
The Combined Influence of Magnesium and Insulin on Central Metabolic Functions and Expression of Genes Involved in Magnesium Homeostasis of Cultured Bovine Adipocytes

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effect of Insulin and Mg on the Content of Non-Polar Lipids (Lipid/Nuclei Ratio)
2.2. Influence of Insulin and Magnesium on Glucose Uptake of Adipocytes (Glucose/Nuclei Ratio)
2.3. Influence of Insulin and Magnesium on GPDH Activity of Adipocytes
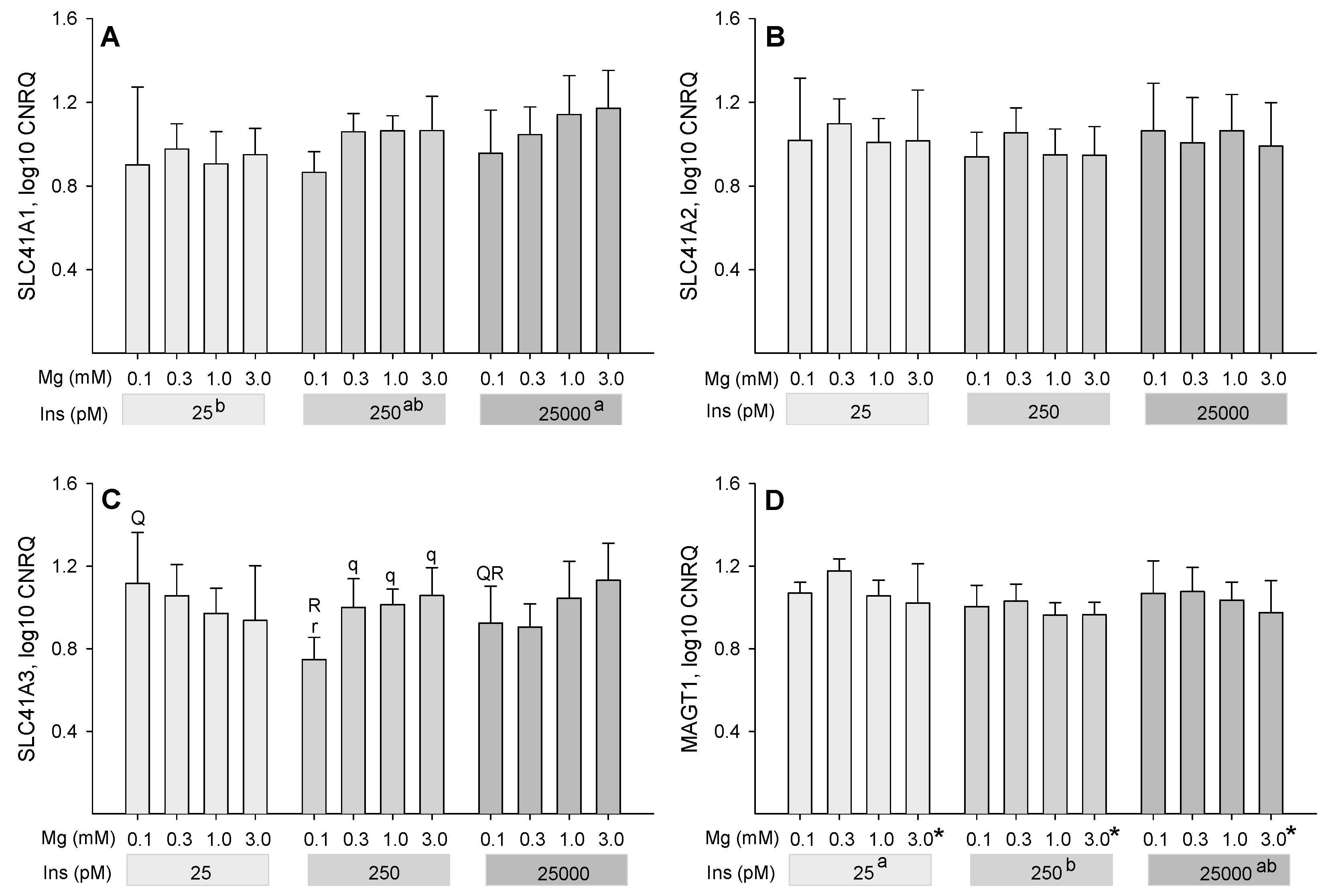
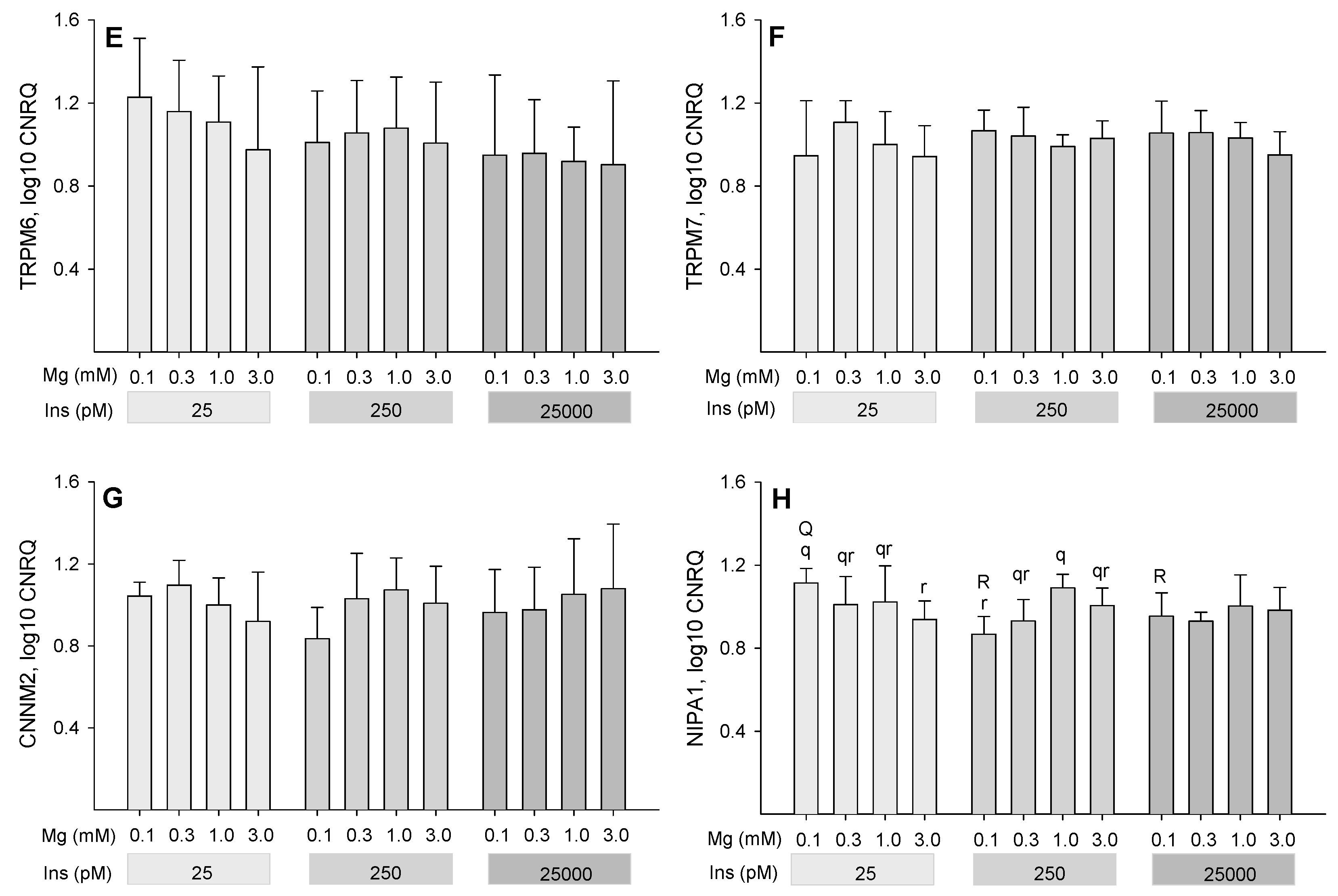
2.4. Influence of Insulin and Magnesium on the Expression of Magnesium-Responsive Genes
3. Discussion
4. Materials and Methods
4.1. Adipose Tissue Collection
4.2. Induction and Differentiation of Adipocytes
4.3. General Procedures
4.3.1. Measurement of Non-Polar Lipids
4.3.2. Measurement of 6-NBDG Uptake
4.3.3. Glycerol 3-Phosphate Dehydrogenase (GPDH) Assay
4.3.4. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR) of Magnesium-Responsive Genes
4.4. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 6-NBDG | 6-(N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino)-6-deoxyglucose |
| ACTB | β-actin |
| Akt (PKB) | protein kinase B |
| ANOVA | analysis of variance |
| BSL | bovine serum lipids |
| cAMP | cyclic adenosine monophosphate |
| cDNA | complementary DNA |
