
Improvement of Glucose Tolerance by Food Factors Having Glucagon-Like Peptide-1 Releasing Activity

Abstract
:1. Introduction
2. Dietary Factors and GLP-1
2.1. Macronutrients
2.2. Micronutrients
2.3. Other Food Factors
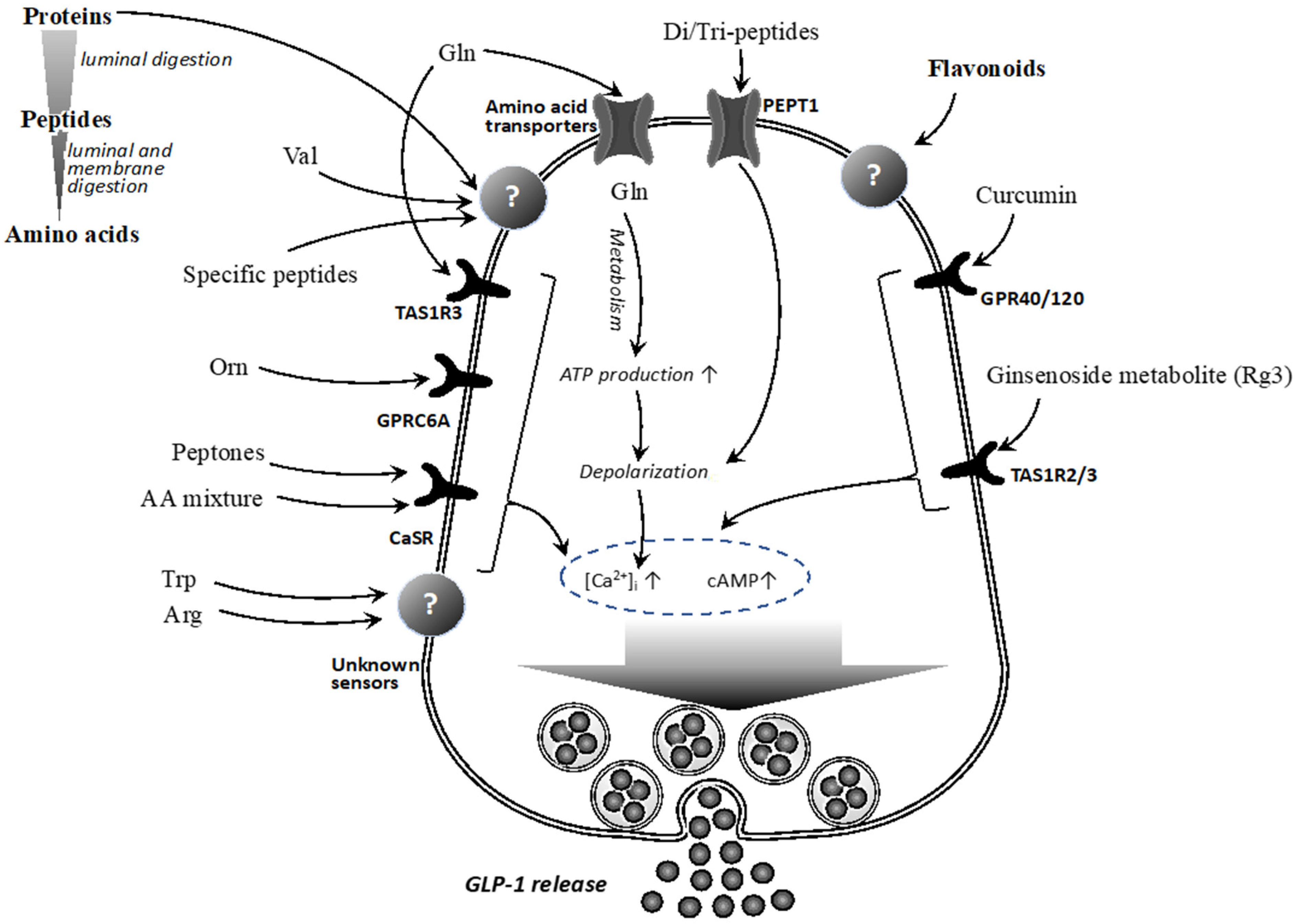
3. Dietary Proteins/Peptides/Amino Acids and GLP-1
3.1. Proteins
3.2. Peptides
3.3. Amino Acids
4. Phytochemicals and GLP-1
4.1. Anti-Diabetic Effects of Phytochemicals, and Related Mechanisms
4.2. Phytochemicals and GLP-1
5. Concluding Remarks
Funding
Conflicts of Interest
References
- Müller, T.; Finan, B.; Bloom, S.; D’Alessio, D.; Drucker, D.; Flatt, P.; Fritsche, A.; Gribble, F.; Grill, H.; Habener, J.; et al. Glucagon-like peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef]
- Meier, J.J.; Nauck, M.A.; Kranz, D.; Holst, J.J.; Deacon, C.F.; Gaeckler, D.; Schmidt, W.E.; Gallwitz, B. Secretion, degradation, and elimination of glucagon-like peptide 1 and gastric inhibitory polypeptide in patients with chronic renal insufficiency and healthy control subjects. Diabetes 2004, 53, 654–662. [Google Scholar] [CrossRef] [Green Version]
- Deacon, C.F. Physiology and Pharmacology of DPP-4 in Glucose Homeostasis and the Treatment of Type 2 Diabetes. Front. Endocrinol. 2019, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Holst, J.J. The incretin system in healthy humans: The role of GIP and GLP-1. Metabolism 2019, 96, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Nauck, M.A.; Quast, D.R.; Wefers, J.; Meier, J.J. GLP-1 receptor agonists in the treatment of type 2 diabetes-state-of-the-art. Mol. Metab. 2021, 46, 101102. [Google Scholar] [CrossRef]
- Buckley, S.T.; Bækdal, T.A.; Vegge, A.; Maarbjerg, S.J.; Pyke, C.; Ahnfelt-Rønne, J.; Madsen, K.G.; Schéele, S.G.; Alanentalo, T.; Kirk, R.K.; et al. Transcellular stomach absorption of a derivatized glucagon-like peptide-1 receptor agonist. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef]
- Deacon, C.F. Dipeptidyl peptidase 4 inhibitors in the treatment of type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2020, 16, 642–653. [Google Scholar] [CrossRef]
- Toft-Nielsen, M.B.; Damholt, M.B.; Madsbad, S.; Hilsted, L.M.; Hughes, T.E.; Michelsen, B.K.; Holst, J.J. Determinants of the Impaired Secretion of Glucagon-Like Peptide-1 in Type 2 Diabetic Patients. J. Clin. Endocrinol. Metab. 2001, 86, 3717–3723. [Google Scholar] [CrossRef]
- Muscelli, E.; Mari, A.; Casolaro, A.; Camastra, S.; Seghieri, G.; Gastaldelli, A.; Holst, J.J.; Ferrannini, E. Separate Impact of Obesity and Glucose Tolerance on the Incretin Effect in Normal Subjects and Type 2 Diabetic Patients. Diabetes 2007, 57, 1340–1348. [Google Scholar] [CrossRef] [Green Version]
- Nauck, M.; Vardarli, I.; Deacon, C.F.; Holst, J.J.; Meier, J.J. Secretion of glucagon-like peptide-1 (GLP-1) in type 2 diabetes: What is up, what is down? Diabetologia 2011, 54, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Meier, J.J.; Nauck, M.A. Is secretion of glucagon-like peptide-1 reduced in type 2 diabetes mellitus? Nat. Clin. Pract. Endocrinol. Metab. 2008, 4, 606–607. [Google Scholar] [CrossRef]
