
The Evolution of Rag Gene Enhancers and Transcription Factor E and Id Proteins in the Adaptive Immune System


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Regulation of Rag1/2 Gene by T or B Cell-Specific Enhancers
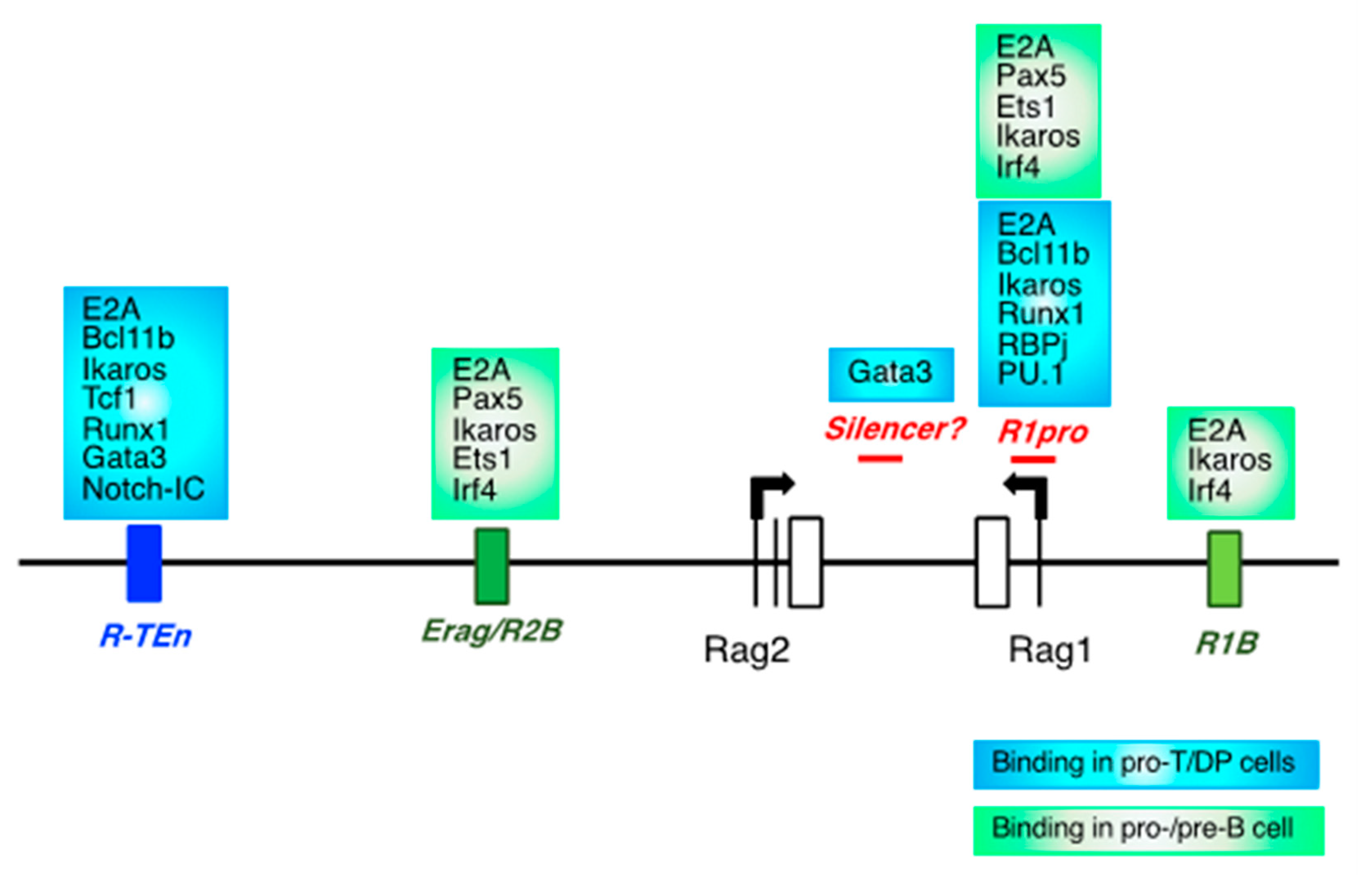
2.1. CREs for the Rag Gene, and Lineage-Specific Transcription Factors
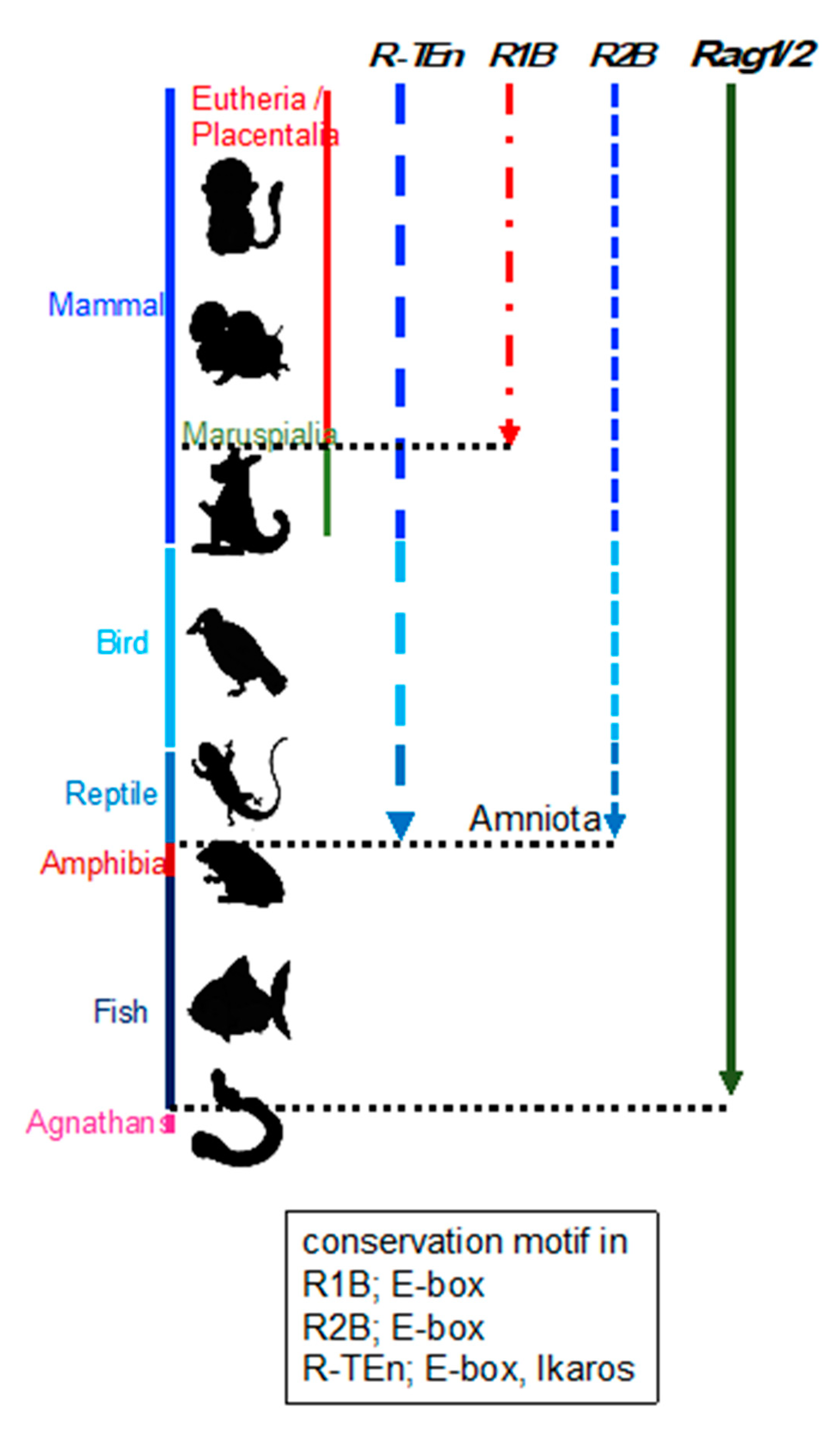
2.2. Evolution of Rag Gene Enhancer
3. E Proteins and Id Proteins in Adaptive Lymphocyte Development
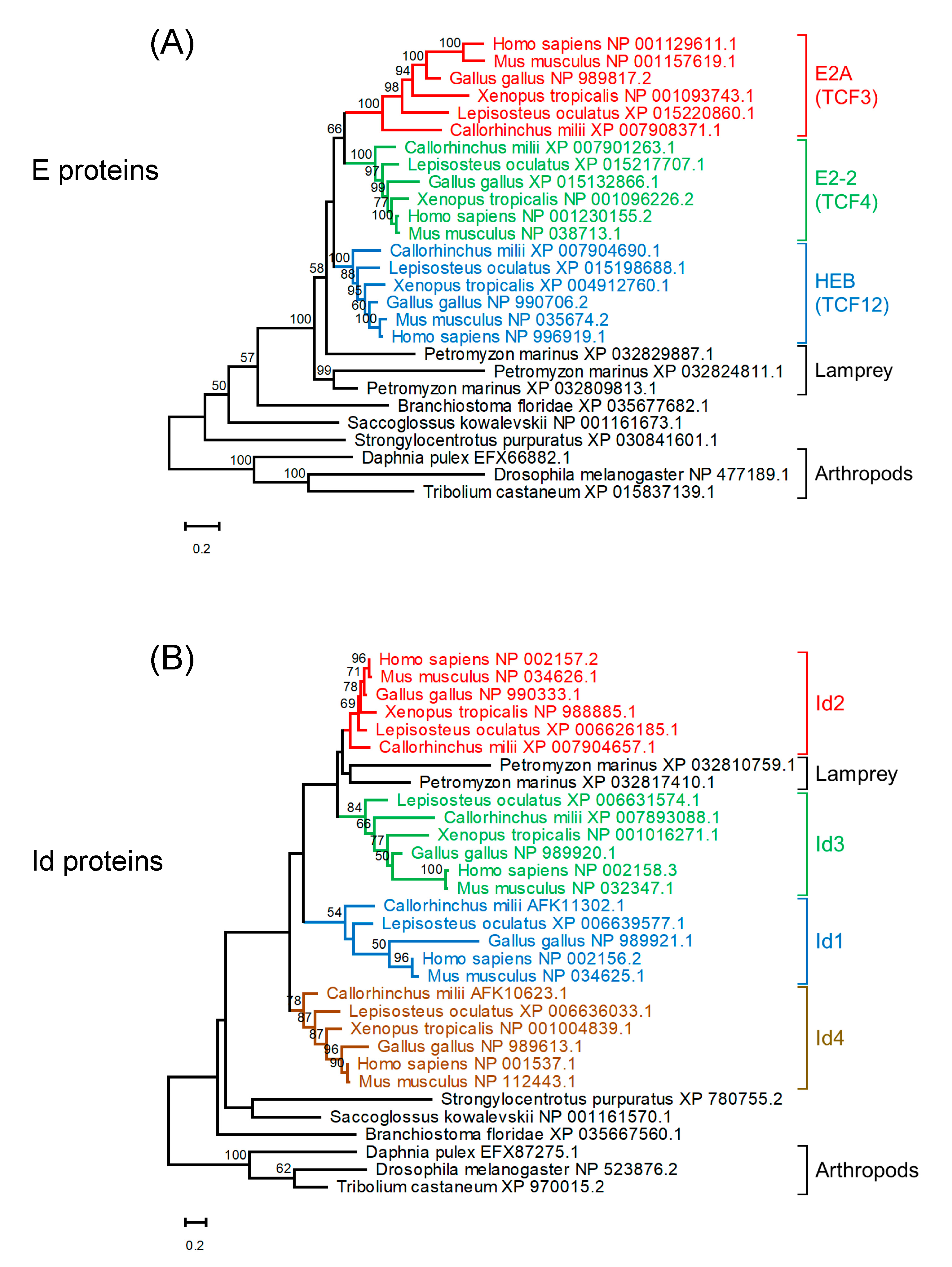
4. Evolution of E and Id Proteins
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tonegawa, S. Somatic generation of antibody diversity. Nature 1983, 302, 575–581. [Google Scholar] [CrossRef]
- Schatz, D.G.; Oettinger, M.A.; Baltimore, D. The V(D)J recombination activating gene, RAG-1. Cell 1989, 59, 1035–1048. [Google Scholar] [CrossRef]
- Schatz, D.G.; Ji, Y. Recombination centres and the orchestration of V(D)J recombination. Nat. Rev. Immunol. 2011, 11, 251–263. [Google Scholar] [CrossRef]
- Kuo, T.C.; Schlissel, M.S. Mechanisms controlling expression of the RAG locus during lymphocyte development. Curr. Opin. Immunol. 2009, 21, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Fugmann, S.D. The origins of the Rag genes—From transposition to V(D)J recombination. Semin. Immunol. 2010, 22, 10–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmona, L.M.; Schatz, D.G. New insights into the evolutionary origins of the recombination-activating gene proteins and V(D)J recombination. FEBS J. 2017, 284, 1590–1605. [Google Scholar] [CrossRef]
- Huang, S.; Tao, X.; Yuan, S.; Zhang, Y.; Li, P.; Beilinson, H.; Zhang, Y.; Yu, W.; Pontarotti, P.; Escriva, H.; et al. Discovery of an Active RAG Transposon Illuminates the Origins of V(D)J Recombination. Cell 2016, 166, 102–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Cheng, T.C.; Huang, G.; Lu, Q.; Surleac, M.D.; Mandell, J.D.; Pontarotti, P.; Petrescu, A.J.; Xu, A.; Xiong, Y.; et al. Transposon molecular domestication and the evolution of the RAG recombinase. Nat. Cell Biol. 2019, 569, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Rothenberg, E.V. Programming for T-lymphocyte fates: Modularity and mechanisms. Genes Dev. 2019, 33, 1117–1135. [Google Scholar] [CrossRef] [Green Version]
- Muljo, S.A.; Schlissel, M.S. Pre-B and pre-T-cell receptors: Conservation of strategies in regulating early lymphocyte development. Immunol. Rev. 2000, 175, 80–93. [Google Scholar] [CrossRef]
- Mandel, E.M.; Grosschedl, R. Transcription control of early B cell differentiation. Curr. Opin. Immunol. 2010, 22, 161–167. [Google Scholar] [CrossRef]