| CNNM2 | cyclin and CBS domain divalent metal cation transport mediator 2 |
| CNRQ | calibrated normalized relative quantity |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| DPBS | Dulbecco’s phosphate-buffered saline |
| em. | emission |
| ex. | excitation |
| FBS | fetal bovine serum |
| GLUT | glucose transporter |
| GPDH | glycerol 3-phosphate dehydrogenase |
| HEPES | 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid |
| Ins | insulin |
| IRC | inter-run calibrator |
| IRS | insulin receptor substrate |
| LSM | least square means |
| MAGT1 | magnesium transporter 1 |
| NAD+ | nicotinamide adenine dinucleotide |
| NADH | reduced nicotinamide adenine dinucleotide |
| NIPA1 | Nipa magnesium transporter 1 |
| NTC | no template control |
| PDE3b | phosphodiesterase 3b |
| PDK1 | phosphoinositide-dependent kinase-1 |
| PI3K | phosphoinositide 3-kinase |
| PKA | protein kinase A |
| RIN | RNA integrity number |
| RNA | ribonucleic acid |
| ROX | carboxy-X-rhodamine |
| RPS19 | ribosomal protein S19 |
| RT-qPCR | reverse-transcription quantitative polymerase chain reaction |
| SD | standard deviation |
| SEM | standard error of mean |
| SLC | solute carrier family |
| STT3B | STT3 oligosaccharyltransferase complex catalytic subunit B |
| TRPM | transient receptor potential cation channel subfamily M |
| YWHAZ | tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein zeta |