- Holst, J.J.; Knop, F.K.; Vilsbøll, T.; Krarup, T.; Madsbad, S. Loss of Incretin Effect Is a Specific, Important, and Early Characteristic of Type 2 Diabetes. Diabetes Care 2011, 34, S251–S257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calanna, S.; Christensen, M.; Holst, J.J.; Laferrère, B.; Gluud, L.L.; Vilsbøll, T.; Knop, F.K. Secretion of glucagon-like peptide-1 in patients with type 2 diabetes mellitus: Systematic review and meta-analyses of clinical studies. Diabetologia 2013, 56, 965–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faerch, K.; Torekov, S.; Vistisen, D.; Johansen, N.B.; Witte, D.R.; Jonsson, A.; Pedersen, O.; Hansen, T.; Lauritzen, T.; Sandbaek, A.; et al. GLP-1 Response to Oral Glucose Is Reduced in Prediabetes, Screen-Detected Type 2 Diabetes, and Obesity and Influenced by Sex: The ADDITION-PRO Study. Diabetes 2015, 64, 2513–2525. [Google Scholar] [CrossRef] [Green Version]
- Hira, T.; Pinyo, J.; Hara, H. What Is GLP-1 Really Doing in Obesity? Trends Endocrinol. Metab. 2020, 31, 71–80. [Google Scholar] [CrossRef]
- Nakajima, S.; Hira, T.; Hara, H. Postprandial glucagon-like peptide-1 secretion is increased during the progression of glucose intolerance and obesity in high-fat/high-sucrose diet-fed rats. Br. J. Nutr. 2015, 113, 1477–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hira, T.; Suto, R.; Kishimoto, Y.; Kanahori, S.; Hara, H. Resistant maltodextrin or fructooligosaccharides promotes GLP-1 production in male rats fed a high-fat and high-sucrose diet, and partially reduces energy intake and adiposity. Eur. J. Nutr. 2017, 57, 965–979. [Google Scholar] [CrossRef] [PubMed]
- Pinyo, J.; Hira, T.; Hara, H. Continuous feeding of a combined high-fat and high-sucrose diet, rather than an individual high-fat or high-sucrose diet, rapidly enhances the glucagon-like peptide-1 secretory response to meal ingestion in diet-induced obese rats. Nutrition 2019, 62, 122–130. [Google Scholar] [CrossRef]
- Pinyo, J.; Hara, H.; Hira, T. Diet-induced obesity enhances postprandial glucagon-like peptide-1 secretion in Wistar rats, but not in diabetic Goto-Kakizaki rats. Br. J. Nutr. 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pinyo, J.; Hira, T.; Hara, H. Enhanced postprandial glucagon-like peptide-1 secretion during obesity development has a protective role against glucose intolerance induction in rats. Br. J. Nutr. 2019, 122, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Svendsen, B.; Pedersen, J.; Albrechtsen, N.J.W.; Hartmann, B.; Toräng, S.; Rehfeld, J.F.; Poulsen, S.S.; Holst, J.J. An Analysis of Cosecretion and Coexpression of Gut Hormones from Male Rat Proximal and Distal Small Intestine. Endocrinology 2015, 156, 847–857. [Google Scholar] [CrossRef]
- Fothergill, L.J.; Furness, J.B. Diversity of enteroendocrine cells investigated at cellular and subcellular levels: The need for a new classification scheme. Histochem. Cell Biol. 2018, 150, 693–702. [Google Scholar] [CrossRef]
- Reimann, F.; Gribble, F.M. Mechanisms underlying glucose-dependent insulinotropic polypeptide and glucagon-like peptide-1 secretion. J. Diabetes Investig. 2016, 7, 13–19. [Google Scholar] [CrossRef]
