
CMAS and ST3GAL4 Play an Important Role in the Adsorption of Influenza Virus by Affecting the Synthesis of Sialic Acid Receptors

 ,
,
Abstract
:1. Introduction
2. Results
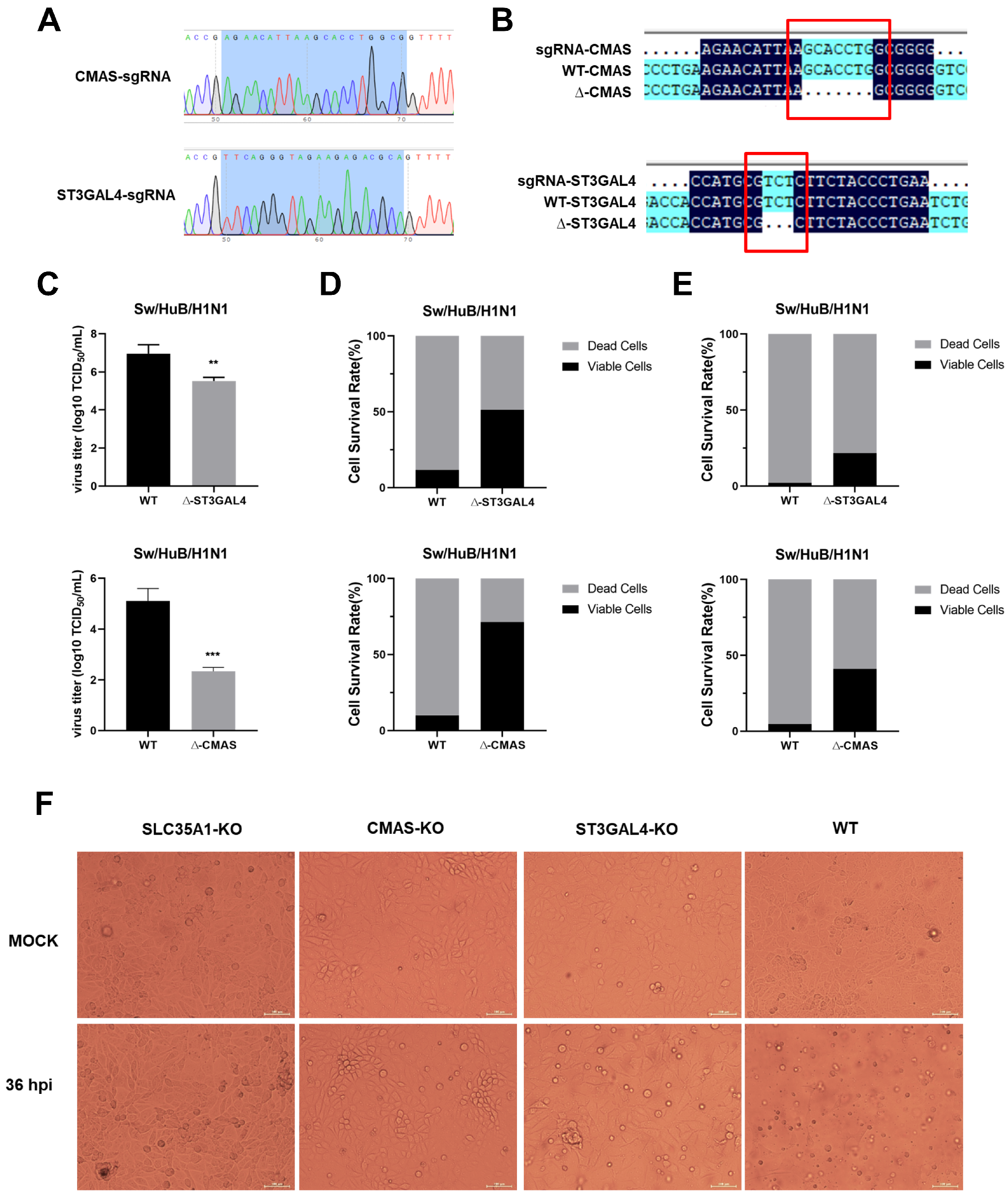
2.1. Generation of Gene Knockout Cell Lines
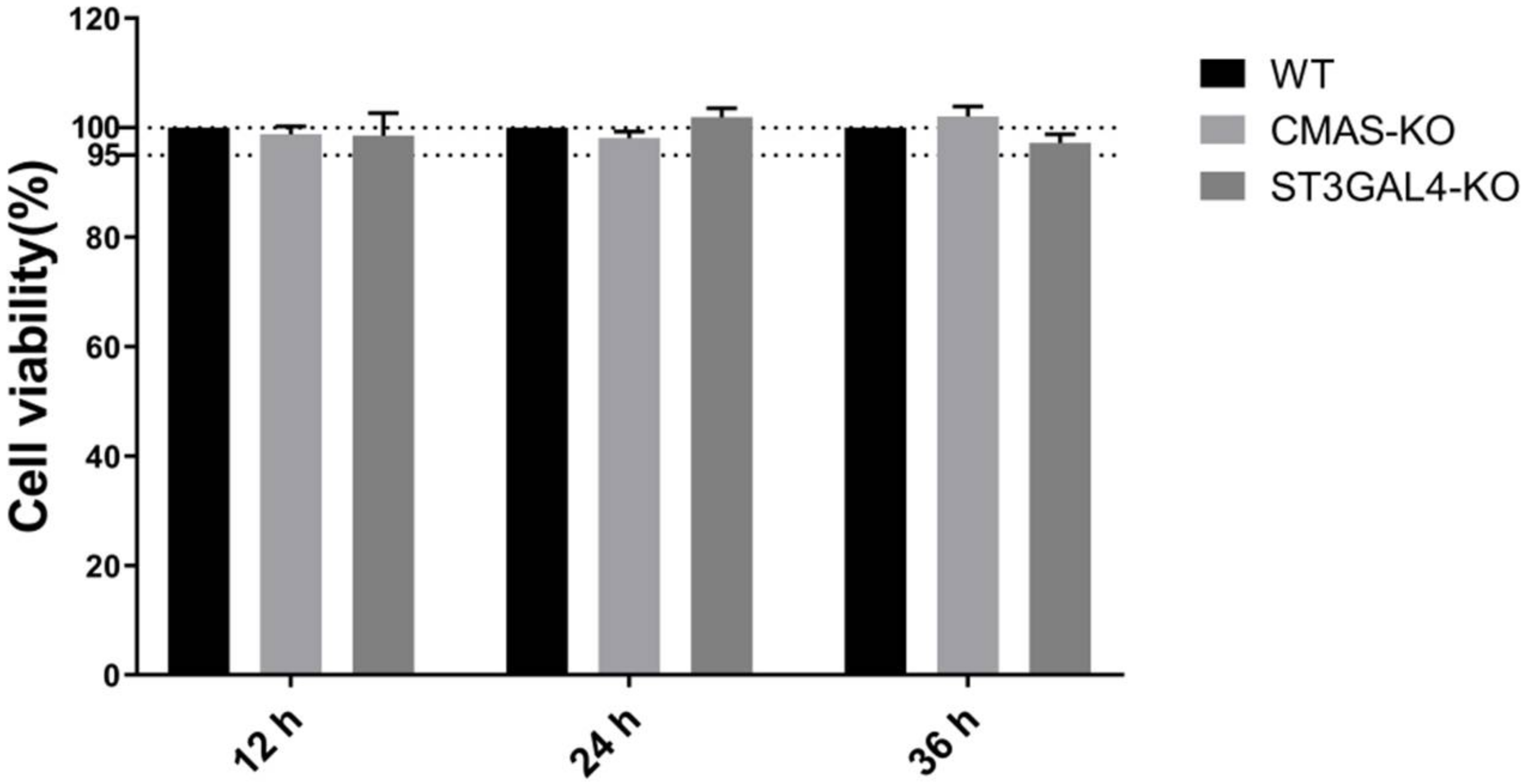
2.2. Knockout of CMAS or ST3GAL4 Has No Effect on Cell Viability
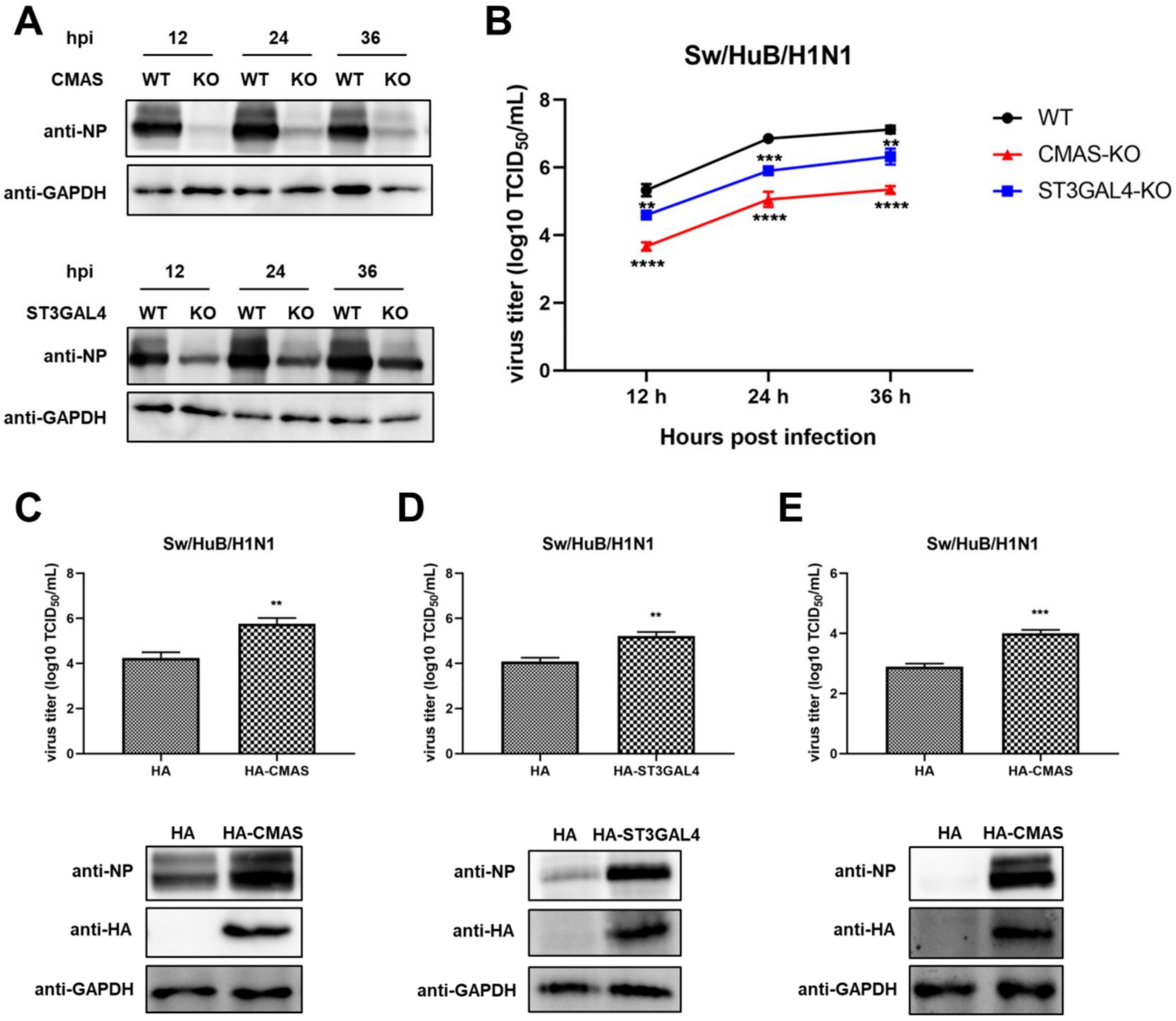
2.3. Knockout of CMAS or ST3GAL4 Negatively Modulates the Proliferation of Swine IAV
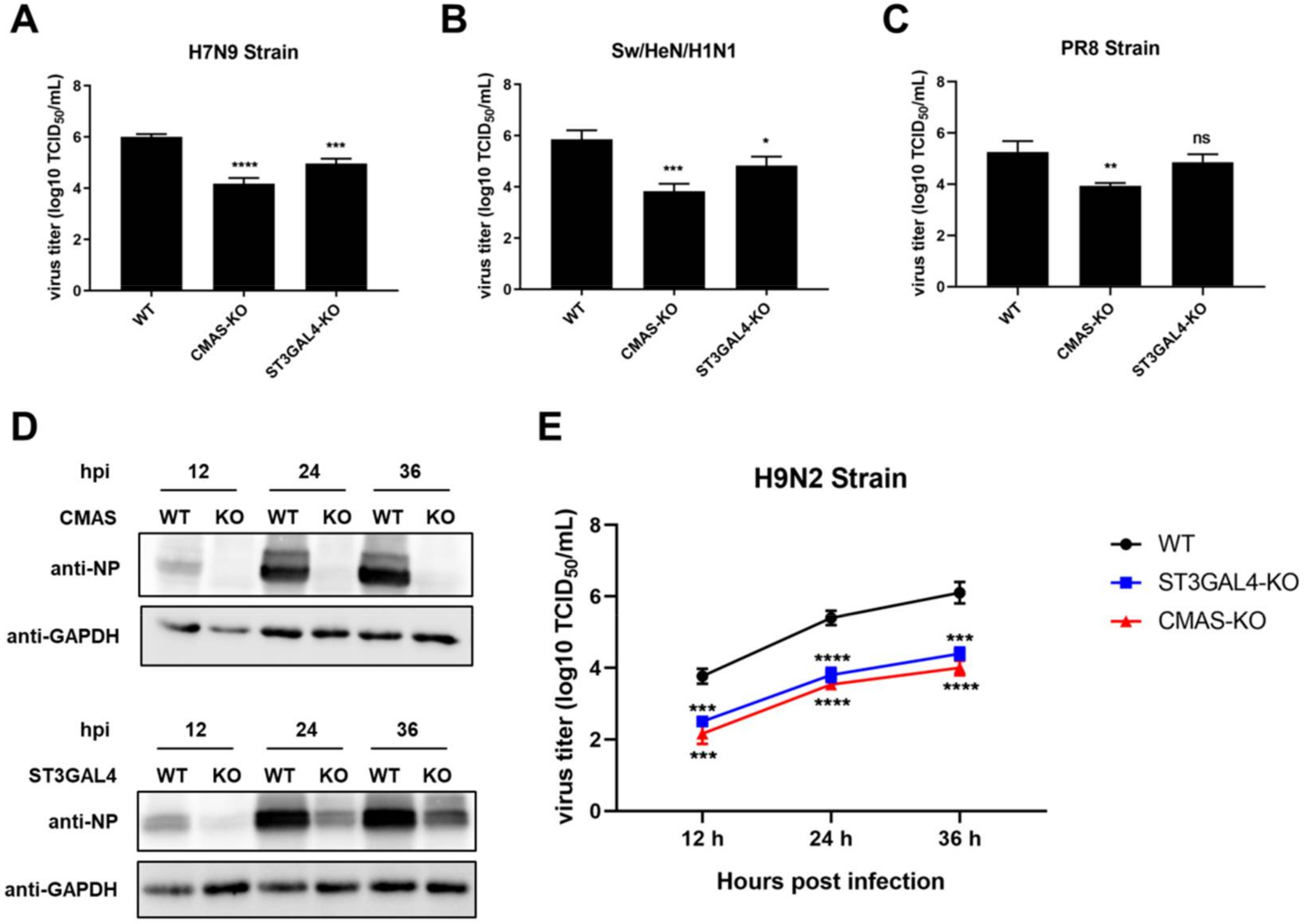
2.4. Effects of Gene Knockout on the Proliferation of IAVs of Different Species
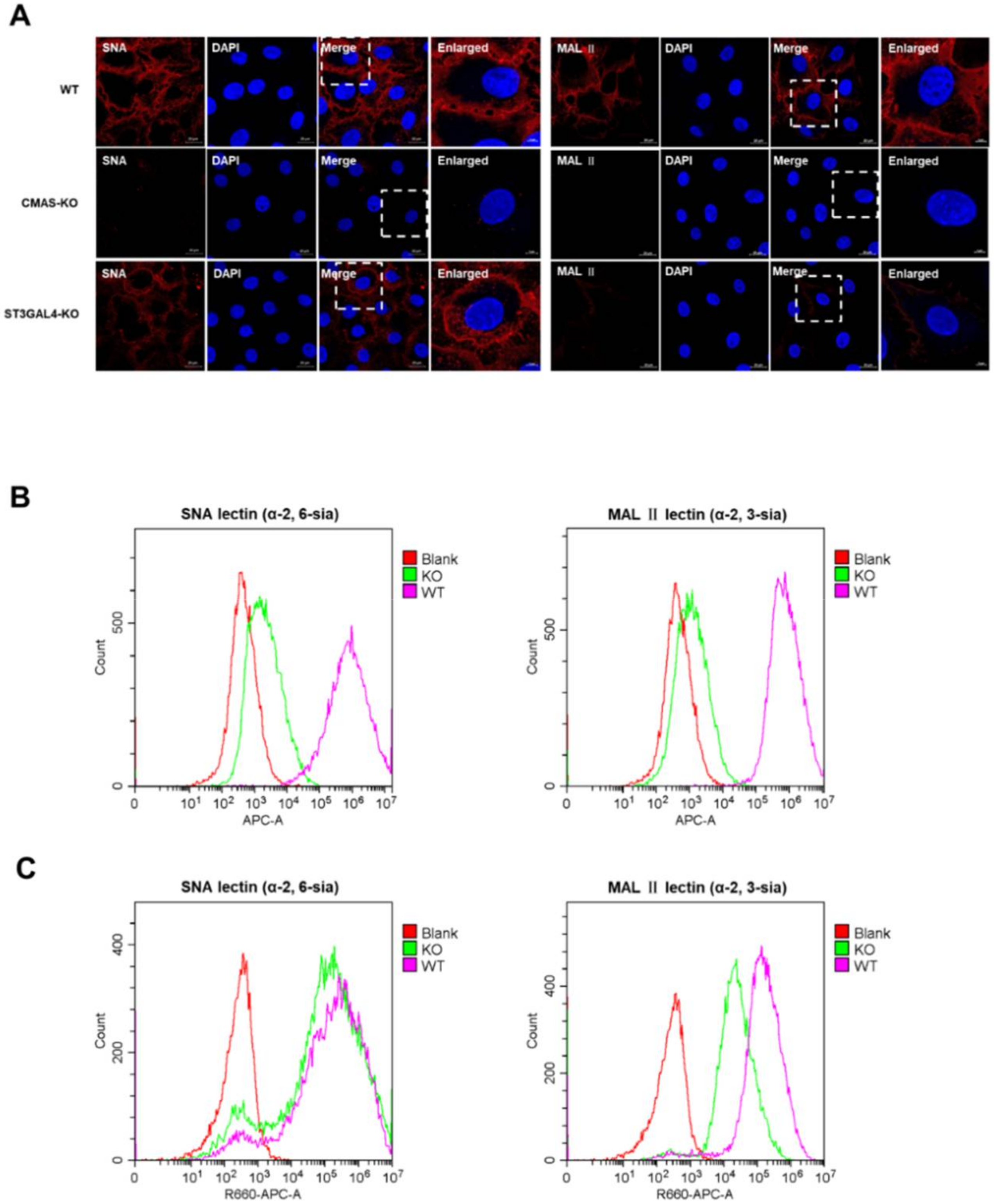
2.5. The Effect of CMAS and ST3GAL4 on the Synthesis of Different Types of Sialic Acid Receptors
2.6. CMAS and ST3GAL4 Are Essential for IAVs Attachment
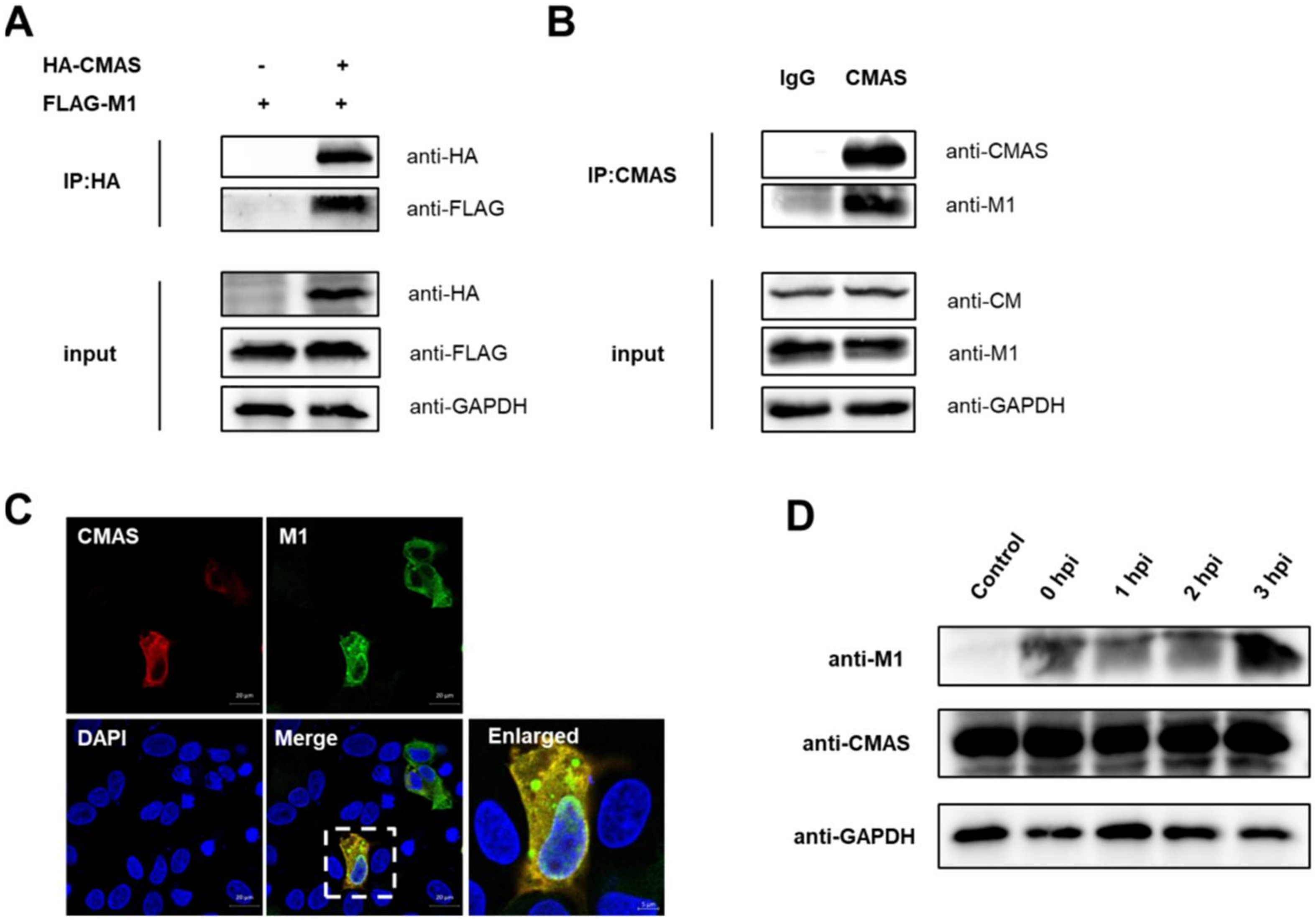
2.7. CMAS Interacts with IAV M1
3. Discussion
4. Materials and Methods
4.1. Cells and Viruses
4.2. Antibodies and Reagents
4.3. Plasmids
4.4. Generation of the KO NPTr Cells
4.5. Transfection
4.6. Virus Titration
4.7. Western Blot Analysis
4.8. Lectin and HA Binding Assays
4.9. Cell Viability Assay
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Medina, R.A.; Garcia-Sastre, A. Influenza A viruses: New research developments. Nat. Rev. Microbiol. 2011, 9, 590–603. [Google Scholar] [CrossRef]
- Brown, I.H. The epidemiology and evolution of influenza viruses in pigs. Vet. Microbiol. 2000, 74, 29–46. [Google Scholar] [CrossRef]