References
- Contreras, G.A.; Sordillo, L.M. Lipid mobilization and inflammatory responses during the transition period of dairy cows. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 281–289. [Google Scholar] [CrossRef]
- Oikawa, S.; Oetzel, G.R. Decreased insulin response in dairy cows following a four-day fast to induce hepatic lipidosis. J. Dairy Sci. 2006, 89, 2999–3005. [Google Scholar] [CrossRef]
- De Koster, J.D.; Opsomer, G. Insulin resistance in dairy cows. Vet. Clin. N. Am. Food Anim. Pract. 2013, 29, 299–322. [Google Scholar] [CrossRef]
- Aschenbach, J.R.; Kristensen, N.B.; Donkin, S.S.; Hammon, H.M.; Penner, G.B. Gluconeogenesis in dairy cows: The secret of making sweet milk from sour dough. IUBMB Life 2010, 62, 869–877. [Google Scholar] [CrossRef]
- Sundrum, A. Metabolic disorders in the transition period indicate that the dairy cows’ ability to adapt is overstressed. Animals 2015, 5, 978–1020. [Google Scholar] [CrossRef] [PubMed]
- Contreras, G.A.; Strieder-Barboza, C.; Raphael, W. Adipose tissue lipolysis and remodeling during the transition period of dairy cows. J. Anim. Sci. Biotechnol. 2017, 8, 41. [Google Scholar] [CrossRef] [Green Version]
- Oetzel, G. Understanding the Impact of Subclinical Ketosis. Available online: https://animal.ifas.ufl.edu/apps/dairymedia/rns/2013/2_oetzel.pdf (accessed on 28 May 2021).
