM 100731 | SCGB 574 | ||
| γ-PGA (cm) | 31 ± 0.86 | 27 ± 100 | 30 ± 0.57 | 55 ± 1.00 | 0 |
| Flavor | D (++) | D (++) | D (++) | D (++) | ND |
| Protease activity (cm) | 2.84 ± 0.04 | 1.94 ± 0.08 | 1.92 ± 0.12 | 2.29 ± 0.04 | 1.76 ± 0.01 |
| Cellulase activity (cm) | 2.08 ± 0.04 | 1.58 ± 0.04 | 1.44 ± 0.04 | 1.95 ± 0.07 | 1.78 ± 0.04 |
| Amylase activity (cm) | 2.84 ± 0.04 | 2.29 ± 0.05 | 2.42 ± 0.11 | 2.29 ± 0.04 | 1.99 ± 0.01 |
| Thrombolytic activity (halo size, cm) | 1.83 ± 0.06 | 3.85 ± 0.02 | 4.07 ± 0.14 | 1.95 ± 0.15 | ND |
| Antibacterial activity | D (+++) | D (+++) | D (+++) | D (+++) | ND |
| Iturin A | D (+) | D (+++) | D (+++) | D (++) | ND |
| Bacillomycin D | D (+++) | D (+++) | D (+++) | D (+++) | ND |
| Model | Strain Used in Fermentation | Action Mode | Reference |
|---|---|---|---|
| Raw soybean | B. amyloliquefaciens MJ1-4 | Repressing the growth of Aspergillus spp. producing aflatoxin B1 and Penicillium spp. producing ochratoxin by producing iturin or actively modified amino acids with 12,000–14,000 Da as a bacteriocin-like substance and another non-proteinaceous compound | [70] |
| Raw soybean | B. amyloliquefancies CH51 | Producing fibrinolytic enzyme AprE51 similar to AprE homolog from B. subtilis | [71] |
| Raw soybean | B. subtilis 168 | Improving fibrinolytic activity by genomic integration (amyE site) of aprE2, aprE176, and aprE179 gene | [72] |
| Raw soybean | B. subtilis CH3-5 | Producing fibrinolytic enzyme AprE2 (29 kDa) | [74] |
| Raw soybean | B. subtilis HK176 | Producing thermostable fibrinolytic enzyme AprE176 | [73] |
| Raw soybean | B. amyloliquefaciens CH86-1 | Producing fibrinolytic enzyme Bpr86-1 | [75] |
| Raw soybean | B. amyloliquefaciens CB1 | Producing fibrinolytic enzyme AprECB1 (28 kDa) as a serine metalloprotease in B. subtilis WB600; exhibiting the highest specificity for N-succinyl-Ala-Ala-Pro-Phe-p-nitroanilide, known as a substrate | [76] |
| Male Sprague–Dawley rats | B. licheniformis ATCC1071 | Activating a plasmin-like protease that degrades fibrin clots, but not plasminogen; inhibiting collagen-induced platelet aggregation | [67] |
| Model | Strain Used in Fermentation | Action Mode | Reference |
|---|---|---|---|
| Adult males and females (25–60 years) | B. subtilis | Lowering concentrations of plasma low-density lipoprotein cholesterol, non-high-density lipoprotein cholesterol, and blood glucose; reducing the ratio of apolipoprotein B to apolipoprotein A-1 | [93] |
| Male mice | B. licheniformis ATCC1071 Bacillus spp. | Lowering total cholesterol and triglyceride levels by controlling lipid metabolism, such as lysophosphatidylcholines and phosphatidylcholines | [49] |
| Model | Strain Used in Fermentation | Action Mode | Reference |
|---|---|---|---|
| Raw soybean | B. subtilis MYCO10001 B. subtilis ATCC 21228 | Enhancing levels of isoflavones (daidzein and genistein), their derivatives (isoflavone-β-glucosides and isoflavone-aglycones), or succinyl derivatives (succinyl-β-daidzin and succinyl-β- genistin), and β-glucosidase | [104,105] |
