
Agrobacterium tumefaciens-Mediated Nuclear Transformation of a Biotechnologically Important Microalga—Euglena gracilis


Abstract
:1. Introduction
2. Results
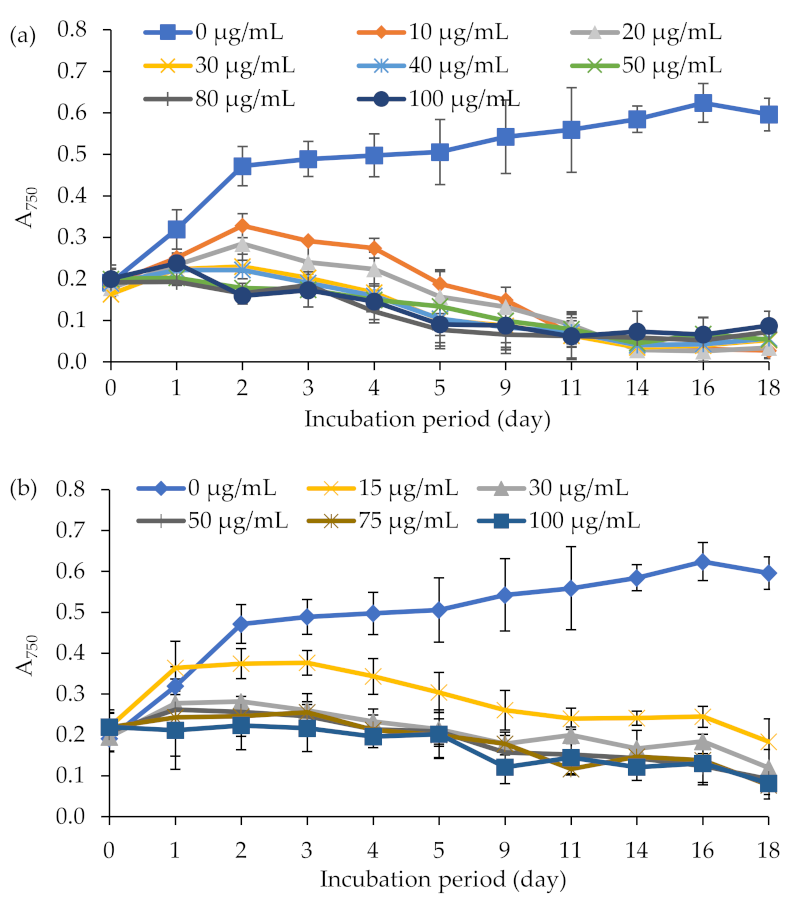
2.1. Sensitivity of E. gracilis and A. tumefaciens to Different Antibiotics
2.2. Transformation of E. gracilis via A. tumefaciens
2.3. Analysis of Transgene Integration and Expression
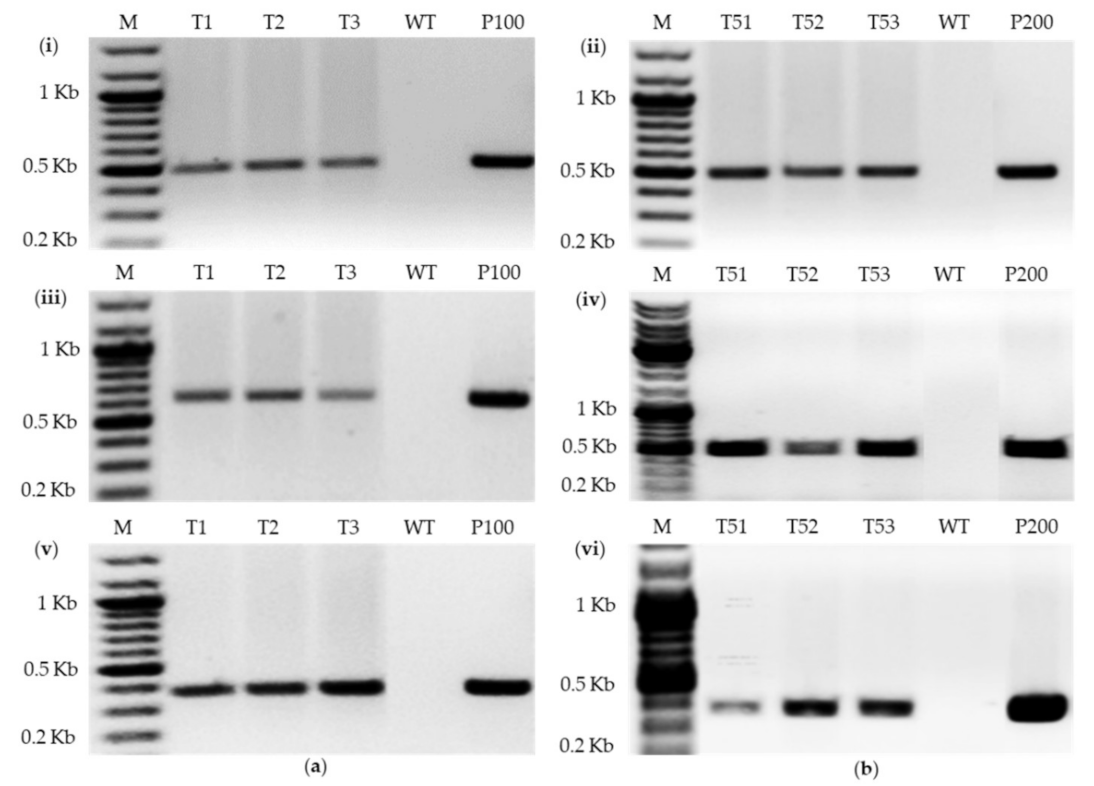
2.3.1. PCR of Transformants
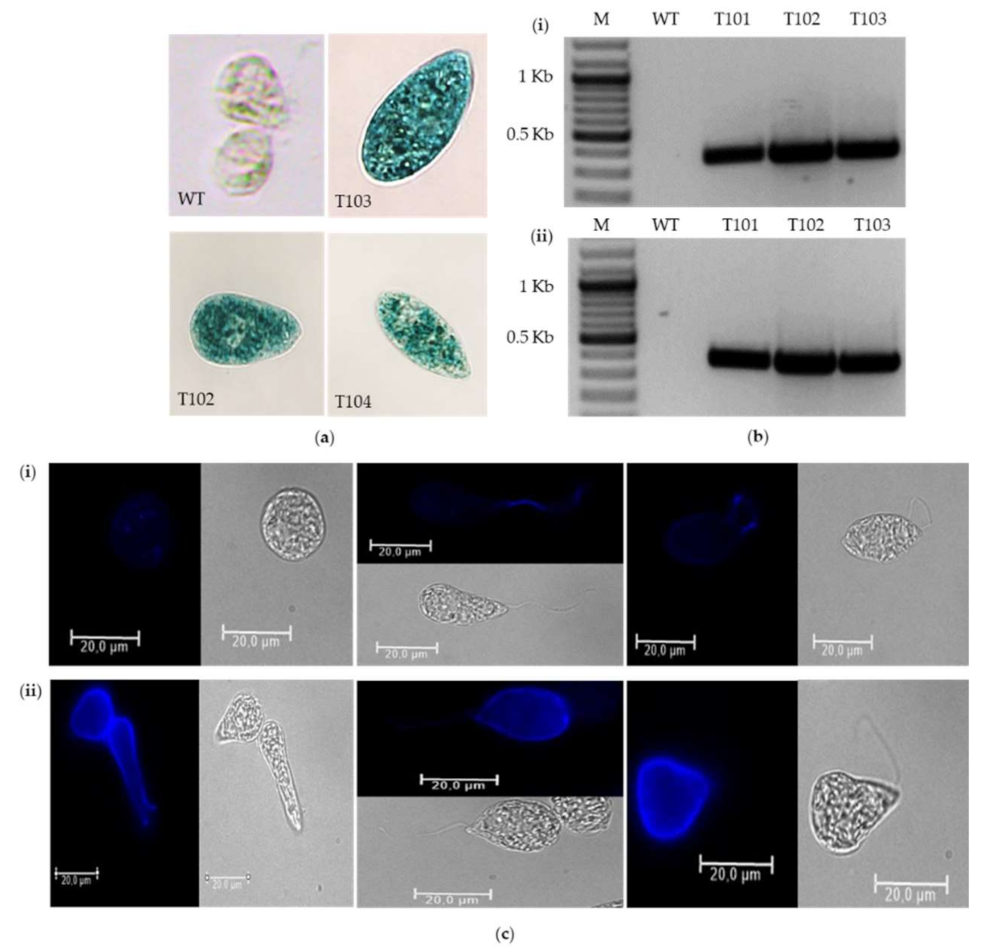
2.3.2. Expression of Reporter Genes
2.4. Optimization of Factors in the ATMT Procedure of E. gracilis
2.5. Genetic Stability of Transformed Clones
3. Discussion
4. Materials and Methods
4.1. Growth Conditions of E. gracilis
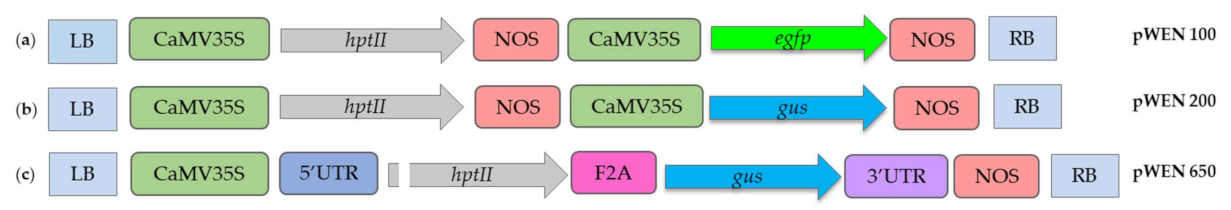
4.2. Vector Systems and Bacterial Strains
4.3. Sensitivity of Microbial Strains to Antibiotics
4.4. Transformation of Euglena
4.5. Confirmation of A. tumefaciens’ Absence in Hygromycin-Resistant E. gracilis
4.6. PCR Analysis of Transformants
4.7. Analysis of Reporter Genes Expression in Hygromycin-Resistant Cells
4.7.1. EGFP Fluorescence Analysis
4.7.2. Determination of GUS Activity
4.8. Indirect Immunofluorescence Assay
4.9. Optimization of Transformation Parameters
4.10. Maintenance and Stability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zakryś, B.; Milanowski, R.; Karnkowska, A. Evolutionary Origin of Euglena. Adv. Exp. Med. Biol. 2017, 979, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Daiker, V.; Häder, D.-P.; Richter, P.R.; Lebert, M. The involvement of a protein kinase in phototaxis and gravitaxis of Euglena gracilis. Planta 2011, 233, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Daiker, V.; Lebert, M.; Richter, P.; Häder, D.-P. Molecular characterization of a calmodulin involved in the signal transduction chain of gravitaxis in Euglena gracilis. Planta 2010, 231, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Häder, D.-P.; Richter, P.R.; Schuster, M.; Daiker, V.; Lebert, M. Molecular analysis of the graviperception signal transduction in the flagellate Euglena gracilis: Involvement of a transient receptor potential-like channel and a calmodulin. Adv. Space Res. 2009, 43, 1179–1184. [Google Scholar] [CrossRef]
- Ntefidou, M.; Iseki, M.; Watanabe, M.; Lebert, M.; Häder, D.-P. Photoactivated adenylyl cyclase controls phototaxis in the flagellate Euglena gracilis. Plant Physiol. 2003, 133, 1517–1521. [Google Scholar] [CrossRef] [Green Version]
- Lebert, M.; Richter, P.; Häder, D.-P. Signal perception and transduction of gravitaxis in the flagellate Euglena gracilis. J. Plant Physiol. 1997, 150, 685–690. [Google Scholar] [CrossRef]
- Nasir, A.; Strauch, S.M.; Becker, I.; Sperling, A.; Schuster, M.; Richter, P.R.; Weißkopf, M.; Ntefidou, M.; Daiker, V.; An, Y.A.; et al. The influence of microgravity on Euglena gracilis as studied on Shenzhou 8. Plant Biol. 2014, 16 (Suppl. 1), 113–119. [Google Scholar] [CrossRef]