- Ospina, P.A.; Nydam, D.V.; Stokol, T.; Overton, T.R. Associations of elevated nonesterified fatty acids and beta-hydroxybutyrate concentrations with early lactation reproductive performance and milk production in transition dairy cattle in the northeastern United States. J. Dairy Sci. 2010, 93, 1596–1603. [Google Scholar] [CrossRef] [Green Version]
- Leno, B.M.; LaCount, S.E.; Ryan, C.M.; Briggs, D.; Crombie, M.; Overton, T.R. The effect of source of supplemental dietary calcium and magnesium in the peripartum period, and level of dietary magnesium postpartum, on mineral status, performance, and energy metabolites in multiparous Holstein cows. J. Dairy Sci. 2017, 100, 7183–7197. [Google Scholar] [CrossRef] [Green Version]
- Erdman, R.A.; Botts, R.L.; Hemken, R.W.; Bull, L.S. Effect of dietary sodium bicarbonate and magnesium oxide on production and physiology in early lactation. J. Dairy Sci. 1980, 63, 923–930. [Google Scholar] [CrossRef]
- Ataollahi, F.; Friend, M.; McGrath, S.; Bhanugopan, M. Effect of maternal mineral supplementation with calcium and magnesium on immune response and energy regulation of ewes and their lambs. In Proceedings of the 2018 MerinoLink Annual Conference and Field Day, Goulburn, Australia, 20 June 2018. [Google Scholar]
- Ahmed, M.H.; Wilkens, M.R.; Ganter, M.; Breves, G. Serum parameters related to mineral homeostasis and energy metabolism in ewes kept on different dietary magnesium supply during the transition period. Res. Vet. Sci. 2021, 134, 19–26. [Google Scholar] [CrossRef]
- Sano, H.; Takebayashi, A.; Kodama, Y.; Nakamura, K.; Ito, H.; Arino, Y.; Fujita, T.; Takahashi, H.; Ambo, K. Effects of feed restriction and cold exposure on glucose metabolism in response to feeding and insulin in sheep. J. Anim. Sci. 1999, 77, 2564–2573. [Google Scholar] [CrossRef] [PubMed]
- Matsunobu, S.; Terashima, Y.; Senshu, T.; Sano, H.; Itoh, H. Insulin secretion and glucose uptake in hypomagnesemic sheep fed a low magnesium, high potassium diet. J. Nutr. Biochem. 1990, 1, 167–171. [Google Scholar] [CrossRef]
- Günther, T. The biochemical function of Mg²+ in insulin secretion, insulin signal transduction and insulin resistance. Magnes. Res. 2010, 23, 5–18. [Google Scholar] [PubMed]
- Kurstjens, S.; de Baaij, J.H.; Overmars-Bos, C.; van den Munckhof, I.C.; Garzero, V.; de Vries, M.A.; Burggraaf, B.; van Diepen, J.A.; Riksen, N.P.; Rutten, J.H. Increased NEFA levels reduce blood Mg2+ in hypertriacylglycerolaemic states via direct binding of NEFA to Mg2+. Diabetologia 2019, 62, 311–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbagallo, M.; Dominguez, L.J.; Galioto, A.; Ferlisi, A.; Cani, C.; Malfa, L.; Pineo, A.; Busardo, A.; Paolisso, G. Role of magnesium in insulin action, diabetes and cardio-metabolic syndrome X. Mol. Aspects Med. 2003, 24, 39–52. [Google Scholar] [CrossRef]
- Kolisek, M.; Launay, P.; Beck, A.; Sponder, G.; Serafini, N.; Brenkus, M.; Froschauer, E.M.; Martens, H.; Fleig, A.; Schweigel, M. SLC41A1 is a novel mammalian Mg2+ carrier. J. Biol. Chem. 2008, 283, 16235–16247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, H.; Garg, R. Effect of magnesium supplementation on type 2 diabetes associated cardiovascular risk factors: A systematic review and meta-analysis. J. Hum. Nutr. Diet 2017, 30, 621–633. [Google Scholar] [CrossRef]
