
Association of MALAT1 and PVT1 Variants, Expression Profiles and Target miRNA-101 and miRNA-186 with Colorectal Cancer: Correlation with Epithelial-Mesenchymal Transition


Abstract
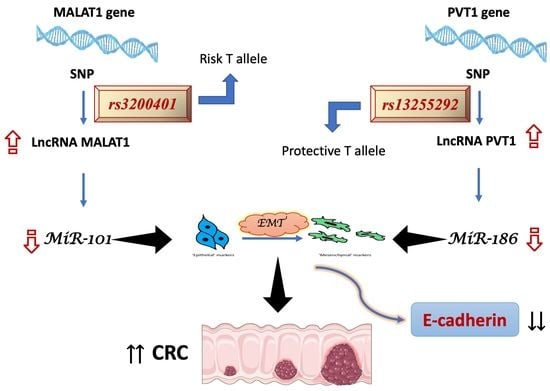
:
1. Introduction
2. Results
2.1. The Studied Groups’ Demographical and Clinicopathological Properties
2.2. Association of rs3200401 (C/T) and rs13255292 (C/T) with the Risk of CRC and AP
2.3. Association of MALAT1 rs3200401 and PVT1 rs13255292 with CRC vs. Non-CRC
2.4. Selection of the Best Fit Models
2.5. Haplotype Analysis
2.6. Serum Expression Levels of MALAT1, PVT1, miRNA-101 and miRNA-186 in CRC and AP
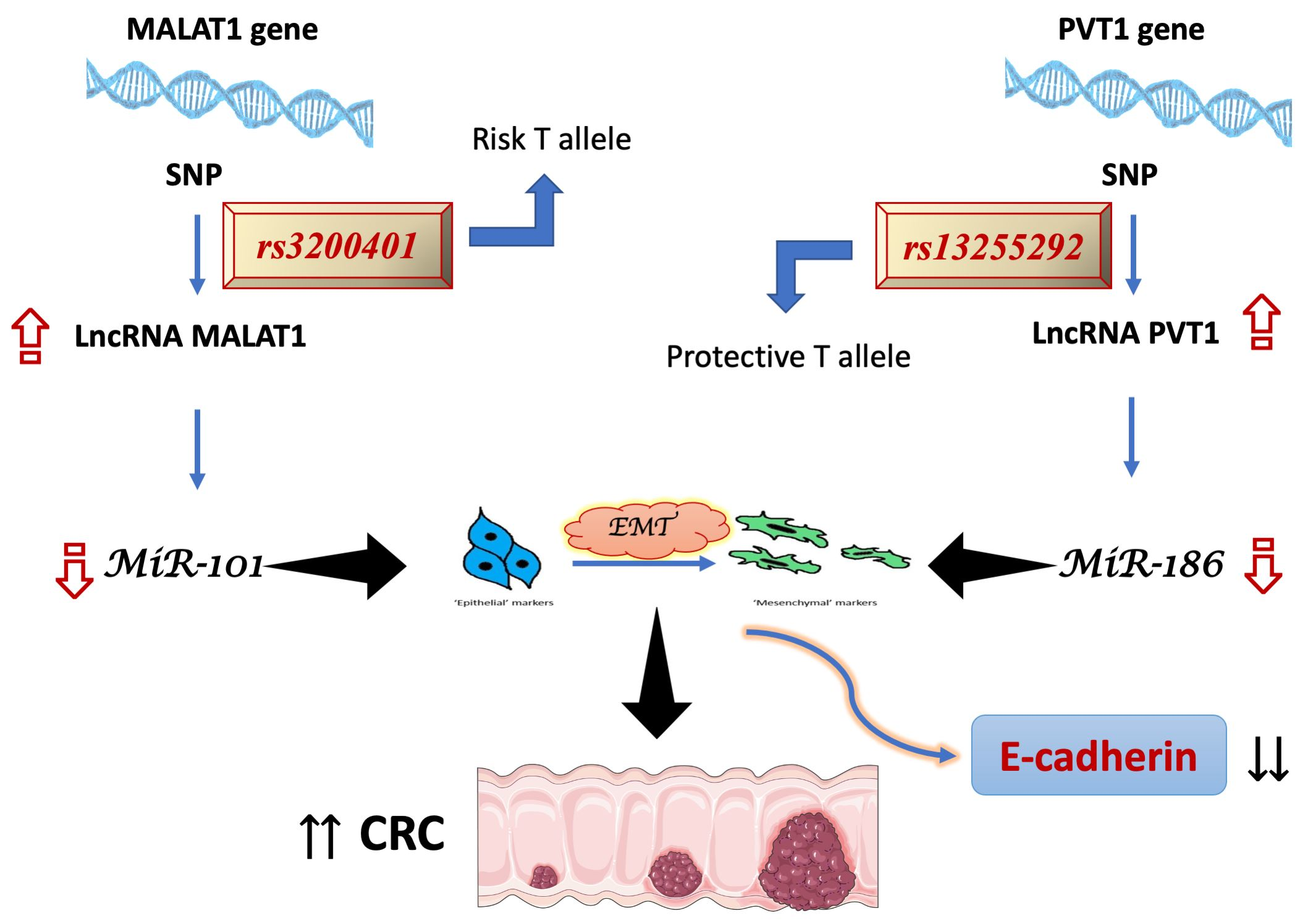
2.7. Serum Levels of E-Cadherin
2.8. Association of rs3200401 and rs13255292 with Serum MALAT1, PVT1, miRNA-101, miRNA-186 and E-Cadherin Levels in CRC Patients
2.9. Correlation between rs3200401 and rs13255292 and the Clinicopathological Characteristics
2.10. Diagnostic Performance of the Studied Parameters between the Studied Groups
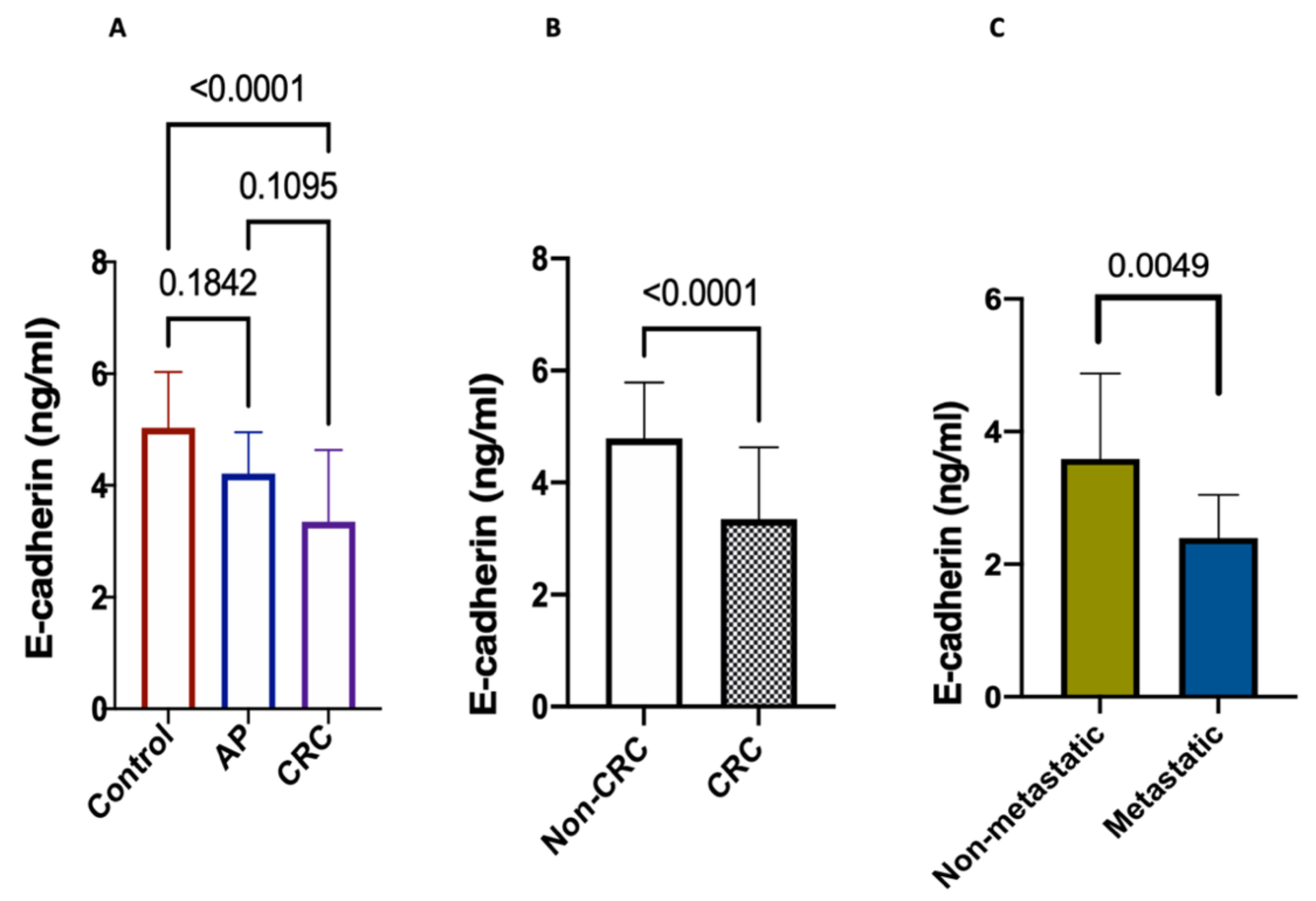
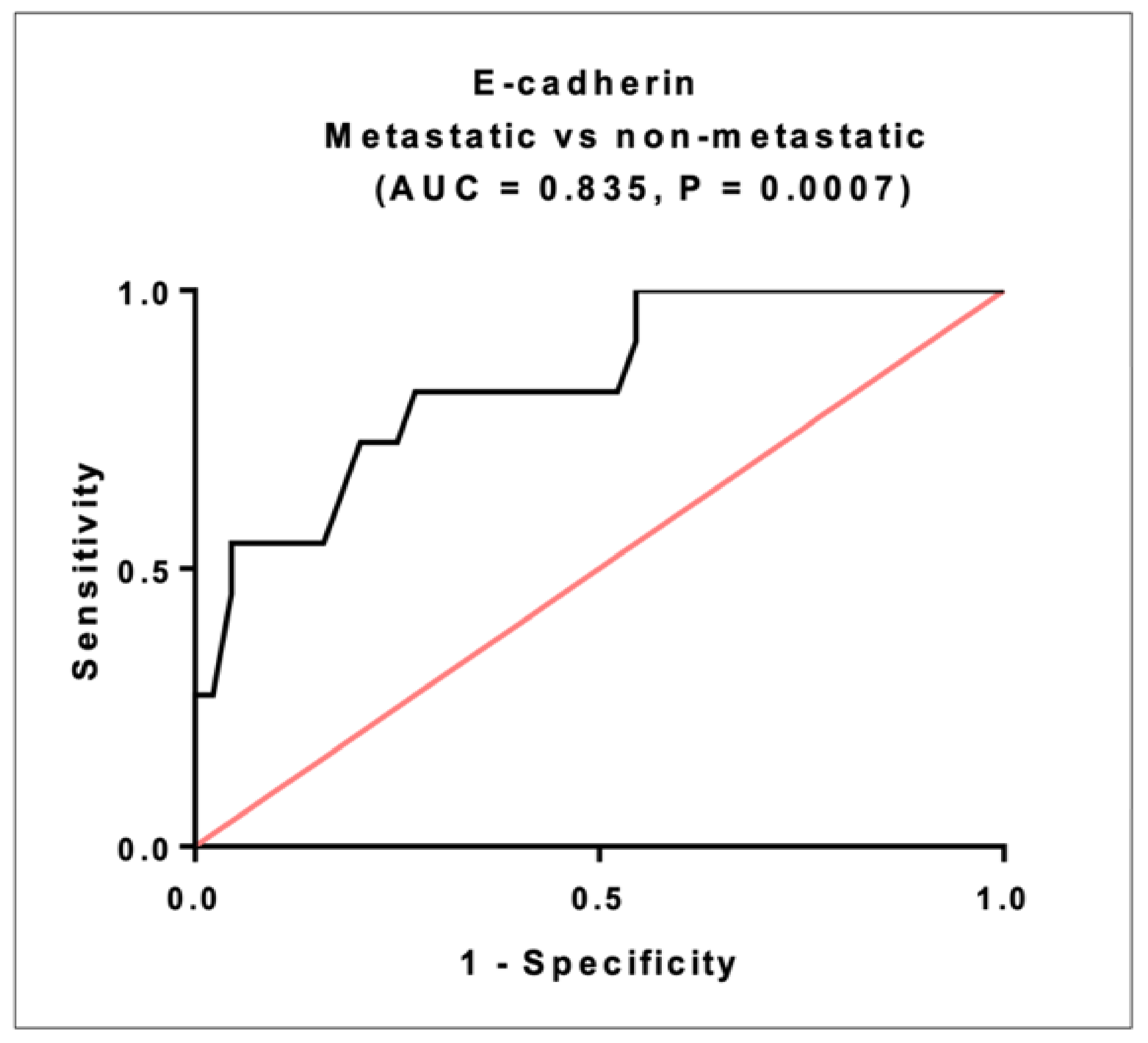
2.11. Prognostic Significance of E-Cadherin in CRC
2.12. Diagnostic Performance of the Studied Parameters between CRC and Non-CRC
2.13. Logistic Regression Analysis of the Studied Parameters
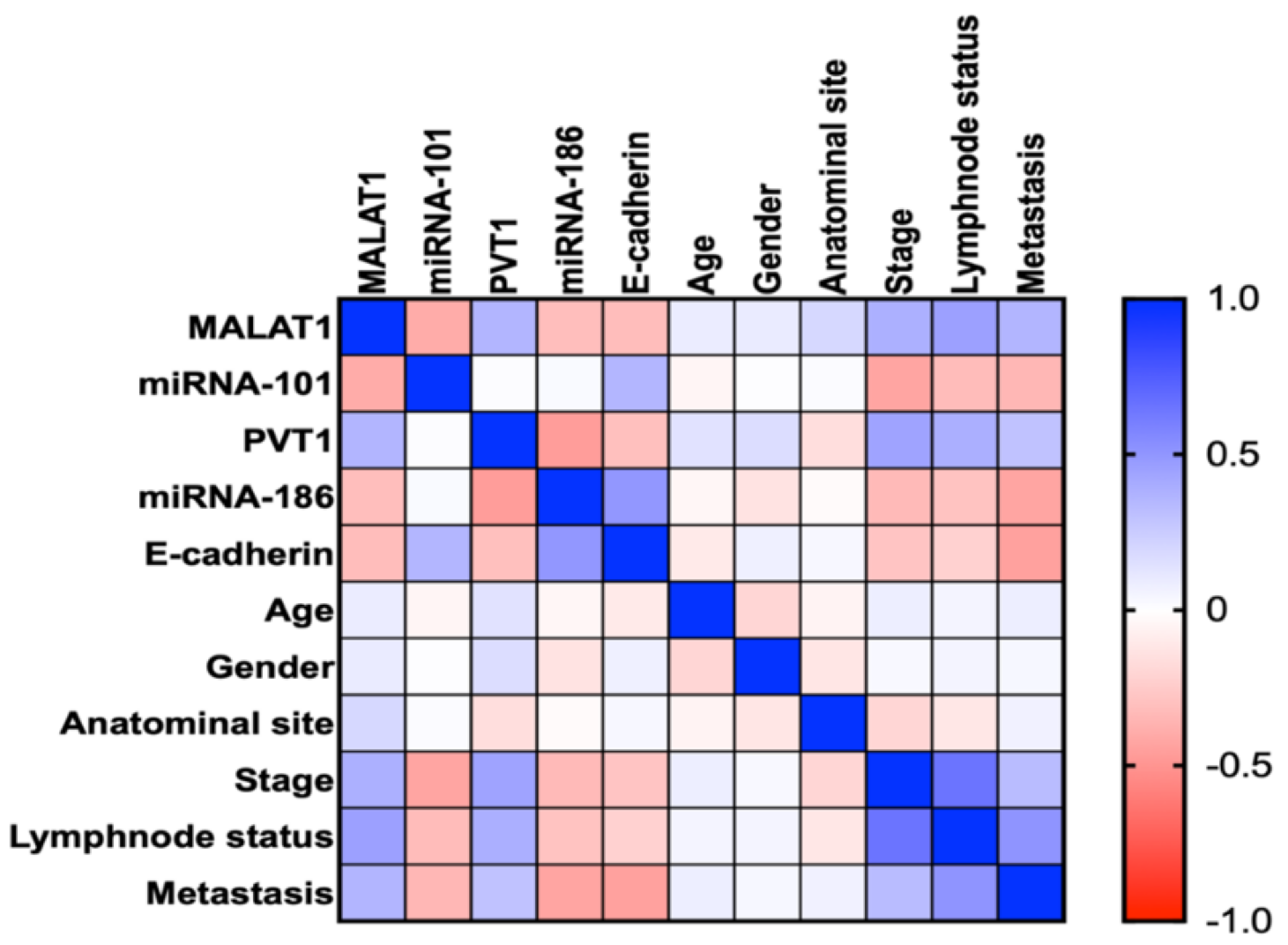
2.14. Correlation between the Studied Parameters and the Clinicopathological Characteristics
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Blood Collection and Storage
4.3. SNPs Position
4.4. Selection of SNPs
4.5. DNA Extraction and Genotyping
4.6. Assay of Serum MALAT1, PVT1, miRNA-186 and miRNA-101 by RT-qPCR
4.7. Assessment of Serum E-Cadherin by Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Statistical Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Mph, K.D.M.; Sauer, A.G.; Fedewa, S.A.; Butterly, L.F.; Anderson, J.C.; Cercek, A.; Smith, R.A.; Jemal, A. Colorectal cancer statistics, 2020. CA A Cancer J. Clin. 2020, 70, 145–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keum, N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef] [PubMed]
- Grillone, K.; Riillo, C.; Scionti, F.; Rocca, R.; Tradigo, G.; Guzzi, P.H.; Alcaro, S.; Di Martino, M.T.; Tagliaferri, P.; Tassone, P. Non-coding RNAs in cancer: Platforms and strategies for investigating the genomic “dark matter”. J. Exp. Clin. Cancer Res. 2020, 39, 1–19. [Google Scholar] [CrossRef]
- Ji, X.; Zhang, J.; Liu, L.; Lin, Z.; Pi, L.; Lin, Z.; Tian, N.; Lin, X.; Chen, S.; Yu, X.; et al. Association of tagSNPs at lncRNA MALAT-1 with HCC Susceptibility in a Southern Chinese Population. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, M.; Zhao, S.; Jiang, Z.; Wang, S.; Sun, P.; Quan, J.; Yan, D.; Wang, X. MALAT1 sponges miR-106b-5p to promote the invasion and metastasis of colorectal cancer via SLAIN2 enhanced microtubules mobility. EBioMedicine 2019, 41, 286–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.-X.; Zhu, Q.-N.; Zhang, H.-B.; Hu, Y.; Wang, G.; Zhu, Y.-S. MALAT1: A potential biomarker in cancer. Cancer Manag. Res. 2018, ume 10, 6757–6768. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-Z.; Deng, F.; Li, H.; Wang, D.-D.; Zhang, W.; Ding, L.; Tang, J.-H. MiR-101: A potential therapeutic target of cancers. Am. J. Transl. Res. 2018, 10, 3310–3321. [Google Scholar]