- Lin, Y.C.; Jhunjhunwala, S.; Benner, C.; Heinz, S.; Welinder, E.; Mansson, R.; Sigvardsson, M.; Hagman, J.; Espinoza, C.; Dutkowski, J.; et al. A global network of transcription factors, involving E2A, EBF1 and Foxo1, that orchestrates B cell fate. Nat. Immunol. 2010, 11, 635–643. [Google Scholar] [CrossRef] [Green Version]
- Serafini, N.; Vosshenrich, C.A.J.; Di Santo, J.P. Transcriptional regulation of innate lymphoid cell fate. Nat. Rev. Immunol. 2015, 15, 415–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, K.; Miyazaki, M. The interplay between chromatin architecture and lineage-specific transcription factors and the regulation of Rag gen expression. Front. Immunol. 2021, 12, 6597612021. [Google Scholar] [CrossRef] [PubMed]
- Boller, S.; Li, R.; Grosschedl, R. Defining B Cell Chromatin: Lessons from EBF1. Trends Genet. 2018, 34, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, H.; Rothenberg, E.V. How transcription factors drive choice of the T cell fate. Nat. Rev. Immunol. 2021, 21, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Eberl, G.; Colonna, M.; Di Santo, J.P.; McKenzie, A.N.J. Innate lymphoid cells: A new paradigm in immunology. Science 2015, 348, aaa6566. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, M.; Miyazaki, K.; Chen, K.; Jin, Y.; Turner, J.; Moore, A.J.; Saito, R.; Yoshida, K.; Ogawa, S.; Rodewald, H.-R.; et al. The E-Id Protein Axis Specifies Adaptive Lymphoid Cell Identity and Suppresses Thymic Innate Lymphoid Cell Development. Immunity 2017, 46, 818–834.e4. [Google Scholar] [CrossRef] [Green Version]
- Nechanitzky, R.; Akbas, D.; Scherer, S.; Györy, I.; Hoyler, T.; Ramamoorthy, S.; Diefenbach, A.; Grosschedl, R. Transcription factor EBF1 is essential for the maintenance of B cell identity and prevention of alternative fates in committed cells. Nat. Immunol. 2013, 14, 867–875. [Google Scholar] [CrossRef]
- Yannoutsos, N.; Barreto, V.M.; Misulovin, Z.; Gazumyan, A.; Yu, W.; Rajewsky, N.; Peixoto, B.R.; Eisenreich, T.; Nussenzweig, M.C. A cis element in the recombination activating gene locus regulates gene expression by counteracting a distant silencer. Nat. Immunol. 2004, 5, 443–450. [Google Scholar] [CrossRef]
- Miyazaki, K.; Watanabe, H.; Yoshikawa, G.; Chen, K.; Hidaka, R.; Aitani, Y.; Osawa, K.; Takeda, R.; Ochi, Y.; Tani-Ichi, S.; et al. The transcription factor E2A activates multiple enhancers that drive Rag expression in developing T and B cells. Sci. Immunol. 2020, 5, eabb1455. [Google Scholar] [CrossRef] [PubMed]
