
Cryopreservation of Agronomic Plant Germplasm Using Vitrification-Based Methods: An Overview of Selected Case Studies

 ,
,  ,
, 
Abstract
:1. Introduction
2. Preservation of Plant Genetic Resources
3. Cryopreservation
4. Plant Material Used for Cryostorage
5. Cryoprotective Agents (CPAs)
5.1. Penetrating CPAs
5.2. Non-Penetrating CPAs
6. Osmoprotective Solutions
7. Plant Vitrification Solution
8. Modern CPAs
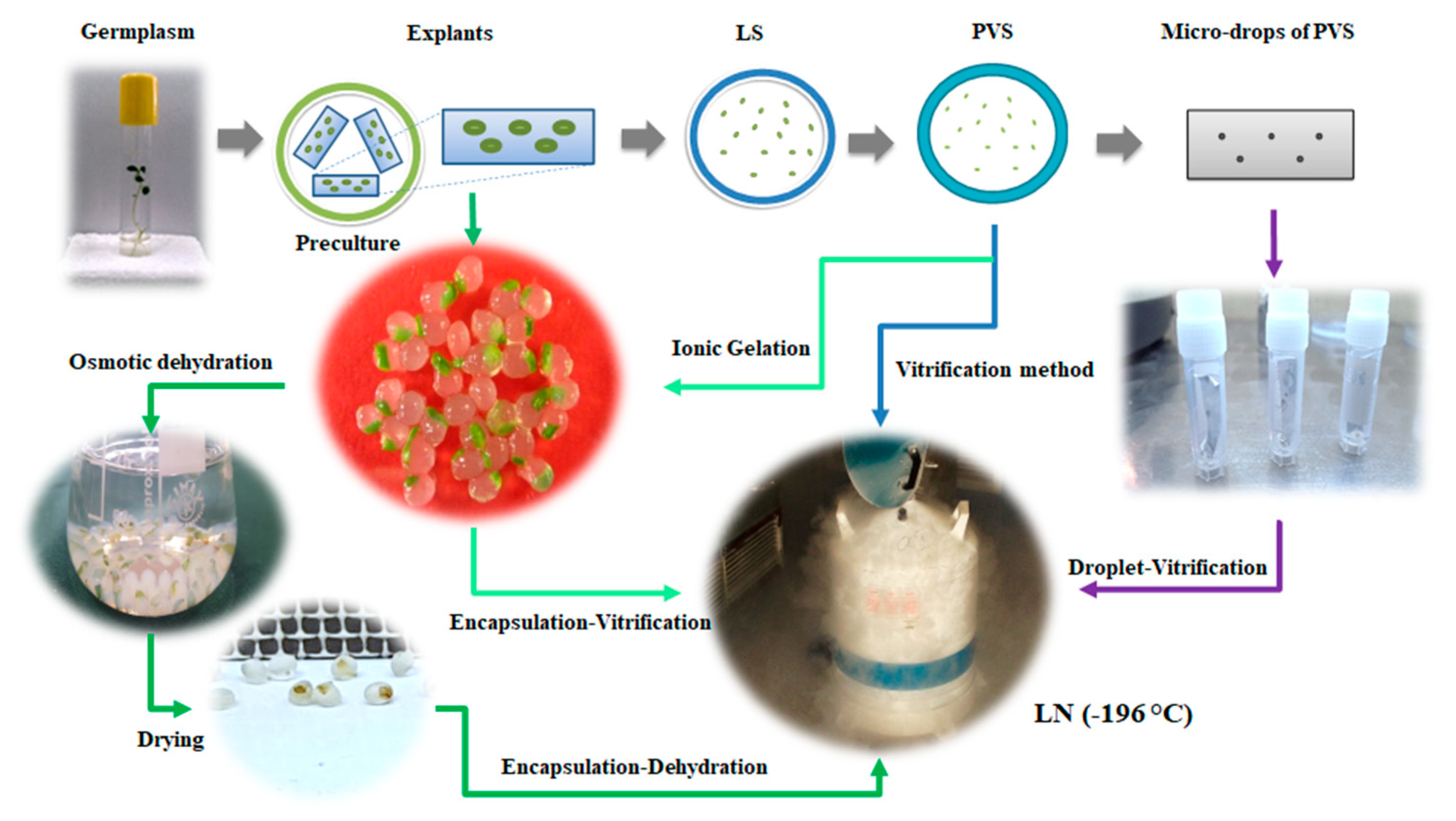
9. Cryopreservation Methods
9.1. Vitrification
9.2. Encapsulation-Dehydration
9.3. Encapsulation-Vitrification
9.4. Droplet-Vitrification
9.5. Cryo-Plate Techniques
9.6. Vacuum-Infiltration-Vitrification
9.7. Post-Cryopreservation
10. Cryopreservation Applications of Agronomic Interest

{kind=link}
{kind=link}
| Biological Material | Explant | Preculture | Pretreatment | Storage and Rewarming | Recovery | S(R) (%) | Remarks | Ref. |
|---|---|---|---|---|---|---|---|---|
| Vitrification | ||||||||
| Allium cepa var. aggregatum (Shallot ‘10603’) | Shoot tips (2–3 mm) with 4–5 leaf primordia | MS medium + 30 g/L sucrose, 0.5 mg/L BA, 0.1 mg/L NAA and 8 g/L agar (pH, 5.8) | PVS3 at 24 °C for 3 h | LN storage for 1 h | Preculture medium for 8 weeks | >95 | Rooting, vegetative growth, bulb production, genetic stability, and biochemical compounds were maintained after LN storage. | [151] |
| Allium sativum ‘Gailiangsuan’ cv. G064 | Shoot apices (2 mm) | MS medium with 6.5 g/L agar and 0.5 M sucrose for 4 d at 23 °C and 12 h photoperiod | LS: 18.4% glycerol + 20.5% sucrose in MS medium without agar for 20 min at 24 °C. PVS2 for 30 min at 0 °C | LN storage for 1 h. Rapid rewarming by directly plunging the samples into RS for 10 min at RT | B5 + 0.1 mg/L NAA + 2.0 mg/L 6-BA, with 30 g/L sucrose, 6.5 g/L agar in the dark for 4 d | 82.6 (75.9) | The LN-recovered plants were stable at the genetic and structural levels. | [152] |
| Solanum tuberosum ‘Superior’ | Shoot tips (2–3 mm) | Liquid MS medium with 0.3 M sucrose on a rotary shaker at 60 rpm for 24 h | LS: AFP III, (0–2000 ng/mL) + liquid MS medium, 0.6 M sucrose, 2.0 M glycerol at 25 °C for 1.5 h. Or PVS2 with AFP III for 30 min at 25 °C | LN storage for 30 min in and at −20 °C for 1 h. Rewarmed in a water bath (38 °C) for 2 min, and washed for 20 min in NH4+-free MS medium with 1.2 M sucrose at 25 °C | MS medium with 88 mM sucrose, agar 8 g/L | 26–39 (12–30) | This finding suggests that AFP increased cryopreservation efficiency by transcriptional regulation of these genes, which might protect plant cell membranes from cold stress during cryopreservation. | [153] |
| Solanum tuberosum (28 genotypes) | Apical shoots (2–3 mm) | 16–20 h in liquid MS medium with 0.09 M sucrose | liquid MS medium with 0.09 M sucrose and 10% DMSO for 2 h at RT | LN storage for 1 h. Rewarming in a water bath at 40 °C | MS medium with 0.09 M sucrose, 0.5 mg/L ZR, 0.2 mg/L GA, 0.5 mg/L IAA at 25/20 °C-d/n temperature 16 h photoperiod at low light intensity | (54) | All 28 genotypes had higher regrowth after cryopreservation using PVS3 instead of DMSO. | [154] |
| S. tuberosum ‘Zihuabai’ | Nodal segments (1 cm) | MS with 0.45 M sucrose in the dark at 5 °C for 1 d | 60–80% PVS2 for 30 min and 100% PVS2 for 40 min at 0 °C | LN storage for 1 h. Rewarming in a water bath at 38 °C for 2 min and then in RS at 25 °C for 20 min | MS with 0.5 mg/L IAA, 0.5 mg/L ZR and 0.2 mg/L GA and kept in the dark at 22 ± 2 °C for 3 d | ~80 (45.5) | No genetic alterations were detected in the recovered shoots by ISSR and AFLP. | [39] |
| Vitis vinifera L. ‘Flame Seedless’ | Axillary buds | MS medium | PVS2 at 25 °C with agitation for 3 h | LN storage for 1 h, 1 week, and 1 month. Rewarming in a water bath at 38 °C for 3 min | NR | NR | Cryopreservation affect genetic stability in grapevine, regardless of storage duration. | [155] |
| Droplet-Vitrification | ||||||||
| Allium cepa ‘Kverve’ ‘Lunteviga’ | Shoot tips (2–3 mm), 4 weeks old | MS medium with 0.3 M and 0.5 M sucrose, 1 d each | LS: 2.0 M glycerol + 0.6 M sucrose (20 min) and PVS3 at 24 °C (3 h) | LN storage for 1 h. Rewarming in RS at 25 °C for 20 min | MS medium with 0.3 M sucrose for 2 d in the dark and then transferred to light | (45–70) | PVS2 was more effective than PVS2 in securing the explants. Cryopreservation of shoot tips was more effective than of meristemoids. | [140] |
| Allium cepa var. aggregatum | Shoot tips (2–3 mm) | MS supplemented with 30 g/L sucrose, 0.5 mg/L BA, 0.1 mg/L NAA and 8 g/L agar | PVS3 at 24 °C for 3 h | LN storage for 1 h | Preculture medium for 8 weeks | NR | No differences in rooting, vegetative growth, bulb production, and contents of soluble sugars and flavonols between the cryo- and in vitro-derived plants. No polymorphisms found in the cryo-derived plants by ISSR and AFLP markers. | [151] |
| Helianthus tuberosus ‘M6′ ‘Relikt’ ‘Shudi’ ‘Stampede’ | Shoot tips (2–3 mm) | Liquid MS medium with 0.4 M sucrose (3 d) | LS: 2.0 M glycerol + 0.4 M sucrose (30 min) and PVS2 at 0 °C (15 min) | LN storage for 1 d. Rewarming in RS at 25 °C for 20 min | MS medium with 0.29 µM GA, 15% sucrose, and 8% agar cultured in the dark for 3–5 d, and then under 14 h photoperiod | 93 (83) | Minimal cellular damage observed within the meristem cells of the shoot tips. No polymorphism detected by SSRs. | [62] |
| Oxalis tuberosa and Ullucus tuberosus | Shoot tips (2 mm) | MS semisolid medium with 2% sucrose, 2 mg/L calcium pantothenate, 10 mg/L putrescine, 0.25 mg/L GA for oca; and with 2% sucrose and 2 mg/L calcium pantothenate for ulluco | LS: 2 M glycerol and 0.4 M sucrose in MS medium at RT for 20 min. PVS2 for 60 min, at 0 °C | LN storage for 30 min. Rewarming in RS for 20 min at RT | MS with, 0.3 M sucrose, 0.04 mg/L KIN, 0.1 mg/L GA, and 0.28% phytagel, incubated in the dark for 2 d. Then, 0.1 M sucrose/dark/2 d. Finally, MS medium with 0.07 M sucrose+ 2 m /L calcium pantothenate at 20 °C | 15 and 35, respectively | The protocols efficiency requires further improvement. | [131] |
| S. tuberosum ‘Agrie Dzeltenie’ ‘Anti’ ‘Bintje’ ‘Désirée‘ ‘Maret’ | Shoot tips (1–3 mm) | MS solution with 3% sucrose | 10% DMSO in liquid MS | LN storage for 1 h. Rewarming at RT in liquid MS | MS medium with 0.5 mg/L zeatin riboside, 0.2 mg/L GA, 0.5 mg/L IAA, 30 g/L sucrose | 17.1–52.6 | The optimization of light spectra during the recovery phase is a promising tool for increasing the recovery of potato shoot tips after cryopreservation. | [156] |
| S. tuberosum ‘Désirée’ S. commersonii Dun. | Shoot-tips (1 × 0.5 mm), 3 weeks old | MS medium with 0.21 M sucrose at 6 °C, 16/8-hour d/n and a light intensity of 50 μmol m−2 s−1 for 2 weeks | LS: 2 M glycerol and 0.4 M sucrose in MS medium (20 min). PVS2 for 50 min at 0 °C | LN storage for 30 min. Rewarming at RT in RS (20 min) | MS medium with 0.3 M sucrose (1 d). and then 0.09 M sucrose in the dark for the first 7 d | >80 | First study in which cryopreservation experiments are combined with the observation of the responses to abiotic stress exposure. | [157] |
| S. tuberosum ‘Zihuabai’ | Nodal segments (1 cm) | MS with 0.3 M sucrose in the dark at 5 °C for 3 d | LS: 2 M glycerol and 0.4 M sucrose in MS. PVS2 at 0 °C for 40 min | LN storage for 1 h. Rewarming in a water bath at 38 °C for 2 min and then in 1.2 M sucrose at 25 °C for 20 min | MS supplemented with 0.5 mg/L IAA, 0.5 mg/L ZR and 0.2 mg/L GA and kept in the dark at 22 ± 2 °C for 3 d | ~80 (72.5) | No genetic alterations were detected in the recovered shoots by ISSR and AFLP. | [39] |