- Wang, X.; Li, M.; Wang, Z.; Han, S.; Tang, X.; Ge, Y.; Zhou, L.; Zhou, C.; Yuan, Q.; Yang, M. Silencing of Long Noncoding RNA MALAT1 by miR-101 and miR-217 Inhibits Proliferation, Migration, and Invasion of Esophageal Squamous Cell Carcinoma Cells. J. Biol. Chem. 2015, 290, 3925–3935. [Google Scholar] [CrossRef] [Green Version]
- Si, Y.; Yang, Z.; Ge, Q.; Yu, L.; Yao, M.; Sun, X.; Ren, Z.; Ding, C. Long non-coding RNA Malat1 activated autophagy, hence promoting cell proliferation and inhibiting apoptosis by sponging miR-101 in colorectal cancer. Cell. Mol. Biol. Lett. 2019, 24, 1–12. [Google Scholar] [CrossRef]
- Wang, J.; Zeng, H.; Li, H.; Chen, T.; Wang, L.; Zhang, K.; Chen, J.; Wang, R.; Li, Q.; Wang, S. MicroRNA-101 Inhibits Growth, Proliferation and Migration and Induces Apoptosis of Breast Cancer Cells by Targeting Sex-Determining Region Y-Box 2. Cell. Physiol. Biochem. 2017, 43, 717–732. [Google Scholar] [CrossRef]
- Xu, C.; Li, B.; Zhao, S.; Jin, B.; Jia, R.; Ge, J.; Xu, H. MicroRNA-186-5p Inhibits Proliferation And Metastasis Of Esophageal Cancer By Mediating HOXA9. OncoTargets Ther. 2019, ume 12, 8905–8914. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.; Zhu, J.-H.; Yao, X.-Q. Long non-coding RNA PVT1 as a novel potential biomarker for predicting the prognosis of colorectal cancer. Int. J. Biol. Markers 2018, 33, 415–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ping, G.; Xiong, W.; Zhang, L.; Li, Y.; Zhang, Y.; Zhao, Y. Silencing long noncoding RNA PVT1 inhibits tumorigenesis and cisplatin resistance of colorectal cancer. Am. J. Transl. Res. 2018, 10, 138–149. [Google Scholar] [PubMed]
- Martínez-Barriocanal, Á.; Arango, D.; Dopeso, H. PVT1 Long Non-coding RNA in Gastrointestinal Cancer. Front. Oncol. 2020, 10, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Wang, P.; Xue, Y.; Qu, C.; Zheng, J.; Liu, X.; Ma, J.; Liu, Y. PVT1 affects growth of glioma microvascular endothelial cells by negatively regulating miR-186. Tumor Biol. 2017, 39, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Z.; Cui, J.; Song, Y. Long noncoding RNA PVT1 promotes EMT via mediating microRNA-186 targeting of Twist1 in prostate cancer. Gene 2018, 654, 36–42. [Google Scholar] [CrossRef]
- Zhu, X.; Shen, H.; Yin, X.; Long, L.; Xie, C.; Liu, Y.; Hui, L.; Lin, X.; Fang, Y.; Cao, Y.; et al. miR-186 regulation of Twist1 and ovarian cancer sensitivity to cisplatin. Oncogene 2016, 35, 323–332. [Google Scholar] [CrossRef]
- Cepowicz, D.; Zaręba, K.; Pryczynicz, A.; Dawidziuk, T.; Żurawska, J.; Hołody-Zaręba, J.; Gryko, M.; Kędra, B. Blood serum levels of E-cadherin in patients with colorectal cancer. Gastroenterol. Rev. 2017, 12, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Yin, Q.; Feng, W.; Shen, X.; Ju, S. Regulatory effects of lncRNAs and miRNAs on autophagy in malignant tumorigenesis. Biosci. Rep. 2018, 38, 1–12. [Google Scholar] [CrossRef]