- Hsu, L.-Y.; Lauring, J.; Liang, H.-E.; Greenbaum, S.; Cado, D.; Zhuang, Y.; Schlissel, M.S. A Conserved Transcriptional Enhancer Regulates RAG Gene Expression in Developing B Cells. Immunity 2003, 19, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Murre, C. Helix-loop-helix proteins and the advent of cellular diversity: 30 years of discovery. Genes Dev. 2019, 33, 6–25. [Google Scholar] [CrossRef] [Green Version]
- Wilson, A.; Held, W.; Macdonald, H.R. Two waves of recombinase gene expression in developing thymocytes. J. Exp. Med. 1994, 179, 1355–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krangel, M.S. Mechanics of T cell receptor gene rearrangement. Curr. Opin. Immunol. 2009, 21, 133–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mombaerts, P.; Iacomini, J.; Johnson, R.S.; Herrup, K.; Tonegawa, S.; Papaioannou, V.E. RAG-1-deficient mice have no mature B and T lymphocytes. Cell 1992, 68, 869–877. [Google Scholar] [CrossRef]
- Shinkai, Y.; Rathbun, G.; Lam, K.P.; Oltz, E.M.; Stewart, V.; Mendelsohn, M.; Charron, J.; Datta, M.; Young, F.; Stall, A.M.; et al. RAG-2-deficient mice lack mature lymphocytes owing to inability to initiate V(D)J rearrangement. Cell 1992, 68, 855–867. [Google Scholar] [CrossRef]
- Wayne, J.; Suh, H.; A Sokol, K.; Petrie, H.T.; Witmer-Pack, M.; Edelhoff, S.; Disteche, C.M.; Nussenzweig, M.C. TCR selection and allelic exclusion in RAG transgenic mice that exhibit abnormal T cell localization in lymph nodes and lymphatics. J. Immunol. 1994, 153, 5491–5502. [Google Scholar] [PubMed]
- Hao, B.; Naik, A.; Watanabe, A.; Tanaka, H.; Chen, L.; Richards, H.W.; Kondo, M.; Taniuchi, I.; Kohwi, Y.; Kohwi-Shigematsu, T.; et al. An anti-silencer– and SATB1-dependent chromatin hub regulates Rag1 and Rag2 gene expression during thymocyte development. J. Exp. Med. 2015, 212, 809–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naik, A.; Byrd, A.T.; Lucander, A.C.; Krangel, M.S. Hierarchical assembly and disassembly of a transcriptionally active RAG locus in CD4+CD8+ thymocytes. J. Exp. Med. 2019, 216, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Amin, R.H.; Schlissel, M.S. Foxo1 directly regulates the transcription of recombination-activating genes during B cell development. Nat. Immunol. 2008, 9, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Long, H.K.; Prescott, S.L.; Wysocka, J. Ever-Changing Landscapes: Transcriptional Enhancers in Development and Evolution. Cell 2016, 167, 1170–1187. [Google Scholar] [CrossRef] [Green Version]