- Strauch, S.M.; Richter, P.; Schuster, M.; Häder, D.-P. The beating pattern of the flagellum of Euglena gracilis under altered gravity during parabolic flights. J. Plant Physiol. 2010, 167, 41–46. [Google Scholar] [CrossRef]
- Furuhashi, T.; Ogawa, T.; Nakai, R.; Nakazawa, M.; Okazawa, A.; Padermschoke, A.; Nishio, K.; Hirai, M.Y.; Arita, M.; Ohta, D. Wax ester and lipophilic compound profiling of Euglena gracilis by gas chromatography-mass spectrometry: Toward understanding of wax ester fermentation under hypoxia. Metabolomics 2015, 11, 175–183. [Google Scholar] [CrossRef]
- Krajčovič, J.; Matej, V.; Schwartzbach, S.D. Euglenoid flagellates: A multifaceted biotechnology platform. J. Biotechnol. 2015, 202, 135–145. [Google Scholar] [CrossRef]
- O’Neill, E.C.; Trick, M.; Hill, L.; Rejzek, M.; Dusi, R.G.; Hamilton, C.J.; Zimba, P.V.; Henrissat, B.; Field, R.A. The transcriptome of Euglena gracilis reveals unexpected metabolic capabilities for carbohydrate and natural product biochemistry. Mol. Biosyst. 2015, 11, 2808–2820. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, E.C.; Trick, M.; Henrissat, B.; Field, R.A. Euglena in time: Evolution, control of central metabolic processes and multi-domain proteins in carbohydrate and natural product biochemistry. Perspect. Sci. 2015, 6, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Schwartzbach, S.D.; Shigeoka, S. Euglena: Biochemistry, Cell and Molecular Biology; Springer International Publishing: Cham, Switzerland, 2017; ISBN 978-3-319-54908-8. [Google Scholar]
- Grimm, P.; Risse, J.M.; Cholewa, D.; Müller, J.M.; Beshay, U.; Friehs, K.; Flaschel, E. Applicability of Euglena gracilis for biorefineries demonstrated by the production of α-tocopherol and paramylon followed by anaerobic digestion. J. Biotechnol. 2015, 215, 72–79. [Google Scholar] [CrossRef]
- Kondo, Y.; Kato, A.; Hojo, H.; Nozoe, S.; Takeuchi, M.; Ochi, K. Cytokine-related immunopotentiating activities of paramylon, a beta-(1→3)-D-glucan from Euglena gracilis. J. Pharmacobiodyn. 1992, 15, 617–621. [Google Scholar] [CrossRef] [Green Version]
- Koizumi, N.; Sakagami, H.; Utsumi, A.; Fujinaga, S.; Takeda, M.; Asano, K.; Sugawara, I.; Ichikawa, S.; Kondo, H.; Mori, S.; et al. Anti-HIV (human immunodeficiency virus) activity of sulfated paramylon. Antivir. Res. 1993, 21, 1–14. [Google Scholar] [CrossRef]
- Nakashima, H.; Ohshiro, Y.; Miyano, N.; Yamamoto, N.; Ichikawa, S.; Kondo, H.; Takeda, M.; Sakagami, H. Synergistic inhibition of human immunodeficiency virus type 1 (HIV-1) replication in vitro by sulphated paramylon and 3′-azido-2′, 3′-dideoxythymidine (AZT). Lett. Appl. Microbiol. 1994, 18, 24–26. [Google Scholar] [CrossRef]