- Senousy, M.A.; Helmy, H.S.; Fathy, N.; Shaker, O.G.; Ayeldeen, G.M. Association of MTMR3 rs12537 at miR-181a binding site with rheumatoid arthritis and systemic lupus erythematosus risk in Egyptian patients. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Orlandi, E.; Zanot, C.; Poli, A.; Nicolis, M.; Rodolfo, M.; Turco, A.; Sangalli, A.; Lira, M.G. Lack of association of metastasis-associated lung adenocarcinoma transcript 1 variants with melanoma skin cancer risk. Melanoma Res. 2019, 29, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Wang, M.; Tyrer, J.P.; Jensen, A.; Wiensch, A.; Liu, G.; Lee, A.; Ness, R.B.; Salvatore, M.; Tworoger, S.S.; et al. A comprehensive gene-environment interaction analysis in Ovarian Cancer using genome-wide significant common variants. Int. J. Cancer 2019, 144, 2192–2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinkins, B.; Nicholson, B.D.; Primrose, J.; Perera, R.; James, T.; Pugh, S.; Mant, D. The diagnostic accuracy of a single CEA blood test in detecting colorectal cancer recurrence: Results from the FACS trial. PLoS ONE 2017, 12, e0171810. [Google Scholar] [CrossRef]
- Radwan, A.F.; Ismael, O.E.; Fawzy, A.; El-Mesallamy, H.O. Evaluation of Serum Integrin αvβ3 & Vitronectin in the Early Diagnosis of Breast Cancer. Clin. Lab. 2019, 65, 1185–1193. [Google Scholar] [CrossRef]
- Goldstein, M.J.; Mitchell, E.P. Carcinoembryonic Antigen in the Staging and Follow-up of Patients with Colorectal Cancer. Cancer Investig. 2005, 23, 338–351. [Google Scholar] [CrossRef] [PubMed]
- Tong, G.; Xu, W.; Zhang, G.; Liu, J.; Zheng, Z.; Chen, Y.; Niu, P.; Xu, X. The role of tissue and serum carcinoembryonic antigen in stages I to III of colorectal cancer-A retrospective cohort study. Cancer Med. 2018, 7, 5327–5338. [Google Scholar] [CrossRef]
- Beck, D.E.; Roberts, P.L.; Rombeau, J.L.; Stamos, M.J.; Wexner, S.D. Anatomy and Embryology of the Colon, Rectum, and Anus; Springer Science and Business Media: New York, NY, USA, 2009; pp. 1–29. [Google Scholar]
- Bassig, B.A.; Cerhan, J.R.; Au, W.-Y.; Kim, H.N.; Sangrajrang, S.; Hu, W.; Tse, J.; Berndt, S.; Zheng, T.; Zhang, H.; et al. Genetic susceptibility to diffuse large B-cell lymphoma in a pooled study of three Eastern Asian populations. Eur. J. Haematol. 2015, 95, 442–448. [Google Scholar] [CrossRef]
- Andrii, V.; Yaroslav, C.; Viktoriia, H.; Alexander, A. Analysis of Association Between Rs3200401 Long Non- Coding RNA MALAT1 Gene Polymorphism and Prostate Adenocarcinoma Development in Ukrainian Population. J. Urol. Nephrol. Stud. 2019, 1, 99–102. [Google Scholar] [CrossRef]
- Qu, Y.; Shao, N.; Yang, W.; Wang, J.; Cheng, Y. Association of polymorphisms in MALAT1 with the risk of esophageal squamous cell carcinoma in a Chinese population. OncoTargets Ther. 2019, 12, 2495–2503. [Google Scholar] [CrossRef] [Green Version]
- Petkevicius, V.; Streleckiene, G.; Balciute, K.; Link, A.; Leja, M.; Malfertheiner, P.; Skieceviciene, J.; Kupcinskas, J. Association of Long Non-Coding RNA Polymorphisms with Gastric Cancer and Atrophic Gastritis. Genes 2020, 11, 1505. [Google Scholar] [CrossRef]