- Rogozin, I.B.; Iyer, L.M.; Liang, L.; Glazko, G.V.; Liston, V.G.; I Pavlov, Y.; Aravind, L.; Pancer, Z. Evolution and diversification of lamprey antigen receptors: Evidence for involvement of an AID-APOBEC family cytosine deaminase. Nat. Immunol. 2007, 8, 647–656. [Google Scholar] [CrossRef]
- Trancoso, I.; Morimoto, R.; Boehm, T. Co-evolution of mutagenic genome editors and vertebrate adaptive immunity. Curr. Opin. Immunol. 2020, 65, 32–41. [Google Scholar] [CrossRef]
- Morimoto, R.; O’Meara, C.P.; Holland, S.J.; Trancoso, I.; Souissi, A.; Schorpp, M.; Vassaux, D.; Iwanami, N.; Giorgetti, O.B.; Evanno, G.; et al. Cytidine deaminase 2 is required for VLRB antibody gene assembly in lampreys. Sci. Immunol. 2020, 5, eaba0925. [Google Scholar] [CrossRef]
- Hsu, E.; Pulham, N.; Rumfelt, L.L.; Flajnik, M.F. The plasticity of immunoglobulin gene systems in evolution. Immunol. Rev. 2006, 210, 8–26. [Google Scholar] [CrossRef]
- Caudy, M.; Vässin, H.; Brand, M.; Tuma, R.; Jah, L.Y.; Jan, Y.N. daughterless, a Drosophila gene essential for both neurogenesis and sex determination, has sequence similarities to myc and the achaete-scute complex. Cell 1988, 55, 1061–1067. [Google Scholar] [CrossRef]
- Sallee, M.D.; Littleford, H.E.; Greenwald, I. A bHLH Code for Sexually Dimorphic Form and Function of the C. elegans Somatic Gonad. Curr. Biol. 2017, 27, 1853–1860.e5. [Google Scholar] [CrossRef] [Green Version]
- Kee, B.L. E and ID proteins branch out. Nat. Rev. Immunol. 2009, 9, 175–184. [Google Scholar] [CrossRef]
- Benezra, R.; Davis, R.L.; Lockshon, D.; Turner, D.L.; Weintraub, H. The protein Id: A negative regulator of helix-loop-helix DNA binding proteins. Cell 1990, 61, 49–59. [Google Scholar] [CrossRef]
- Belle, I.; Zhuang, Y. E Proteins in Lymphocyte Development and Lymphoid Diseases. Curr. Top. Dev. Biol. 2014, 110, 153–187. [Google Scholar] [CrossRef]
- Bain, G.; Maandag, E.C.; Izon, D.J.; Amsen, D.; Kruisbeek, A.M.; Weintraub, B.C.; Krop, I.; Schlissel, M.S.; Feeney, A.J.; Van Roon, M.; et al. E2A proteins are required for proper B cell development and initiation of immunoglobulin gene rearrangements. Cell 1994, 79, 885–892. [Google Scholar] [CrossRef]
- Zhuang, Y.; Soriano, P.; Weintraub, H. The helix-loop-helix gene E2A is required for B cell formation. Cell 1994, 79, 875–884. [Google Scholar] [CrossRef]
- Murre, C.; McCaw, P.S.; Baltimore, D. A new DNA binding and dimerization motif in immunoglobulin enhancer binding, daughterless, MyoD, and myc proteins. Cell 1989, 56, 777–783. [Google Scholar] [CrossRef]