| S. ajanhuiri ‘Wila Yari’ CIP702650 S. commersonii CGN18024, S. juzepcukii ‘Piñaza’ CIP702445 S. tuberosum ‘Désirée’ CIP800048 | Shoot tips (1 × 0.5 mm) | MS medium with 0.3 M/0.09 M sucrose at 6 °C for 14 d | LS: 2 M glycerol and 0.4 M sucrose in MS for 20 min in the dark, at RT. PVS2 for 50 min at 0 °C | LN storage for 30 min. Rewarming in RS at RT (20 min) | MS medium with 0.3 M sucrose solidified with 0.25% gerlite for 1 d and transfer to MS with 0.09 M sucrose | 90–100 | The increased accumulation of sucrose and raffinose family of oligosaccharides play a fundamental role in the response to stress in potato and may help to acquire tolerance to cryopreservation. | [158] |
| S. commersonii, S. tuberosum spp. andigena, S. tuberosum spp. tuberosum, S. × ajanhuiri, S. × juzepczukii | Shoot tips (1.8–2.5 mm), 3 weeks old | MS salts with 0.04 mg/L KIN, 0.1 mg/L GA, 0.07 M or 0.03 M sucrose and 2.8 g/L Phytagel at 6 °C and RT for 1 h | LS: 2 M glycerol and 0.4 M sucrose for 15 min at RT PVS2 for 50 min on ice | LN storage for 1 h. Rewarming in RS at RT (15–20 min). in the dark | MS salts with 0.04 mg/L KIN 0.1 mg/L GA, 2.8 g/L phytagel + 0.3 M sucrose. Daily transfers from 0.3, to 0.2, to 0.1 M and finally maintained on 0.07 M sucrose at 22 °C, 16 h photoperiod | 40–100 | This method is recommended for the long-term conservation of diverse accessions of potato germplasm. | [99] |
| Solanum tuberosum ‘Agrie’ ‘Anti’ ‘Bintje’ ‘Désirée’ ‘Dzeltenie’ ‘Maret’ | Shoot tips (1–3 mm) | MS medium with 2% sucrose and 6.4 g/L agar under various light spectra conditions at 22 °C | LS: MS-solution with 3% sucrose under the original light quality treatments overnight. Dehydration for 2 h in MS-solution with 10% DMSO | LN storage for 1 h. Rewarmed by dipping the foils with shoot tips into 30 mL of MS solution at RT | MS medium with 0.5 mg/L ZR, 0.2 mg/L GA, 0.5 mg/L IAA, 30 g/L sucrose and (1 mL /L) PPM | 31–66 using blue light | Light spectral quality before cryopreservation can significantly affect the cryopreservation success of potato shoot tips. | [119] |
| S. tuberosum ‘Blue Congo’ ‘E03-2677′ | Buds (1.0–1.4 mm) | MS with 0.3 M sucrose at 4 °C and 16 h photoperiod for 3 weeks | LS: MS with 0.1 M sucrose and 2 M glycerol for 30 min on ice. PVS2 for 30 to 40 min on ice | LN storage for 1 h. Rewarming in a water bath at 38 °C for 2 min | MS medium with 0.2 mg/L GA, 0.5 mg/L IAA, and ZR for 3 d in the dark. Then, MS with 0.05 mg/L GA in light | (70–80) | First report on cryopreservation of purple-fleshed potato by vitrification-based procedures. Larger explants (1.5–2.0 mm) were less effective. | [104] |
| S. tuberosum (28 genotypes) | Shoot apices (2–3 mm) | Liquid MS medium with 0.3 M sucrose for 16–20 h | LS: 0.4 M sucrose and 2 M glycerol in MS for 20 min and PVS3 for 2 h | LN storage for 1 h. Rewarming at 40 °C, unloading in RS (20 min) recovered 25/20 °C d/n or only 20 °C for 7 d | MS medium with 0.09 M sucrose, 0.5 mg/L ZR, 0.2 mg/L GA, 0.5 mg/L IAA at 25/20 °C d/n temperature in the dark (7 d) | (71) | All 28 genotypes had higher regrowth after cryopreservation using PVS3 instead of DMSO. | [154] |
| Potato varieties (Solanum spp.) | Shoot tips with 3–4 leaf primordia (length: 0.8–1.2 mm; width: 0.4–0.7 mm) | MS medium with 0.04 mg/L KIN, 0.1 mg/L GA, 25 g/L sucrose and 2.8 g/L Phytagel | LS: 2.0 M glycerol and 0.4 M sucrose in MS at RT for 20 min. PVS2 on ice for 50 min | LN storage for 24 h. Rewarming in liquid MS medium with 0.0–1.2 M sucrose for 20 min at RT | MS medium with 0.4 mg/1 KIN, 0.1 mg/L GA, 20 mL /L coconut water, 0.3 M sucrose, 2.8 g/L Phytagelfor 4 d under diffuse light at 18–22 °C with a 16 h photoperiod | 74.6–90.7 (66.5–86.8) | The specific response to low (0.0 M) and high (1.2 M) concentrations of sucrose in the unloading solution was highly variable within species/subspecies and appears to be genotype-specific. | [67] |
| Encapsulation-Vitrification | ||||||||
| Solanum tuberosum ‘Zihuabai’ | Nodal segments (1 cm) | MS with 0.3 M sucrose (1 d) and then suspended in MS with 2.5% (w/v) alginate and 0.4 M sucrose | LS: 2 M glycerol and 0.6 M sucrose in MS for 90 min and then dehydrated with PVS2 at 0 °C for 4 h | LN storage for 1 h. Rewarming in a water bath at 38 °C for 2 min and then in 1.2 M sucrose at 25 °C for 20 min | MS supplemented with 0.5 mg/L IAA, 0.5 mg/L ZR and 0.2 mg/L GA and kept in the dark at 22 ± 2 °C for 3 d | 70 | No genetic alterations were detected by ISSR and AFLP. | [39] |
| Solanum tuberosum ‘Blue Congo’ ‘E03-2677′ | Buds of 1.5–2.0 mm | MS with 0.3 M sucrose at 4 °C and 16 h photoperiod for 3 weeks, then suspended in 1/2 MS with 3% alginate, 2 M glycerol, and 0.4 M sucrose | MS containing 2 M glycerol and 0.6 M sucrose at RT for 90 min and PVS2 on ice for 6–7 h | LN storage for 1 h. Rewarming in a water bath at 38 °C for 2 min | MS medium with 0.2 mg/L GA, 0.5 mg/L IAA, and ZR for 3 d in the dark. Then, MS with 0.05 mg/L GA in light | (40–80) | First report on cryopreservation of purple-fleshed potato by vitrification-based procedures. Smaller explants (1.0–1.4 mm) were less effective. | [104] |
| S. tuberosum ‘Zihuabai’ | Nodal segments (1 cm) | MS with 0.45 M sucrose in the dark at 5 °C for 1 d | LS: 2 M glycerol and 0.6 M sucrose in MS for 90 min. PVS2 at 0 °C for 4 h | LN storage for 1 h. Rewarming in a water bath at 38 °C for 2 min and then in 1.2 M sucrose at 25 °C for 20 min | MS with 0.5 mg/L IAA, 0.5 mg/L ZR, 0.2 mg/L GA, kept in the dark at 22 °C for 3 d | ~80 (70) | No genetic alterations detected in the recovered shoots by ISSR and AFLP markers. | [39] |
| Encapsulation-Dehydration | ||||||||
| A. officinalis ‘Morado de Huétor’ | Rhizome buds | MS with 0.3 M sucrose at 25 °C for 24 or 48 h | Explants embedded in alginate (3%), sucrose (0.4 M) and glycerol (2 M). Desiccation over silica gel for 24 h | LN storage for 24 h. Rewarming in distilled water at RT | MS modified with Ferric Sodium EDDHA 85.7 mg/L, 0.5 mg/L NAA, 0.7 mg/L kinetin, 2 mg/L Ancymidol, and 6% sucrose | 34.5–84 | Confirmation of ploidy and molecular stability of LN-recovered plantlets. | [50] |
| Mentha × piperita ‘MEN 198′ | Axillary buds (1–2 mm) | MS medium with 0.3 M sucrose for 1 d at RT and 16 h photoperiod at low light intensity. Explants suspended in MS with 3% alginate and 0.35 M sucrose (30 min) | Osmotic dehydration in MS liquid + 0.75 M sucrose at 120 rpm for 18–20 h. Then, drying in a flow chamber for 5 h to a level of 22% initial water content | LN storage for 1 d. Rewarming in a water bath at 40 °C for 2 min | NR | NR | RAPD and AFLP showed an almost complete genetic stability of the recovered plants. | [103] |
| Solanum phureja | Single node stem cuttings (1 cm) | 0.1 M sucrose in TR medium, MS microelements, vitamins of MW medium for 7 d at RT, and 16 h photoperiod. Suspension in 3% alginate and 0.1 M sucrose | PC medium supplemented with (sugars and polyols) (0.65 M) for 2 d at RT. Dehydration for 4.5 h on silica gel to a level of 22% initial water content | LN storage for 1 h. Rewarming at RT | PC medium | 88 (maltose) and 91 (trehalose) | Maltose and trehalose were the most effective cryoprotectants. Shoot tips precultured in sucrose, trehalose or glucose, indicated an increase in total soluble sugars, especially when sucrose was applied as a cryoprotectant. | [146] |
| Cryo-plate | ||||||||
| Allium spp. A. sativum A. fistulosum var. caespitosum A. chinense | Shoot tips (2.5 mm), 2 days old | ½MS medium with 0.3 M sucrose for 2 d. Explants embedded on cryo-plates with 3% alginate and 0.4 M sucrose (15 min) | LS: 1.0 M sucrose and 2.0 M glycerol (30 min) at 25 °C. Air desiccation for 30 to 180 min | LN storage for 1 h. Rewarming in 1.0 M sucrose solution in 1/2 MS medium at 25 °C for 30 min | ½MS medium with 3.0% sucrose and 0.8% agar | 94 | The glass transition temperature of shoot tips after air desiccation was −39.4 °C. | [159] |