| Raw soybean | L. acidophilus KCTC 3925 L. rhamnosus KCTC 3929 | Producing aglycone-formed isoflavones and exopolysaccharide | [106] |
| Raw soybean | B. subtilis | Enhancing isoflavone levels by controlling CGJ fermentation conditions, including temperature, time, germination, and osmolarity | [107] |
| Healthy Korean men (age range 21–29 years) | Bacillus sp. | Improving human health function of isoflavones following CGJ ingestion | [108] |
| HT-29 human colorectal cancer cells | Bacillus sp. FBL-2 | Promoting poly-γ-glutamic acid (γ-PGA)-mediated apoptosis inhibiting pro-proliferative functions leading to DNA fragmentation, and inactivation of poly (ADP-ribose) polymerase (PARP), cyclooxygenase (COX-2), and inducible nitric oxide synthase (iNOS); activating 5’-adenosine monophosphate-activated protein kinase (AMPK) | [109,110] |
| Raw soybean | B. subtilis 168 | Producing arginase, an arginine-degrading and ornithine-producing enzyme, used to treat arginine-dependent cancer | [111] |
| Model | Strain Used in Fermentation | Action Mode | Reference |
|---|---|---|---|
| Raw soybean | B. licheniformis KCCM 11053P B. licheniformis 11054P B. amyliquefaciens CH86-1 | Increasing amounts of amino acids, phenolics, flavonoids, saponins, and isoflavones (isoflavone aglycone and isoflavone glucoside); increasing bacterial population associated with CGJ fermentation; enhancing 2,2-diphenyl-1-picrylhydrazyl (DPPH), 2,2-azinobis (3-ethyl-benzothiazoline-6-sulfonic acid (ABST), and ferric reducing/antioxidant power (FRAP) activity | [3,31,115,116,117,118] |
| Raw soybean | B. amyloliquefaciens RWL-1 | Increasing levels of phenolics, isoflavones, amino acids, and minerals | [31] |
| Raw soybean | B. subtilis KCTC 13241 B. amyloliquefaciens MJ1-4 | Increasing levels of total phenolics, isoflavones (isoflavone-malonylglycoside, isoflavone-acetylglycoside, and isoflavone-aglycone), and amino acids; exhibiting highest amounts of palmitic acid, stearic acid, and linoleic acid; elevating DPPH, ABST, FRAP, and superoxide dismutase (SOD)-like activity; augmenting levels of calcium, iron, sodium, and zinc | [24,122,123] |
| Male Sprague–Dawley rats | Bacillus sp. | Inducing antioxidative mechanism by redox homeostasis via inactivation of nuclear factor kappa B (NF-κB) and modulating the expression of genes involved in NF-κB-related inflammation | [3,120] |
| C57BL/6J mice | B. subtilis HCD02 B. amyloliquefaciens EMD17 B. amyloliquefaciens MJ1-4 | Producing total phenolics and flavonoids, including quercetin (quercetin glycoside) Enhancing Nrf2-mediated transcription of gene encoding antioxidant molecules (GSH) and enzymes (i.e., NQQ1, GSH reductase, HO-1, γ-glutamylcysteine ligase) → improving DPPH, FRAP, ABST, and TBARS activity | [3,121] |
| Raw soybean | B. subtilis | Enhancing tyrosinase inhibitory activity through arbutin production | [124] |
| Anti-inflammatory activity | |||
| Mice | L. acidophilusKCTC3925 | Suppressing serum triglyceride levels; increasing serum HDL-C, GOT, GPT, and leptin levels; downregulating expression of COX-2, IL-4, and ICAM1-1 genes | [44] |