- Yamazaki, T.; Liu, L.; Lazarev, D.; Al-Zain, A.; Fomin, V.; Yeung, P.L.; Chambers, S.M.; Lu, C.-W.; Studer, L.; Manley, J.L. TCF3 alternative splicing controlled by hnRNP H/F regulates E-cadherin expression and hESC pluripotency. Genes Dev. 2018, 32, 1161–1174. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.C.; Benner, C.; Mansson, R.; Heinz, S.; Miyazaki, K.; Miyazaki, M.; Chandra, V.; Bossen, C.; Glass, C.K.; Murre, C. Global changes in the nuclear positioning of genes and intra- and interdomain genomic interactions that orchestrate B cell fate. Nat. Immunol. 2012, 13, 1196–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’cruz, L.M.; Knell, J.; Fujimoto, J.K.; Goldrath, A.W. An essential role for the transcription factor HEB in thymocyte survival, Tcra rearrangement and the development of natural killer T cells. Nat. Immunol. 2010, 11, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.E.; Zhuang, Y. Acquisition of a Functional T Cell Receptor during T Lymphocyte Development Is Enforced by HEB and E2A Transcription Factors. Immunity 2007, 27, 860–870. [Google Scholar] [CrossRef] [Green Version]
- Cisse, B.; Caton, M.L.; Lehner, M.; Maeda, T.; Scheu, S.; Locksley, R.; Holmberg, D.; Zweier, C.; Hollander, N.S.D.; Kant, S.G.; et al. Transcription Factor E2-2 Is an Essential and Specific Regulator of Plasmacytoid Dendritic Cell Development. Cell 2008, 135, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, H.S.; Cisse, B.; Bunin, A.; Lewis, K.L.; Reizis, B. Continuous Expression of the Transcription Factor E2-2 Maintains the Cell Fate of Mature Plasmacytoid Dendritic Cells. Immunity 2010, 33, 905–916. [Google Scholar] [CrossRef] [Green Version]
- Grajkowska, L.T.; Ceribelli, M.; Lau, C.M.; Warren, M.E.; Tiniakou, I.; Higa, S.N.; Bunin, A.; Haecker, H.; Mirny, L.A.; Staudt, L.M.; et al. Isoform-Specific Expression and Feedback Regulation of E Protein TCF4 Control Dendritic Cell Lineage Specification. Immunity 2017, 46, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Verykokakis, M.; Zook, E.C.; Kee, B.L. ID’ing innate and innate-like lymphoid cells. Immunol. Rev. 2014, 261, 177–197. [Google Scholar] [CrossRef] [Green Version]
- Kee, B.L.; Rivera, R.R.; Murre, C. Id3 inhibits B lymphocyte progenitor growth and survival in response to TGF-beta. Nat. Immunol. 2001, 2, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Miyazaki, M.; Chandra, V.; Fisch, K.M.; Chang, A.N.; Murre, C. Id3 Orchestrates Germinal Center B Cell Development. Mol. Cell. Biol. 2016, 36, 2543–2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gloury, R.; Zotos, D.; Zuidscherwoude, M.; Masson, F.; Liao, Y.; Hasbold, J.; Corcoran, L.M.; Hodgkin, P.D.; Belz, G.T.; Shi, W.; et al. Dynamic changes in Id3 and E-protein activity orchestrate germinal center and plasma cell development. J. Exp. Med. 2016, 213, 1095–1111. [Google Scholar] [CrossRef]