- Peng, R.; Luo, C.; Guo, Q.; Cao, J.; Yang, Q.; Dong, K.; Wang, S.; Wang, K.; Song, C. Association analyses of genetic variants in long non-coding RNA MALAT1 with breast cancer susceptibility and mRNA expression of MALAT1 in Chinese Han population. Gene 2018, 642, 241–248. [Google Scholar] [CrossRef]
- Wang, J.-Z.; Xiang, J.-J.; Wu, L.-G.; Bai, Y.-S.; Chen, Z.-W.; Yin, X.-Q.; Wang, Q.; Guo, W.-H.; Peng, Y.; Guo, H.; et al. A genetic variant in long non-coding RNA MALAT1 associated with survival outcome among patients with advanced lung adenocarcinoma: A survival cohort analysis. BMC Cancer 2017, 17, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.-C.; Dang, Y.-W.; Wei, D.-M.; Chen, P.; Tang, R.-X.; Huang, Q.; Liu, J.-H.; Luo, D.-Z. Clinical significance and prospective molecular mechanism of MALAT1 in pancreatic cancer exploration: A comprehensive study based on the GeneChip, GEO, Oncomine, and TCGA databases. OncoTargets Ther. 2017, 10, 3991–4005. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Liang, L.; Ouyang, K.; Li, Z.; Yi, X. MALAT1 induces tongue cancer cells’ EMT and inhibits apoptosis through Wnt/β-catenin signaling pathway. J. Oral Pathol. Med. 2017, 46, 98–105. [Google Scholar] [CrossRef]
- Niu, J.; Song, X.; Zhang, X. Regulation of lncRNA PVT1 on miR-125 in metastasis of gastric cancer cells. Oncol. Lett. 2019, 19, 1261–1266. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, R.; Wu, Y.; Liu, Y.; Su, W.; Xiong, W.; Zeng, Z. PVT1 Promotes Cancer Progression via MicroRNAs. Front. Oncol. 2019, 9, 609. [Google Scholar] [CrossRef] [Green Version]
- Roelands, J.; Kuppen, P.J.K.; Vermeulen, L.; Maccalli, C.; Decock, J.; Wang, E.; Marincola, F.M.; Bedognetti, D.; Hendrickx, W. Immunogenomic Classification of Colorectal Cancer and Therapeutic Implications. Int. J. Mol. Sci. 2017, 18, 2229. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Xia, Z.; Chen, C.; Hu, W.; Yuan, Y. LncRNA MALAT1 promotes epithelial-to-mesenchymal transition of esophageal cancer through Ezh2-Notch1 signaling pathway. Anti-Cancer Drugs 2018, 29, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Xiao, X.; Yang, Y.; Mishra, A.; Liang, Y.; Zeng, X.; Yang, X.; Xu, D.; Blackburn, M.R.; Henke, C.A.; et al. MicroRNA-101 attenuates pulmonary fibrosis by inhibiting fibroblast proliferation and activation. J. Biol. Chem. 2017, 292, 16420–16439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.; Luo, W.; Wang, J.; Peng, T.; Sun, G.; Shi, J.; Li, Z.; Zhang, B. Malat1 activates autophagy and promotes cell proliferation by sponging miR-101 and upregulating STMN1, RAB5A and ATG4D expression in glioma. Biochem. Biophys. Res. Commun. 2017, 492, 480–486. [Google Scholar] [CrossRef]