- Martens, H.; Leonhard-Marek, S.; Röntgen, M.; Stumpff, F. Magnesium homeostasis in cattle: Absorption and excretion. Nutr. Res. Rev. 2018, 31, 114–130. [Google Scholar] [CrossRef]
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose tissue remodeling: Its role in energy metabolism and metabolic disorders. Front. Endocrinol. (Lausanne) 2016, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Weber, M.; Locher, L.; Huber, K.; Kenéz, Á.; Rehage, J.; Tienken, R.; Meyer, U.; Dänicke, S.; Sauerwein, H.; Mielenz, M. Longitudinal changes in adipose tissue of dairy cows from late pregnancy to lactation. Part 1: The adipokines apelin and resistin and their relationship to receptors linked with lipolysis. J. Dairy Sci. 2016, 99, 1549–1559. [Google Scholar] [CrossRef] [Green Version]
- Ji, P.; Drackley, J.K.; Khan, M.J.; Loor, J.J. Inflammation- and lipid metabolism-related gene network expression in visceral and subcutaneous adipose depots of Holstein cows. J. Dairy Sci. 2014, 97, 3441–3448. [Google Scholar] [CrossRef] [Green Version]
- Weber, M.; Locher, L.; Huber, K.; Rehage, J.; Tienken, R.; Meyer, U.; Dänicke, S.; Webb, L.; Sauerwein, H.; Mielenz, M. Longitudinal changes in adipose tissue of dairy cows from late pregnancy to lactation. Part 2: The SIRT-PPARGC1A axis and its relationship with the adiponectin system. J. Dairy Sci. 2016, 99, 1560–1570. [Google Scholar] [CrossRef] [Green Version]
- Jurek, S.; Sandhu, M.A.; Trappe, S.; Bermudez-Pena, M.C.; Kolisek, M.; Sponder, G.; Aschenbach, J.R. Optimizing adipogenic transdifferentiation of bovine mesenchymal stem cells: A prominent role of ascorbic acid in FABP4 induction. Adipocyte 2020, 9, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Laviola, L.; Perrini, S.; Cignarelli, A.; Natalicchio, A.; Leonardini, A.; De Stefano, F.; Cuscito, M.; De Fazio, M.; Memeo, V.; Neri, V.; et al. Insulin signaling in human visceral and subcutaneous adipose tissue in vivo. Diabetes 2006, 55, 952–961. [Google Scholar] [CrossRef]
- Weber, C.; Schäff, C.T.; Kautzsch, U.; Börner, S.; Erdmann, S.; Görs, S.; Röntgen, M.; Sauerwein, H.; Bruckmaier, R.M.; Metges, C.C.; et al. Insulin-dependent glucose metabolism in dairy cows with variable fat mobilization around calving. J. Dairy Sci. 2016, 99, 6665–6679. [Google Scholar] [CrossRef] [PubMed]
- Loftus, T.M.; Kuhajda, F.P.; Lane, M.D. Insulin depletion leads to adipose-specific cell death in obese but not lean mice. Proc. Natl. Acad. Sci. USA 1998, 95, 14168–14172. [Google Scholar] [CrossRef] [Green Version]
- Winegrad, A.I.; Shaw, W.N.; Lukens, F.D.W.; Stadie, W.C. Lipogenesis in adipose tissue. Am. J. Clin. Nutr. 1960, 8, 651–665. [Google Scholar] [CrossRef]