- Petrova, V.N.; Russell, C.A. The evolution of seasonal influenza viruses. Nat. Rev. Microbiol. 2018, 16, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, G.N.; Paulson, J.C. Receptor determinants of human and animal influenza virus isolates: Differences in receptor specificity of the H3 hemagglutinin based on species of origin. Virology 1983, 127, 361–373. [Google Scholar] [CrossRef]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco, S.; Gomez, J.; et al. New world bats harbor diverse influenza A viruses. PLoS Pathog. 2013, 9, e1003657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Clohisey, S.M.; Chia, B.S.; Wang, B.; Cui, A.; Eisenhaure, T.; Schweitzer, L.D.; Hoover, P.; Parkinson, N.J.; Nachshon, A.; et al. Genome-wide CRISPR screen identifies host dependency factors for influenza A virus infection. Nat. Commun. 2020, 11, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Dang, Y.; Wu, Y.; Jia, G.; Anaya, E.; Zhang, J.; Abraham, S.; Choi, J.G.; Shi, G.; Qi, L.; et al. A CRISPR-Based Screen Identifies Genes Essential for West-Nile-Virus-Induced Cell Death. Cell Rep. 2015, 12, 673–683. [Google Scholar] [CrossRef] [Green Version]
- Sellmeier, M.; Weinhold, B.; Munster-Kuhnel, A. CMP-Sialic Acid Synthetase: The Point of Constriction in the Sialylation Pathway. Top. Curr. Chem. 2015, 366, 139–167. [Google Scholar] [CrossRef]
- Gillespie, W.; Kelm, S.; Paulson, J.C. Cloning and expression of the Gal beta 1, 3GalNAc alpha 2,3-sialyltransferase. J. Biol. Chem. 1992, 267, 21004–21010. [Google Scholar] [CrossRef]