| RAW 264.7 macrophage and HaCaT cells | B. subtilis | Inhibiting the expression of lipopolysaccharide-induced iNOS and COX-2 protein; suppressing TNF-α, IL-6, and prostaglandin E2 secretion; inducing the production of hyaluronic acid and the expression of filaggrin and serine palmitoyltransferase | [125] |
| Male Sprague–Dawley rats | Bacillus sp. | Suppressing NF-κB-related activities of genes associated with inflammatory proteins, including iNOS, COX-2, and vascular cell adhesion molecule-1 (VCAM-1) | [3,120] |
| Model | Strain Used in Fermentation | Action Mode | Reference |
|---|---|---|---|
| RAW264.7 macrophage | B. subtilis | Increasing levels of functional polysaccharides Activating mRNA expression of nitric oxide (NO) synthase and tumor necrosis factor-α (TNF-α) by inducing nuclear factor-kappa B (NF-κB) → increased production of NO and TNF-α | [126,127] |
| Mast cells | – | Inhibiting allergen permeation through paracellular diffusion into epithelial cells; suppressing Th2 cells-related cytokine production by modulating Th1/Th2 homeostasis; inhibiting CD4+ T cell differentiation by activating regulatory T cells (Treg); inhibiting degranulation of mast cells | [128] |
| Male C57BL/6 mice | B. amyloliquefaciens CJ3-27 B. amyloliquefaciens CJ1526 B. subtilis CJ1553 | Enhancing T helper type-1-mediated immune response by upregulating production ratio of IFN-γ vs. IL-4 and IgG2a vs. IgG1 in T and B cells; improving enhanced splenic natural killer cell activity → activating humoral and cellular immunity to Th1 response | [129] |
| Human primary immune cells | – | Modulating isoflavone-derived immunostimulatory activity by enhancing cell proliferation, nitrite, and transcriptional activation of TNF-α, IL-6, iNOS, and COX-2 gene | [130] |
| Male BALB/cByJ mice | B. subtilis | Improving lymphocyte proliferation, natural killer cell activity, and white blood cell population through activation of NO and immunostimulatory cytokines (IL-6 and IL-12) | [126] |
| MCF7 cells | B. licheniformis B1 | Decreasing growth of breast cancer MCF7 cells via upregulation of TGFβ1 and SMAD3, downregulation of inflammation-related genes (CSF2, CSF2RA, and CSF3), and differential expression of the genes encoding chemokines (CCL2, CCL3, CCL3L3, CXCL1, and CXCL2) → preventing breast cancer by TGFβ-dependent signaling mechanism and inhibiting inflammation | [131] |
| ICR mice | B. subtilis MC31 L. sakei 383 | Not inducing any specific toxicity in liver and kidney organs ICR at dose 100 mg/kg body weight/day as no observed effect level | [132] |
| Model | Strain Used in Fermentation | Action Mode | Reference |
|---|---|---|---|
| Anti-obesity | |||
| Mice | L. acidophilus KCTC3925 | Suppressing serum triglyceride; increasing serum of high-density lipoprotein cholesterol (HDL-C), glutamic-oxaloacetic transaminase (GOT), glutamic-pyruvic transaminase (GPT), and leptin levels; downregulating expression of COX-2, IL-4, and ICAM1-1 genes | [44] |
