
Calreticulin Deficiency Disturbs Ribosome Biogenesis and Results in Retardation in Embryonic Kidney Development

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
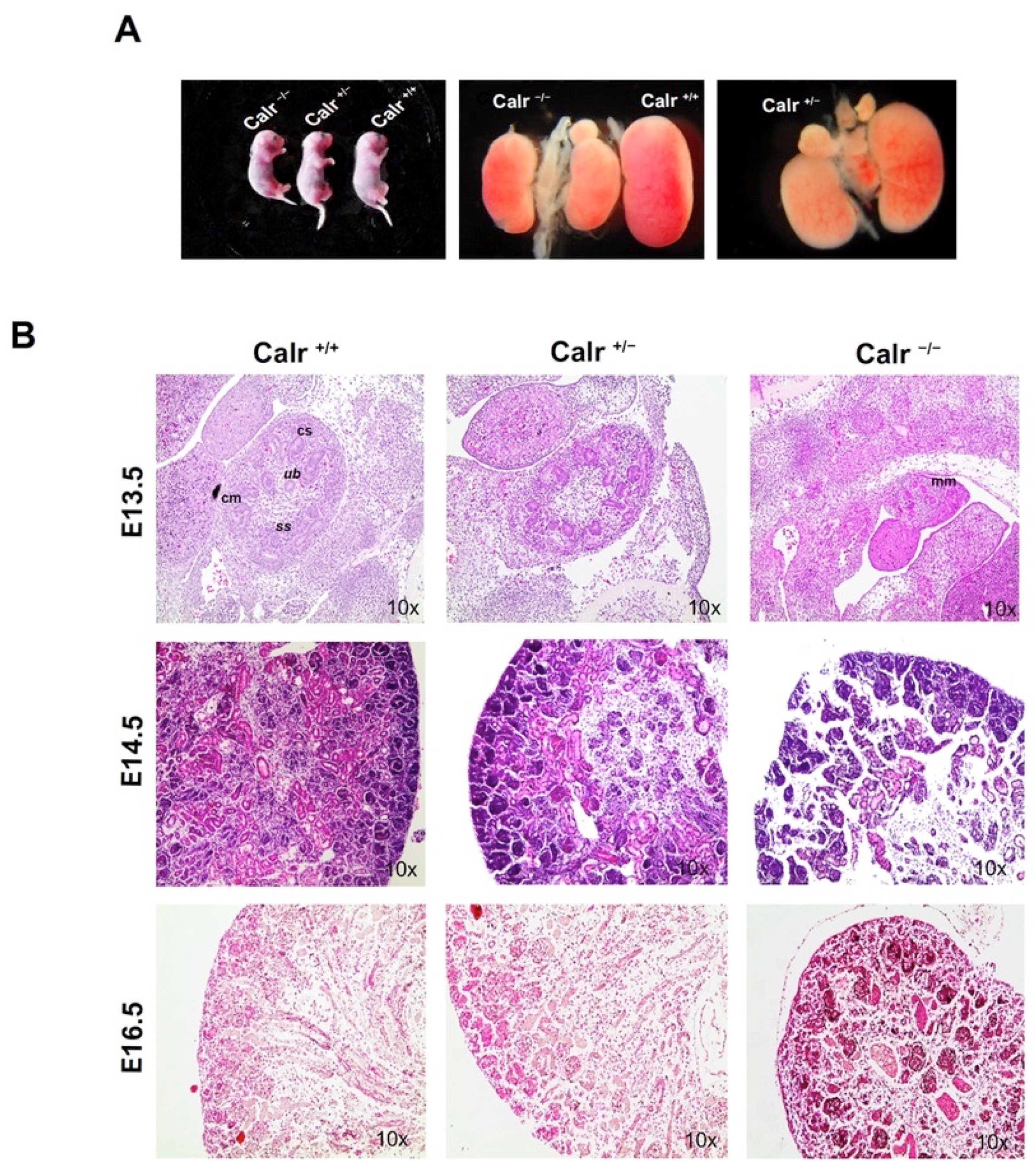
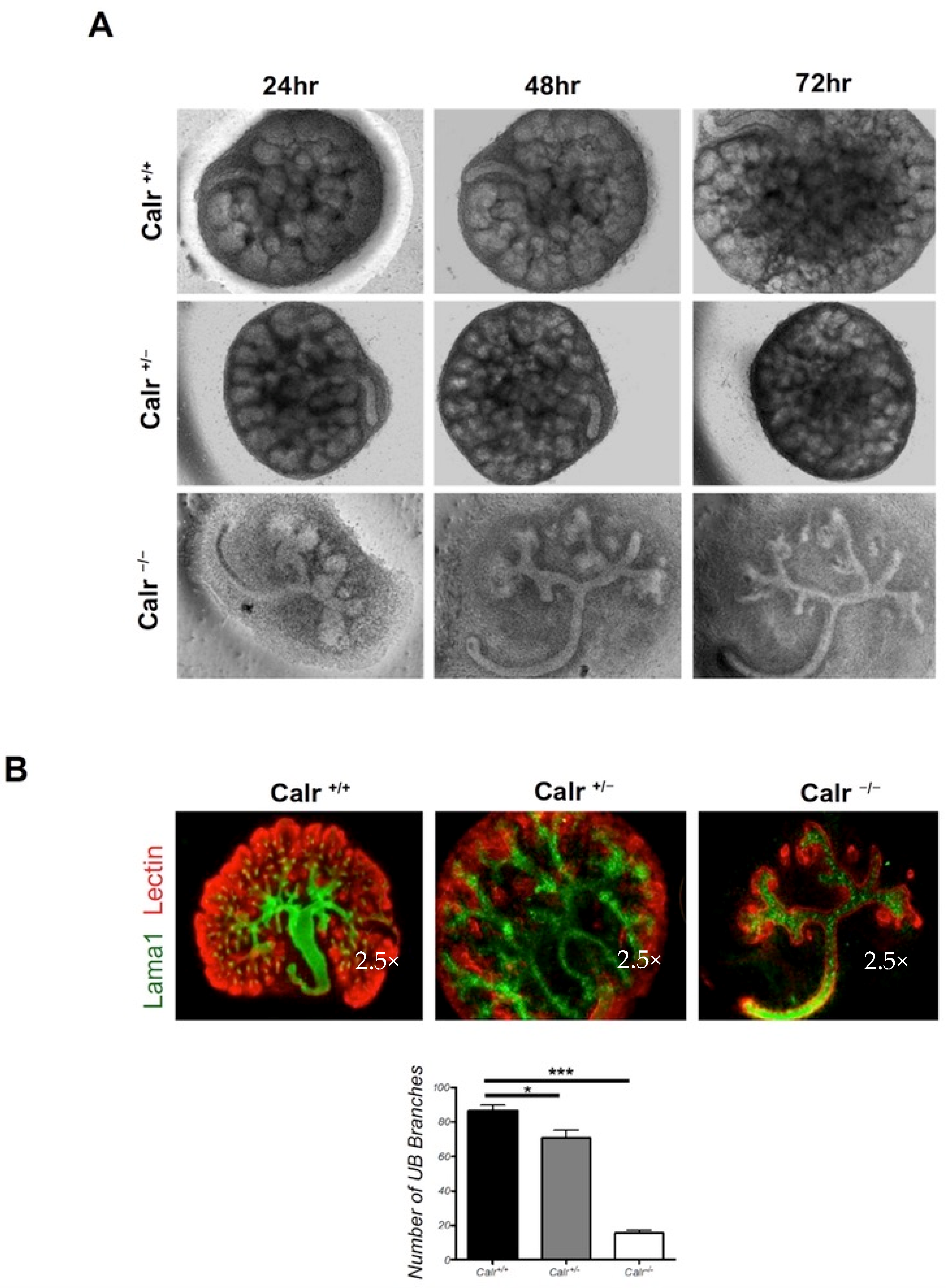
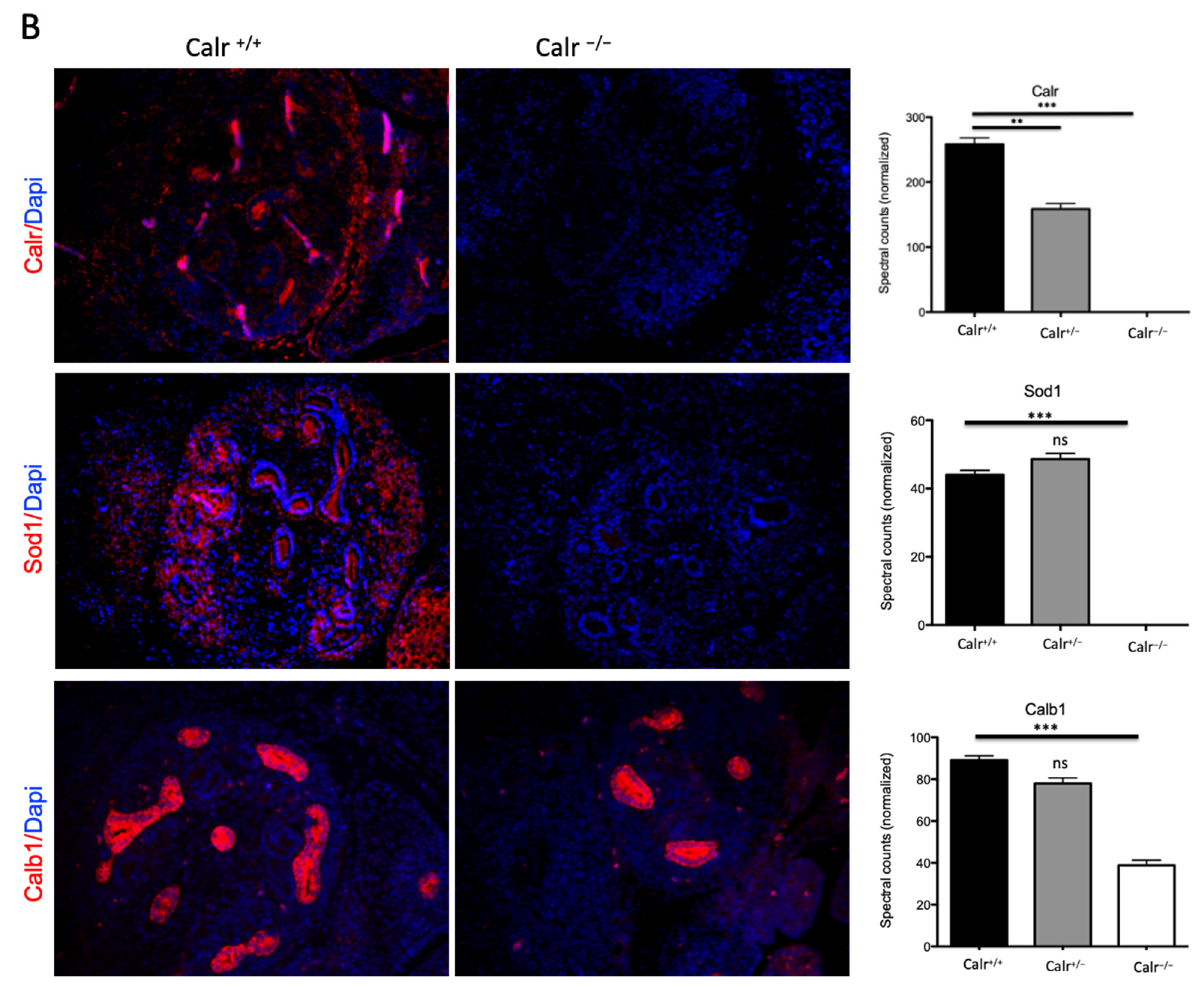
2.1. Calreticulin Knockout Results in Morphological and Histological Abnormalities of the Kidney and in Impaired Renal Branching
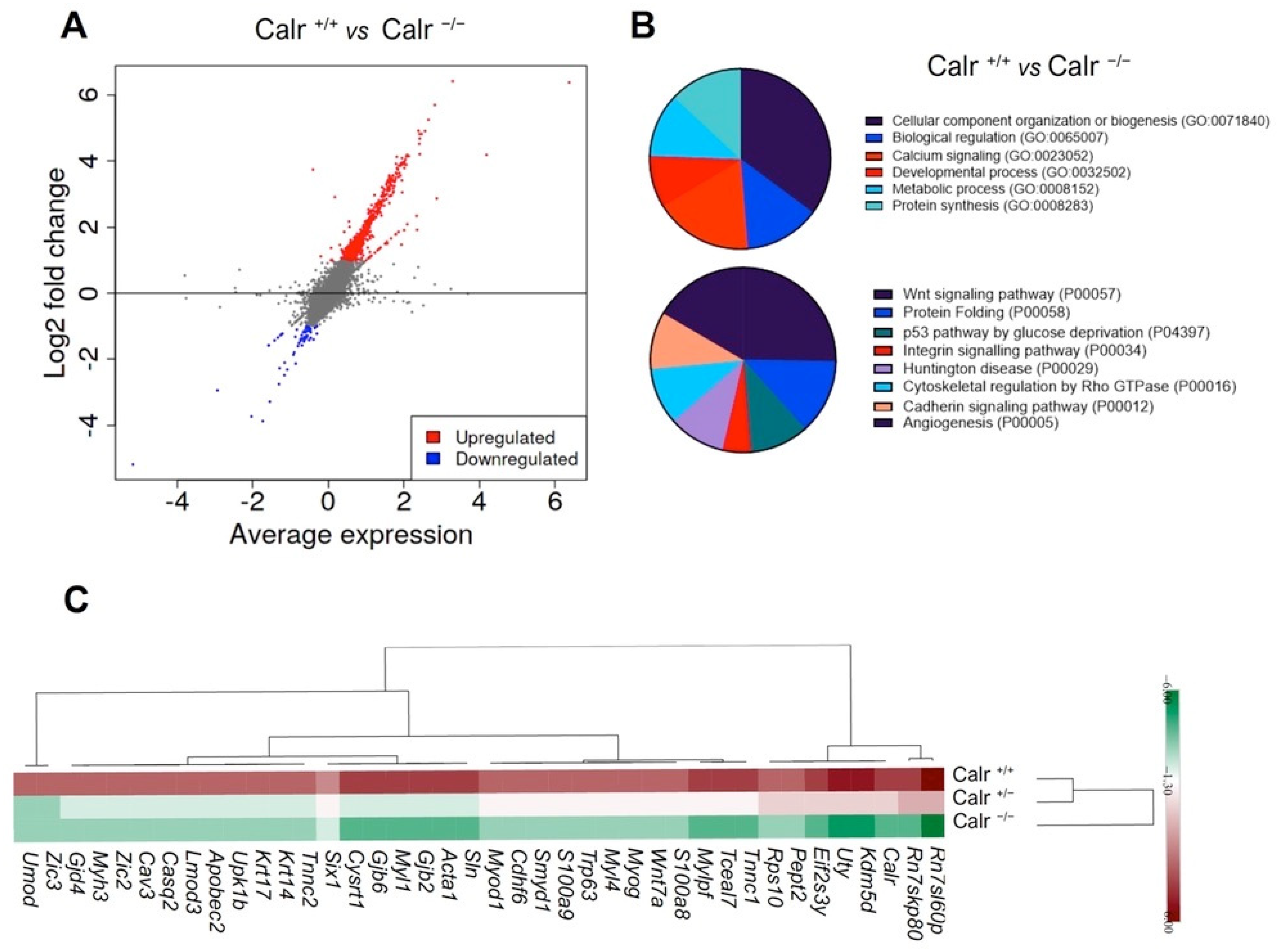
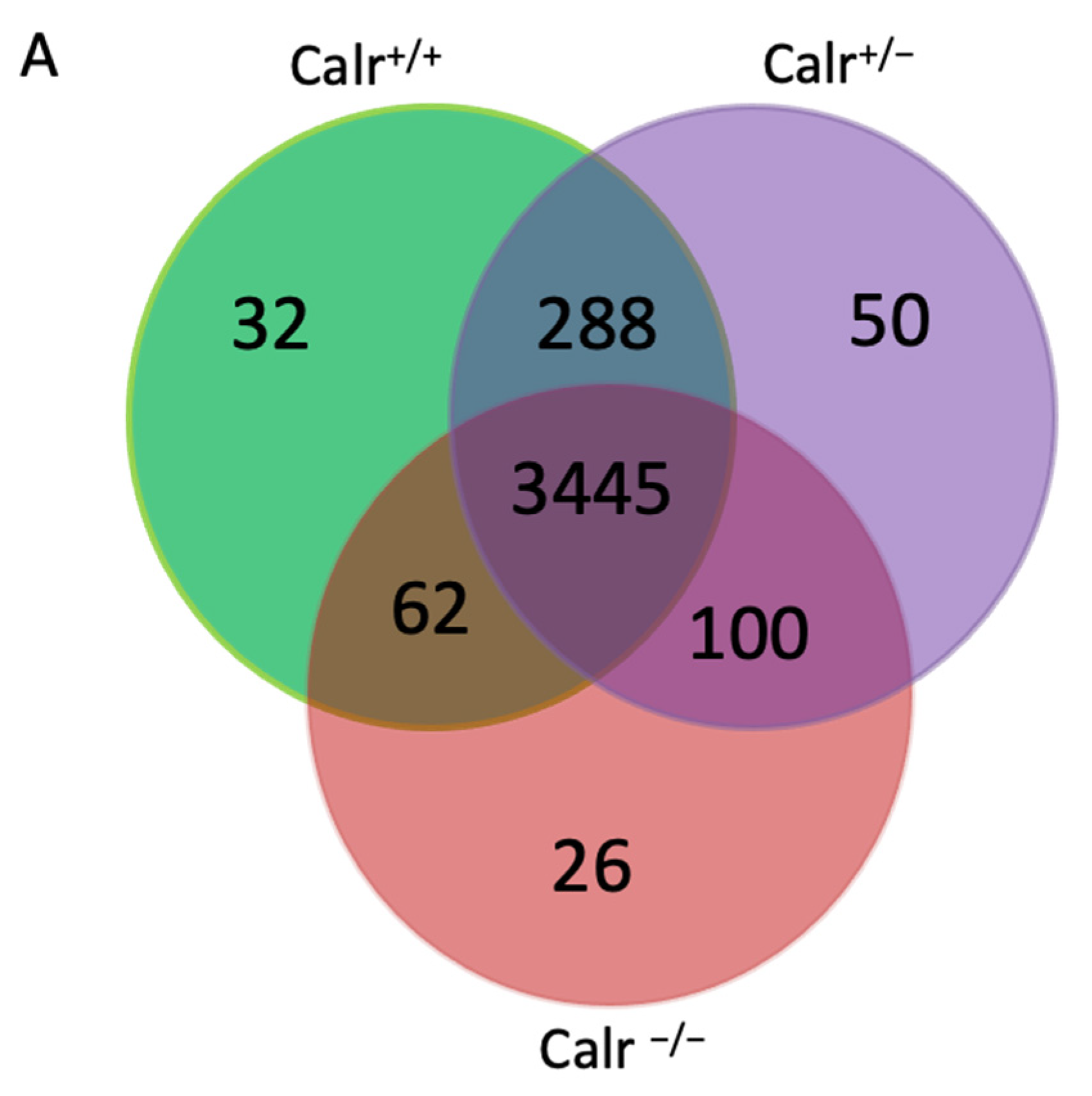
2.2. Calr Deficiency Is Associated with Large and Significant Transcriptome Alterations