- Nakashima, A.; Suzuki, K.; Asayama, Y.; Konno, M.; Saito, K.; Yamazaki, N.; Takimoto, H. Oral administration of Euglena gracilis Z and its carbohydrate storage substance provides survival protection against influenza virus infection in mice. Biochem. Biophys. Res. Commun. 2017, 494, 379–383. [Google Scholar] [CrossRef]
- Sugiyama, A.; Hata, S.; Suzuki, K.; Yoshida, E.; Nakano, R.; Mitra, S.; Arashida, R.; Asayama, Y.; Yabuta, Y.; Takeuchi, T. Oral administration of paramylon, a beta-1,3-D-glucan isolated from Euglena gracilis Z inhibits development of atopic dermatitis-like skin lesions in NC/Nga mice. J. Vet. Med. Sci. 2010, 72, 755–763. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, A.; Suzuki, K.; Mitra, S.; Arashida, R.; Yoshida, E.; Nakano, R.; Yabuta, Y.; Takeuchi, T. Hepatoprotective effects of paramylon, a beta-1, 3-D-glucan isolated from Euglena gracilis Z, on acute liver injury induced by carbon tetrachloride in rats. J. Vet. Med. Sci. 2009, 71, 885–890. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Shimada, R.; Matsuyama, A.; Yuasa, M.; Sawamura, H.; Yoshida, E.; Suzuki, K. Antitumor activity of the β-glucan paramylon from Euglena against preneoplastic colonic aberrant crypt foci in mice. Food Funct. 2013, 4, 1685–1690. [Google Scholar] [CrossRef]
- Sakagami, H.; Kikuchi, K.; Takeda, M.; Sato, T.; Ichikawa, S.; Fujimaki, M.; Wada, C.; Komatsu, N. Macrophage stimulation activity of antimicrobial N,N-dimethylaminoethyl paramylon. In Vivo 1991, 5, 101–105. [Google Scholar]
- Yamada, K.; Suzuki, H.; Takeuchi, T.; Kazama, Y.; Mitra, S.; Abe, T.; Goda, K.; Suzuki, K.; Iwata, O. Efficient selective breeding of live oil-rich Euglena gracilis with fluorescence-activated cell sorting. Sci. Rep. 2016, 6, 26327. [Google Scholar] [CrossRef] [Green Version]
- Miazek, K.; Iwanek, W.; Remacle, C.; Richel, A.; Goffin, D. Effect of Metals, Metalloids and Metallic Nanoparticles on Microalgae Growth and Industrial Product Biosynthesis: A Review. Int. J. Mol. Sci. 2015, 16, 23929–23969. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Zavala, J.S.; García-García, J.D.; Ortiz-Cruz, M.A.; Moreno-Sánchez, R. Molecular mechanisms of resistance to heavy metals in the protist Euglena gracilis. J. Environ. Sci. Health Part A 2007, 42, 1365–1378. [Google Scholar] [CrossRef]
- Khatiwada, B.; Sunna, A.; Nevalainen, H. Molecular tools and applications of Euglena gracilis: From biorefineries to bioremediation. Biotechnol. Bioeng. 2020, 117, 3952–3967. [Google Scholar] [CrossRef]
- Ebenezer, T.E.; Carrington, M.; Lebert, M.; Kelly, S.; Field, M.C. Euglena gracilis Genome and Transcriptome: Organelles, Nuclear Genome Assembly Strategies and Initial Features. Adv. Exp. Med. Biol. 2017, 979, 125–140. [Google Scholar] [CrossRef]