- Schauer, R.; Kamerling, J.P. Exploration of the Sialic Acid World. Adv. Carbohydr. Chem. Biochem. 2018, 75, 1–213. [Google Scholar] [CrossRef] [PubMed]
- Varki, A. Essentials of Glycobiology, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2017; p. 823. [Google Scholar]
- Kean, E.L.; Munster-Kuhnel, A.K.; Gerardy-Schahn, R. CMP-sialic acid synthetase of the nucleus. Biochim. Biophys. Acta 2004, 1673, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Bullough, P.A.; Hughson, F.M.; Skehel, J.J.; Wiley, D.C. Structure of influenza haemagglutinin at the pH of membrane fusion. Nature 1994, 371, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Dou, D.; Revol, R.; Ostbye, H.; Wang, H.; Daniels, R. Influenza A Virus Cell Entry, Replication, Virion Assembly and Movement. Front. Immunol. 2018, 9, 1581. [Google Scholar] [CrossRef] [PubMed]
- Bui, M.; Whittaker, G.; Helenius, A. Effect of M1 protein and low pH on nuclear transport of influenza virus ribonucleoproteins. J. Virol. 1996, 70, 8391–8401. [Google Scholar] [CrossRef] [Green Version]
- Matrosovich, M.N.; Gambaryan, A.S.; Teneberg, S.; Piskarev, V.E.; Yamnikova, S.S.; Lvov, D.K.; Robertson, J.S.; Karlsson, K.A. Avian influenza A viruses differ from human viruses by recognition of sialyloligosaccharides and gangliosides and by a higher conservation of the HA receptor-binding site. Virology 1997, 233, 224–234. [Google Scholar] [CrossRef] [Green Version]