2.3. Comparative Proteomic Analyses Identified Significant Alterations in the Proteome of the Calr−/− Embryonic Kidney
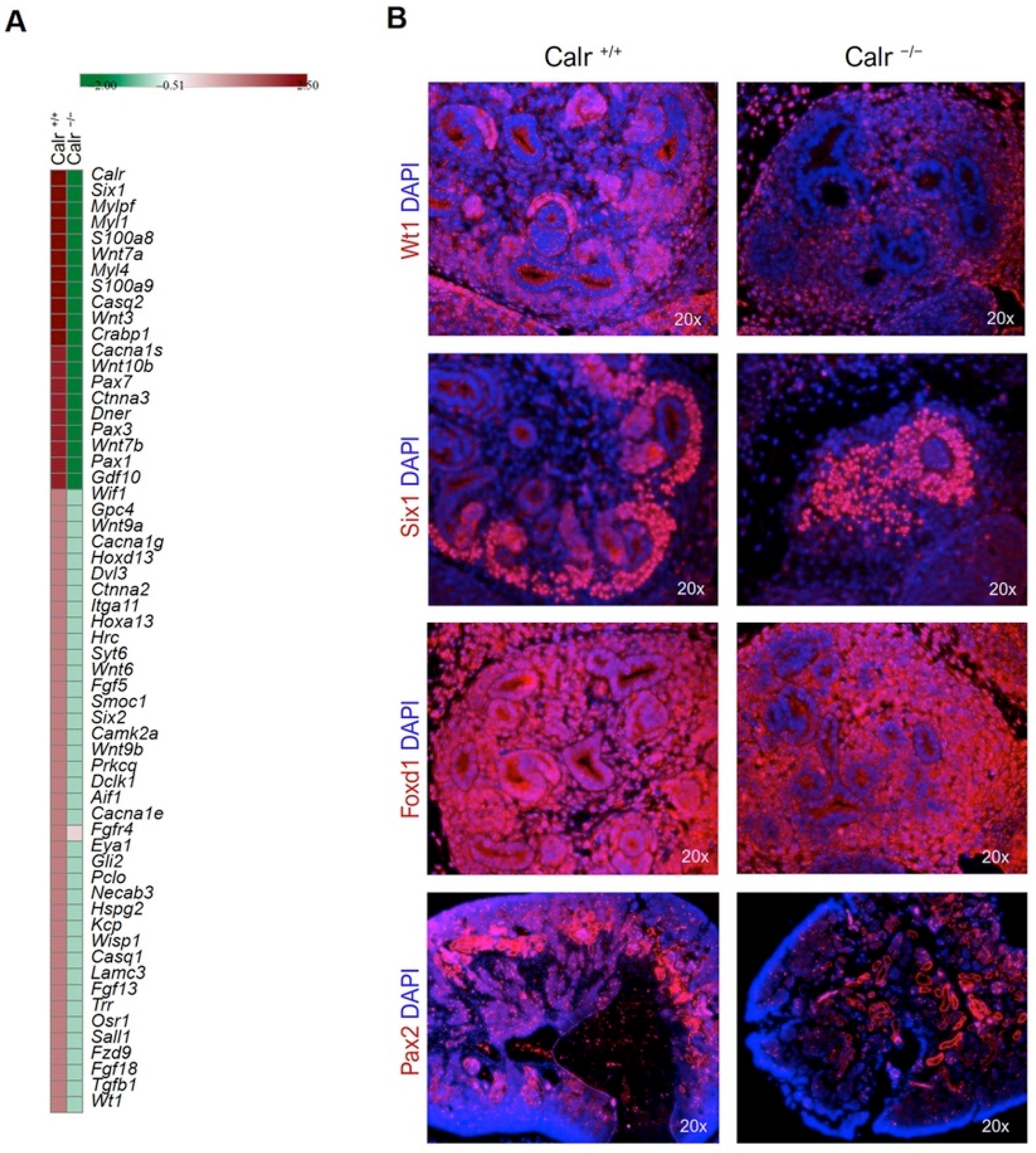
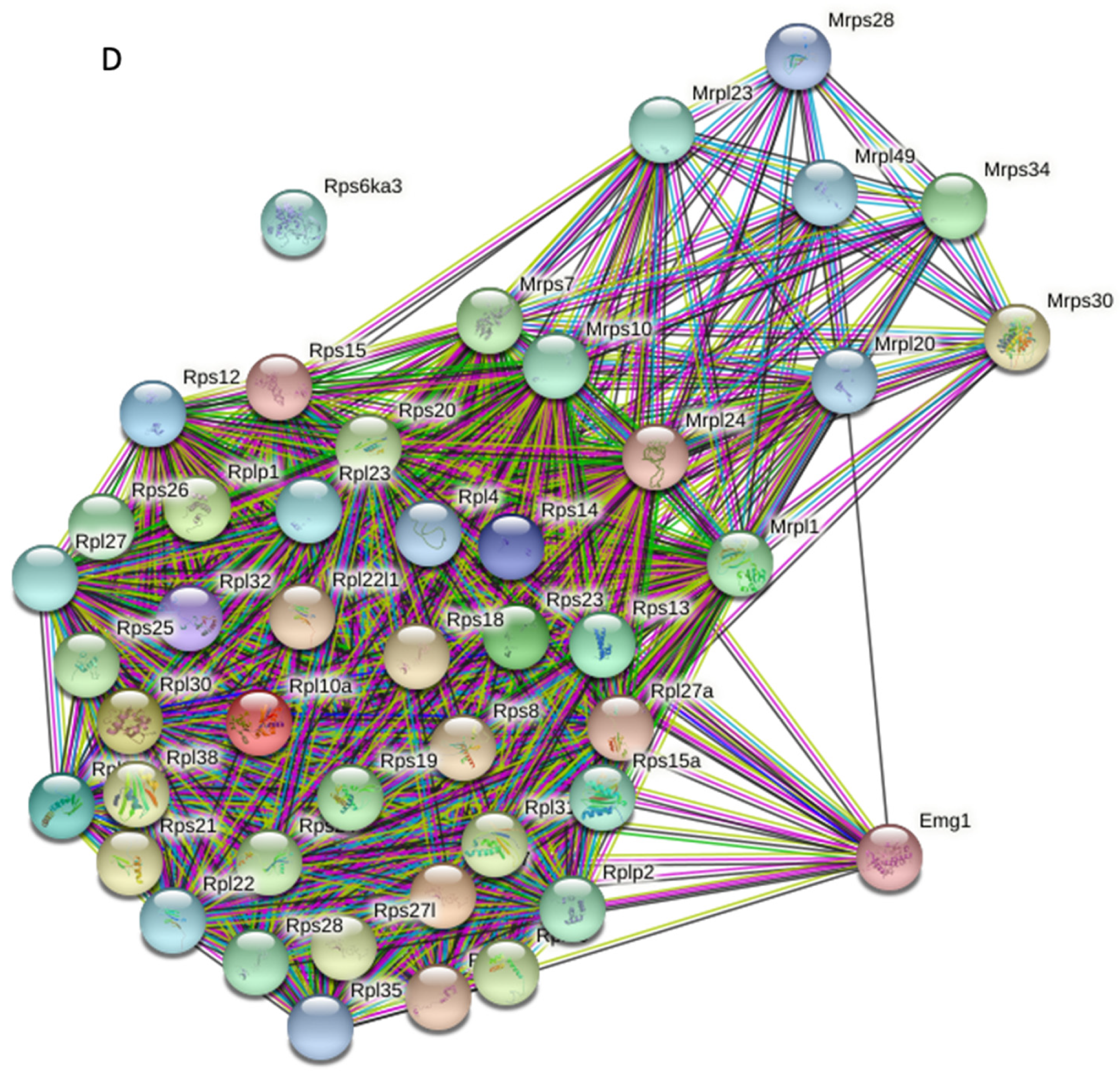
2.4. Gene Ontology Classification and Protein–Protein Interaction Network Analyses
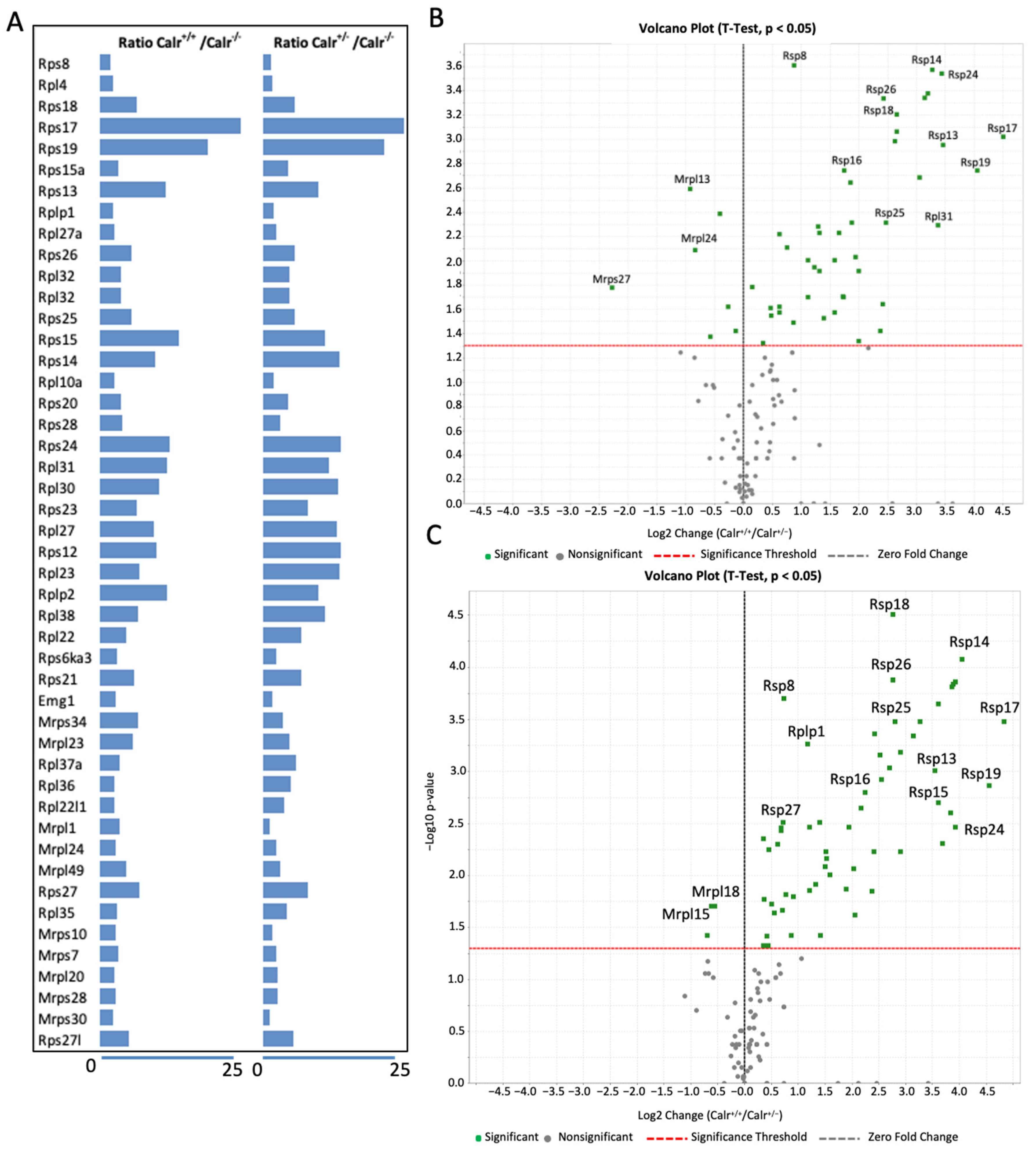
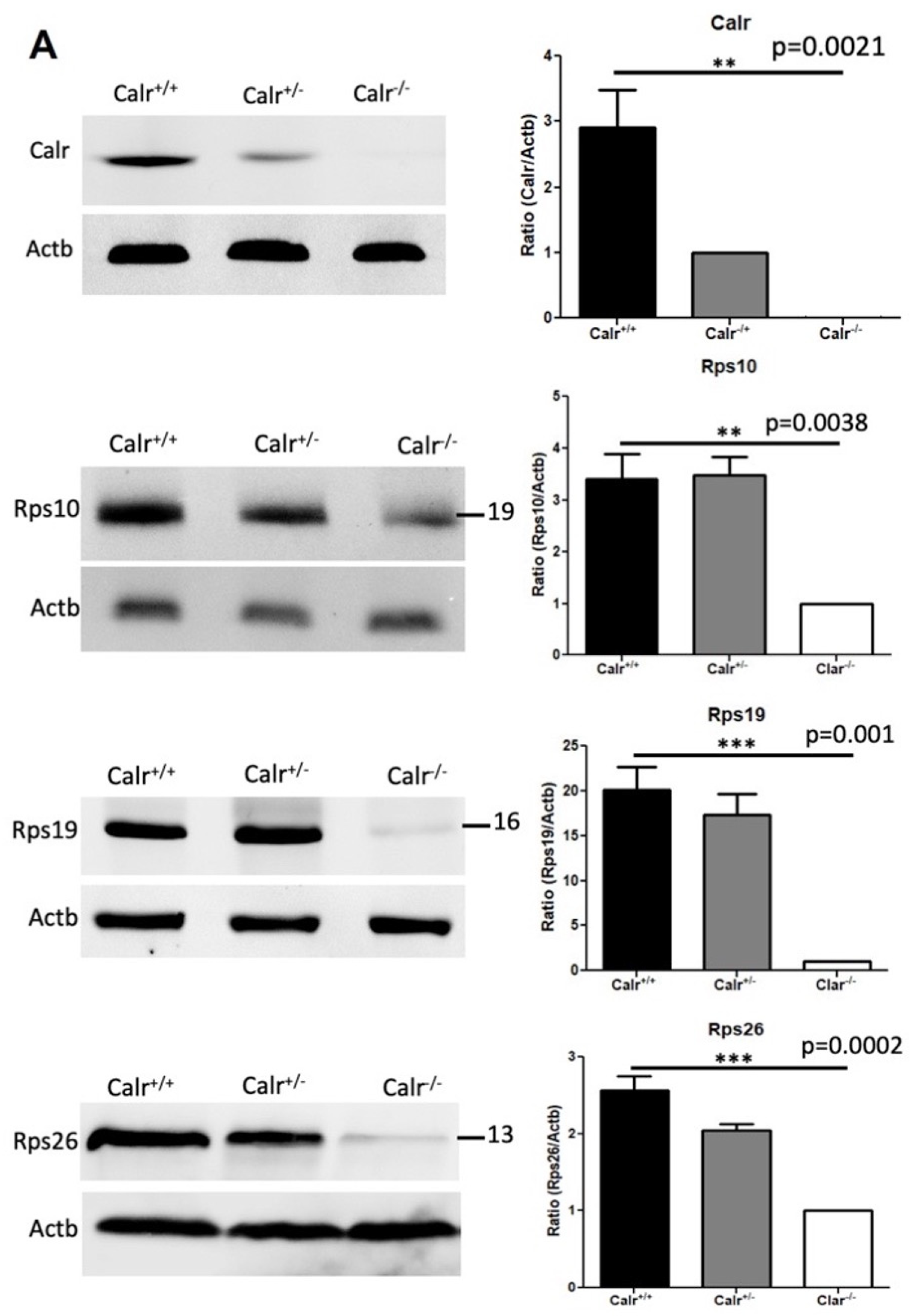
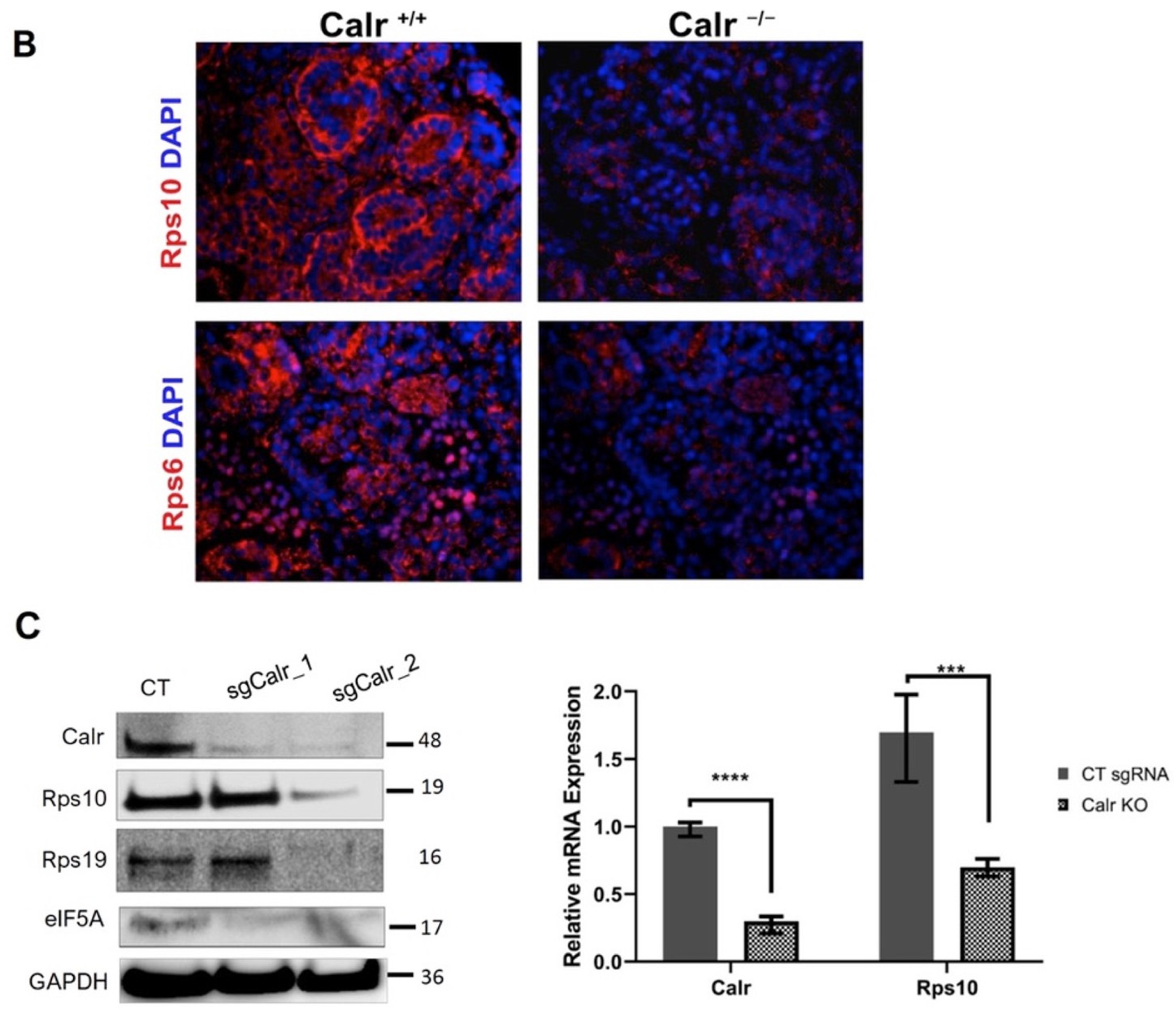
2.5. Calr Knockout Results in an Alteration in Ribosomal Protein Expression
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Histochemical Staining