- Miyazaki, M.; Rivera, R.R.; Miyazaki, K.; Lin, Y.C.; Agata, Y.; Murre, C. The opposing roles of the transcription factor E2A and its antagonist Id3 that orchestrate and enforce the naive fate of T cells. Nat. Immunol. 2011, 12, 992–1001. [Google Scholar] [CrossRef]
- Engel, I.; Murre, C. E2A proteins enforce a proliferation checkpoint in developing thymocytes. EMBO J. 2003, 23, 202–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritsen, J.P.; Wong, G.W.; Lee, S.Y.; Lefebvre, J.M.; Ciofani, M.; Rhodes, M.; Kappes, D.J.; Zúñiga-Pflücker, J.C.; Wiest, D.L. Marked induction of the helix-loop-helix protein Id3 promotes the gammadelta T cell fate and renders their functional maturation Notch independent. Immunity 2009, 31, 565–575. [Google Scholar] [CrossRef] [Green Version]
- Fahl, S.P.; Contreras, A.V.; Verma, A.; Qiu, X.; Harly, C.; Radtke, F.; Zúñiga-Pflücker, J.C.; Murre, C.; Xue, H.H.; Sen, J.M.; et al. The E protein-TCF1 axis controls γδ T cell development and effector fate. Cell Rep. 2021, 34, 108716. [Google Scholar] [CrossRef]
- Miyazaki, M.; Miyazaki, K.; Chen, S.; Chandra, V.; Wagatsuma, K.; Agata, Y.; Rodewald, H.-R.; Saito, R.; Chang, A.N.; Varki, N. The E–Id protein axis modulates the activities of the PI3K–AKT–mTORC1–Hif1a and c-myc/p19Arf pathways to suppress innate variant TFH cell development, thymocyte expansion, and lymphomagenesis. Genes Dev. 2015, 29, 409–425. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Chen, X.; Zhong, B.; Wang, A.; Wang, X.; Chu, F.; Nurieva, R.I.; Yan, X.; Chen, P.; Van Der Flier, L.G.; et al. Transcription factor achaete-scute homologue 2 initiates follicular T-helper-cell development. Nat. Cell Biol. 2014, 507, 513–518. [Google Scholar] [CrossRef]
- Leong, Y.A.; Chen, Y.; Ong, H.S.; Wu, D.; Man, K.; Deleage, C.; Minnich, M.; Meckiff, B.J.; Wei, Y.; Hou, Z.; et al. CXCR5+ follicular cytotoxic T cells control viral infection in B cell follicles. Nat. Immunol. 2016, 17, 1187–1196. [Google Scholar] [CrossRef]
- Yang, C.Y.; Best, J.A.; Knell, J.; Yang, E.; Sheridan, A.D.; Jesionek, A.K.; Li, H.S.; Rivera, R.R.; Lind, K.C.; D’Cruz, L.M.; et al. The transcriptional regulators Id2 and Id3 control the formation of distinct memory CD8+ T cell subsets. Nat. Immunol. 2011, 12, 1221–1229. [Google Scholar] [CrossRef]