- Ebenezer, T.E.; Zoltner, M.; Burrell, A.; Nenarokova, A.; Novák Vanclová, A.M.G.; Prasad, B.; Soukal, P.; Santana-Molina, C.; O’Neill, E.; Nankissoor, N.N.; et al. Transcriptome, proteome and draft genome of Euglena gracilis. BMC Biol. 2019, 17, 11. [Google Scholar] [CrossRef] [Green Version]
- Khatiwada, B.; Kautto, L.; Sunna, A.; Sun, A.; Nevalainen, H. Nuclear transformation of the versatile microalga Euglena gracilis. Algal Res. 2019, 37, 178–185. [Google Scholar] [CrossRef]
- Iseki, M.; Matsunaga, S.; Murakami, A.; Ohno, K.; Shiga, K.; Yoshida, K.; Sugai, M.; Takahashi, T.; Hori, T.; Watanabe, M. A blue-light-activated adenylyl cyclase mediates photoavoidance in Euglena gracilis. Nature 2002, 415, 1047–1051. [Google Scholar] [CrossRef]
- Tanaka, Y.; Ogawa, T.; Maruta, T.; Yoshida, Y.; Arakawa, K.; Ishikawa, T. Glucan synthase-like 2 is indispensable for paramylon synthesis in Euglena gracilis. FEBS Lett. 2017, 591, 1360–1370. [Google Scholar] [CrossRef] [Green Version]
- Tomiyama, T.; Kurihara, K.; Ogawa, T.; Maruta, T.; Ogawa, T.; Ohta, D.; Sawa, Y.; Ishikawa, T. Wax Ester Synthase/Diacylglycerol Acyltransferase Isoenzymes Play a Pivotal Role in Wax Ester Biosynthesis in Euglena gracilis. Sci. Rep. 2017, 7, 13504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakazawa, M.; Haruguchi, D.; Ueda, M.; Miyatake, K. Transformed Euglena and Process for Producing Same. U.S. Patent Application US 14/655,228, 24 December 2015. [Google Scholar]
- Nomura, T.; Yoshikawa, M.; Suzuki, K.; Mochida, K. Highly Efficient CRISPR-Associated Protein 9 Ribonucleoprotein-Based Genome Editing in Euglena gracilis. STAR Protoc. 2020, 1, 100023. [Google Scholar] [CrossRef] [PubMed]
- Cha, T.S.; Yee, W.; Aziz, A. Assessment of factors affecting Agrobacterium-mediated genetic transformation of the unicellular green alga, Chlorella vulgaris. World J. Microbiol. Biotechnol. 2012, 28, 1771–1779. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.; Vadakedath, N.; Jeong, H.-J.; General, T.; Cho, M.-G.; Lein, W. Agrobacterium tumefaciens-mediated genetic transformation of haptophytes (Isochrysis species). Appl. Microbiol. Biotechnol. 2014, 98, 8629–8639. [Google Scholar] [CrossRef]
- Doron, L.; Segal, N.; Shapira, M. Transgene Expression in Microalgae-From Tools to Applications. Front. Plant Sci. 2016, 7, 505. [Google Scholar] [CrossRef]
- Pratheesh, P.T.; Vineetha, M.; Kurup, G.M. An efficient protocol for the Agrobacterium-mediated genetic transformation of microalga Chlamydomonas reinhardtii. Mol. Biotechnol. 2014, 56, 507–515. [Google Scholar] [CrossRef]
- Kathiresan, S.; Chandrashekar, A.; Ravishankar, G.A.; Sarada, R. Agrobacterium-mediated transformation in the green alga Haematococcus Pluvialis (Chlorophyceae, Volvocales)(1). J. Phycol. 2009, 45, 642–649. [Google Scholar] [CrossRef]