4.3. Ex-Vivo Organ Culture of Embryonic Kidney
4.4. Immunofluorescence Staining of the Cultured Kidneys and Immunohistological Analysis of the Kidney Sections
4.5. Cell Culture
4.6. Generation of Knockout Cells
4.7. Transcriptome Analysis
4.8. Kidney Lysis, Protein Extraction and Two -Dimensional Gel Electrophoresis (2-DE)
4.9. Mass Spectrometric Analysis and Protein Identification
4.10. Protein Extraction, SDS-PAGE, in Gel Tryptic Digestion, and Mass Spectrometric Analyses
4.11. Western Blot Analysis
4.12. Bioinformatics
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Saxen, L.; Saxén, L. Organogenesis of the Kidney; Cambridge University Press: Cambridge, UK, 1987. [Google Scholar]
- Burrow, C.R. Regulatory Molecules in Kidney Development. Pediatr. Nephrol. 2000, 14, 240–253. [Google Scholar] [CrossRef]
- Costantini, F.; Kopan, R. Patterning a Complex Organ: Branching Morphogenesis and Nephron Segmentation in Kidney Development. Dev. Cell 2010, 18, 698–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichel, J.G.; Shen, L.; Sheng, H.Z.; Granholm, A.C.; Drago, J.; Grinberg, A.; Lee, E.J.; Huang, S.P.; Saarma, M.; Hoffer, B.J.; et