- Yu, F.; Lu, Z.; Cai, J.; Huang, K.; Chen, B.; Li, G.; Dong, P.; Zheng, J. MALAT1 functions as a competing endogenous RNA to mediate Rac1 expression by sequestering miR-101b in liver fibrosis. Cell Cycle 2015, 14, 3885–3896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Zhao, G.; Zhao, X.; Zhan, X.; Cai, M.; Geng, C.; Pu, Q.; Zhao, Q.; Fu, Q.; Huang, C.; et al. Elevated soluble E-cadherin during the epithelial-mesenchymal transition process and as a diagnostic marker in colorectal cancer. Gene 2020, 754, 144899. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, K.; Liao, Y.; Zeng, Q.; Li, Y.; Yuhua, L.; Liu, Y.; Meng, K.; Qian, C.; Zhang, Q.; et al. MicroRNA-101a Inhibits Cardiac Fibrosis Induced by Hypoxia via Targeting TGFβRI on Cardiac Fibroblasts. Cell. Physiol. Biochem. 2015, 35, 213–226. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Song, Z.; Chen, H.; Chen, Z.; Yang, P.; Li, W.; Yang, Z.; Zhang, T.; Wang, F.; Wei, J.; et al. Long noncoding RNA PVT1-214 promotes proliferation and invasion of colorectal cancer by stabilizing Lin28 and interacting with miR-128. Oncogene 2019, 38, 164–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waller, P.; Blann, A. Non-coding RNAs–A primer for the laboratory scientist. Br. J. Biomed. Sci. 2019, 76, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Wang, T.; Liu, Y.; Su, Z.; Lu, P.; Chen, X.; Hu, D. LncRNA PVT1 as an effective biomarker for cancer diagnosis and detection based on transcriptome data and meta-analysis. Oncotarget 2017, 8, 75455–75466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, L.D.; Pedersen, S.K.; Brown, G.S.; Ho, T.; Kassir, Z.; Moynihan, A.T.; Vizgoft, E.K.; Dunne, R.; Pimlott, L.; Young, G.; et al. Colorectal Neoplasia Differentially Expressed (CRNDE), a Novel Gene with Elevated Expression in Colorectal Adenomas and Adenocarcinomas. Genes Cancer 2011, 2, 829–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, X.; Cao, D.; Yang, L.; Li, X.; Zhang, W.; Xiao, Y.; Xi, Y.; Li, F.; Li, D.; Pan, Z. High Expression of long non-coding RNA PVT1 predicts metastasis in Han and Uygur Patients with Gastric Cancer in Xinjiang, China. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, T.; Huang, H.-Q.; Li, W.; Cheng, X.-L.; Yang, J. Human MALAT-1 long non-coding RNA is overexpressed in cervical cancer metastasis and promotes cell proliferation, invasion and migration. J. Balk. Union Oncol. 2016, 20, 1497–1503. [Google Scholar]
- Ji, Q.; Cai, G.; Liu, X.; Zhang, Y.; Wang, Y.; Zhou, L.; Sui, H.; Li, Q. MALAT1 regulates the transcriptional and translational levels of proto-oncogene RUNX2 in colorectal cancer metastasis. Cell Death Dis. 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Wang, S.-H.; Zhang, W.-J.; Wu, X.-C.; Zhang, M.-D.; Weng, M.-Z.; Zhou, D.; Wang, J.-D.; Quan, Z.-W. Long non-coding RNA Malat1 promotes gallbladder cancer development by acting as a molecular sponge to regulate miR-206. Oncotarget 2016, 7, 37857–37867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.-H.; Min-Hui, Y.; Xu, C.; Xie, L.-Y.; Wang, X.-Y.; Chen, S.-Y.; Lin-Ying, X. MALAT1 promotes colorectal cancer cell proliferation/migration/invasion via PRKA kinase anchor protein 9. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2015, 1852, 166–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onagoruwa, O.T.; Pal, G.; Ochu, C.; Ogunwobi, O.O. Oncogenic Role of PVT1 and Therapeutic Implications. Front. Oncol. 