| Solanum tuberosum ‘Sayaka’ | Shoot tips (0.5–1.5 mm) | MS medium + 0.3 M sucrose and 0.3% gellan gum at 25 °C overnight. Explants embedded on cryo-plates with 2% alginate (15 min) | LS: 2.0 M glycerol and 0.8 M sucrose in MS at 25 °C for 30 min. V cryo-plate: PVS2 at 25 °C for 30 min D cryo-plate: air desiccation for 2 h | LN storage for 30 min. Rewarming in MS medium with 1 M sucrose for 15 min at RT | PC medium | D cryo-plate: 93.3; V cryo-plate: 96.7 | Both protocols will facilitate efficient strategies for the preservation, storage, and maintenance of genetic stability of potato germplasm. | [160] |
| Ullucus tuberosus 11 lines | Shoot tips (1.0–1.5 mm) | MS with 0.3 M sucrose for 16 h at 25 °C | D cryo-plate: Alginate with 0.4 M sucrose. Cryo-plates + 2 M glycerol + 1 M sucrose for 90 min at 25 °C. Dehydration at 25 °C for 45 min | LN storage | - | 73–97 | D cryo-plate is a practical and simple procedure for cryostorage of in vitro grown ulluco shoot tips in an ex situ genebank. | [132] |
| Desiccation | ||||||||
| Potato: ‘Avon’ ‘Ceza’ ‘Climax’ | Microtubers (~2 mm) | MS medium without vitamins with 100 g/L sucrose, and 10 g/L agar | Microtuber desiccation using sterile dry silica gel for 3–6 h (17–36% FW) | LN storage for 10 min. Rewarming in a water bath at 45 °C for 60 s and in a liquid MS medium at RT for 5–10 min | MS with vitamins, 30 g/L sucrose and 2 g/L activated charcoal at 25 °C with 16 h photoperiod | 100 | The desiccation technique is a simple approach for cryostorage of microtubers. | [147] |
| S(R) = Survival (recovery). NR = not reported. RT = room temperture. RS = rewarming solution. d/n = day/night. | ||||||||
11. Oxidative Stress Markers in LN-Derived Plant Material
12. Stability and Functional Genomics of LN-Derived Plant Material
13. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| AFLP | Amplified fragment length polymorphism |
| AFP | Antifreeze protein |
| APX | Ascorbic acid peroxidase |
| BA | N6-benzyladenine |
| CAT | Catalase |
| CBD | Convention on Biological Diversity |
| CIAT | International Center for Tropical Agriculture |
| CPAs | Cryoprotective agents |
| CRI | Crop Research Institute in the Czech Republic |
| DMSO | Dimethyl sulfoxide |
| DSC | Differential scanning calorimetry |
| EG | Ethylene glycol |
| EMBRAPA | Brazilian Agricultural Research Corporation |
| FCM | Flow cytometry |
| GA | Gibberellic acid |
| GABA | γ-aminobutyric acid |
| GR | Glutathione reductase |
| HEPES | 4-(2-hydroxyethyl)-1-piperazine ethanesulfonic acid |
| InHORT | Research Institute of Horticulture in Poland |
| IPC | International Potato Center |
| IAA | Indoleacetic acid |
| ISSR | Inter simple sequence repeat |
| KIN | Kinetin |
| LEA | Late embryogenesis abundant |
| LN | Liquid nitrogen |
| LS | Loading solution |
| MDA | Malondialdehyde |
| MS | Murashige and Skoog |
| MSAP | Methylation-sensitive amplified polymorphism |
| NAA | 1-naphthaleneacetic acid |
| NARO | National Agriculture and Food Research Organization in Japan |
| NCSS | National Center for Seeds and Seedlings in Japan |
| NIAS | National Institute of Agrobiological Sciences in Japan |
| NPs | Nanoparticles |
| PC | Preculture |
| PCO | Protein carbonyls |
| PEG | Polyethylene glycol |
| PG | Propylene glycol |
| PGRs | Plant growth regulators |
| POD | Peroxidase |
| PVS | Plant vitrification solution |
| RAPD | Randomly amplified polymorphic DNA |
| ROS | Reactive oxygen species |
| RS | Sakai’s unloading solution |
| SA | Salicylic acid |
| SOD | Superoxidase dismutase |
| SSR | Sequence simple repeat |
| SWCNTs | Single-wall carbon nanotubes |
| TF | Transcription factor |
| ZR | Zeatin riboside |
References
- Milla, R. Crop Origins and Phylo Food: A database and a phylogenetic tree to stimulate comparative analyses on the origins of food crops. Glob. Ecol. Biogeogr. 2020, 29, 606–614. [Google Scholar] [CrossRef]
- CBD, U. Convention on Biological Diversity (with Annexes); Concluded at Rio de Janeiro on 5 June 1992; United Nations: Rio de Janeiro, Brazil, 1992. [Google Scholar]
- Lambers, J.R.H. Extinction risks from climate change. Science (80-) 2015, 348, 501–502. [Google Scholar] [CrossRef]
- Prevéy, J.S.; Parker, L.E.; Harrington, C.A. Projected impacts of climate change on the range and phenology of three culturally-important shrub species. PLoS ONE 2020, 15, e0232537. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Sáenz, D.R.; Ayala-Hernández, D.D.; Niino, T.; Cruz-Gutiérrez, E.J.; Aquino-Martínez, J.G.; López-Delgado, H.A. Salicylic acid-cryotherapy treatment for elimination of potato virus S from Solanum tuberosum. Am. J. Potato Res. 2019, 96, 225–234. [Google Scholar] [CrossRef]
- Urban, M.C. Accelerating extinction risk from climate change. Science (80-) 2015, 348, 571–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, M.C.; Bocedi, G.; Hendry, A.P.; Mihoub, J.B.; Peer, G.; Singer, A.; Bridle, J.R.; Crozier, L.G.; De Meester, L.; Godsoe, W.; et al. Improving the forecast for biodiversity under climate change. Science (80-) 2016, 353, aad8466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pullman, G.S.; Bai, K.; Hane, M.; Ruland, D.; Cruse-Sanders, J.M.; Boyd, R.S.; Johnson, S. Seed cryopreservation and micropropagation of the federally threatened species, Price’s potato-bean (Apios priceana B.L. Robins.). Vitro Cell. Dev. Biol. Plant 2019, 55, 558–568. [Google Scholar] [CrossRef]
- Ruta, C.; Lambardi, M.; Ozudogru, E.A. Biobanking of vegetable genetic resources by in vitro conservation and cryopreservation. Biodivers. Conserv. 2020, 29, 3495–3532. [Google Scholar] [CrossRef]
- Fitzgerald, H.; Palmé, A.; Asdal, Å.; Endresen, D.; Kiviharju, E.; Lund, B.; Rasmussen, M.; Thorbjörnsson, H.; Weibull, J. A regional approach to Nordic crop wild relative in situ conservation planning. Plant Genet. Resour. Charact. Util. 2019, 17, 196–207. [Google Scholar] [CrossRef] [Green Version]
- Vincent, H.; Amri, A.; Castañeda-Álvarez, N.P.; Dempewolf, H.; Dulloo, E.; Guarino, L.; Hole, D.; Mba, C.; Toledo, A.; Maxted, N. Modeling of crop wild relative species identifies areas globally for in situ conservation. Commun. Biol. 2019, 2, 136. [Google Scholar] [CrossRef] [Green Version]
- Ayuso, M.; García-Pérez, P.; Ramil-Rego, P.; Gallego, P.P.; Barreal, M.E. In vitro culture of the endangered plant Eryngium viviparum as dual strategy for its ex situ conservation and source of bioactive compounds. Plant Cell. Tissue Organ Cult. 2019, 138, 427–435. [Google Scholar] [CrossRef]
- Amano, R.; Nakayama, H.; Momoi, R.; Omata, E.; Gunji, S.; Takebayashi, Y.; Kojima, M.; Ikematsu, S.; Ikeuchi, M.; Iwase, A.; et al. Molecular basis for natural vegetative propagation via regeneration in North American Lake Cress, Rorippa aquatica (Brassicaceae). Plant Cell Physiol. 2020, 61, 353–369. [Google Scholar] [CrossRef]
- Guimarães, R.N.; Souza, E.R.B.d.; Naves, R.V.; Melo, A.P.C.d.; Rubio Neto, A. Vegetative propagation of pequi (souari nut) by cutting. Ciência Rural 2019, 49. [Google Scholar] [CrossRef]
- Nagahama, N.; Manifesto, M.M.; Fortunato, R.H. Vegetative propagation and proposal for sustainable management of Valeriana carnosa Sm., a traditional medicinal plant from Patagonia. J. Appl. Res. Med. Aromat. Plants 2019, 14, 100218. [Google Scholar] [CrossRef]
- Onyebuchi Okonkwo, H. Amenability of Cola Lepidota to vegetative propagation by marcotting. Niger. J. Agric. Food Environ. 2020, 16, 7–13. [Google Scholar]
- Kulus, D. Genetic resources and selected conservation methods of tomato. J. Appl. Bot. Food Qual. 2018, 91, 135–144. [Google Scholar] [CrossRef]
- Gonzalez-Arnao, M.T.; Martinez-Montero, M.E.; Cruz-Cruz, C.A.; Engelmann, F. Advances in Cryogenic Techniques for the Long-Term Preservation of Plant Biodiversity; Springer: Berlin/Heidelberg, Germany, 2014; pp. 129–170. [Google Scholar]