- Omilusik, K.D.; Shaw, L.A.; Goldrath, A.W. Remembering one’s ID/E-ntity: E/ID protein regulation of T cell memory. Curr. Opin. Immunol. 2013, 25, 660–666. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, M.; Miyazaki, K.; Chen, S.; Itoi, M.; Miller, M.; Lu, L.-F.; Varki, N.; Chang, A.N.; Broide, D.H.; Murre, C. Id2 and Id3 maintain the regulatory T cell pool to suppress inflammatory disease. Nat. Immunol. 2014, 15, 767–776. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Baker, N.E. A Network of Broadly Expressed HLH Genes Regulates Tissue-Specific Cell Fates. Cell 2011, 147, 881–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simakov, O.; Marlétaz, F.; Yue, J.-X.; O’Connell, B.; Jenkins, J.; Brandt, A.; Calef, R.; Tung, C.-H.; Huang, T.-K.; Schmutz, J.; et al. Deeply conserved synteny resolves early events in vertebrate evolution. Nat. Ecol. Evol. 2020, 4, 820–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Cannon, J.P.; Haire, R.N.; Rast, J.P.; Litman, G.W. The phylogenetic origins of the antigen-binding receptors and somatic diversification mechanisms. Immunol. Rev. 2004, 200, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Mayer, W.E.; Uinuk-Ool, T.; Tichy, H.; Gartland, L.A.; Klein, J.; Cooper, M.D. Isolation and characterization of lymphocyte-like cells from a lamprey. Proc. Natl. Acad. Sci. USA 2002, 99, 14350–14355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, P.; Hirano, M.; Herrin, B.R.; Li, J.; Yu, C.; Sadlonova, A.; Cooper, M.D. Dual nature of the adaptive immune system in lampreys. Nat. Cell Biol. 2009, 459, 796–801. [Google Scholar] [CrossRef] [Green Version]
- Boehm, T.; Hirano, M.; Holland, S.J.; Das, S.; Schorpp, M.; Cooper, M.D. Evolution of Alternative Adaptive Immune Systems in Vertebrates. Annu. Rev. Immunol. 2018, 36, 19–42. [Google Scholar] [CrossRef]
- Rothenberg, E.V.; Pant, R. Origins of lymphocyte developmental programs: Transcription factor evidence. Semin. Immunol. 2004, 16, 227–238. [Google Scholar] [CrossRef]
- Cooper, M.D.; Alder, M.N. The Evolution of Adaptive Immune Systems. Cell 2006, 124, 815–822. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Shin, I.T.; Kohara, Y.; Kasahara, M. Transcriptome analysis of hagfish leukocytes: A framework for understanding the immune system of jawless fishes. Dev. Comp. Immunol. 2004, 28, 993–1003. [Google Scholar] [CrossRef]
- Pancer, Z.; Amemiya, C.T.; Ehrhardt, G.R.A.; Ceitlin, J.; Gartland, G.L.; Cooper, M.D. Somatic diversification of variable lymphocyte receptors in the agnathan sea lamprey. Nat. Cell Biol. 2004, 430, 174–180. [Google Scholar] [CrossRef]
- Muramatsu, M.; Kinoshita, K.; Fagarasan, S.; Yamada, S.; Shinkai, Y.; Honjo, T. Class Switch Recombination and Hypermutation Require Activation-Induced Cytidine Deaminase (AID), a Potential RNA Editing Enzyme. Cell 2000, 102, 553–563. [Google Scholar] [CrossRef] [Green Version]