| C57BL/6J mice | B. licheniformis-67 Bacillus spp. | Downregulating lipid anabolic gene and upregulating lipid catabolic gene; lowering serum and hepatic lipid profiles, blood glucose, insulin, and leptin levels Lowering triglyceride and cholesterol levels in blood by increasing the expression of CPT-1 encoding carnitine palmitoyltransferase, ACO encoding acyl-CoA oxidase, and UCP2 encoding uncoupling protein 2 associated with fatty acid oxidation in liver | [41,42] |
| Women | – | Increasing the ratio of apolipoprotein B to apolipoprotein A1 in CGJ-supplemented women having a body mass index ≥23 kg/m2; waist-to-hip ratio of ≥0.90 for men and ≥0.85 for women | [143] |
| Men and women between 19 and 29 years of age | Bacillus licheniformis | Improving percentage body fat, lean body mass, waist circumference and waist-to-hip ratio of CGJ-supplemented women | [144] |
| Men:women (mean age, 44.9 ± 3.1 years) | Bacillus spp. | Lowering fasting blood glucose level; improving plasma lipid profile | [93] |
| 3T3-L1 cells | Bacillus spp. | Exhibiting anti-lipogenic effect by suppressing lipid accumulation | [135] |
| Anti-diabetic activity | |||
| KK-Ay/TaJcl mice | Bacillus spp. KCTC11351BP B. subtilis KCTC 11352BP B. sonolensis KCTC 11354BP B. circulans KCTC 11355BP | Enhancing decrease in glycosylated hemoglobin level, improvement in insulin sensitivity, and decrease in serum glycerides and low-density lipoprotein-cholesterol levels Activating peroxisome proliferator-activated receptor-γ (PPARγ); regulating glucose uptake via AMP-activated kinase stimulation | [3,134] |
| C57BL/KsJ-db/db mice | B. subtilis | Improving insulin resistance and hyperglycemia in type 2 diabetic animals that are partly medicated by regulating hepatic glucose enzymes, including glucokinase, glucose-6-phosphatase, and phosphoenolpyruvate carboxykinase), hepatic glycogen content, and insulin sensitivity in peripheral tissues | [133] |
| Type 2 diabetic male rats | B. lichemiformis | Enhancing glucose-stimulated insulin secretion during hyperglycemic clamp through regulation of hepatic insulin signaling pathway (i.e., Akt and AMPK) | [136,137] |
| 3T3-L1 cell | B. subtilis | Stimulating translocation of glucose transporter-4 into the plasma membrane through phosphorylation of insulin receptor substrate-1 and Akt → enhancing glucose utilization via activating insulin signaling and stimulating peroxisome proliferator-activated receptor (PPAR-γ) activity by increasing isoflavonoid aglycones (especially daidzein) in adipocytes Improving glucose-stimulated insulin secretion by increasing levels of small peptides with low polarity in insulinoma cells | [145] |
| Model | Strain Used in Fermentation | Action Mode | Reference |
|---|---|---|---|
| Neuroprotection effect | |||
| ICR mice | B. subtilis MC31 L. sakei 383 | Decreasing the number of dead cells in the granule cell layer of the dentate gyrus; suppressing acetylcholinesterase activity; activating nerve growth factor (NGF) and its receptor signaling pathway, including TrkA high affinity receptor and p75 low affinity receptor; downregulating Bax/Bcl2 and caspase 3 expression; promoting increase in superoxide dismutase activity and decrease in lipid peroxidation | [146] |