- Úbeda-Mínguez, P.; Chileh, T.; Dautor, Y.; García-Maroto, F.; Alonso, D.L. Tools for microalgal biotechnology: Development of an optimized transformation method for an industrially promising microalga—Tetraselmis chuii. J. Appl. Phycol. 2015, 27, 223–232. [Google Scholar] [CrossRef]
- Rathod, J.P.; Prakash, G.; Pandit, R.; Lali, A.M. Agrobacterium-mediated transformation of promising oil-bearing marine algae Parachlorella kessleri. Photosynth. Res. 2013, 118, 141–146. [Google Scholar] [CrossRef]
- Ng, I.-S.; Tan, S.-I.; Kao, P.-H.; Chang, Y.-K.; Chang, J.-S. Recent Developments on Genetic Engineering of Microalgae for Biofuels and Bio-Based Chemicals. Biotechnol. J. 2017, 12. [Google Scholar] [CrossRef]
- Cheng, R.; Ma, R.; Li, K.; Rong, H.; Lin, X.; Wang, Z.; Yang, S.; Ma, Y. Agrobacterium tumefaciens mediated transformation of marine microalgae Schizochytrium. Microbiol. Res. 2012, 167, 179–186. [Google Scholar] [CrossRef]
- Prasad, B.; Lein, W.; Lindenberger, C.P.; Buchholz, R.; Vadakedath, N. An optimized method and a dominant selectable marker for genetic engineering of an industrially promising microalga—Pavlova lutheri. J. Appl. Phycol. 2019, 31, 1163–1174. [Google Scholar] [CrossRef]
- Prasad, B.; Lein, W.; Thiyam, G.; Lindenberger, C.P.; Buchholz, R.; Vadakedath, N. Stable nuclear transformation of rhodophyte species Porphyridium purpureum: Advanced molecular tools and an optimized method. Photosynth. Res. 2019, 140, 173–188. [Google Scholar] [CrossRef]
- Bourras, S.; Rouxel, T.; Meyer, M. Agrobacterium tumefaciens Gene Transfer: How a Plant Pathogen Hacks the Nuclei of Plant and Nonplant Organisms. Phytopathology 2015, 105, 1288–1301. [Google Scholar] [CrossRef] [Green Version]
- Tzfira, T.; Citovsky, V. Agrobacterium-mediated genetic transformation of plants: Biology and biotechnology. Curr. Opin. Biotechnol. 2006, 17, 147–154. [Google Scholar] [CrossRef]
- Jefferson, R.A. Assaying chimeric genes in plants: The GUS gene fusion system. Plant Mol. Biol. Rep. 1987, 5, 387–405. [Google Scholar] [CrossRef]
- Starr, R.C. The Culture Collection of Algae at Indiana University. Am. J. Bot. 1964, 51, 1013. [Google Scholar] [CrossRef]
- Gangl, D.; Zedler, J.A.Z.; Rajakumar, P.D.; Martinez, E.M.R.; Riseley, A.; Włodarczyk, A.; Purton, S.; Sakuragi, Y.; Howe, C.J.; Jensen, P.E.; et al. Biotechnological exploitation of microalgae. J. Exp. Bot. 2015, 66, 6975–6990. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, T.; Tamoi, M.; Kimura, A.; Mine, A.; Sakuyama, H.; Yoshida, E.; Maruta, T.; Suzuki, K.; Ishikawa, T.; Shigeoka, S. Enhancement of photosynthetic capacity in Euglena gracilis by expression of cyanobacterial fructose-1,6-/sedoheptulose-1,7-bisphosphatase leads to increases in biomass and wax ester production. Biotechnol. Biofuels 2015, 8, 80. [Google Scholar] [CrossRef] [Green Version]