- Kuhre, R.E.; Gribble, F.M.; Hartmann, B.; Reimann, F.; Windeløv, J.A.; Rehfeld, J.F.; Holst, J.J. Fructose stimulates GLP-1 but not GIP secretion in mice, rats, and humans. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G622–G630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, H.J.; Kokrashvili, Z.; Theodorakis, M.J.; Carlson, O.D.; Kim, B.-J.; Zhou, J.; Kim, H.H.; Xu, X.; Chan, S.L.; Juhaszova, M.; et al. Gut-expressed gustducin and taste receptors regulate secretion of glucagon-like peptide-1. Proc. Natl. Acad. Sci. USA 2007, 104, 15069–15074. [Google Scholar] [CrossRef] [Green Version]
- Margolskee, R.F.; Dyer, J.; Kokrashvili, Z.; Salmon, K.S.H.; Ilegems, E.; Daly, K.; Maillet, E.; Ninomiya, Y.; Mosinger, B.; Shirazi-Beechey, S.P. T1R3 and gustducin in gut sense sugars to regulate expression of Na+-glucose cotransporter 1. Proc. Natl. Acad. Sci. USA 2007, 104, 15075–15080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinert, R.E.; Gerspach, A.C.; Gutmann, H.; Asarian, L.; Drewe, J.; Beglinger, C. The functional involvement of gut-expressed sweet taste receptors in glucose-stimulated secretion of glucagon-like peptide-1 (GLP-1) and peptide YY (PYY). Clin. Nutr. 2011, 30, 524–532. [Google Scholar] [CrossRef]
- Feltrin, K.L.; Little, T.J.; Meyer, J.H.; Horowitz, M.; Smout, A.J.P.M.; Wishart, J.; Pilichiewicz, A.N.; Rades, T.; Chapman, I.M.; Feinle-Bisset, C. Effects of intraduodenal fatty acids on appetite, antropyloroduodenal motility, and plasma CCK and GLP-1 in humans vary with their chain length. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R524–R533. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, C.; Rasmussen, O.; Lousen, T.; Holst, J.J.; Fenselau, S.; Schrezenmeir, J.; Hermansen, K. Differential effects of saturated and monounsaturated fatty acids on postprandial lipemia and incretin responses in healthy subjects. Am. J. Clin. Nutr. 1999, 69, 1135–1143. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, C.; Storm, H.; Holst, J.J.; Hermansen, K. Differential effects of saturated and monounsaturated fats on postprandial lipemia and glucagon-like peptide 1 responses in patients with type 2 diabetes. Am. J. Clin. Nutr. 2003, 77, 605–611. [Google Scholar] [CrossRef]
- Hansen, K.B.; Rosenkilde, M.M.; Knop, F.K.; Wellner, N.; Diep, T.A.; Rehfeld, J.F.; Andersen, U.B.; Holst, J.J.; Hansen, H.S. 2-Oleoyl Glycerol Is a GPR119 Agonist and Signals GLP-1 Release in Humans. J. Clin. Endocrinol. Metab. 2011, 96, E1409–E1417. [Google Scholar] [CrossRef] [Green Version]
- Mandøe, M.J.; Hansen, K.B.; Hartmann, B.; Rehfeld, J.F.; Holst, J.J.; Hansen, H.S. The 2-monoacylglycerol moiety of dietary fat appears to be responsible for the fat-induced release of GLP-1 in humans. Am. J. Clin. Nutr. 2015, 102, 548–555. [Google Scholar] [CrossRef]
- Diakogiannaki, E.; Pais, R.; Tolhurst, G.; Parker, H.E.; Horscroft, J.; Rauscher, B.; Zietek, T.; Daniel, H.; Gribble, F.M.; Reimann, F. Oligopeptides stimulate glucagon-like peptide-1 secretion in mice through proton-coupled uptake and the calcium-sensing receptor. Diabetologia 2013, 56, 2688–2696. [Google Scholar] [CrossRef] [Green Version]