- Skehel, J.J.; Wiley, D.C. Receptor binding and membrane fusion in virus entry: The influenza hemagglutinin. Annu. Rev. Biochem. 2000, 69, 531–569. [Google Scholar] [CrossRef]
- Baum, L.G.; Paulson, J.C. Sialyloligosaccharides of the respiratory epithelium in the selection of human influenza virus receptor specificity. Acta Histochem. Suppl. 1990, 40, 35–38. [Google Scholar]
- Ito, T.; Suzuki, Y.; Suzuki, T.; Takada, A.; Horimoto, T.; Wells, K.; Kida, H.; Otsuki, K.; Kiso, M.; Ishida, H.; et al. Recognition of N-glycolylneuraminic acid linked to galactose by the alpha2,3 linkage is associated with intestinal replication of influenza A virus in ducks. J. Virol. 2000, 74, 9300–9305. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Ito, T.; Suzuki, T.; Holland, R.E., Jr.; Chambers, T.M.; Kiso, M.; Ishida, H.; Kawaoka, Y. Sialic acid species as a determinant of the host range of influenza A viruses. J. Virol. 2000, 74, 11825–11831. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Perez, J.T.; Chen, C.; Li, Y.; Benitez, A.; Kandasamy, M.; Lee, Y.; Andrade, J.; TenOever, B.; Manicassamy, B. Genome-wide CRISPR/Cas9 Screen Identifies Host Factors Essential for Influenza Virus Replication. Cell Rep. 2018, 23, 596–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Graaf, M.; Fouchier, R.A. Role of receptor binding specificity in influenza A virus transmission and pathogenesis. EMBO J. 2014, 33, 823–841. [Google Scholar] [CrossRef] [Green Version]