| ICR mice | B. subtilis B. licheniformis | Modulating the gut–microbiome–brain–axis, brain insulin sensitivity, and neuroinflammation by producing bioactive peptides, dietary fiber, poly-γ-glutamic acid (γ-PGA), and isoflavone aglycones | [3] |
| C57BL/ 6J mice | B. subtilis HCD02 B. amyloliquefaciens EMD17 B. amyloliquefaciens MJ1-4 | Inhibiting growth of mouse hippocampal HT22 and human neuroblast-like SHSY5Y cells Reducing the frequency of behavioral dysfunction induced by D-galactose; improving cognitive abnormal function | [3,121] |
| Raw soybean | B. subtilis | Enhancing tyrosinase inhibitory activity through arbutin production | [124] |
| Anti-osteoporotic effect | |||
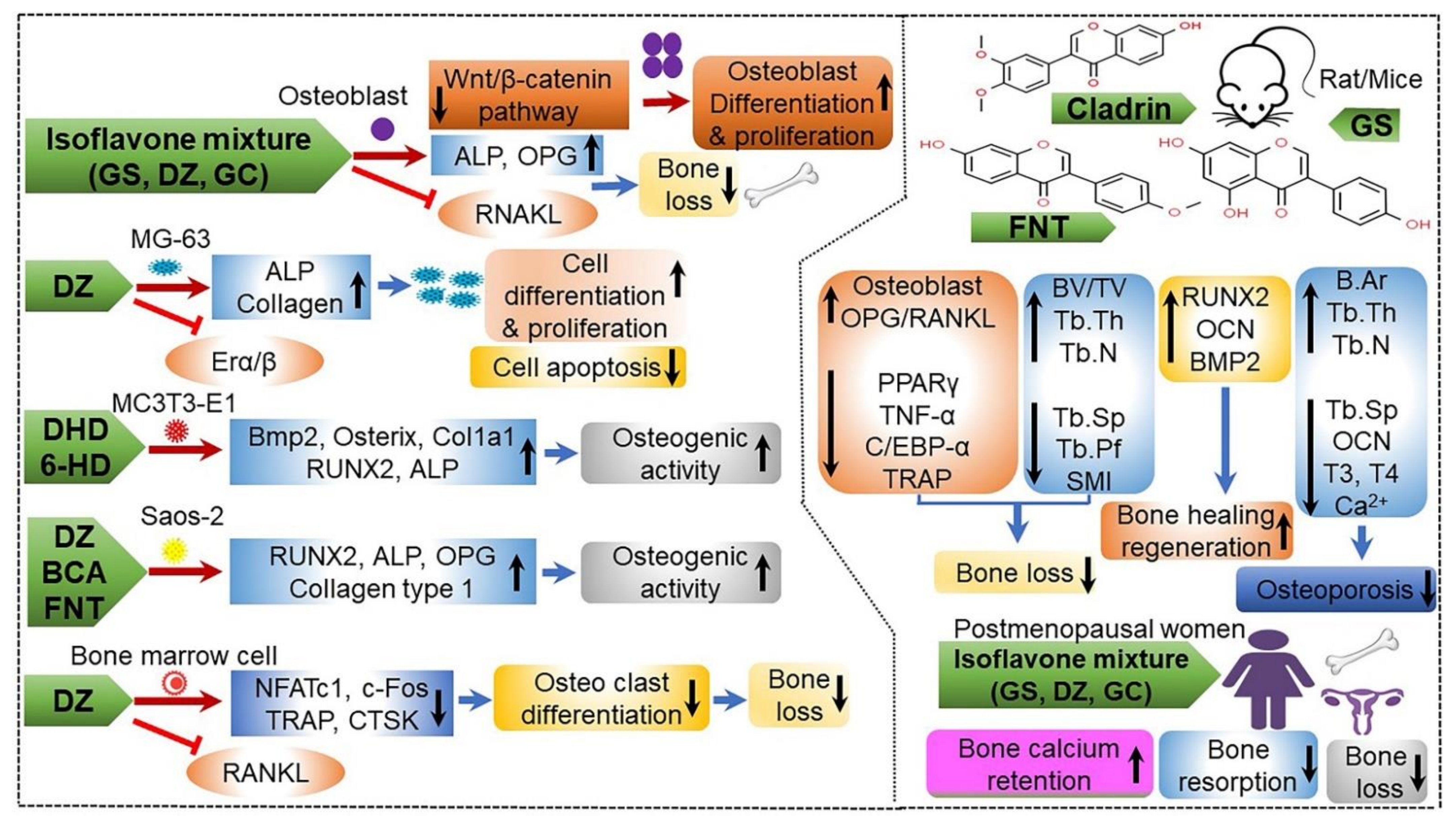
| Senescence-accelerated mouse prone 6 | B. subtilis | Increasing bone mineral density (BMD) and relative bone length; enhancing osteopontin reactivity; upregulating expression of Alp, Col1a1, Fak, Bmp2/4, Smad1/5/8, and Runx2; downregulating expression of Rnakl and Nfatc1; decreasing Cathepsin K level; increasing osteoprotegerin/Rankl ratio | [35,40] |
| MC3T3-E1 subclone 4 (CRL-2593) cells | B. subtilis KCTC 12392BP | Suppressing osteoclast formation and increasing cell proliferation, cellular alkaline phosphatase activity, osteocalcin, and calcium deposition by upregulating the ratio of osteoprotegerin/receptor activator of NF-κB ligand; promoting bone health | [36] |
| Female/male Sprague-Dawley rat | B. subtilis | Preventing bone loss; increasing BMD, bone mineral content (BMC), and trabecular number | [35,147] |
| Female sham-operated and OVX mice | B. subtilis | Enhancing various isoflavone metabolites, including intact isoflavones, 3-hydroxygenistein, genistein 4′-sulfate, and equol 7-glucuronide → promoting bone health of postmenopausal women | [151] |
| Male/female Sprague-Dawley rat | B. subtilis | Accumulating total flavonoids, phenolics, and isoflavones; increasing spine BMD and femur BMC; stimulating growth hormone (GH) secretion, which leads to activation of the GH receptor downstream signaling pathway via induction of phosphorylation of Akt and Erk, but not STAT5 | [147,148,149] |
| C57BL/6J mice | B. amyloliquefaciens KCTC11712BP | Increasing BMD through upregulation of bone morphogenic protein 2 (Bmp2) and osteopontin in bone tissues Increasing isoflavone-mediated osteogenesis via the Bmp2 signaling pathway; reducing receptor activator of NF-κB induced by NF-κB inactivation and MAPK dephosphorylation | [150] |
| Osteoblastic cell line MC3T3-E1 | – | Producing vitamin K1 and K2 Stimulating osteoblast mineralization mechanism by elevating transcriptional ratio of osteoprotegerin and the receptor activator of NF-κB ligand | [36,152] |
| Model | Strain Used in Fermentation | Action Mode | Reference |
|---|---|---|---|
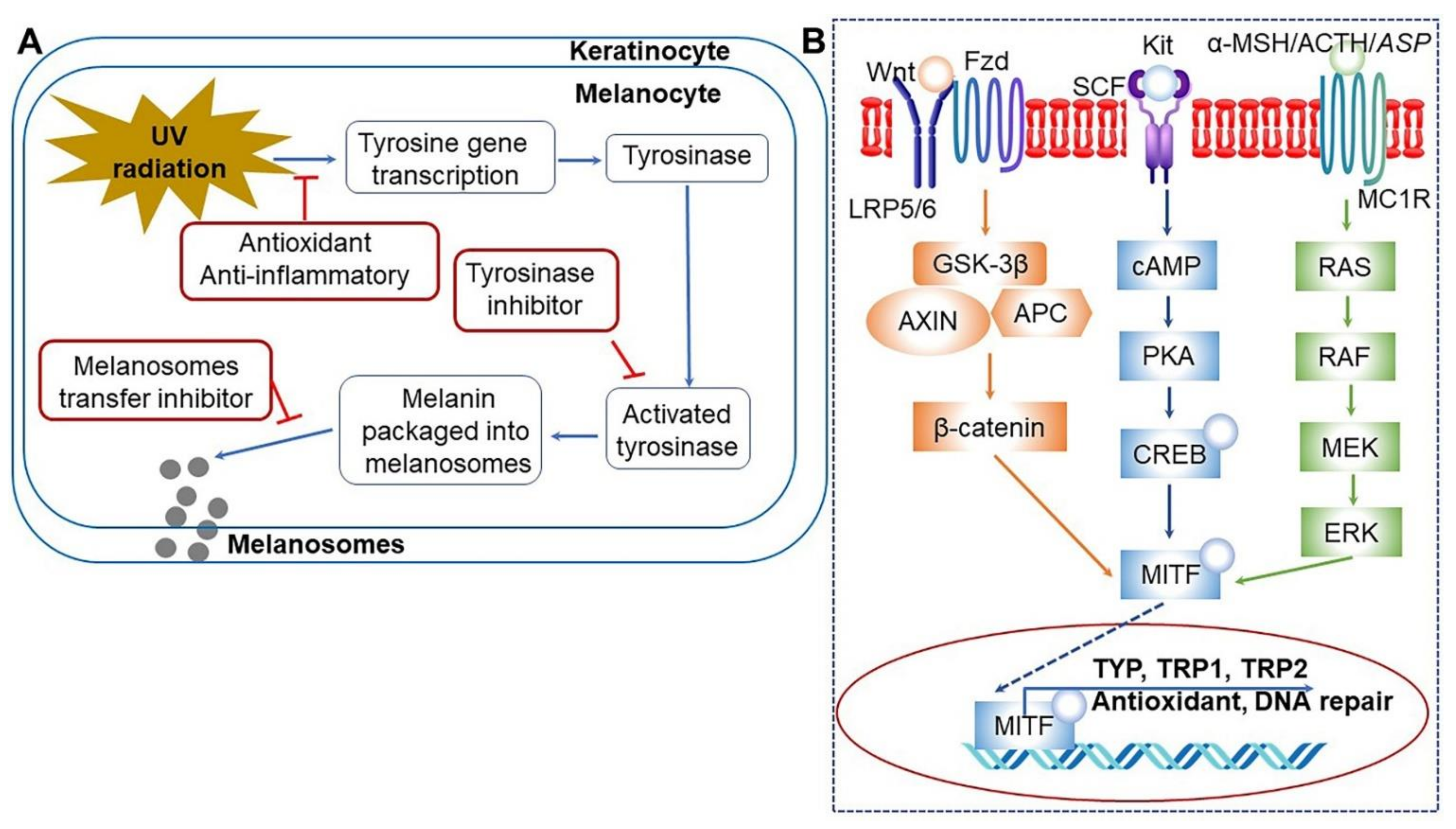