2020, 10, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, L.; Cai, Q.; Wang, S.; Wang, S.; Wang, J.; Quan, Z. Long noncoding RNA PVT1 promoted gallbladder cancer proliferation by epigenetically suppressing miR-18b-5p via DNA methylation. Cell Death Dis. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zu, X.; Zhang, H.; Qi, F.; Cao, Y.; Chen, M. Down-regulated microRNA-101 in bladder transitional cell carcinoma is associated with poor prognosis. Med. Sci. Monit. 2014, 20, 812–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Sha, H.-H.; Li, H.-J. Functions and mechanisms of miR-186 in human cancer. Biomed. Pharmacother. 2019, 119, 109428. [Google Scholar] [CrossRef]
- Roshchupkin, A.; Chumachenko, Y.; Ataman, A. Association of MALAT1 rs3200401 gene polymorphism with kidney cancer in Ukrainian population. East. Ukr. Med. J. 2019, 7, 121–125. [Google Scholar]
- Senousy, M.A.; El-Abd, A.M.; Abdel-Malek, R.R.; Rizk, S.M. Circulating long non-coding RNAs HOTAIR, Linc-p21, GAS5 and XIST expression profiles in diffuse large B-cell lymphoma: Association with R-CHOP responsiveness. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Shaker, O.G.; Khalefa, A.A.; Abdelwahed, M.Y.; Ali, E.; Ezzat, E.M.; Elghobary, H.A.; Awaji, A.A.; Fouad, N.A.; Ayoub, S.E. Serum long noncoding RNAs FAS-AS1 & PVT1 are novel biomarkers for systemic lupus erythematous. Br. J. Biomed. Sci. 2020, 77, 208–212. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CRC (n = 140) | AP (n = 40) | Healthy Controls (n = 100) | p Value | ||
|---|---|---|---|---|---|
| Age (years) Age range | 50 ± 12 (25–73) | 37 ± 16 (19–75) | 49 ± 8.2 (30–65) | <0.0001 a 0.888 b 0.006 c | |
| Sex, n (%) | Male | 90 (64.3%) | 24 (60%) | 62 (62%) | 0.861 |
| Female | 50 (35.7%) | 16 (40%) | 38 (38%) | ||
| Smokers | 42 (30%) | 8 (20%) | 19 (19%) | 0.084 | |
| Hemoglobin (g/dL) | 10.94 ± 2.75 | 11.98 ± 1.37 | 12.22 ± 1.45 | 0.04 a 0.009 b 0.88 c | |
| Platelet count × 103/mm3 | 276.6 ± 94.48 | 261.1 ± 40 | 248.6 ± 39.98 | 0.156 | |
| TLC × 103/mm3 | 6.98 ± 2.63 | 5.66 ± 1.05 | 6.44 ± 1.33 | 0.071 | |
| ESR (mm/h) | 46 ± 31.38 | 22 ± 15 | 20.1 ± 11 | <0.001 a,b >0.99 | |
| Anatomical Site, n (%) | |||||
| Proximal colon | 58 (41.4%) | 8 (20%) | - | - | |
| Distal colon | 45 (32.1%) | 17 (42.5%) | - | - | |
| Rectum | 37 (26.5%) | 15 (37.5%) | - | - | |
| Stage, n (%) | |||||
| Stage I, II (Early) | 88 (62.8%) | - | - | - | |
| Stage III, IV (Late) | 52 (37.2%) | - | - | - | |
| Tumor Grade, n (%) | |||||
| Well-differentiated | 26 (18.6%) | - | - | - | |
| Moderately differentiated | 98 (70%) | - | - | - | |
| Poorly-differentiated | 16 (11.4%) | - | - | - | |
| Lymphatic Metastasis, n (%) | |||||
| Present | 52 (37.1%) | - | - | - | |
| Absent | 85 (57.8%) | - | - | - | |
| Cannot be Assessed | 3 (2.1%) | - | - | - | |
| Distant Metastasis, n (%) | |||||
| Present | 24 (17.1%) | - | - | - | |
| Absent | 116 (82.9%) | - | - | - | |
| Tissue Type | |||||
| Adenocarcinoma | 131 (93.5%) | - | - | - | |
| Non-adenocarcinoma | 9 (6.5%) | - | - | - | |
| MALAT1 rs3200401 (C/T) (n = 280, Adjusted Analysis) | |||||||
|---|---|---|---|---|---|---|---|
| Model | Genotype | Control (n = 100) | CRC (n = 140) | Adjusted OR (95% CI) | p Value a | AIC a | BIC a |