- Fiqa, A.P.; Fauziah, F.; Lestari, D.A.; Budiharta, S. The importance of in-situ conservation area in mining concession in preserving diversity, threatened and potential floras in East Kalimantan, Indonesia. Biodiversitas J. Biol. Divers. 2018, 20, 198–210. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.; Xu, G.; Li, D.; Clements, D.R.; Zhang, F.; Jin, G.; Wu, J.; Wei, P.; Lin, S.; Xue, D. Agrobiodiversity and in situ conservation in ethnic minority communities of Xishuangbanna in Yunnan Province, Southwest China. J. Ethnobiol. Ethnomed. 2017, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bekheet, S.A.; Sota, V.; El-Shabrawi, H.M.; El-Minisy, A.M. Cryopreservation of shoot apices and callus cultures of globe artichoke using vitrification method. J. Genet. Eng. Biotechnol. 2020, 18, 2. [Google Scholar] [CrossRef]
- Ramírez-Mosqueda, M.A.; Cruz-Cruz, C.A.; Atlahua-Temoxtle, J.; Bello-Bello, J.J. In vitro conservation and regeneration of Laelia anceps Lindl. S. Afr. J. Bot. 2019, 121, 219–223. [Google Scholar] [CrossRef]
- Sánchez-Chiang, N.; Jiménez, V.M. Técnicas de conservación in vitro para el establecimiento de bancos de germoplasma en cultivos tropicales. Agron. Mesoam. 2010, 21, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Engelmann, F. Cryopreservation of embryos: An overview. Methods Mol. Biol. 2011, 710, 155–184. [Google Scholar] [PubMed]
- Acosta, Y.; Pérez, L.; Linares, C.; Hernández, L.; Escalante, D.; Pérez, A.; Zevallos, B.E.; Yabor, L.; Martínez-Montero, M.E.; Cejas, I.; et al. Effects of Teramnus labialis (L.f.) Spreng seed cryopreservation on subsequent seed and seedling growth and biochemistry. Acta Physiol. Plant. 2020, 42, 7. [Google Scholar] [CrossRef]
- Agrawal, A.; Singh, S.; Malhotra, E.V.; Meena, D.P.S.; Tyagi, R.K. In Vitro Conservation and Cryopreservation of Clonally Propagated Horticultural Species. In Conservation and Utilization of Horticultural Genetic Resources; Springer: Singapore, 2019; pp. 529–578. [Google Scholar]
- Quatrano, R.S. Freeze-preservation of cultured flax cells utilizing dimethyl sulfoxide. Plant Physiol. 1968, 43, 2057–2061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. var. brasiliensis Tanaka) by vitrification. Plant Cell Rep. 1990, 9, 30–33. [Google Scholar] [CrossRef]
- Sugawara, Y.; Sakai, A. Survival of Suspension-cultured sycamore cells cooled to the temperature of liquid nitrogen. Plant Physiol. 1974, 54, 722–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panis, B.; Thinh, N.T. Cryopreservation of Musa germplasm; INIBAP Technical Guideline 5; Bioversity International: Montpellier, France, 2001. [Google Scholar]
- Pence, V.C.; Clark, J.R. Desiccation, cryopreservation and germination of seeds of the rare wetland species, Plantago cordata Lam. Seed Sci. Technol. 2005, 33, 767–770. [Google Scholar] [CrossRef]
- Berjak, P.; Pammenter, N.W. Implications of the lack of desiccation tolerance in recalcitrant seeds. Front. Plant Sci. 2013, 4, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, C.; Pammenter, N.W.; Berjak, P.; Crane, J. Desiccation damage, accelerated ageing and respiration in desiccation tolerant and sensitive seeds. Seed Sci. Res. 2001, 11, 135–148. [Google Scholar] [CrossRef]
- Pence, V. Cryopreservation of immature embryos of Theobroma cacao. Plant Cell Rep. 1991, 10, 144–147. [Google Scholar] [CrossRef]
- Pence, V.C. Evaluating costs for the in vitro propagation and preservation of endangered plants. Vitro Cell. Dev. Biol. Plant 2011, 47, 176–187. [Google Scholar] [CrossRef]
- Agrawal, A.; Sanayaima, R.; Singh, R.; Tandon, R.; Verma, S.; Tyagi, R.K. Phenotypic and molecular studies for genetic stability assessment of cryopreserved banana meristems derived from field and in vitro explant sources. Vitro Cell. Dev. Biol. Plant 2014, 50, 345–356. [Google Scholar] [CrossRef]
- Sakai, A. Development of Cryopreservation Techniques. In Cryopreservation of Tropical Plant Germplasm—Current Research Progress and Applications; Engelmann, F., Takagi, H., Eds.; International Plant Genetic Resources Institute: Roma, Italy, 2000; pp. 1–7. ISBN 9290434287. [Google Scholar]
- Kim, H.H.; Cho, E.G.; Baek, H.J.; Kim, C.Y.; Keller, E.R.J.; Engelmann, F. Cryopreservation of garlic shoot tips by vitrification: Effects of dehydration, rewarming, unloading and regrowth conditions. Cryo Lett. 2004, 25, 59–70. [Google Scholar]
- Wang, B.; Li, J.W.; Zhang, Z.B.; Wang, R.R.; Ma, Y.L.; Blystad, D.R.; Keller, E.R.J.; Wang, Q.C. Three vitrification-based cryopreservation procedures cause different cryo-injuries to potato shoot tips while all maintain genetic integrity in regenerants. J. Biotechnol. 2014, 184, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Nadarajan, J.; van der Walt, K.; Lehnebach, C.A.; Saeiahagh, H.; Pathirana, R. Integrated ex situ conservation strategies for endangered New Zealand Myrtaceae species. N. Zeal. J. Bot. 2020, 1–18. [Google Scholar] [CrossRef]
- Volk, G.M.; Walters, C. Plant vitrification solution 2 lowers water content and alters freezing behavior in shoot tips during cryoprotection. Cryobiology 2006, 52, 48–61. [Google Scholar] [CrossRef]
- Tanaka, D.; Niino, T.; Uemura, M. Cryopreservation of Plant Genetic Resources. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2018; Volume 1081, pp. 355–369. [Google Scholar]
- Kulus, D. Application of Cryogenic Technologies and Somatic Embryogenesis in the Storage and Protection of valuable Genetic Resources of Ornamental Plants. In Somatic Embryogenesis in Ornamentals and Its Applications; Springer: New Delhi, India, 2015; pp. 1–25. ISBN 9788132226833. [Google Scholar]
- Panta, A.; Panis, B.; Ynouye, C.; Swennen, R.; Roca, W. Development of a PVS2 droplet vitrification method for potato cryopreservation. Cryo Lett. 2014, 35, 255–266. [Google Scholar]
- Yoon, J.W.; Kim, H.H.; Cho, E.G.; Ko, H.C.; Hwang, H.S.; Park, Y.E.; Engelmann, F. Cryopreservation of cultivated and wild potato varieties by droplet vitrification procedure. Acta Hortic. 2007, 760, 203–208. [Google Scholar]
- Engelmann, F. Plant cryopreservation: Progress and prospects. Vitro Cell. Dev. Biol. Plant 2004, 40, 427–433. [Google Scholar] [CrossRef]
- Wang, M.-R.; Zhang, Z.; Zámečník, J.; Bilavčík, A.; Blystad, D.-R.; Haugslien, S.; Wang, Q.-C. Droplet-vitrification for shoot tip cryopreservation of shallot (Allium cepa var. aggregatum): Effects of PVS3 and PVS2 on shoot regrowth. Plant Cell Tissue Organ Cult. 2020, 140, 185–195. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.-H. Cryopreservation of carrot (Daucus carota l.) cell suspensions and protoplasts by vitrification. Cryo Lett. 2003, 24, 57–64. [Google Scholar]
- Fábián, A.; Jäger, K.; Darkó, É.; Barnabás, B. Cryopreservation of wheat (Triticum aestivum L.) egg cells by vitrification. Acta Physiol. Plant. 2008, 30, 737–744. [Google Scholar] [CrossRef]
- Carmona-Martín, E.; Regalado, J.J.; Perán-Quesada, R.; Encina, C.L. Cryopreservation of rhizome buds of Asparagus officinalis L. (cv. Morado de Huétor) and evaluation of their genetic stability. Plant Cell. Tissue Organ Cult. 2018, 133, 395–403. [Google Scholar] [CrossRef]
- Lambert, E.; Goossens, A.; Panis, B.; Van Labeke, M.C.; Geelen, D. Cryopreservation of hairy root cultures of Maesa lanceolata and Medicago truncatula. Plant Cell. Tissue Organ Cult. 2009, 96, 289–296. [Google Scholar] [CrossRef]
- Dolce, N.R.; Hernández-Ramírez, F.; González-Arnao, M.T. Cryopreservation of vanilla (Vanilla planifolia) root-tips: A new alternative for in vitro long-term storage of its germplasm. Acta Hortic. 2019, 1234, 203–210. [Google Scholar] [CrossRef]
- Popova, E.; Shukla, M.; Kim, H.H.; Saxena, P.K. Root cryobanking: An important tool in plant cryopreservation. Plant Cell. Tissue Organ Cult. 2021, 144, 49–66. [Google Scholar] [CrossRef]