- Van Poucke, S.G.; Nicholls, J.M.; Nauwynck, H.J.; Van Reeth, K. Replication of avian, human and swine influenza viruses in porcine respiratory explants and association with sialic acid distribution. Virol. J. 2010, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Connor, R.J.; Kawaoka, Y.; Webster, R.G.; Paulson, J.C. Receptor specificity in human, avian, and equine H2 and H3 influenza virus isolates. Virology 1994, 205, 17–23. [Google Scholar] [CrossRef]
- Qu, R.; Sang, Q.; Wang, X.; Xu, Y.; Chen, B.; Mu, J.; Zhang, Z.; Jin, L.; He, L.; Wang, L. A homozygous mutation in CMAS causes autosomal recessive intellectual disability in a Kazakh family. Ann. Hum. Genet. 2020, 84, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Stray, S.J.; Cummings, R.D.; Air, G.M. Influenza virus infection of desialylated cells. Glycobiology 2000, 10, 649–658. [Google Scholar] [CrossRef] [Green Version]
- Londrigan, S.L.; Tate, M.D.; Brooks, A.G.; Reading, P.C. Cell-surface receptors on macrophages and dendritic cells for attachment and entry of influenza virus. J. Leukoc. Biol. 2012, 92, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Gambaryan, A.S.; Tuzikov, A.B.; Pazynina, G.V.; Desheva, J.A.; Bovin, N.V.; Matrosovich, M.N.; Klimov, A.I. 6-sulfo sialyl Lewis X is the common receptor determinant recognized by H5, H6, H7 and H9 influenza viruses of terrestrial poultry. Virol. J. 2008, 5, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.T.; Lichtenberg, B.; Rick, O. Involvement of annexin V in the entry of influenza viruses and role of phospholipids in infection. FEBS Lett. 1996, 392, 59–62. [Google Scholar] [CrossRef] [Green Version]