| Codominant | CC | 42 (42%) | 31 (22.1%) | 1 | <0.0001 | 306 | 323.4 |
| CT | 48 (48%) | 62 (44.3%) | 1.78 (0.97–3.27) | ||||
| TT | 10 (10%) | 47 (33.6%) | 6.79 (2.92–15.80) | ||||
| Dominant | CC | 42 (42%) | 31 (22.1%) | 1 | <0.0001 | 316.3 | 330.2 |
| CT-TT | 58 (58%) | 109 (77.9%) | 2.62 (1.48–4.64) | ||||
| Recessive | CC-CT | 90 (90%) | 93 (66.4%) | 1 | <0.0001 | 307.5 | 321.4 |
| TT | 10 (10%) | 47 (33.6%) | 4.82 (2.25–10.31) | ||||
| Overdominant | CC-TT | 52 (52%) | 78 (55.7%) | 1 | 0.6 | 327 | 341 |
| CT | 48 (58%) | 62 (44.3%) | 0.87 (0.51–1.47) | ||||
| Allelic | C | 132 (53%) | 124 (44%) | 2.43 (1.64–3.61) | <0.0001 | 305.8 | 319.7 |
| T | 68 (34%) | 156 (56%) | |||||
| PVT1 rs13255292 (C/T) (n = 280, Adjusted Analysis) | |||||||
| Model | Genotype | Control (n = 100) | CRC (n = 140) | Adjusted OR (95% CI) | p Value a | AIC a | BIC a |
| Codominant | CC | 31 (31%) | 64 (45.7%) | 1 | 0.086 | 324.4 | 341.8 |
| CT | 55 (55%) | 62 (44.3%) | 0.56 (0.31–0.98) | ||||
| TT | 14 (14%) | 14 (10%) | 0.50 (0.21–1.19) | ||||
| Dominant | CC | 31 (31%) | 64 (45.7%) | 1 | 0.028 | 322.5 | 336.4 |
| CT-TT | 69 (69%) | 76 (54.3%) | 0.54 (0.32–0.94) | ||||
| Recessive | CC-CT | 86 (86%) | 126 (90%) | 1 | 0.38 | 326.6 | 340.5 |
| TT | 14 (14%) | 14 (10%) | 0.70 (0.31–1.56) | ||||
| Overdominant | CC-TT | 45 (45%) | 78 (55.7%) | 1 | 0.12 | 324.8 | 338.8 |
| CT | 55 (55%) | 62 (44.3%) | 0.66 (0.39–1.11) | ||||
| Allelic | C | 117 (58%) | 190 (68%) | 0.66 (0.44–0.98) | 0.04 | 323.1 | 337 |
| T | 83 (42%) | 90 (32%) | |||||
| MALAT1 rs3200401 (C/T) (n = 280, Adjusted Analysis) | |||||||
|---|---|---|---|---|---|---|---|
| Model | Genotype | Non-CRC (n = 140) | CRC (n = 140) | Adjusted OR (95% CI) | p Value a | AIC a | BIC a |
| Codominant | CC | 59 (42.1%) | 31 (22.1%) | 1 | <0.0001 | 362.2 | 380.4 |
| CT | 63 (45%) | 62 (44.3%) | 1.82 (1.03–3.23) | ||||
| TT | 18 (12.9%) | 47 (33.6%) | 5.12 (2.49–10.51) | ||||
| Dominant | CC | 59 (42.1%) | 31 (22.1%) | 1 | <0.0001 | 369.8 | 384.4 |
| CT-TT | 81 (57.9%) | 109 (77.9%) | 2.53 (1.48–4.32) | ||||
| Recessive | CC-CT | 122 (87.1%) | 93 (66.4%) | 1 | <0.0001 | 364.5 | 379 |
| TT | 18 (12.9%) | 47 (33.6%) | 3.60 (1.92–6.77) | ||||
| Overdominant | CC-TT | 77 (55%) | 78 (55.7%) | 1 | 0.83 | 381.8 | 396.3 |
| CT | 63 (45%) | 62 (44.3%) | 0.95 (0.58–1.54) | ||||
| Allelic | C | 181 (65%) | 124 (44%) | 2.21 (1.55–3.14) | <0.0001 | 360.9 | 375.4 |
| T | 99 (35%) | 156 (56%) | |||||
| PVT1 rs13255292 (C/T) (n = 280, Adjusted Analysis) | |||||||
| Model | Genotype | Non-CRC (n = 140) | CRC (n = 140) | OR (95% CI) | p Value | AIC | BIC |
| Codominant | CC | 47 (33.6%) | 64 (45.7%) | 1 | 0.1 | 379.3 | 397.4 |
| CT | 75 (53.6%) | 62 (44.3%) | 0.59 (0.35–1.00) | ||||
| TT | 18 (12.9%) | 14 (10%) | 0.55 (0.24–1.24) | ||||
| Dominant | CC | 47 (33.6%) | 64 (45.7%) | 1 | 0.034 | 377.3 | 391.9 |
| CT-TT | 93 (66.4%) | 76 (54.3%) | 0.59 (0.36–0.96) | ||||
| Recessive | CC-CT | 122 (87.1%) | 126 (90%) | 1 | 0.42 | 381.2 | 395.7 |
| TT | 18 (12.9%) | 14 (10%) | 0.73 (0.34–1.57) | ||||
| Overdominant | CC-TT | 65 (46.4%) | 78 (55.7%) | 1 | 0.12 | 379.4 | 393.9 |