- Abe, H.; Kawakit, Y.; Hodate, K.; Saito, M. Postnatal development of glucose transporter proteins in bovine skeletal muscle and adipose tissue. J. Vet. Med. Sci. 2001, 63, 1071–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaakson, H.; Karis, P.; Ling, K.; Ilves-Luht, A.; Samarütel, J.; Henno, M.; Jõudu, I.; Waldmann, A.; Reimann, E.; Pärn, P.; et al. Adipose tissue insulin receptor and glucose transporter 4 expression, and blood glucose and insulin responses during glucose tolerance tests in transition Holstein cows with different body condition. J. Dairy Sci. 2018, 101, 752–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.Q.; Moseley, W.M.; Tucker, H.A.; Kennelly, J.J. Regulation of glucose transporter gene expression in mammary gland, muscle, and fat of lactating cows by administration of bovine growth hormone and bovine growth hormone-releasing factor. J. Anim. Sci. 1996, 74, 183–189. [Google Scholar] [CrossRef]
- Komatsu, T.; Itoh, F.; Kushibiki, S.; Hodate, K. Changes in gene expression of glucose transporters in lactating and nonlactating cows. J. Anim. Sci. 2005, 83, 557–564. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Q.; Zhang, Z.; Fu, R.; Zhou, T.; Long, C.; He, T.; Yang, D.; Li, Z.; Peng, S. Magnesium supplementation enhances mTOR signalling to facilitate myogenic differentiation and improve aged muscle performance. Bone 2021, 146, 115886. [Google Scholar] [CrossRef] [PubMed]
- Beg, M.; Abdullah, N.; Thowfeik, F.S.; Altorki, N.K.; McGraw, T.E. Distinct Akt phosphorylation states are required for insulin regulated Glut4 and Glut1-mediated glucose uptake. Elife 2017, 6, e26896. [Google Scholar] [CrossRef]
- De Valk, H.W. Magnesium in diabetes mellitus. Neth. J. Med. 1999, 54, 139–146. [Google Scholar] [CrossRef]
- Sales, C.H.; Pedrosa, L.d.F.C. Magnesium and diabetes mellitus: Their relation. Clin. Nutr. 2006, 25, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.K.; Madsen, F.C.; Lentz, D.E.; Wong, W.O.; Ramsey, N.; Tysinger, C.E.; Hansard, S.L. Blood plasma magnesium, potassium, glucose, and immunoreactive insulin changes in cows moved abruptly from barn feeding to early spring pasture. J. Dairy Sci. 1980, 63, 1073–1079. [Google Scholar] [CrossRef]
- Abuelo, A.; Hernández, J.; Benedito, J.L.; Castillo, C. A comparative study of the metabolic profile, insulin sensitivity and inflammatory response between organically and conventionally managed dairy cattle during the periparturient period. Animal 2014, 8, 1516–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moustaïd, N.; Jones, B.H.; Taylor, J.W. Insulin increases lipogenic enzyme activity in human adipocytes in primary culture. J. Nutr. 1996, 126, 865–870. [Google Scholar] [CrossRef]
- Grant, A.; Ortiz-Colon, G.; Doumit, M.; Buskirk, D. Optimization of in vitro conditions for bovine subcutaneous and intramuscular preadipocyte differentiation. J. Anim. Sci. 2008, 86, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Kolisek, M.; Sponder, G.; Pilchova, I.; Cibulka, M.; Tatarkova, Z.; Werner, T.; Racay, P. Magnesium extravaganza: A critical compendium of current research into cellular Mg2+ transporters other than TRPM6/7. Rev. Physiol. Biochem. Pharmacol. 2019, 176, 65–105. [Google Scholar]
- Dai, L.J.; Ritchie, G.; Bapty, B.W.; Kerstan, D.; Quamme, G.A. Insulin stimulates Mg2+ uptake in mouse distal convoluted tubule cells. Am. J. Physiol. 1999, 277, F907–F913. [Google Scholar] [CrossRef]
- Sponder, G.; Mastrototaro, L.; Kurth, K.; Merolle, L.; Zhang, Z.; Abdulhanan, N.; Smorodchenko, A.; Wolf, K.; Fleig, A.; Penner, R. Human CNNM2 is not a Mg2+ transporter per se. Pflugers Arch. 2016, 468, 1223–1240. [Google Scholar] [CrossRef]