- Pais, R.; Gribble, F.M.; Reimann, F. Signalling pathways involved in the detection of peptones by murine small intestinal enteroendocrine L-cells. Peptides 2016, 77, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Mansour, A.; Hosseini, S.; Larijani, B.; Pajouhi, M.; Mohajeri-Tehrani, M.R. Nutrients related to GLP1 secretory responses. Nutrition 2013, 29, 813–820. [Google Scholar] [CrossRef]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-Chain Fatty Acids Stimulate Glucagon-Like Peptide-1 Secretion via the G-Protein-Coupled Receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Psichas, A.; Sleeth, M.L.; Murphy, K.; Brooks, L.; Bewick, G.; Hanyaloglu, A.C.; A Ghatei, M.; Bloom, S.R.; Frost, G. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int. J. Obes. 2015, 39, 424–429. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, C.B.; Gabe, M.B.N.; Svendsen, B.; Dragsted, L.O.; Rosenkilde, M.M.; Holst, J.J. The impact of short-chain fatty acids on GLP-1 and PYY secretion from the isolated perfused rat colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G53–G65. [Google Scholar] [CrossRef] [Green Version]
- Hira, T.; Ikee, A.; Kishimoto, Y.; Kanahori, S.; Hara, H. Resistant maltodextrin promotes fasting glucagon-like peptide-1 secretion and production together with glucose tolerance in rats. Br. J. Nutr. 2015, 114, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, M.; Hira, T.; Nakamura, M.; Iida, T.; Kishimoto, Y.; Hara, H. Secretion of GLP-1 but not GIP is potently stimulated by luminal d-Allulose (d-Psicose) in rats. Biochem. Biophys. Res. Commun. 2018. [Google Scholar] [CrossRef]
- Iwasaki, Y.; Sendo, M.; Dezaki, K.; Hira, T.; Sato, T.; Nakata, M.; Goswami, C.; Aoki, R.; Arai, T.; Kumari, P.; et al. GLP-1 release and vagal afferent activation mediate the beneficial metabolic and chronotherapeutic effects of D-allulose. Nat. Commun. 2018, 9, 113. [Google Scholar] [CrossRef]
- Miguéns-Gómez, A.; Casanova-Martí, À.; Blay, M.T.; Terra, X.; Beltrán-Debón, R.; Rodríguez-Gallego, E.; Ardévol, A.; Pinent, M. Glucagon-like peptide-1 regulation by food proteins and protein hydrolysates. Nutr. Res. Rev. 2021, 1–17. [Google Scholar] [CrossRef]
- Belza, A.; Ritz, C.; Sørensen, M.Q.; Holst, J.J.; Rehfeld, J.F.; Astrup, A. Contribution of gastroenteropancreatic appetite hormones to protein-induced satiety. Am. J. Clin. Nutr. 2013, 97, 980–989. [Google Scholar] [CrossRef]
- Van Der Klaauw, A.A.; Keogh, J.M.; Henning, E.; Trowse, V.M.; Dhillo, W.; Ghatei, M.A.; Farooqi, I.S. High protein intake stimulates postprandial GLP1 and PYY release. Obesity 2013, 21, 1602–1607. [Google Scholar] [CrossRef]
- Dougkas, A.; Östman, E. Protein-Enriched Liquid Preloads Varying in Macronutrient Content Modulate Appetite and Appetite-Regulating Hormones in Healthy Adults. J. Nutr. 2016, 146, 637–645. [Google Scholar] [CrossRef] [Green Version]
- Ohlsson, L.; Alsalim, W.; Carr, R.D.; Tura, A.; Pacini, G.; Mari, A.; Ahrén, B. Glucose-lowering effect of the DPP-4 inhibitor sitagliptin after glucose and non-glucose macronutrient ingestion in non-diabetic subjects. Diabetes Obes. Metab. 2013, 15, 531–537. [Google Scholar] [CrossRef]