- Wowk, B. Thermodynamic aspects of vitrification. Cryobiology 2010, 60, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Elliott, G.D.; Wang, S.; Fuller, B.J. Cryoprotectants: A review of the actions and applications of cryoprotective solutes that modulate cell recovery from ultra-low temperatures. Cryobiology 2017, 76, 74–91. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Oldenhof, H.; Dai, X.; Haverich, A.; Hilfiker, A.; Harder, M.; Wolkers, W.F. Protein stability in stored decellularized heart valve scaffolds and diffusion kinetics of protective molecules. Acta Proteins Proteom. 2014, 1844, 430–438. [Google Scholar] [CrossRef]
- Teixeira, A.S.; Elena González-Benito, M.; Molina-García, A.D. Glassy state and cryopreservation of mint shoot tips. Biotechnol. Prog. 2013, 29, 707–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, A.S.; Faltus, M.; Zámečník, J.; González-Benito, M.E.; Molina-García, A.D. Glass transition and heat capacity behaviors of plant vitrification solutions. Thermochim. Acta 2014, 593, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Tanne, E.; Arav, A.; Gafny, R. Cryopreservation of in vitro-grown shoot tips of grapevine by encapsulation-dehydration. Plant Cell. Tissue Organ Cult. 2000, 63, 41–46. [Google Scholar] [CrossRef]
- Bahr, M.M.; Amer, M.S.; Abo-El-Sooud, K.; Abdallah, A.N.; El-Tookhy, O.S. Preservation techniques of stem cells extracellular vesicles: A gate for manufacturing of clinical grade therapeutic extracellular vesicles and long-term clinical trials. Int. J. Vet. Sci. Med. 2020, 8, 1–8. [Google Scholar] [CrossRef]
- Teixeira, A.S.; González, M.E. Tissue and cytoplasm vitrification in cryopreservation monitored by low temperature scanning electron microscopy (cryo-SEM). Curr. Microsc. Contrib. Adv. Sci. Technol. 2012, 2, 872–879. [Google Scholar]
- Zhang, J.M.; Han, L.; Lu, X.X.; Volk, G.M.; Xin, X.; Yin, G.K.; He, J.J.; Wang, L.; Chen, X.L. Cryopreservation of Jerusalem artichoke cultivars using an improved droplet-vitrification method. Plant Cell. Tissue Organ Cult. 2017, 128, 577–587. [Google Scholar] [CrossRef]
- Nishizawa, S. Cryopreservation of asparagus (Asparagus officinalis L.) embryogenic suspension cells and subsequent plant regeneration by a simple freezing method. Cryo Lett. 1992, 13, 379–388. [Google Scholar]
- Suranthran, P.; Gantait, S.; Sinniah, U.R.; Subramaniam, S.; Alwee, S.S.R.S.; Roowi, S.H. Effect of loading and vitrification solutions on survival of cryopreserved oil palm polyembryoids. Plant Growth Regul. 2012, 66, 101–109. [Google Scholar] [CrossRef]
- Kim, H.-H.; Lee, Y.-G.; Park, S.-U.; Lee, S.-C.; Baek, H.-J.; Cho, E.-G.; Engelmann, F. Development of alternative loading solutions in droplet-vitrification procedures. Cryo Lett. 2009, 30, 291–299. [Google Scholar]
- Panis, B.; Strosse, H.; Van Den Hende, S.; Swennen, R. Sucrose preculture to simplify cryopreservation of banana meristem cultures. Cryo Lett. 2002, 23, 375–384. [Google Scholar]
- Vollmer, R.; Villagaray, R.; Castro, M.; Anglin, N.L.; Ellis, D. Cryopreserved potato shoot tips showed genotype-specific response to sucrose concentration in rewarming solution (RS). Plant Cell Tissue Organ Cult. 2019, 136, 353–363. [Google Scholar] [CrossRef]
- Santos, R.R.; Tharasanit, T.; Figueiredo, J.R.; van Haeften, T.; van den Hurk, R. Preservation of caprine preantral follicle viability after cryopreservation in sucrose and ethylene glycol. Cell Tissue Res. 2006, 325, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Nadarajan, J.; Mansor, M.; Krishnapillay, B.; Staines, H.J.; Benson, E.E.; Harding, K. Applications of differential scanning calorimetry in developing cryopreservation strategies for Parkia speciosa, a tropical tree producing recalcitrant seeds. Cryo Lett. 2008, 29, 95–110. [Google Scholar]
- Tao, D.; Li, P.H. Classification of plant cell cryoprotectants. J. Theor. Biol. 1986, 123, 305–310. [Google Scholar] [CrossRef]
- Jiwu, Z.; Ganjun, Y.; Qiuming, Z. Micropropagation and cryopreservation of in vitro shoot tips of “Suizhonghong” papaya. Acta Hortic. 2007, 760, 217–224. [Google Scholar] [CrossRef]
- Nishizawa, S.; Sakai, A.; Amano, Y.; Matsuzawa, T. Cryopreservation of asparagus (Asparagus officinalis L.) embryogenic suspension cells and subsequent plant regeneration by vitrification. Plant Sci. 1993, 91, 67–73. [Google Scholar] [CrossRef]
- Matsumoto, T.; Sakai, A.; Yamada, K. Cryopreservation of in vitro-grown apical meristems of lily by vitrification. Plant Cell. Tissue Organ Cult. 1995, 41, 237–241. [Google Scholar] [CrossRef]
- Uragami, A.; Sakai, A.; Nagai, M.; Takahashi, T. Survival of cultured cells and somatic embryos of Asparagus officinalis cryopreserved by vitrification. Plant Cell Rep. 1989, 8, 418–421. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Survival by vitrification of nucellar cells of navel orange (Citrus sinensis var. brasiliensis Tanaka) Cooled to −196 °C. J. Plant Physiol. 1991, 137, 465–470. [Google Scholar] [CrossRef]
- Sakai, A.; Matsumoto, T.; Hirai, D.; Niino, T. Newly developed encapsulation-dehydration protocol for plantcryopreservation. Cryo Lett. 2000, 21, 53–62. [Google Scholar]
- Meryman, H.T. Cryoprotective agents. Cryobiology 1971, 8, 173–183. [Google Scholar] [CrossRef]
- Ishikawa, K.; Harata, K.; Mii, M.; Sakai, A.; Yoshimatsu, K.; Shimomura, K. Cryopreservation of zygotic embryos of a Japanese terrestrial orchid (Bletilla striata) by vitrification. Plant Cell Rep. 1997, 16, 754–757. [Google Scholar] [CrossRef]
- Towill, L.E. Cryopreservation of isolated mint shoot tips by vitrification. Plant Cell Rep. 1990, 9, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Steponkus, P. Vitrification of Oryza sativa L. cell suspensions. Cryo Lett. 1995, 16, 255–262. [Google Scholar]
- Speeckaert, G.; Borges, A.V.; Gypens, N. Salinity and growth effects on dimethylsulfoniopropionate (DMSP) and dimethylsulfoxide (DMSO) cell quotas of Skeletonema costatum, Phaeocystis globosa and Heterocapsa triquetra. Estuar. Coast. Shelf Sci. 2019, 226, 106275. [Google Scholar] [CrossRef] [Green Version]
- Wohnhaas, C.T.; Leparc, G.G.; Fernandez-Albert, F.; Kind, D.; Gantner, F.; Viollet, C.; Hildebrandt, T.; Baum, P. DMSO cryopreservation is the method of choice to preserve cells for droplet-based single-cell RNA sequencing. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kulus, D.; Zalewska, M. Cryopreservation as a tool used in long-term storage of ornamental species—A review. Sci. Hortic. (Amsterdam) 2014, 168, 88–107. [Google Scholar] [CrossRef]
- Teixeira, A.S.; González-Benito, M.E.; Molina-García, A.D. Determination of glassy state by cryo-SEM and DSC in cryopreservation of mint shoot tips by encapsulation–dehydration. Plant Cell Tissue Organ Cult. 2014, 119, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Pomeisl, K.; Richter, J.; Golan, M.; Kratochvílová, I. Simple syntheses of new pegylated trehalose derivatives as a chemical tool for potential evaluation of cryoprotectant effects on cell membrane. Molecules 2020, 25, 497. [Google Scholar] [CrossRef] [Green Version]
- Shishova, N.V.; Davydova, G.A.; Kombarova, N.A.; Davydova, I.B.; Poltavtsev, A.M.; Zaraisky, E.I.; Poltavtseva, R.A. The use of nanosized liposomes from vegetable phospholipids in combination with albumine and some polysaccharides as cryoprotective agents in the course of cryopreservation. IOP Conf. Ser. Mater. Sci. Eng. 2020, 747, 012074. [Google Scholar] [CrossRef]
- Grondin, M.; Hamel, F.; Averill-Bates, D.A.; Sarhan, F. Wheat proteins improve cryopreservation of rat hepatocytes. Biotechnol. Bioeng. 2009, 103, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Bian, H.W.; Zhang, Y.X.; Cheng, H.P. The dual effect of antifreeze protein on cryopreservation of rice (Oryza sativa l.) embryogenic suspension cells. Cryo Lett. 2001, 22, 175–182. [Google Scholar]