- Anila, N.; Chandrashekar, A.; Ravishankar, G.A.; Sarada, R. Establishment of Agrobacterium tumefaciens-mediated genetic transformation in Dunaliella bardawil. Eur. J. Phycol. 2011, 46, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Karami, O. Factors affecting Agrobacterium-mediated transformation of plants. Transgenic Plant J. 2008, 2, 127–137. [Google Scholar]
- Specht, E.; Miyake-Stoner, S.; Mayfield, S. Micro-algae come of age as a platform for recombinant protein production. Biotechnol. Lett. 2010, 32, 1373–1383. [Google Scholar] [CrossRef] [Green Version]
- Cerutti, H.; Ma, X.; Msanne, J.; Repas, T. RNA-mediated silencing in Algae: Biological roles and tools for analysis of gene function. Eukaryot. Cell 2011, 10, 1164–1172. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.-J.; Ma, X.; Cerutti, H. Gene silencing in microalgae: Mechanisms and biological roles. Bioresour. Technol. 2015, 184, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Neupert, J.; Karcher, D.; Bock, R. Generation of Chlamydomonas strains that efficiently express nuclear transgenes. Plant J. 2009, 57, 1140–1150. [Google Scholar] [CrossRef]
- Zaslavskaia, L.A.; Lippmeier, J.C.; Kroth, P.G.; Grossman, A.R.; Apt, K.E. Transformation of the diatom Phaeodactylum tricornutum (Bacillariophyceae) with a variety of selectable marker and reporter genes. J. Phycol. 2000, 36, 379–386. [Google Scholar] [CrossRef]
- Elmayan, T.; Vaucheret, H. Expression of single copies of a strongly expressed 35S transgene can be silenced post-transcriptionally. Plant J. 1996, 9, 787–797. [Google Scholar] [CrossRef]
- Liu, Q.; Feng, Y.; Zhu, Z. Dicer-like (DCL) proteins in plants. Funct. Integr. Genom. 2009, 9, 277–286. [Google Scholar] [CrossRef]
- Hutvagner, G.; Simard, M.J. Argonaute proteins: Key players in RNA silencing. Nat. Rev. Mol. Cell Biol. 2008, 9, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Jinkerson, R.E.; Jonikas, M.C. Molecular techniques to interrogate and edit the Chlamydomonas nuclear genome. Plant J. 2015, 82, 393–412. [Google Scholar] [CrossRef]
- Rasala, B.A.; Lee, P.A.; Shen, Z.; Briggs, S.P.; Mendez, M.; Mayfield, S.P. Robust expression and secretion of Xylanase1 in Chlamydomonas reinhardtii by fusion to a selection gene and processing with the FMDV 2A peptide. PLoS ONE 2012, 7, e43349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasala, B.A.; Barrera, D.J.; Ng, J.; Plucinak, T.M.; Rosenberg, J.N.; Weeks, D.P.; Oyler, G.A.; Peterson, T.C.; Haerizadeh, F.; Mayfield, S.P. Expanding the spectral palette of fluorescent proteins for the green microalga Chlamydomonas reinhardtii. Plant J. 2013, 74, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, M.L.L.; Luke, G.; Mehrotra, A.; Li, X.; Hughes, L.E.; Gani, D.; Ryan, M.D. Analysis of the aphthovirus 2A/2B polyprotein ‘cleavage’ mechanism indicates not a proteolytic reaction, but a novel translational effect: A putative ribosomal ‘skip’. J. Gen. Virol. 