- Geraedts, M.C.; Troost, F.J.; Fischer, M.A.; Edens, L.; Saris, W.H. Direct induction of CCK and GLP-1 release from murine endocrine cells by intact dietary proteins. Mol. Nutr. Food Res. 2010, 55, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, A.L.; Calderwood, D.; Hobson, L.; Green, B.D. Whey proteins have beneficial effects on intestinal enteroendocrine cells stimulating cell growth and increasing the production and secretion of incretin hormones. Food Chem. 2015, 189, 120–128. [Google Scholar] [CrossRef]
- Gillespie, A.L.; Green, B.D. The bioactive effects of casein proteins on enteroendocrine cell health, proliferation and incretin hormone secretion. Food Chem. 2016, 211, 148–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, Y.; Wada, Y.; Izumi, H.; Shimizu, T.; Takeda, Y.; Hira, T.; Hara, H. Casein materials show different digestion patterns using an In Vitro gastrointestinal model and different release of glucagon-like peptide-1 by enteroendocrine GLUTag cells. Food Chem. 2019, 277, 423–431. [Google Scholar] [CrossRef]
- Ryan, A.T.; Feinle-Bisset, C.; Kallas, A.; Wishart, J.M.; Clifton, P.; Horowitz, M.; Luscombe-Marsh, N.D. Intraduodenal protein modulates antropyloroduodenal motility, hormone release, glycemia, appetite, and energy intake in lean men. Am. J. Clin. Nutr. 2012, 96, 474–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchison, A.T.; Feinle-Bisset, C.; Fitzgerald, P.C.; Standfield, S.; Horowitz, M.; Clifton, P.M.; Luscombe-Marsh, N.D. Comparative effects of intraduodenal whey protein hydrolysate on antropyloroduodenal motility, gut hormones, glycemia, appetite, and energy intake in lean and obese men. Am. J. Clin. Nutr. 2015, 102, 1323–1331. [Google Scholar] [CrossRef]
- Nobile, V.; Duclous, E.; Michelotti, A.; Bizzaro, G.; Negro, M.; Soisson, F. Supplementation with a fish protein hydrolysate (Micromesistius poutassou): Effects on body weight, body composition, and CCK/GLP-1 secretion. Food Nutr. Res. 2016, 60, 29857. [Google Scholar] [CrossRef] [Green Version]
- Jensen, C.; Dale, H.F.; Hausken, T.; Lied, E.; Hatlebakk, J.G.; Brønstad, I.; Lied, G.A.; Hoff, D.A.L. Supplementation with cod protein hydrolysate in older adults: A dose range cross-over study. J. Nutr. Sci. 2019, 8, e40. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, N.; Hira, T.; Yamada, N.; Hara, H. Oral Administration of Corn Zein Hydrolysate Stimulates GLP-1 and GIP Secretion and Improves Glucose Tolerance in Male Normal Rats and Goto-Kakizaki Rats. Endocrinology 2013, 154, 3089–3098. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, Y.; Hira, T.; Inoue, D.; Harada, Y.; Hashimoto, H.; Fujii, M.; Kadowaki, M.; Hara, H. Rice protein hydrolysates stimulate GLP-1 secretion, reduce GLP-1 degradation, and lower the glycemic response in rats. Food Funct. 2015, 6, 2525–2534. [Google Scholar] [CrossRef]
- De Cicco, M.; Mamone, G.; Di Stasio, L.; Ferranti, P.; Addeo, F.; Picariello, G. Hidden “Digestome”: Current Analytical Approaches Provide Incomplete Peptide Inventories of Food Digests. J. Agric. Food Chem. 2019, 67, 7775–7782. [Google Scholar] [CrossRef]