- Kajiwara, N.; Nomura, N.; Ukaji, M.; Yamamoto, N.; Kohara, M.; Yasui, F.; Sakoda, Y.; Kida, H.; Shibasaki, F. Cell-penetrating peptide-mediated cell entry of H5N1 highly pathogenic avian influenza virus. Sci. Rep. 2020, 10, 18008. [Google Scholar] [CrossRef]
- Monteerarat, Y.; Suptawiwat, O.; Boonarkart, C.; Uiprasertkul, M.; Auewarakul, P.; Viprakasit, V. Inhibition of H5N1 highly pathogenic influenza virus by suppressing a specific sialyltransferase. Arch. Virol. 2010, 155, 889–893. [Google Scholar] [CrossRef]
- Saito, S.; Yamashita, S.; Endoh, M.; Yamato, T.; Hoshi, S.; Ohyama, C.; Watanabe, R.; Ito, A.; Satoh, M.; Wada, T.; et al. Clinical significance of ST3Gal IV expression in human renal cell carcinoma. Oncol. Rep. 2002, 9, 1251–1255. [Google Scholar] [CrossRef] [PubMed]
- Roa-de La Cruz, L.; Martinez-Morales, P.; Moran-Cruz, I.; Milflores-Flores, L.; Rosas-Murrieta, N.; Gonzalez-Ramirez, C.; Ortiz-Mateos, C.; Monterrosas-Santamaria, R.; Gonzalez-Frias, C.; Rodea-Avila, C.; et al. Expression analysis of ST3GAL4 transcripts in cervical cancer cells. Mol. Med. Rep. 2018, 18, 617–621. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.H.; Li, Y.F.; Juang, C.M.; Lee, Y.R.; Chao, H.T.; Tsai, Y.C.; Yuan, C.C. Altered mRNA expression of sialyltransferase in squamous cell carcinomas of the cervix. Gynecol. Oncol. 2001, 83, 121–127. [Google Scholar] [CrossRef]