2001, 82, 1013–1025. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.D.; King, A.M.; Thomas, G.P. Cleavage of foot-and-mouth disease virus polyprotein is mediated by residues located within a 19 amino acid sequence. J. Gen. Virol. 1991, 72, 2727–2732. [Google Scholar] [CrossRef]
- Lumbreras, V.; Stevens, D.R.; Purton, S. Efficient foreign gene expression in Chlamydomonas reinhardtii mediated by an endogenous intron. Plant J. 1998, 14, 441–447. [Google Scholar] [CrossRef]
- Jakobiak, T.; Mages, W.; Scharf, B.; Babinger, P.; Stark, K.; Schmitt, R. The bacterial paromomycin resistance gene, aphH, as a dominant selectable marker in Volvox carteri. Protist 2004, 155, 381–393. [Google Scholar] [CrossRef]
- Fischer, N.; Rochaix, J.D. The flanking regions of PsaD drive efficient gene expression in the nucleus of the green alga Chlamydomonas reinhardtii. Mol. Genet. Genom. 2001, 265, 888–894. [Google Scholar] [CrossRef]
- Srinivasan, R.; Gothandam, K.M. Synergistic Action of D-Glucose and Acetosyringone on Agrobacterium Strains for Efficient Dunaliella Transformation. PLoS ONE 2016, 11, e0158322. [Google Scholar] [CrossRef]
- Kumar, S.; Raj, S.K.; Sharma, A.K.; Varma, H.N. Genetic transformation and development of Cucumber mosaic virus resistant transgenic plants of Chrysanthemum morifolium cv. Kundan. Sci. Hortic. 2012, 134, 40–45. [Google Scholar] [CrossRef]
- Naing, A.H.; Ai, T.N.; Jeon, S.M.; Lim, S.H.; Kim, C.K. An efficient protocol for Agrobacterium-mediated genetic transformation of recalcitrant chrysanthemum cultivar Shinma. Acta Physiol. Plant. 2016, 38. [Google Scholar] [CrossRef]
- Kumar, S.V.; Misquitta, R.W.; Reddy, V.S.; Rao, B.J.; Rajam, M.V. Genetic transformation of the green alga—Chlamydomonas reinhardtii by Agrobacterium tumefaciens. Plant Sci. 2004, 166, 731–738. [Google Scholar] [CrossRef]
- Hu, Z.; Wu, Y.-R.; Li, W.; Gao, H.-H. Factors affecting Agrobacterium tumefaciens-mediated genetic transformation of Lycium barbarum L. In Vitro Cell. Dev. Biol. Plant 2006, 42, 461–466. [Google Scholar] [CrossRef]
- Schlösser, U.G. SAG—Sammlung von Algenkulturen at the University of Göttingen Catalogue of Strains 1994. Bot. Acta 1994, 107, 113–186. [Google Scholar] [CrossRef]
- Checcucci, A.; Colombetti, G.; Ferrara, R.; Lenci, F. Action spectra for photoaccumulation of green and colorless Euglena: Evidence for identification of receptor pigments. Photochem. Photobiol. 1976, 23, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Höfgen, R.; Willmitzer, L. Storage of competent cells for Agrobacterium transformation. Nucleic Acids Res. 1988, 16, 9877. [Google Scholar] [CrossRef] [Green Version]