| CT | 75 (53.6%) | 62 (44.3%) | 0.68 (0.42–1.10) | ||||
| Allelic | C | 169 (60%) | 190 (68%) | 0.70 (0.49–1.01) | 0.048 | 377.9 | 392.5 |
| T | 111 (40%) | 90 (32%) | |||||
| Haplotype | Total Frequency | Frequency in Non-CRC Group (n = 140) | Frequency in CRC Patients (n = 140) | Adjusted OR (95% CI) | p Value | |
|---|---|---|---|---|---|---|
| PVT1 rs13255292 | MALAT1 rs3200401 | |||||
| C a | C | 0.3574 | 0.404 | 0.3185 | 1.00 | - |
| C a | T a | 0.2836 | 0.1995 | 0.3601 | 2.21 (1.31–3.72) | 0.0032 * |
| T | C | 0.1872 | 0.2424 | 0.1244 | 0.64 (0.34–1.20) | 0.17 |
| T | T a | 0.1717 | 0.154 | 0.197 | 1.50 (0.89–2.51) | 0.13 |
| Marker | Cutoff | AUC | p Value | Sensitivity | Specificity | PPV | NPV | 95% CI |
|---|---|---|---|---|---|---|---|---|
| MALAT1 | >23.71-fold | 0.907 | <0.0001 | 82% | 88% | 90% | 78% | 0.8435 to 0.9707 |
| PVT1 | >13.96-fold | 0.848 | <0.0001 | 90% | 70% | 80% | 84% | 0.7641 to 0.9316 |
| miRNA-101 | <0.28-fold | 0.686 | 0.0049 | 62% | 67% | 72% | 56% | 0.5697 to 0.8015 |
| miRNA-186 | <0.20-fold | 0.702 | 0.0028 | 54% | 80% | 78% | 55% | 0.5861 to 0.8172 |
| E-Cadherin | <3.81 ng/mL | 0.864 | <0.0001 | 76% | 90% | 91% | 73% | 0.7837 to 0.9442 |
| Parameter | Beta Coefficient | SE | p Value | OR | OR (95% CI) |
|---|---|---|---|---|---|
| Univariate | |||||
| MALAT1 | 0.05 | 0.013 | <0.0001 | 1.052 | 1.052–1.078 |
| PVT1 | 0.0316 | 0.376 | 0.0027 | 1.032 | 1.011–1.053 |
| miRNA-101 | −0.54 | 0.254 | 0.0325 | 0.58 | 0.3529–0.955 |
| miRNA-186 | −1.12 | 0.39 | 0.0039 | 0.324 | 0.1512–0.697 |
| E-Cadherin | −1.26 | 0.313 | <0.0001 | 0.2811 | 0.1523–0.518 |
| Multivariate | |||||
| MALAT1 | 0.0398 | 0.015 | 0.0064 | 1.0403 | 1.0112–1.070 |
| PVT1 | 0.0187 | 0.011 | 0.0877 | 1.0188 | 0.9973–1.040 |
| miRNA-101 | −1.3931 | 0.759 | 0.0664 | 0.2483 | 0.0561–1.386 |
| miRNA-186 | −1.2410 | 0.750 | 0.0980 | 0.2891 | 0.0665–1.208 |
| E-Cadherin | −0.7858 | 0.450 | 0.0805 | 0.4532 | 0.1856–1.106 |
| Constant | 2.62 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radwan, A.F.; Shaker, O.G.; El-Boghdady, N.A.; Senousy, M.A. Association of MALAT1 and PVT1 Variants, Expression Profiles and Target miRNA-101 and miRNA-186 with Colorectal Cancer: Correlation with Epithelial-Mesenchymal Transition. Int. J. Mol. Sci. 2021, 22, 6147. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116147
Radwan AF, Shaker OG, El-Boghdady NA, Senousy MA. Association of MALAT1 and PVT1 Variants, Expression Profiles and Target miRNA-101 and miRNA-186 with Colorectal Cancer: Correlation with Epithelial-Mesenchymal Transition. International Journal of Molecular Sciences. 2021; 22(11):6147. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116147
Chicago/Turabian StyleRadwan, Abdullah F., Olfat G. Shaker, Noha A. El-Boghdady, and Mahmoud A. Senousy. 2021. "Association of MALAT1 and PVT1 Variants, Expression Profiles and Target miRNA-101 and miRNA-186 with Colorectal Cancer: Correlation with Epithelial-Mesenchymal Transition" International Journal of Molecular Sciences 22, no. 11: 6147. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116147