al. Defects in Enteric Innervation and Kidney Development in Mice Lacking GDNF. Nature 1996, 382, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Wnt/β-Catenin Signaling Regulates Nephron Induction during Mouse Kidney Development|Development. Available online: https://dev.biologists.org/content/134/13/2533.short (accessed on 19 August 2020).
- Bridgewater, D.; Di Giovanni, V.; Cain, J.E.; Cox, B.; Jakobson, M.; Sainio, K.; Rosenblum, N.D. β-Catenin Causes Renal Dysplasia via Upregulation of Tgfβ2 and Dkk1. J. Am. Soc. Nephrol. 2011, 22, 718–731. [Google Scholar] [CrossRef] [Green Version]
- Dihazi, G.H.; Jahn, O.; Tampe, B.; Zeisberg, M.; Müller, C.; Müller, G.A.; Dihazi, H. Proteomic Analysis of Embryonic Kidney Development: Heterochromatin Proteins as Epigenetic Regulators of Nephrogenesis. Sci. Rep. 2015, 5, 13951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calreticulin: One Protein, One Gene, Many Functions | Biochemical Journal|Portland Press. Available online: https://portlandpress.com/biochemj/article/344/2/281/36172/Calreticulin-one-protein-one-gene-many-functions (accessed on 28 June 2020).