| Korean middle-aged women | Bacillus sp. | Decreasing melanin level and red spots on facial skin; reducing body weight and body fat ratio; increasing muscle mass | [153,154] |
| RAW 264.7 macrophage and HaCaT cell lines | B. subtilis | Anti-dry skin activity: inhibiting the expression of lipopolysaccharide-induced iNOS and COX-2 protein; suppressing TNF-α, IL-6, and prostaglandin E2 secretion; inducing the production of hyaluronic acid and expression of filaggrin and serine palmitoyltransferase | [125,155] |
| Raw soybean | B. subtilis | Skin lightening activity: enhancing tyrosinase inhibitory activity through arbutin production | [124] |
| Model | Strain Used in Fermentation | Action Mode | Reference |
|---|---|---|---|
| Raw soybean | B. amyloliquefaciens EMD17 B. subtilis SC-8 | Inhibiting growth of B. cereus ATCC14579, Listeria monocytogenes ATCC19111, and Penicillium spp. producing an ochratoxin by activating the gene encoding surfactin synthetase A (SrfAA), fengycin, and iturin | [45,158] |
| Raw soybean | B. licheniformis KCCM 11053P | Inhibiting growth of Xanthomonas oryzae pv. Oryzae causing rice bacterial blight by producing daidzein, glycitein, genistein, and surfactins A and B | [161] |
| Raw soybean | B. subtilis W42 | Inhibiting growth of Listeria monocytogenes ATCC 19111 and B. cereus ATCC 14579 by producing Bac W42 (5.4 kDa) as a bacteriocin | [159] |
| Raw soybean | B. subtilis-infecting bacteriophage | Reducing L. monocytogenes infection | [45] |
| Raw soybean | E. faecium | Inhibiting growth of B. subtilis ATCC 15245 by enhancing poly-γ-glutamic acid (γ-PGA) hydrolase activity capable of degrading γ-PGA | [164,165] |
| Raw soybean | B. subtilis SC-8 | Inhibiting growth of B. cereus by producing an antibiotic-like lipopeptidal compound (BSAP-254) as an antagonistic effector | [158] |
| Raw soybean | Bacteriophages BCP1-1 and 8-2 | Inhibiting growth of B. cereus through absorption of divalent cation ions, such as Ca2+, Mg2+, or Mn2+ before monovalent cation (Na+)-mediated post-absorption phase | [160] |
| Raw soybean | E.faecium | Inhibiting bacterial growth by regulating tyramine content | [162] |
| Raw soybean | Lactobacillus curvatus PA40 | Antibacterial activity against B. cereus, Listeria monocytogenes, Escherichia coli O157:H7, and Salmonella enterica | [163] |
| – | E. faecium E. durans, E. sanguinicola | Inhibiting growth of Escherichia coli O157:H7, E. faecalis, Salmonella choleraesuis, Staphylococcus aureus, and Listeria monocytogenes | – |
| Raw soybean | B. amyloliquefaciens MJ1-4 | Repressing growth of Aspergillus spp. producing aflatoxin B1 and Penicillium spp. producing ochratoxin by producing iturin or actively modified amino acid with 12,000–14,000 Da as a bacteriocin-like substance and another non-proteinaceous compound | [70] |
| Raw soybean | B. licheniformis B. subtilis B.amyloliquefaciens | Developing beneficial bacterial strains that overproduce S-adenosyl-L-methionine | [12] |
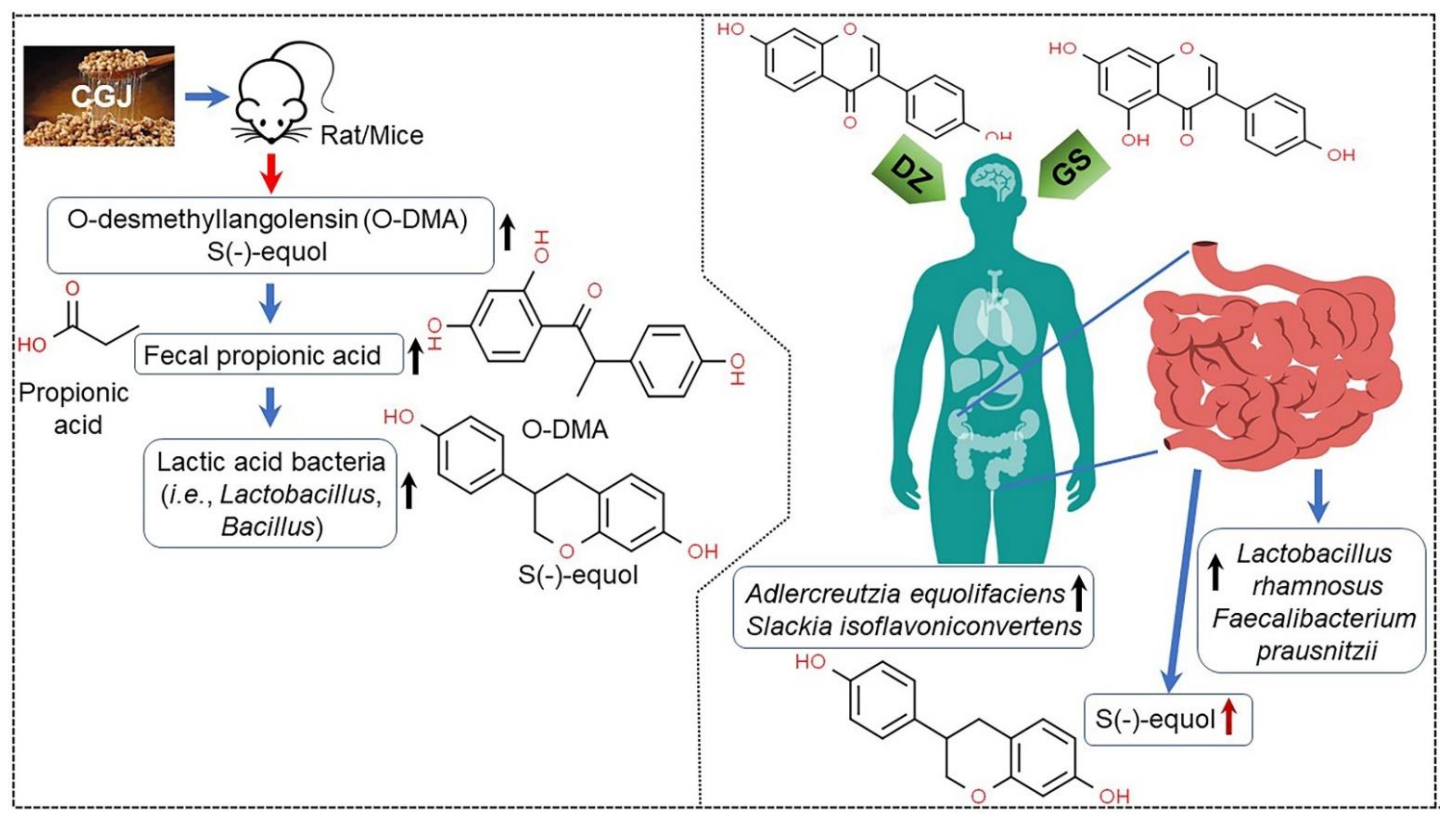