- Torriani, G.; Galan-Navarro, C.; Kunz, S. Lassa Virus Cell Entry Reveals New Aspects of Virus-Host Cell Interaction. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Jae, L.T.; Raaben, M.; Herbert, A.S.; Kuehne, A.I.; Wirchnianski, A.S.; Soh, T.K.; Stubbs, S.H.; Janssen, H.; Damme, M.; Saftig, P.; et al. Virus entry. Lassa virus entry requires a trigger-induced receptor switch. Science 2014, 344, 1506–1510. [Google Scholar] [CrossRef] [Green Version]
- Qi, F.; Isaji, T.; Duan, C.; Yang, J.; Wang, Y.; Fukuda, T.; Gu, J. ST3GAL3, ST3GAL4, and ST3GAL6 differ in their regulation of biological functions via the specificities for the alpha2,3-sialylation of target proteins. FASEB J. 2020, 34, 881–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanjana, N.E.; Shalem, O.; Zhang, F. Improved vectors and genome-wide libraries for CRISPR screening. Nat. Methods 2014, 11, 783–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Yang, C.; Ren, C.; Zhang, S.; Gao, X.; Jin, M.; Chen, H.; Ma, W.; Zhou, H. Eukaryotic Translation Elongation Factor 1 Delta Inhibits the Nuclear Import of the Nucleoprotein and PA-PB1 Heterodimer of Influenza A Virus. J. Virol. 2020, 95. [Google Scholar] [CrossRef]
- Ramakrishnan, M.A. Determination of 50% endpoint titer using a simple formula. World J. Virol. 2016, 5, 85–86. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequences (5′ to 3′) |
|---|---|
| sgRNA Primers | |
| ST3GAL4-sgRNA | F: CACCGTTCAGGGTAGAAGAGACGCATGG |