- Groenestege, W.M.; Hoenderop, J.G.; van den Heuvel, L.; Knoers, N.; Bindels, R.J. The epithelial Mg2+ channel transient receptor potential melastatin 6 is regulated by dietary Mg2+ content and estrogens. J. Am. Soc. Nephrol. 2006, 17, 1035–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.H.; Xu, X.H.; Liu, Y.; Hu, Y.; Jin, M.W.; Li, G.R. TRPM7 channels regulate proliferation and adipogenesis in 3T3-L1 preadipocytes. J. Cell. Physiol. 2014, 229, 60–67. [Google Scholar] [CrossRef]
- Kolisek, M.; Nestler, A.; Vormann, J.; Schweigel-Röntgen, M. Human gene SLC41A1 encodes for the Na+/Mg2+ exchanger. Am. J. Physiol. Cell Physiol. 2012, 302, C318–C326. [Google Scholar] [CrossRef] [Green Version]
- Mastrototaro, L.; Tietjen, U.; Sponder, G.; Vormann, J.; Aschenbach, J.R.; Kolisek, M. Insulin modulates the Na+/Mg2+ exchanger SLC41A1 and influences Mg2+ efflux from intracellular stores in transgenic HEK293 cells. J. Nutr. 2015, 145, 2440–2447. [Google Scholar] [CrossRef] [Green Version]
- Mastrototaro, L.; Smorodchenko, A.; Aschenbach, J.R.; Kolisek, M.; Sponder, G. Solute carrier 41A3 encodes for a mitochondrial Mg2+ efflux system. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kubota, T.; Shindo, Y.; Tokuno, K.; Komatsu, H.; Ogawa, H.; Kudo, S.; Kitamura, Y.; Suzuki, K.; Oka, K. Mitochondria are intracellular magnesium stores: Investigation by simultaneous fluorescent imagings in PC12 cells. Biochim. Biophys. Acta 2005, 1744, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Cherepanova, N.A.; Gilmore, R. Mammalian cells lacking either the cotranslational or posttranslocational oligosaccharyltransferase complex display substrate-dependent defects in asparagine linked glycosylation. Sci. Rep. 2016, 6, 20946. [Google Scholar] [CrossRef] [Green Version]
- Blommaert, E.; Péanne, R.; Cherepanova, N.A.; Rymen, D.; Staels, F.; Jaeken, J.; Race, V.; Keldermans, L.; Souche, E.; Corveleyn, A.; et al. Mutations in MAGT1 lead to a glycosylation disorder with a variable phenotype. Proc. Natl. Acad. Sci. USA 2019, 116, 9865–9870. [Google Scholar] [CrossRef] [Green Version]
- Goytain, A.; Hines, R.M.; El-Husseini, A.; Quamme, G.A. NIPA1 (SPG6), the basis for autosomal dominant form of hereditary spastic paraplegia, encodes a functional Mg2+ transporter. J. Biol. Chem. 2007, 282, 8060–8068. [Google Scholar] [CrossRef] [Green Version]
- Kurth, K. Validation of Magnesium Sensitive Genes as Biomarkers for the Intracellular Magnesium Status of Patients with Diabetes Mellitus. Ph.D. Thesis, Institute of Veterinary-Physiology Freie Universität Berlin, Berlin, Germany, 30 January 2013. [Google Scholar]
- Sandhu, M.A.; Jurek, S.; Trappe, S.; Kolisek, M.; Sponder, G.; Aschenbach, J.R. Influence of bovine serum lipids and fetal bovine serum on the expression of cell surface markers in cultured bovine preadipocytes. Cells Tissues Organs 2017, 204, 13–24. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ins/Mg | 0.1 MM | 0.3 MM | 1 MM | 3 MM | LSM | SEM | |
|---|---|---|---|---|---|---|---|
| 25 PM | 1.58 | 1.45 | 1.26 | 3.24 | 1.88 | 0.170 | Factor insulin, P = 0.045 Mg × insulin, P = 0.38 |