- Nadri, T.; Towhidi, A.; Zeinoaldini, S.; Martínez-Pastor, F.; Mousavi, M.; Noei, R.; Tar, M.; Mohammadi Sangcheshmeh, A. Lecithin nanoparticles enhance the cryosurvival of caprine sperm. Theriogenology 2019, 133, 38–44. [Google Scholar] [CrossRef]
- Hou, Y.; Lu, C.; Dou, M.; Zhang, C.; Chang, H.; Liu, J.; Rao, W. Soft liquid metal nanoparticles achieve reduced crystal nucleation and ultrarapid rewarming for human bone marrow stromal cell and blood vessel cryopreservation. Acta Biomater. 2020, 102, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, H.; Stewart, S.; Jiang, B.; Ou, W.; Zhao, G.; He, X. Cold-responsive nanoparticle enables intracellular delivery and rapid release of trehalose for organic-solvent-free cryopreservation. Nano Lett. 2019, 19, 9051–9061. [Google Scholar] [CrossRef] [PubMed]
- Li, M. Enhancing the efficiency of a PCR using gold nanoparticles. Nucleic Acids Res. 2005, 33, e184. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Hartland, G.V. Heat dissipation for Au particles in aqueous solution: Relaxation time versus size. J. Phys. Chem. B 2003, 107, 1284–1284. [Google Scholar] [CrossRef] [Green Version]
- Montalbán, I.A.; Olarieta, A.C.; Casillas-Figueroa, F.; Arellano-García, M.E.; Chávez-Santoscoy, A.R.; Pestryakov, A.; Bogdanchikova, N.; Moncaleán, P. Simplified method to store embryogenic cells: Silver nanoparticles and cryoprotectors elimination effect. Cryobiology 2018, 85, 134. [Google Scholar] [CrossRef]
- Ren, L.; Deng, S.; Chu, Y.; Zhang, Y.; Zhao, H.; Chen, H.; Zhang, D. Single-wall carbon nanotubes improve cell survival rate and reduce oxidative injury in cryopreservation of Agapanthus praecox embryogenic callus. Plant Methods 2020, 16, 130. [Google Scholar] [CrossRef]
- Kulus, D.; Tymoszuk, A. Gold nanoparticles affect the cryopreservation efficiency of in vitro-derived shoot tips of bleeding heart. Plant Cell. Tissue Organ Cult. 2021, 1–15. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Digilio, A. Crioconservación de variedades nativas de Solanum tuberosum ssp. andigenum Juz. & Bukasov. Master’s Thesis, Universidad Nacional de Mar del Plata, Buenos Aires, Argentina, 2014. [Google Scholar]
- Panta, A.; Panis, B.; Ynouye, C.; Swennen, R.; Roca, W.; Tay, D.; Ellis, D. Improved cryopreservation method for the long-term conservation of the world potato germplasm collection. Plant Cell. Tissue Organ Cult. 2015, 120, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Niino, T.; Arizaga, M.V. Cryopreservation for preservation of potato genetic resources. Breed. Sci. 2015, 65, 41–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathew, L.; Burritt, D.J.; McLachlan, A.; Pathirana, R. Combined pre-treatments enhance antioxidant metabolism and improve survival of cryopreserved kiwifruit shoot tips. Plant Cell. Tissue Organ Cult. 2019, 138, 193–205. [Google Scholar] [CrossRef]
- Bi, W.L.; Pan, C.; Hao, X.Y.; Cui, Z.H.; Kher, M.M.; Marković, Z.; Wang, Q.C.; Teixeira da Silva, J.A. Cryopreservation of grapevine (Vitis spp.)—A review. Vitro Cell. Dev. Biol. Plant 2017, 53, 449–460. [Google Scholar] [CrossRef]
- Ibáñez, M.A.; Alvarez-Mari, A.; Rodríguez-Sanz, H.; Kremer, C.; González-Benito, M.E.; Martín, C. Genetic and epigenetic stability of recovered mint apices after several steps of a cryopreservation protocol by encapsulation-dehydration. A new approach for epigenetic analysis. Plant Physiol. Biochem. 2019, 143, 299–307. [Google Scholar] [CrossRef]
- Li, J.W.; Chen, H.Y.; Li, X.Y.; Zhang, Z.; Blystad, D.R.; Wang, Q.C. Cryopreservation and evaluations of vegetative growth, microtuber production and genetic stability in regenerants of purple-fleshed potato. Plant Cell. Tissue Organ Cult. 2017, 128, 641–653. [Google Scholar] [CrossRef]
- Yamamoto, S.; Rafique, T.; Priyantha, W.S.; Fukui, K.; Matsumoto, T.; Niino, T. Development of a cryopreservation procedure using aluminium cryo-plates. Cryo Lett. 2011, 32, 256–265. [Google Scholar]
- Niino, T.; Yamamoto, S.-I.; Fukui, K.; Martínez, C.R.C.; Arizaga, M.V.; Matsumoto, T.; Engelmann, F. Dehydration improves cryopreservation of mat rush (Juncus decipiens Nakai) basal stem buds on cryo-plates. Cryo Lett. 2013, 549–560. [Google Scholar]
- Matsumoto, T.; Yamamoto, S.-I.; Fukui, K.; Rafique, T.; Engelmann, F.; Niino, T. Cryopreservation of persimmon shoot tips from dormant buds using the D cryo-plate technique. Hortic. J. 2015, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Niino, T.; Watanabe, K.; Nohara, N.; Rafique, T.; Yamamoto, S.; Fukui, K.; Arizaga, M.V.; Martinez, C.R.C.; Matsumoto, T.; Engelmann, F.; et al. Cryopreservation of mat rush lateral buds by air dehydration using aluminum cryo-plate. Plant Biotechnol. 2014, 31, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Rafique, T.; Yamamoto, S.I.; Fukui, K.; Tanaka, D.; Arizaga, M.V.; Abbas, M.; Matsumoto, T.; Niino, T. Cryopreservation of shoot-tips from different sugarcane varieties using D cryo-plate technique. Pakistan J. Agric. Sci. 2016, 53, 151–158. [Google Scholar] [CrossRef]
- Nadarajan, J.; Pritchard, H.W. Biophysical characteristics of successful oilseed embryo cryoprotection and cryopreservation using vacuum infiltration vitrification: An innovation in plant cell preservation. PLoS ONE 2014, 9, 96169. [Google Scholar] [CrossRef] [PubMed]
- Vozovyk, K.; Bobrova, O.; Prystalov, A.; Shevchenko, N.; Kuleshova, L. Amorphous state stability of plant vitrification solutions. Biologija 2020, 66. [Google Scholar] [CrossRef]
- Kulus, D. Managing plant genetic resources using low and ultra-low temperature storage: A case study of tomato. Biodivers. Conserv. 2019, 28, 1003–1027. [Google Scholar] [CrossRef] [Green Version]
- Sgueglia, A.; Gentile, A.; Frattarelli, A.; Germanà, M.A.; Caboni, E. Cryopreservation of Italian cultivars of hazelnut. Acta Hortic. 2021, 1307, 159–162. [Google Scholar] [CrossRef]
- Bidabadi, S.S.; Mohan Jain, S. Cellular, molecular, and physiological aspects of in vitro plant regeneration. Plants 2020, 9, 702. [Google Scholar] [CrossRef]
- Manokari, M.; Priyadharshini, S.; Jogam, P.; Dey, A.; Shekhawat, M.S. Meta-topolin and liquid medium mediated enhanced micropropagation via ex vitro rooting in Vanilla planifolia Jacks. ex Andrews. Plant Cell. Tissue Organ Cult. 2021, 1–14. [Google Scholar] [CrossRef]
- Zaytseva, Y.G.; Ambros, E.V.; Novikova, T.I. Meta-topolin:Advantages and Disadvantages for In Vitro Propagation. In Meta-topolin: A Growth Regulator for Plant Biotechnology and Agriculture; Springer: Singapore, 2021; pp. 119–141. [Google Scholar]
- Ahmad, A.; Anis, M. Meta-topolin improves in vitro morphogenesis, rhizogenesis and biochemical analysis in Pterocarpus marsupium Roxb.: A potential drug-yielding tree. J. Plant Growth Regul. 2019, 38, 1007–1016. [Google Scholar] [CrossRef]
- Khanam, M.N.; Javed, S.B.; Anis, M.; Alatar, A.A. Meta-topolin induced in vitro regeneration and metabolic profiling in Allamanda cathartica L. Ind. Crops Prod. 2020, 145, 111944. [Google Scholar] [CrossRef]
- Edesi, J.; Maria Pirttilä, A.; Häggman, H. Modified light spectral conditions prior to cryopreservation alter growth characteristics and cryopreservation success of potato (Solanum tuberosum L.) shoot tips in vitro. Plant Cell Tissue Organ Cult. 2017, 128, 409–421. [Google Scholar] [CrossRef]
- Mølmann, J.A.B.; Johansen, T.J. Sprout growth inhibition and photomorphogenic development of potato seed tubers (Solanum tuberosum L.) under different LED light colours. Potato Res. 2020, 63, 199–215. [Google Scholar] [CrossRef]
- Volk, G.M. Application of functional genomics and proteomics to plant cryopreservation. Curr. Genomics 2010, 11, 24–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchendu, E.E.; Muminova, M.; Gupta, S.; Reed, B.M. Antioxidant and anti-stress compounds improve regrowth of cryopreserved Rubus shoot tips. Vitro Cell. Dev. Biol. Plant 2010, 46, 386–393. [Google Scholar] [CrossRef]