| R: AAACCCATGCGTCTCTTCTACCCTGAAC | |
| CMAS-sgRNA | F: CACCGAGAACATTAAGCACCTGGCGGGG |
| R: AAACCCCCGCCAGGTGCTTAATGTTCTC | |
| PCR primers to verify mutations | |
| ST3GAL4-YZ | F: CGCCTCCCAGGCTAAACAAT |
| R: GACACAGATCTCAAGTGTTCT | |
| CMAS-YZ | F: TGGGAGGAAGATGGACTCAGT |
| R: GCCTCCTTTCCTTTACCCAAT | |
| PCR primers for gene cloning | |
| ST3GAL4-HA | F: CCATCGATATGGAGGAGGCTGGCCAGAG |
| R: CTAGCTAGCTCAGAAGTATGTGAGGTTCTT | |
| CMAS-HA | F: CCGGAATTCATGCCCCACAAGGGTGAA |
| R: GGGGTACCTTAATTAATAGAAGGGCATGTCTTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Zou, J.; Gao, Q.; Xie, S.; Cao, J.; Zhou, H. CMAS and ST3GAL4 Play an Important Role in the Adsorption of Influenza Virus by Affecting the Synthesis of Sialic Acid Receptors. Int. J. Mol. Sci. 2021, 22, 6081. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116081
Zhao Y, Zou J, Gao Q, Xie S, Cao J, Zhou H. CMAS and ST3GAL4 Play an Important Role in the Adsorption of Influenza Virus by Affecting the Synthesis of Sialic Acid Receptors. International Journal of Molecular Sciences. 2021; 22(11):6081. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116081
Chicago/Turabian StyleZhao, Yaxin, Jiahui Zou, Qingxia Gao, Shengsong Xie, Jiyue Cao, and Hongbo Zhou. 2021. "CMAS and ST3GAL4 Play an Important Role in the Adsorption of Influenza Virus by Affecting the Synthesis of Sialic Acid Receptors" International Journal of Molecular Sciences 22, no. 11: 6081. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116081