- Theysgeur, S.; Cudennec, B.; Deracinois, B.; Perrin, C.; Guiller, I.; Lepoudère, A.; Flahaut, C.; Ravallec, R. New Bioactive Peptides Identified from a Tilapia Byproduct Hydrolysate Exerting Effects on DPP-IV Activity and Intestinal Hormones Regulation After Canine Gastrointestinal Simulated Digestion. Molecules 2020, 26, 136. [Google Scholar] [CrossRef]
- Le Nevé, B.; Daniel, H. Selected tetrapeptides lead to a GLP-1 release from the human enteroendocrine cell line NCI-H716. Regul. Pept. 2011, 167, 14–20. [Google Scholar] [CrossRef]
- Santos-Hernández, M.; Amigo, L.; Recio, I. Induction of CCK and GLP-1 release in enteroendocrine cells by egg white peptides generated during gastrointestinal digestion. Food Chem. 2020, 329, 127188. [Google Scholar] [CrossRef]
- Chandra, R.; Liddle, R.A. Recent advances in pancreatic endocrine and exocrine secretion. Curr. Opin. Gastroenterol. 2011, 27, 439–443. [Google Scholar] [CrossRef] [Green Version]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static In Vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Modvig, I.M.; Kuhre, R.E.; Holst, J.J. Peptone-mediated glucagon-like peptide-1 secretion depends on intestinal absorption and activation of basolaterally located Calcium-Sensing Receptors. Physiol. Rep. 2019, 7, e14056. [Google Scholar] [CrossRef]
- Proença, C.; Freitas, M.; Ribeiro, D.; Tomé, S.M.; Araújo, A.N.; Silva, A.M.S.; Fernandes, P.A.; Fernandes, E. The dipeptidyl peptidase-4 inhibitory effect of flavonoids is hindered in protein rich environments. Food Funct. 2019, 10, 5718–5731. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Fitzgerald, R.J. Features of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides from dietary proteins. J. Food Biochem. 2017, 43, e12451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, S.P.; Goswami, A.; Kalia, K.; Kate, A.S. Plant-Derived Bioactive Peptides: A Treatment to Cure Diabetes. Int. J. Pept. Res. Ther. 2020, 26, 955–968. [Google Scholar] [CrossRef]
- Mochida, T.; Hira, T.; Hara, H. The Corn Protein, Zein Hydrolysate, Administered into the Ileum Attenuates Hyperglycemia via Its Dual Action on Glucagon-Like Peptide-1 Secretion and Dipeptidyl Peptidase-IV Activity in Rats. Endocrinology 2010, 151, 3095–3104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, Y.; Hara, H.; Hira, T. Glucagon-like peptide-1 response to whey protein is less diminished by dipeptidyl peptidase-4 in comparison with responses to dextrin, a lipid and casein in rats. Br. J. Nutr. 2021, 125, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Gunnarsson, P.T.; Winzell, M.S.; Deacon, C.F.; Larsen, M.O.; Jelic, K.; Carr, R.D.; Ahrén, B. Glucose-Induced Incretin Hormone Release and Inactivation Are Differently Modulated by Oral Fat and Protein in Mice. Endocrinology 2006, 147, 3173–3180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.; Davies, K.A.B.; Stevenson, E.J.; West, D.J. The Clinical Application of Mealtime Whey Protein for the Treatment of Postprandial Hyperglycaemia for People With Type 2 Diabetes: A Long Whey to Go. Front. Nutr. 2020, 7, 587843. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.L.; Broughton, K.S. Insulinotropic Effects of Whey: Mechanisms of Action, Recent Clinical Trials, and Clinical Applications. Ann. Nutr. Metab. 2016, 69, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Caron, J.; Cudennec, B.; Domenger, D.; Belguesmia, Y.; Flahaut, C.; Kouach, M.; Lesage, J.; Goossens, J.F.; Dhulster, P.; Ravallec, R. Simulated GI digestion of dietary protein: Release of new bioactive peptides involved in gut hormone secretion. Food Res. Int. 2016, 89, 382–390. [Google Scholar] [CrossRef]
- Reimann, F.; Williams, L.; da Silva Xavier, G.D.S.; Rutter, G.; Gribble, F.M. Glutamine potently stimulates glucagon-like peptide-1 secretion from GLUTag cells. Diabetologia 2004, 47, 1592–1601. [Google Scholar] [CrossRef] [Green Version]
- Tolhurst, G.; Zheng, Y.; Parker, H.E.; Habib, A.M.; Reimann, F.; Gribble, F.M. Glutamine Triggers and Potentiates Glucagon-Like Peptide-1 Secretion by Raising Cytosolic Ca2+ and cAMP. Endocrinology 2011, 152, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Harada, K.; Kamiya, T.; Takizawa, M.; Küppers, J.; Nakajima, K.; Gütschow, M.; Kitaguchi, T.; Ohta, K.; Kato, T.; et al. Glutamine-induced signaling pathways via amino acid receptors in enteroendocrine L cell lines. J. Mol. Endocrinol. 2020, 64, 133–143. [Google Scholar] [CrossRef]
- Andersson, L.E.; Shcherbina, L.; Al-Majdoub, M.; Vishnu, N.; Arroyo, C.B.B.; Carrara, J.A.; Wollheim, C.B.; Fex, M.; Mulder, H.; Wierup, N.; et al. Glutamine-Elicited Secretion of Glucagon-Like Peptide 1 Is Governed by an Activated Glutamate Dehydrogenase. Diabetes 2017, 67, 372–384. [Google Scholar] [CrossRef] [Green Version]
- Oya, M.; Kitaguchi, T.; Pais, R.; Reimann, F.; Gribble, F.; Tsuboi, T. The G Protein-coupled Receptor Family C Group 6 Subtype A (GPRC6A) Receptor Is Involved in Amino Acid-induced Glucagon-like Peptide-1 Secretion from GLUTag Cells. J. Biol. Chem. 2013, 288, 4513–4521. [Google Scholar] [CrossRef] [Green Version]
- Clemmensen, C.; Jørgensen, C.V.; Smajilovic, S.; Bräuner-Osborne, H. Robust GLP-1 secretion by basic L-amino acids does not require the GPRC6A receptor. Diabetes Obes. Metab. 2017, 19, 599–603. [Google Scholar] [CrossRef]
- Modvig, I.M.; Kuhre, R.E.; Jepsen, S.L.; Xu, S.F.S.; Engelstoft, M.S.; Egerod, K.L.; Schwartz, T.W.; Ørskov, C.; Rosenkilde, M.M.; Holst, J.J. Amino acids differ in their capacity to stimulate GLP-1 release from the perfused rat small intestine and stimulate secretion by different sensing mechanisms. Am. J. Physiol. Endocrinol. Metab. 2021. [Google Scholar] [CrossRef]
- Greenfield, J.R.; Farooqi, I.S.; Keogh, J.M.; Henning, E.; Habib, A.M.; Blackwood, A.; Reimann, F.; Holst, J.J.; Gribble, F.M. Oral glutamine increases circulating glucagon-like peptide 1, glucagon, and insulin concentrations in lean, obese, and type 2 diabetic subjects. Am. J. Clin. Nutr. 2008, 89, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Samocha-Bonet, D.; Wong, O.; Synnott, E.-L.; Piyaratna, N.; Douglas, A.; Gribble, F.M.; Holst, J.J.; Chisholm, D.J.; Greenfield, J.R. Glutamine Reduces Postprandial Glycemia and Augments the Glucagon-Like Peptide-1 Response in Type 2 Diabetes Patients. J. Nutr. 2011, 141, 1233–1238. [Google Scholar] [CrossRef]