- Bedard, K.; Szabo, E.; Michalak, M.; Opas, M. Cellular Functions of Endoplasmic Reticulum Chaperones Calreticulin, Calnexin, and ERp57. In International Review of Cytology; A Survey of Cell Biology; Academic Press: Cambridge, MA, USA, 2005; Volume 245, pp. 91–121. [Google Scholar] [CrossRef]
- Michalak, M.; Robert Parker, J.M.; Opas, M. Ca2+ Signaling and Calcium Binding Chaperones of the Endoplasmic Reticulum. Cell Calcium 2002, 32, 269–278. [Google Scholar] [CrossRef]
- Molinari, M.; Eriksson, K.K.; Calanca, V.; Galli, C.; Cresswell, P.; Michalak, M.; Helenius, A. Contrasting Functions of Calreticulin and Calnexin in Glycoprotein Folding and ER Quality Control. Mol. Cell 2004, 13, 125–135. [Google Scholar] [CrossRef]
- Okunaga, T.; Urata, Y.; Goto, S.; Matsuo, T.; Mizota, S.; Tsutsumi, K.; Nagata, I.; Kondo, T.; Ihara, Y. Calreticulin, a Molecular Chaperone in the Endoplasmic Reticulum, Modulates Radiosensitivity of Human Glioblastoma U251MG Cells. Cancer Res. 2006, 66, 8662–8671. [Google Scholar] [CrossRef] [Green Version]
- Lu, A.; Pallero, M.A.; Owusu, B.Y.; Borovjagin, A.V.; Lei, W.; Sanders, P.W.; Murphy-Ullrich, J.E. Calreticulin Is Important for the Development of Renal Fibrosis and Dysfunction in Diabetic Nephropathy. Matrix Biol. Plus 2020, 8, 100034. [Google Scholar] [CrossRef]
- Maruyama, K.; Nakagawa, N.; Suzuki, A.; Kabara, M.; Matsuki, M.; Shindo, M.; Iwasaki, S.; Ogawa, Y.; Hasebe, N. Novel Detection of CALR-Mutated Cells in Myeloproliferative Neoplasm-Related Glomerulopathy With Interstitial Extramedullary Hematopoiesis: A Case Report. Am. J. Kidney Dis. 2019, 74, 844–848. [Google Scholar] [CrossRef] [PubMed]
- Mesaeli, N.; Nakamura, K.; Zvaritch, E.; Dickie, P.; Dziak, E.; Krause, K.H.; Opas, M.; MacLennan, D.H.; Michalak, M. Calreticulin Is Essential for Cardiac Development. J. Cell Biol. 1999, 144, 857–868. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, S.; Rehn, M.; Itaranta, P.; Tuukkanen, J.; Heljasvaara, R.; Peltoketo, H.; Pihlajaniemi, T.; Vainio, S. Induced Repatterning of Type XVIII Collagen Expression in Ureter Bud from Kidney to Lung Type: Association with Sonic Hedgehog and Ectopic Surfactant Protein C. Development 2001, 128, 1573–1585. [Google Scholar] [CrossRef] [PubMed]
- Kurtzeborn, K.; Cebrian, C.; Kuure, S. Regulation of Renal Differentiation by Trophic Factors. Front Physiol 2018, 9, 1588. [Google Scholar] [CrossRef] [PubMed]
- Rowan, C.J.; Li, W.; Martirosyan, H.; Erwood, S.; Hu, D.; Kim, Y.-K.; Sheybani-Deloui, S.; Mulder, J.; Blake, J.; Chen, L.; et al. Hedgehog-GLI Signaling in Foxd1-Positive Stromal Cells Promotes Murine Nephrogenesis via TGFβ Signaling. Development 2018, 145. [Google Scholar] [CrossRef] [Green Version]
- Groenendyk, J.; Michalak, M. Disrupted WNT Signaling in Mouse Embryonic Stem Cells in the Absence of Calreticulin. Stem. Cell Rev. Rep. 2014, 10, 191–206. [Google Scholar] [CrossRef]
- Wnt11 and Ret/Gdnf Pathways Cooperate in Regulating Ureteric Branching during Metanephric Kidney Development|Development. Available online: https://dev.biologists.org/content/130/14/3175.short (accessed on 28 June 2020).
- Saito, Y.; Ihara, Y.; Leach, M.R.; Cohen-Doyle, M.F.; Williams, D.B. Calreticulin Functions in Vitro as a Molecular Chaperone for Both Glycosylated and Non-Glycosylated Proteins. EMBO J. 1999, 18, 6718–6729. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Nag, S.; Zhang, X.; Wang, M.-H.; Wang, H.; Zhou, J.; Zhang, R. Ribosomal Proteins and Human Diseases: Pathogenesis, Molecular Mechanisms, and Therapeutic Implications. Med. Res. Rev. 2015, 35, 225–285. [Google Scholar] [CrossRef] [PubMed]
- Ruggero, D.; Pandolfi, P.P. Does the Ribosome Translate Cancer? Nat. Rev. Cancer 2003, 3, 179–192. [Google Scholar] [CrossRef]
- Wool, I.G. Extraribosomal Functions of Ribosomal Proteins. Trends Biochem. Sci. 1996, 21, 164–165. [Google Scholar] [CrossRef]
- Warner, J.R.; McIntosh, K.B. How Common Are Extraribosomal Functions of Ribosomal Proteins? Mol. Cell 2009, 34, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindström, M.S.; Zhang, Y. Ribosomal Protein S9 Is a Novel B23/NPM-Binding Protein Required for Normal Cell Proliferation. J. Biol. Chem. 2008, 283, 15568–15576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volarevic, S.; Stewart, M.J.; Ledermann, B.; Zilberman, F.; Terracciano, L.; Montini, E.; Grompe, M.; Kozma, S.C.; Thomas, G. Proliferation, but Not Growth, Blocked by Conditional Deletion of 40S Ribosomal Protein S6. Science 2000, 288, 2045–2047. [Google Scholar] [CrossRef] [PubMed]
- Jang, C.-Y.; Lee, J.Y.; Kim, J. RpS3, a DNA Repair Endonuclease and Ribosomal Protein, Is Involved in Apoptosis. FEBS Lett. 2004, 560, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Hegde, V.; Wang, M.; Deutsch, W.A. Human Ribosomal Protein S3 Interacts with DNA Base Excision Repair Proteins HAPE/Ref-1 and HOGG1. Biochemistry 2004, 43, 14211–14217. [Google Scholar] [CrossRef]
- Kim, J.; Chubatsu, L.S.; Admon, A.; Stahl, J.; Fellous, R.; Linn, S. Implication of Mammalian Ribosomal Protein S3 in the Processing of DNA Damage. J. Biol. Chem. 1995, 270, 13620–13629. [Google Scholar] [CrossRef] [Green Version]
- Zhan, Y.; Melian, N.Y.; Pantoja, M.; Haines, N.; Ruohola-Baker, H.; Bourque, C.W.; Rao, Y.; Carbonetto, S. Dystroglycan and Mitochondrial Ribosomal Protein L34 Regulate Differentiation in the Drosophila Eye. PLoS ONE 2010, 5, e10488. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; You, K.-R.; Kim, I.H.; Cho, B.-H.; Kim, C.-Y.; Kim, D.-G. Over-Expression of the Ribosomal Protein L36a Gene Is Associated with Cellular Proliferation in Hepatocellular Carcinoma. Hepatology 2004, 39, 129–138. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Sun, Y. Ribosomal Protein S27L Is a Direct P53 Target That Regulates Apoptosis. Oncogene 2007, 26, 2707–2716. [Google Scholar] [CrossRef] [Green Version]