- Tymoszuk, A.; Miler, N. Silver and gold nanoparticles impact on in vitro adventitious organogenesis in chrysanthemum, gerbera and Cape Primrose. Sci. Hortic. (Amsterdam) 2019, 257. [Google Scholar] [CrossRef]
- Tymoszuk, A.; Wojnarowicz, J. Zinc oxide and zinc oxide nanoparticles impact on in vitro germination and seedling growth in Allium cepa L. Materials (Basel) 2020, 13, 2784. [Google Scholar] [CrossRef]
- Tian, H.; Ghorbanpour, M.; Kariman, K. Manganese oxide nanoparticle-induced changes in growth, redox reactions and elicitation of antioxidant metabolites in deadly nightshade (Atropa belladonna L.). Ind. Crops Prod. 2018, 126, 403–414. [Google Scholar] [CrossRef]
- Gupta, S.D.; Agarwal, A.; Pradhan, S. Phytostimulatory effect of silver nanoparticles (AgNPs) on rice seedling growth: An insight from antioxidative enzyme activities and gene expression patterns. Ecotoxicol. Environ. Saf. 2018, 161, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Tymoszuk, A.; Kulus, D. Silver nanoparticles induce genetic, biochemical, and phenotype variation in chrysanthemum. Plant Cell. Tissue Organ Cult. 2020, 143, 331–344. [Google Scholar] [CrossRef]
- Zhang, Z.; Ke, M.; Qu, Q.; Peijnenburg, W.J.G.M.; Lu, T.; Zhang, Q.; Ye, Y.; Xu, P.; Du, B.; Sun, L.; et al. Impact of copper nanoparticles and ionic copper exposure on wheat (Triticum aestivum L.) root morphology and antioxidant response. Environ. Pollut. 2018, 239, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Vandenbussche, B.; Weyens, G.; De Proft, M. Cryopreservation of in vitro sugar beet (Beta vulgaris L.) shoot tips by a vitrification technique. Plant Cell Rep. 2000, 19, 1064–1068. [Google Scholar] [CrossRef] [PubMed]
- Yamuna, G.; Sumathi, V.; Geetha, S.P.; Praveen, K.; Swapna, N.; Babu, K.N. Cryopreservation of in vitro grown shoots of ginger (Zingiber officinale Rosc.). Cryo Lett. 2007, 28, 241–252. [Google Scholar]
- Sánchez, D.F.; Panta, A.; Tay, D.; Roca, W. Cryopreservation of ulluco (Ullucus tuberosus cal.) and oca (Oxalis tuberosa Mol.) shoot tips using the PVS2 droplet-vitrification method. Acta Hortic. 2011, 908, 339–346. [Google Scholar] [CrossRef]
- Arizaga, M.V.; Yamamoto, S.-I.; Tanaka, D.; Fukui, K.; Nohara, N.; Nishikawa, T.; Watanabe, K.; Niino, T. Cryopreservation of in vitro shoot tips of ulluco (Ullucus tuberosus Cal.) using D cryo-plate method. Cryo Lett. 2017, 9, 419–427. [Google Scholar]
- Romadanova, N.; Kushnarenko, S.; Karasholakova, L. Development of a common PVS2 vitrification method for cryopreservation of several fruit and vegetable crops. Vitro Cell. Dev. Biol. Plant 2017, 53, 382–393. [Google Scholar] [CrossRef]
- Pipithsangchan, K.; Chareonsap, P.P.; Reanhatthakam, K.; Jutamas, P.; Narkprasert, D.; Thammasiri, K. Cryopreservation technique for lettuce, Chinese cabbage, flowering white cabbage, Chinese kale and pak choi seeds by encapsulation-dehydration. Acta Hortic. 2019, 1234, 189–194. [Google Scholar] [CrossRef]
- Senula, A.; Büchner, D.; Keller, E.R.J.; Nagel, M. An improved cryopreservation protocol for Mentha spp. based on PVS3 as the cryoprotectant. Cryo Lett. 2018, 9, 345–353. [Google Scholar]
- Pipithsangchan, K.; Chareonsap, P.P.; Reanhatthakam, K.; Jutamas, P.; Narkprasert, D.; Thammasiri, K. Cryopreservation technique of chilli seeds by vitrification. Acta Hortic. 2019, 1234, 145–148. [Google Scholar] [CrossRef]
- Pérez, J.; Araya-Valverde, E.; Garro, G.; Abdelnour-Esquivel, A. Analysis of stress indicators during cryopreservation of seeds of landrace maize (Zea mays). Cryo Lett. 2017, 38, 445–454. [Google Scholar]
- Arguedas, M.; Villalobos, A.; Gómez, D.; Hernández, L.; Zevallos, B.E.; Cejas, I.; Yabor, L.; Martínez-Montero, M.E.; Lorenzo, J.C. Field performance of cryopreserved seed-derived maize plants. Cryo Lett. 2018, 39, 366–370. [Google Scholar]
- Bettoni, J.C.; Bonnari, R.; Shepherd, A.; Kretzschmar, A.A.; Volk, G.M. Cryopreservation of grapevine (Vitis spp.) shoot tips from growth chamber-sourced plants and histological observations. Vitis 2019, 58, 71–78. [Google Scholar] [CrossRef]
- Wang, M.-R.; Zhang, Z.; Haugslien, S.; Sivertsen, A.; Rasmussen, M.; Wang, Q.-C.; Blystad, D.-R. Cryopreservation of shallot (Allium cepa var. aggregatum) shoot tips by droplet-vitrification. Acta Hortic. 2019, 1234, 241–248. [Google Scholar] [CrossRef]
- Liu, X.-X.; Mou, S.-W.; Cheng, Z.-H. Effect of cryopreservation on plant growth, bulb characteristics, and virus reduction of garlic (Allium Sativum L.). Cryo Lett. 2019, 40, 322–332. [Google Scholar]
- Keller, E.R.J. Improvement of cryopreservation results in garlic using low temperature preculture and high-quality in vitro plantlets. Cryo Lett. 2005, 26, 357–366. [Google Scholar]
- Kim, H.H.; Lee, J.K.; Yoon, J.W.; Ji, J.J.; Nam, S.S.; Hwang, H.S.; Cho, E.G.; Engelmann, F. Cryopreservation of garlic bulbil primordia by the droplet-vitrification procedure. Cryo Lett. 2006, 27, 143–153. [Google Scholar]
- Kim, H.H.; Popova, E.; Shin, D.J.; Yi, J.Y.; Kim, C.H.; Lee, J.S.; Yoon, M.K.; Engelmann, F. Cryobanking of Korean allium germplasm collections: Results from a 10 year experience. Cryo Lett. 2012, 33, 45–57. [Google Scholar]
- Kaczmarczyk, A.; Rokka, V.-M.; Keller, E.R.J. Potato Shoot Tip Cryopreservation. A Review. Potato Res. 2011, 54, 45–79. [Google Scholar] [CrossRef] [Green Version]
- Bissati, S.; Boudjenah, S.; Morisset, C.; Chenchouni, H. Does pre-culture in sugar-rich media affect carbohydrate content and post-thawing recovery rate of cryopreserved potato (Solanum phureja) shoot tips? J. King Saud Univ. Sci. 2020, 32, 1917–1924. [Google Scholar] [CrossRef]
- Uchendu, E.E.; Shukla, M.; Saxena, P.K.; Keller, J.E.R. Cryopreservation of potato microtubers: The critical roles of sucrose and desiccation. Plant Cell Tissue Organ Cult. 2016, 124, 649–656. [Google Scholar] [CrossRef]
- Keller, E.R.J.; Dreiling, M. Potato cryopreservation in Germany—Using the droplet method for the establishment of a new large collection. Acta Hortic. 2003, 623, 193–200. [Google Scholar] [CrossRef]
- Kushnarenko, S.; Romadanova, N.; Aralbayeva, M.; Zholamanova, S.; Alexandrova, A.; Karpova, O. Combined ribavirin treatment and cryotherapy for efficient Potato virus M and Potato virus S eradication in potato (Solanum tuberosum L.) in vitro shoots. Vitro Cell. Dev. Biol. Plant 2017, 53, 425–432. [Google Scholar] [CrossRef]
- Ayala Hernandez, D.D.; Ruiz Sáenz, D.R.; Cruz Gutiérrez, E.J.; Lopez Delgado, H.A. Ácido salicílico induce tolerancia al estrés por criogenia en Solanum tuberosum. Rev. Mex. Ciencias Agrícolas 2019, 10, 1505–1515. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.-R.; Hamborg, Z.; Slimestad, R.; Elameen, A.; Blystad, D.-R.; Haugslien, S.; Skjeseth, G.; Wang, Q.-C. Assessments of rooting, vegetative growth, bulb production, genetic integrity and biochemical compounds in cryopreserved plants of shallot. Plant Cell Tissue Organ Cult. 2021, 144, 123–131. [Google Scholar] [CrossRef]
- Liu, X.-X.; Wen, Y.-B.; Cheng, Z.-H.; Mou, S.-W. Establishment of a garlic cryopreservation protocol for shoot apices from adventitious buds in vitro. Sci. Hortic. (Amsterdam) 2017, 226, 10–18. [Google Scholar] [CrossRef]