| Raw soybean | Fecal bacteria | Enhancing short-chain fatty acids, methionine biosynthesis; depleting chondroitin sulfate degradation Increasing abundance of probiotics, including Coprococcus, Ruminococcus, and Bifibobacterium; reducing growth of Sutterella | [77] |
| Model | Strain Used in Fermentation | Action Mode | Reference |
|---|---|---|---|
| Raw soybean | – | Anti-tyrosinase, anti-proliferative, and syringic acid-mediated estrogenic activities | [166] |
| Mice | B. amyloliquefaciens SCGB1 | Amelioration of atopic dermatitis-like skin lesion: suppressing mast cell infiltration, immunoglobulin-E expression, and TH2 IL-4 and itch-related IL-31 cytokine; alleviating collagen fiber deposition; activating dephosphorylation of NF-κB and MAPK | [167] |
| IL4/Luc/CNS-1 Tg mice | B. subtilis MC31 | Therapeutic effect on atopic dermatitis: producing high concentration of gamma-aminobutyric acid (GABA); improving common allergic responses, including decreased thickness of ear and dermis, and reduction of auricular lymph node (ALN) weight and infiltrating mast cells, while decreasing IgE levels in epidermis | [168] |
| Men with stage II to V patterns of hair loss | Leuconostoc holzapfelii, Leuconostoc mesenteroides L.sake | Anti-androgenetic alopecia: enhancing hair count and thickness; promoting hair growth without reverse hair loss and side effects such as diarrhea | [169] |
| C57BL/ 6J mice | B. subtilis | Anti-asthmatic activity: suppressing histamine release of rat peritoneal mast cells by inhibiting calcium uptake as well as ear swelling by permeation of inflammatory cells; downregulating the population of eosinophil and monocytes in the lungs of mice; repressing histopathological changes, such as eosinophil infiltration, mucus accumulation, goblet cell hyperplasia, and collagen fiber deposits | [170] |
| ICR mice | B. subtilis MC31 L. sakei 383 | Reduction of toxic effects in liver and kidney; regulating levels of alkaline phosphatase, alanine aminotransferase, aspartate aminotransferase, and lactate dehydrogenase; reducing blood urea nitrogen and serum creatinine levels | [132] |
| Raw soybean | B. amyloliqueciens CH86-1 B. licheniformis 58 B. licheniformis 67 B. amyloliquefaciens RWL-1 | Increasing amounts of most amino acids including proline, sugars (sucrose, fructose, glucose, mannose, and xylose), and organic acids (glutamic acid, succinic acid, and malonic acid) | [175] |
| Raw soybean | B. subtilis MYCO10001 B. subtilis ATCC 21228 | Enhancing levels of isoflavones (daidzein and genistein), their derivatives (isoflavone-β-glucosides and isoflavone-aglycones), or succinyl derivatives (succinyl-β-daidzin and succinyl-β- genistin), and β-glucosidase | [104,105] |
| Raw soybean | L. acidophilus KCTC 3925 L. rhamnosus KCTC 3929 | Producing aglycone-formed isoflavones and exopolysaccharide | [106] |
| Raw soybean | B. subtilis | Enhancing isoflavone levels by controlling CGJ fermentation conditions, including temperature, time, germination, and osmolarity | [107] |
| Raw soybean | Bacillus sp. FBL-2 | Agro-industrial bioresource: production of γ-PGA using rice bran as a renewable substrate | [109] |