- Flygare, J.; Kiefer, T.; Miyake, K.; Utsugisawa, T.; Hamaguchi, I.; Da Costa, L.; Richter, J.; Davey, E.J.; Matsson, H.; Dahl, N.; et al. Deficiency of Ribosomal Protein S19 in CD34+ Cells Generated by SiRNA Blocks Erythroid Development and Mimics Defects Seen in Diamond-Blackfan Anemia. Blood 2005, 105, 4627–4634. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.J.; Lauritsen, J.P.H.; Hartman, M.G.; Foushee, A.M.D.; Lefebvre, J.M.; Shinton, S.A.; Gerhardt, B.; Hardy, R.R.; Oravecz, T.; Wiest, D.L. Ablation of Ribosomal Protein L22 Selectively Impairs Alphabeta T Cell Development by Activation of a P53-Dependent Checkpoint. Immunity 2007, 26, 759–772. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, L.; Narla, G.; Willig, T.-N.; Peters, L.L.; Parra, M.; Fixler, J.; Tchernia, G.; Mohandas, N. Ribosomal Protein S19 Expression during Erythroid Differentiation. Blood 2003, 101, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Draptchinskaia, N.; Gustavsson, P.; Andersson, B.; Pettersson, M.; Willig, T.N.; Dianzani, I.; Ball, S.; Tchernia, G.; Klar, J.; Matsson, H.; et al. The Gene Encoding Ribosomal Protein S19 Is Mutated in Diamond-Blackfan Anaemia. Nat. Genet. 1999, 21, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Ebert, B.L.; Pretz, J.; Bosco, J.; Chang, C.Y.; Tamayo, P.; Galili, N.; Raza, A.; Root, D.E.; Attar, E.; Ellis, S.R.; et al. Identification of RPS14 as a 5q- Syndrome Gene by RNA Interference Screen. Nature 2008, 451, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Costantini, F.; Shakya, R. GDNF/Ret Signaling and the Development of the Kidney. BioEssays 2006, 28, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, A.; Rosenberg, P.S.; Atsidaftos, E.; Alter, B.P.; Lipton, J.M. Incidence of Neoplasia in Diamond Blackfan Anemia: A Report from the Diamond Blackfan Anemia Registry. Blood 2012, 119, 3815–3819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlachos, A.; Rosenberg, P.S.; Kang, J.; Atsidaftos, E.; Alter, B.P.; Lipton, J.M. Myelodysplastic Syndrome and Gastrointestinal Carcinomas Characterize the Cancer Risk in Diamond Blackfan Anemia: A Report from the Diamond Blackfan Anemia Registry. Blood 2016, 128, 333. [Google Scholar] [CrossRef]
- Rumi, E.; Harutyunyan, A.S.; Pietra, D.; Milosevic, J.D.; Casetti, I.C.; Bellini, M.; Them, N.C.C.; Cavalloni, C.; Ferretti, V.V.; Milanesi, C.; et al. CALR Exon 9 Mutations Are Somatically Acquired Events in Familial Cases of Essential Thrombocythemia or Primary Myelofibrosis. Blood 2014, 123, 2416–2419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebinger, D.D.R.; Unbekandt, M.; Ganeva, V.V.; Ofenbauer, A.; Werner, C.; Davies, J.A. A Novel, Low-Volume Method for Organ Culture of Embryonic Kidneys That Allows Development of Cortico-Medullary Anatomical Organization. PLoS ONE 2010, 5, e10550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq--a Python Framework to Work with High-Throughput Sequencing Data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Durinck, S.; Moreau, Y.; Kasprzyk, A.; Davis, S.; De Moor, B.; Brazma, A.; Huber, W. BioMart and Bioconductor: A Powerful Link between Biological Databases and Microarray Data Analysis. Bioinformatics 2005, 21, 3439–3440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene Ontology Analysis for RNA-Seq: Accounting for Selection Bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wessel, D.; Flügge, U.I. A Method for the Quantitative Recovery of Protein in Dilute Solution in the Presence of Detergents and Lipids. Anal. Biochem. 1984, 138, 141–143. [Google Scholar] [CrossRef]
- Dihazi, H.; Dihazi, G.H.; Nolte, J.; Meyer, S.; Jahn, O.; Müller, G.A.; Engel, W. Multipotent Adult Germline Stem Cells and Embryonic Stem Cells: Comparative Proteomic Approach. J. Proteome Res. 2009, 8, 5497–5510. [Google Scholar] [CrossRef]
- Dihazi, H.; Asif, A.R.; Agarwal, N.K.; Doncheva, Y.; Müller, G.A. Proteomic Analysis of Cellular Response to Osmotic Stress in Thick Ascending Limb of Henle’s Loop (TALH) Cells. Mol. Cell. Proteom. 2005, 4, 1445–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic Transfer of Proteins from Polyacrylamide Gels to Nitrocellulose Sheets: Procedure and Some Applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Mering, C.; Jensen, L.J.; Snel, B.; Hooper, S.D.; Krupp, M.; Foglierini, M.; Jouffre, N.; Huynen, M.A.; Bork, P. STRING: Known and predicted protein-protein associations, integrated and transferred across organisms. Nucleic Acids Res. 2005, 33, D433–D437. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serin, N.; Dihazi, G.H.; Tayyeb, A.; Lenz, C.; Müller, G.A.; Zeisberg, M.; Dihazi, H. Calreticulin Deficiency Disturbs Ribosome Biogenesis and Results in Retardation in Embryonic Kidney Development. Int. J. Mol. Sci. 2021, 22, 5858. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115858
Serin N, Dihazi GH, Tayyeb A, Lenz C, Müller GA, Zeisberg M, Dihazi H. Calreticulin Deficiency Disturbs Ribosome Biogenesis and Results in Retardation in Embryonic Kidney Development. International Journal of Molecular Sciences. 2021; 22(11):5858. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115858
Chicago/Turabian StyleSerin, Nazli, Gry H. Dihazi, Asima Tayyeb, Christof Lenz, Gerhard A. Müller, Michael Zeisberg, and Hassan Dihazi. 2021. "Calreticulin Deficiency Disturbs Ribosome Biogenesis and Results in Retardation in Embryonic Kidney Development" International Journal of Molecular Sciences 22, no. 11: 5858. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115858