- Seo, J.H.; Naing, A.H.; Jeon, S.M.; Kim, C.K. Anti-freezing-protein type III strongly influences the expression of relevant genes in cryopreserved potato shoot tips. Plant Mol. Biol. 2018, 97, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Köpnick, C.; Grübe, M.; Stock, J.; Senula, A.; Mock, H.-P.; Nagel, M. Changes of soluble sugars and ATP content during DMSO droplet freezing and PVS3 droplet vitrification of potato shoot tips. Cryobiology 2018, 85, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Lazo Javalera, M.F.; Tiznado Hernández, M.E.; Vargas Arispuro, I.; Martínez Téllez, M.Á.; Islas Osuna, M.A.; Hernández Oñate, M.Á.; Rivera Domínguez, M. Genetic stability of cryopreserved grapevine (Vitis vinifera L.) genome by vitrification method. J. Agric. Sci. Technol. B 2016, 6, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Edesi, J.; Kotkas, K.; Pirttilä, A.M.; Häggman, H. Does light spectral quality affect survival and regeneration of potato (Solanum tuberosum L.) shoot tips after cryopreservation? Plant Cell. Tissue Organ Cult. 2014, 119, 599–607. [Google Scholar] [CrossRef]
- Folgado, R.; Sergeant, K.; Renaut, J.; Swennen, R.; Hausman, J.-F.; Panis, B. Changes in sugar content and proteome of potato in response to cold and dehydration stress and their implications for cryopreservation. J. Proteomics 2014, 98, 99–111. [Google Scholar] [CrossRef]
- Folgado, R.; Panis, B.; Sergeant, K.; Renaut, J.; Swennen, R.; Hausman, J.-F. Unravelling the effect of sucrose and cold pretreatment on cryopreservation of potato through sugar analysis and proteomics. Cryobiology 2015, 71, 432–441. [Google Scholar] [CrossRef]
- Tanaka, D.; Sakuma, Y.; Yamamoto, S.; Valle Arizaga, M.; Niino, T.; Matsumoto, T. Development of –80 °C storage for Allium shoot tips using D cryo-plate method. Plant Cell Tissue Organ Cult. 2021, 144, 115–122. [Google Scholar] [CrossRef]
- Yamamoto, S.I.; Rafique, T.; Arizaga, M.V.; Fukui, K.; Gutierrez, E.J.C.; Martinez, C.R.C.; Watanabe, K.; Niino, T. The aluminum cryo-plate increases efficiency of cryopreservation protocols for potato shoot tips. Am. J. Potato Res. 2015, 92, 250–257. [Google Scholar] [CrossRef]
- Morel, G.; Wetmore, R.H. Fern callus tissue culture. Am. J. Bot. 1951, 38, 141–143. [Google Scholar] [CrossRef]
- Tendille, C.; Lecerf, M. La multiplication vegetative de L’Asperge (Asparagus officinalis L.). action de divers facteurs, en particu-lier de la nutrition minerale, sur le developpement des meristemes d’asperge, sur la croissance des plantu-les issues de ces meristemes et sur la p. ANN. Amelior. Plantes 1974, 24, 269–282. [Google Scholar]
- Ren, R.; Li, Z.; Zhang, L.; Zhou, H.; Jiang, X.; Liu, Y. Enzymatic and nonenzymatic antioxidant systems impact the viability of cryopreserved Paeonia suffruticosa pollen. Plant Cell. Tissue Organ Cult. 2021, 144, 233–246. [Google Scholar] [CrossRef]
- Wen, B.; Cai, C.; Wang, R.; Song, S.; Song, J. Cytological and physiological changes in recalcitrant Chinese fan palm (Livistona chinensis) embryos during cryopreservation. Protoplasma 2012, 249, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Wang, R.; Cheng, H.; Song, S. Cytological and physiological changes in orthodox maize embryos during cryopreservation. Protoplasma 2010, 239, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Subbarayan, K.; Rolletschek, H.; Senula, A.; Ulagappan, K.; Hajirezaei, M.R.; Keller, E.R.J. Influence of oxygen deficiency and the role of specific amino acids in cryopreservation of garlic shoot tips. BMC Biotechnol. 2015, 15, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solov’eva, A.I.; Dolgikh, Y.I.; Vysotskaya, O.N.; Popov, A.S. Patterns of ISSR and REMAP DNA markers after cryogenic preservation of spring wheat calli by dehydration method. Russ. J. Plant Physiol. 2011, 58, 423–430. [Google Scholar] [CrossRef]
- Maki, S.; Hirai, Y.; Niino, T.; Matsumoto, T. Assessment of molecular genetic stability between long-term cryopreserved and tissue cultured wasabi (Wasabia japonica) plants. Cryo Lett. 2015, 36, 318–324. [Google Scholar]
- Zevallos, B.; Cejas, I.; Valle, B.; Yabor, L.; Aragón, C.; Engelmann, F.; Martínez, M.E.; Lorenzo, J.C. Short-term liquid nitrogen storage of wild tomato (Solanum lycopersicum Mill.) seeds modifies the levels of phenolics in 7 day-old seedlings. Sci. Hortic. (Amsterdam) 2013, 160, 264–267. [Google Scholar] [CrossRef]
- Villalobos, A.; Arguedas, M.; Escalante, D.; Martínez, J.; Zevallos, B.E.; Cejas, I.; Yabor, L.; Martínez-Montero, M.S.E.; Lorenzo, J.C. Cryopreservation of sorghum seeds modifies germination and seedling growth but not field performance of adult plants. Artic. J. Appl. Bot. Food Qual. 2019, 92, 94–99. [Google Scholar] [CrossRef]
- Ritonga, F.N.; Chen, S. Physiological and molecular mechanism involved in cold stress tolerance in plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef] [PubMed]


| Cryopreservation Method | |
|---|---|
| Classic Methods (Slow Cooling) | Modern Methods (Rapid Cooling) |
| Establishment of in vitro culture or disinfection of tissues and organs taken directly from the donor plant | |
| Preculture of explants on a medium with an increased osmotic pressure and optional cold-hardening of explants | |
| Treatment with an appropriate cryoprotectant, usually DMSO (concentration and treatment duration vary depending on the plant material) | Dehydration with a mixture of diluted then concentrated cryoprotectants (after an optional encapsulation of explants in alginate) or dehydration in a series of sucrose solutions (with increasing concentration), followed by air drying |
| Gradual and slow cooling at a certain cooling rate (0.2–2 °C·min−1), which can be achieved by commercially available cryostats. This cooling is periodic up to −20 °C, −40 °C, −70 °C, −100 °C and, finally, −196 °C and at defined intervals | Fast cooling by direct immersion of the explants in LN |
| Storage of specimens in vials or straws in cryo-boxes in LN or, less often, its vapor phase | |
| Rewarming of samples (rapid in a water bath or at room temperature) | |
| Elimination of the cryoprotectant by washing with a solution of high sucrose concentration (usually 1.2 M) | |
| Determination of viability (histochemically or by growth observation) | |
| In vitro recovery of plants on a PGRs-supplemented media (usually at reduced light conditions during the first two weeks of culture) | |
| Acclimatization and transfer to ex vitro conditions | |
| Solution | Component (% w/v) | Reference | ||
|---|---|---|---|---|
| DMSO | Glycerol | Sucrose | ||
| LS1 | - | 18.4 | 13.7 | [72] |
| LS2 | 5.0 | 13.8 | 13.7 | [73] |
| LS3 | 10.0 | 4.6 | 10.3 | |
| LS4 | 10.0 | - | 24.0 | |
| Solution | Component (% w/v) | Reference | |||||
|---|---|---|---|---|---|---|---|
| Sorbitol | EG | DMSO | PEG | Glycerol | Sucrose | ||
| PVS1 | 9.1 | 15.0 | 7.0 | 15.0 | 22.0 | - | [74] |
| PVS2 | - | 15.0 | 15.0 | - | 30.0 | 13.7 | [75] |
| PVS3 | - | - | - | - | 50.0 | 50.0 | [72] |
| PVS4 | - | 20.0 | - | - | 35.0 | 20.5 | [37] |
| L-solution | - | 30.0 | 7.0 | - | 22.0 | 15.0 | [78] |
| T-solution | - | 35.0 | 7.8 | 10.0 | - | - | [79] |
| W-solution | 18.7 | - | 44.5 | - | - | - | [80] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roque-Borda, C.A.; Kulus, D.; Vacaro de Souza, A.; Kaviani, B.; Vicente, E.F. Cryopreservation of Agronomic Plant Germplasm Using Vitrification-Based Methods: An Overview of Selected Case Studies. Int. J. Mol. Sci. 2021, 22, 6157. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116157
Roque-Borda CA, Kulus D, Vacaro de Souza A, Kaviani B, Vicente EF. Cryopreservation of Agronomic Plant Germplasm Using Vitrification-Based Methods: An Overview of Selected Case Studies. International Journal of Molecular Sciences. 2021; 22(11):6157. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116157
Chicago/Turabian StyleRoque-Borda, Cesar Augusto, Dariusz Kulus, Angela Vacaro de Souza, Behzad Kaviani, and Eduardo Festozo Vicente. 2021. "Cryopreservation of Agronomic Plant Germplasm Using Vitrification-Based Methods: An Overview of Selected Case Studies" International Journal of Molecular Sciences 22, no. 11: 6157. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116157