- Müller, V.; De Boer, R.J.; Bonhoeffer, S.; Szathmáry, E. An evolutionary perspective on the systems of adaptive immunity. Biol. Rev. 2018, 93, 505–528. [Google Scholar] [CrossRef] [Green Version]
- Kapitonov, V.V.; Koonin, E.V. Evolution of the RAG1-RAG2 locus: Both proteins came from the same transposon. Biol. Direct 2015, 10, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kasahara, M. What do the paralogous regions in the genome tell us about the origin of the adaptive immune system? Immunol. Rev. 1998, 166, 159–175. [Google Scholar] [CrossRef]
- Kasahara, M.; Suzuki, T.; Du Pasquier, L. On the origins of the adaptive immune system: Novel insights from invertebrates and cold-blooded vertebrates. Trends Immunol. 2004, 25, 105–111. [Google Scholar] [CrossRef]
- Okada, K.; Asai, K. Expansion of signaling genes for adaptive immune system evolution in early vertebrates. BMC Genom. 2008, 9, 218. [Google Scholar] [CrossRef] [Green Version]
- Jones-Mason, M.E.; Zhao, X.; Kappes, D.; Lasorella, A.; Iavarone, A.; Zhuang, Y. E Protein Transcription Factors Are Required for the Development of CD4+ Lineage T Cells. Immunity 2012, 36, 348–361. [Google Scholar] [CrossRef] [Green Version]
- Wöhner, M.; Tagoh, H.; Bilic, I.; Jaritz, M.; Poliakova, D.K.; Fischer, M.; Busslinger, M. Molecular functions of the transcription factors E2A and E2-2 in controlling germinal center B cell and plasma cell development. J. Exp. Med. 2016, 213, 1201–1221. [Google Scholar] [CrossRef] [Green Version]
- Litman, G.W.; Rast, J.P.; Fugmann, S.D. The origins of vertebrate adaptive immunity. Nat. Rev. Immunol. 2010, 10, 543–553. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshikawa, G.; Miyazaki, K.; Ogata, H.; Miyazaki, M. The Evolution of Rag Gene Enhancers and Transcription Factor E and Id Proteins in the Adaptive Immune System. Int. J. Mol. Sci. 2021, 22, 5888. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115888
Yoshikawa G, Miyazaki K, Ogata H, Miyazaki M. The Evolution of Rag Gene Enhancers and Transcription Factor E and Id Proteins in the Adaptive Immune System. International Journal of Molecular Sciences. 2021; 22(11):5888. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115888
Chicago/Turabian StyleYoshikawa, Genki, Kazuko Miyazaki, Hiroyuki Ogata, and Masaki Miyazaki. 2021. "The Evolution of Rag Gene Enhancers and Transcription Factor E and Id Proteins in the Adaptive Immune System" International Journal of Molecular Sciences 22, no. 11: 5888. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115888