- Mermelstein, C.S.; Rodrigues, A.; Einicker-Lamas, M.; Navarrete, R.D.B.; Farina, M.; Costa, M.L. Distribution of F-actin, α-actinin, tropomyosin, tubulin and organelles in Euglena gracilis by immunofluorescence microscopy. Tissue Cell 1998, 30, 545–553. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Primer Name | Primer Sequence | Fragment Size (bp) |
|---|---|---|---|
| 1 | Gus-For | 5′GGGTACCCGGGTGGTCAGTC3′ | 1893 |
| Gus-Rev | 5′CGAGCTCGGTAGCAATTCCCG3′ | ||
| 2 | LHP5UTR-For | 5′GACCTGCAAGATCCCGGGcccagggaaacgtctctt3′ | 493 |
| LHP5UTR-Rev | 5′ctttcatccactcacttcaaaATGAAAAAGCCTgagc3′ | ||
| 3 | LHPINT-For | 5′ctttcatccactcacttcaaaATGAAAAAGCCTgagcacaactgccttcagacttg3′ | 1145 |
| LHPINT-Rev | 5′CTCGTCCGGGATCTTGGCGGGTGAAACAGACTTTG3′ | ||
| 4 | F2AGUS-For | 5′CTCGTCCGGGATCTTGGCGGGTGAAACAGACTTTGAATTTTGACCTTCTCAAGTTGGCGGGAGACGTGGAGTCCAACCCTGGACCTATGTTACGTCCTGTAGAAAC3′ | 1895 |
| F2AGUS-Rev | 5′GCAGCAGGGAGGCAAACAATGA3′ | ||
| 5 | LHP3UTR-For | 5′GGAGGCAAACAATGAtgatgtggacaacgcaact3′ | 410 |
| LHP3UTR-Rev | 5′agcaactacggctagaagtAGGCCTACTAGTGAGCTCG3′ | ||
| 6 | HptII-1839-For | 5′GACAGCGTCTCCGACCTG3′ | 495 |
| HptII-1840-Rev | 5′CCAAAGCATCAGCTCATCG3′ | ||
| 7 | HptII-1841-For | 5′GCGGTCATTGACTGGAGC3′ | 359 |
| HptII-1842-Rev | 5′CGTCGGTTTCCACTATCGG3′ | ||
| 8 | GFP-1847-For | 5′GACGTAAACGGCCACAAGTT3′ | 609 |
| GFP-1848-Rev | 5′GAACTCCAGCAGGACCATGT3′ | ||
| 9 | GUS-1851-For | 5′TGAAGATGCGGACTTACGTG3′ | 465 |
| GUS-1852-Rev | 5′TGAGCGTCGCAGAACATTAC3′ | ||
| 10 | GUS-1849-For | 5′CGACGCTCACACCGATAC3′ | 377 |
| GUS-1850-Rev | 5′GGTCGCGAGTGAAGATCC3′ | ||
| 11 | CaMV35S-1853-For | 5′CGCAGCAGGTCTCATCAAG3′ | 382 |
| CaMV35S-1854-Rev | 5′GACAAGTGTGTCGTGCTCC3′ | ||
| 12 | VirC-For | 5′ATCATTTGTAGCGACT3′ | 730 |
| VirC-Rev | 5′AGCTCAAACCTGCTTC3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becker, I.; Prasad, B.; Ntefidou, M.; Daiker, V.; Richter, P.; Lebert, M. Agrobacterium tumefaciens-Mediated Nuclear Transformation of a Biotechnologically Important Microalga—Euglena gracilis. Int. J. Mol. Sci. 2021, 22, 6299. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126299
Becker I, Prasad B, Ntefidou M, Daiker V, Richter P, Lebert M. Agrobacterium tumefaciens-Mediated Nuclear Transformation of a Biotechnologically Important Microalga—Euglena gracilis. International Journal of Molecular Sciences. 2021; 22(12):6299. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126299
Chicago/Turabian StyleBecker, Ina, Binod Prasad, Maria Ntefidou, Viktor Daiker, Peter Richter, and Michael Lebert. 2021. "Agrobacterium tumefaciens-Mediated Nuclear Transformation of a Biotechnologically Important Microalga—Euglena gracilis" International Journal of Molecular Sciences 22, no. 12: 6299. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126299