| 250 PM | 1.01 | 1.12 | 1.35 | 1.52 | 1.25 | 0.129 | |
| 25,000 PM | 1.55 | 1.49 | 1.48 | 1.67 | 1.55 | 0.129 | |
| LSM | 1.38 | 1.35 | 1.36 | 2.14 | |||
| SEM | 0.151 | 0.151 | 0.151 | 0.214 | |||
| Factor Mg, P = 0.037 | |||||||
| Gene | Sense Primer (5′-3′) | nM | Ampl. Size (bp) | Database Accession No. |
|---|---|---|---|---|
| Anti-Sense Primer (3′-5′) | ||||
| Probe | ||||
| SLC41A1 | TGGTGTTCCTCTATACCATCAG | 1000 | 186 | NM_001206036.1 |
| TCAAGTACGGGATGGAGAAG | 3000 | |||
| ATGTAGAGCAGGATCAGCACCTGGAGCAGA | 500 | |||
| SLC41A2 | CTGCTTTTAGTGATACCTGGAC | 500 | 178 | NM_001205910.1 |
| TTCCTTTCCTCCAGAAATGATG | 1500 | |||
| TTGCTGTGGATCGCTGACTGGATG | 150 | |||
| SLC41A3 | CTTCTGCACTATTTCCAGCAC | 1500 | 100 | XM_024983333.1 |
| TCATCTCCAGGTTGCCCTTC | 3000 | |||
| TTCACGGAGATGAAGGACCTGCTGACCTTGG | 500 | |||
| MAGT1 | GCTCAATTTGTAGCTGAAACAC | 500 | 124 | NM_001244318.2 |
| CACACATTATCTTTCGCTTTCC | 1500 | |||
| ATGTGAAGCTGCTACATCTGACATGGATATTG | 150 | |||
| CNNM2 | GCTCCAGAATACTACCTCTACC | 500 | 83 | NM_001191172.1 |
| GCTTCTACTTCACTTTCCCC | 1500 | |||
| CGAAACAAACCTGTAGACTACTTCGTTCTCAT | 150 | |||
| NIPA1 | TCCCCGAAATCTGAGAGTGTG | 1000 | 115 | XM_002685192.6 |
| AGAAGATGAGCAGCAGCAGC | 1500 | |||
| TGGAGGAGAAGCTGACCAATCCAGTGTTTGTG | 150 | |||
| TRPM6 | ACAAACCATTCCCTACACTCC | 500 | 125 | XM_015472505.2 |
| CGTTGTTGTTGTTGTACTTCC | 1500 | |||
| TTGACCATCGAGAAGTATATGACGGGGGAG | 150 | |||
| TRPM7 | ATACAAGAGGGGAGTTACTGG | 500 | 112 | NM_001206166.3 |
| GGGCCAAAAACCATATCACAG | 1500 | |||
| CTGACCCATCTGTGATAAAGGCAGAAGAA | 150 | |||
| RPS19 | GGAAAAGGACCAAGATGGGG | 500 | 136 | NM_001037467.2 |
| CGAACGAGGCAATTTATTAACC | 1500 | |||
| ACAGAGAGATCTGGACAGAATCGCTGGACA | 150 | |||
| YWHAZ | AGAGAGAAAATAGAGACCGAGC | 500 | 144 | NM_174814.2 |
| AGCCAAGTAGCGGTAGTAG | 1500 | |||
| CCAACGCTTCACAAGCAGAGAGCAAA | 150 | |||
| ACTB | GCCAACCGTGAGAAGATGAC | 500 | 124 | NM_173979.3 |
| AGTCCATCACGATGCCAGTG | 1500 | |||
| CCAGATCATGTTTGAGACC TTCAACACCCCTGC | 150 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becker, S.K.; Sponder, G.; Sandhu, M.A.; Trappe, S.; Kolisek, M.; Aschenbach, J.R. The Combined Influence of Magnesium and Insulin on Central Metabolic Functions and Expression of Genes Involved in Magnesium Homeostasis of Cultured Bovine Adipocytes. Int. J. Mol. Sci. 2021, 22, 5897. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115897
Becker SK, Sponder G, Sandhu MA, Trappe S, Kolisek M, Aschenbach JR. The Combined Influence of Magnesium and Insulin on Central Metabolic Functions and Expression of Genes Involved in Magnesium Homeostasis of Cultured Bovine Adipocytes. International Journal of Molecular Sciences. 2021; 22(11):5897. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115897
Chicago/Turabian StyleBecker, Sandra K., Gerhard Sponder, Mansur A. Sandhu, Susanne Trappe, Martin Kolisek, and Jörg R. Aschenbach. 2021. "The Combined Influence of Magnesium and Insulin on Central Metabolic Functions and Expression of Genes Involved in Magnesium Homeostasis of Cultured Bovine Adipocytes" International Journal of Molecular Sciences 22, no. 11: 5897. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115897