| Raw soybean | B. subtilis CSY191 | Fatty acid and volatile compound profiling during CGJ fermentation: identifying 10 fatty acids including C16:0, C18:0, C18:1 ω9, C18:1 ω7, C18:2 ω6, C18:3 ω3, C20:0, C20:1 ω9, C22:0, and C24:0, and 13 different volatile ketones, including acetone, 2,3-butanedione, and 3-hydroxy-3-methyl-2-butanone | [172] |
| Raw soybean | B. amyloliquefaciens CH51 | Production of industrial enzyme: secreting xylanase (19 kDa) after introduction of xynA into B. subtilis WB600 | [173] |
| Raw soybean | B. subtilis NB-NUC1 | Increasing γ-glutamyl transpeptidase and extracellular protein content when supplemented with 1% glycine | [174] |
| Healthy Korean men (age range 21–29 years) | – | Improving human health function of isoflavones following CGJ ingestion | [108] |
| Raw soybean | B. licheniformis KCCM 11053P B. licheniformis KCCM 11054P B. amyloliquefaciens CH86-1 | Improvement of purine metabolism via metabolic profiling during CGJ fermentation: increasing uracil and thymine levels; improving xanthine and adenine levels during CGJ fermentation process; enhancing hypoxanthine and guanine levels | [43,176] |
| Raw soybean | B. amyloliqueciens CH86-1 B. licheniformis 58 B. licheniformis 67 | Enhanced nutritional value: increasing the amounts of most amino acids including proline, sugars (sucrose, fructose, glucose, mannose, and xylose), and organic acids (glutamic acid, succinic acid, and malonic acid) | [175] |
| Raw soybean | Enterococcusfaecium | Establishment of fermentation culture platform for high accumulation of tyramine: reducing the safety risks that may arise when consuming CGJ with high tyramine amount by lowering fermentation temperature and shortening duration period; controlling tyrosine decarboxylase (TDC) activity using nicotinic acid as a TDC inhibitor to produce high amount of tyramine | [162,177,178] |
| Raw soybean | Bacillus spp. | Bacillus taxonomy: developing recQ as a genetic marker for the classification of Bacillus taxa | [179] |
| Raw soybean | – | Phenotype characterization of osmotic (~8% NaCl) tolerant B. glycinifermentans sp. | [180] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, I.-S.; Hwang, C.-W.; Yang, W.-S.; Kim, C.-H. Current Perspectives on the Physiological Activities of Fermented Soybean-Derived Cheonggukjang. Int. J. Mol. Sci. 2021, 22, 5746. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115746
Kim I-S, Hwang C-W, Yang W-S, Kim C-H. Current Perspectives on the Physiological Activities of Fermented Soybean-Derived Cheonggukjang. International Journal of Molecular Sciences. 2021; 22(11):5746. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115746
Chicago/Turabian StyleKim, Il-Sup, Cher-Won Hwang, Woong-Suk Yang, and Cheorl-Ho Kim. 2021. "Current Perspectives on the Physiological Activities of Fermented Soybean-Derived Cheonggukjang" International Journal of Molecular Sciences 22, no. 11: 5746. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115746