- Meek, C.L.; Lewis, H.B.; Vergese, B.; Park, A.; Reimann, F.; Gribble, F. The effect of encapsulated glutamine on gut peptide secretion in human volunteers. Peptides 2016, 77, 38–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotoudeh, G.; Abshirini, M.; Mahaki, B.; Bagheri, F.; Siassi, F.; Koohdani, F. Higher intake of phytochemical-rich foods is inversely related to prediabetes: A case-control study. Int. J. Prev. Med. 2018, 9, 64. [Google Scholar] [CrossRef]
- Eslami, O.; Khoshgoo, M.; Shidfar, F. Dietary phytochemical index and overweight/obesity in children: A cross-sectional study. BMC Res. Notes 2020, 13, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Park, K. Association between phytochemical index and metabolic syndrome. Nutr. Res. Pract. 2020, 14, 252–261. [Google Scholar] [CrossRef]
- Mozaffarian, D. Dietary and policy priorities to reduce the global crises of obesity and diabetes. Nat. Food 2020, 1, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Hameed, A.; Galli, M.; Adamska-Patruno, E.; Krętowski, A.; Ciborowski, M. Select Polyphenol-Rich Berry Consumption to Defer or Deter Diabetes and Diabetes-Related Complications. Nutrients 2020, 12, 2538. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Zhang, W.; Zhen, W.; Lum, H.; Nadler, J.L.; Bassaganya-Riera, J.; Jia, Z.; Wang, Y.; Misra, H.; Liu, D. Genistein Induces Pancreatic β-Cell Proliferation through Activation of Multiple Signaling Pathways and Prevents Insulin-Deficient Diabetes in Mice. Endocrinology 2010, 151, 3026–3037. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Zhen, W.; Yang, Z.; Carter, J.D.; Si, H.; Reynolds, K.A. Genistein Acutely Stimulates Insulin Secretion in Pancreatic α-Cells Through a cAMP-Dependent Protein Kinase Pathway. Diabetes 2006, 55, 1043–1050. [Google Scholar] [CrossRef] [Green Version]
- Braxas, H.; Rafraf, M.; Hasanabad, S.K.; Jafarabadi, M.A. Effectiveness of Genistein Supplementation on Metabolic Factors and Antioxidant Status in Postmenopausal Women with Type 2 Diabetes Mellitus. Can. J. Diabetes. 2019, 43, 490–497. [Google Scholar] [CrossRef]
- Alam, M.; Meerza, D.; Naseem, I. Protective effect of quercetin on hyperglycemia, oxidative stress and DNA damage in alloxan induced type 2 diabetic mice. Life Sci. 2014, 109, 8–14. [Google Scholar] [CrossRef]
- Eitah, H.E.; Maklad, Y.A.; Abdelkader, N.F.; el Din, A.A.G.; Badawi, M.; Kenawy, S.A. Modulating impacts of quercetin/sitagliptin combination on streptozotocin-induced diabetes mellitus in rats. Toxicol. Appl. Pharmacol. 2019, 365, 30–40. [Google Scholar] [CrossRef]
- Cremonini, E.; Fraga, C.G.; Oteiza, P.I. (–)-Epicatechin in the control of glucose homeostasis: Involvement of redox-regulated mechanisms. Free Radic. Biol. Med. 2019, 130, 478–488. [Google Scholar] [CrossRef]
- Tian, R.; Yang, W.; Xue, Q.; Gao, L.; Huo, J.; Ren, D.; Chen, X. Rutin ameliorates diabetic neuropathy by lowering plasma glucose and decreasing oxidative stress via Nrf2 signaling pathway in rats. Eur. J. Pharmacol. 2016, 771, 84–92. [Google Scholar] [CrossRef]
- Prasath, G.S.; Sundaram, S.; Subramanian, S.P. Fisetin averts oxidative stress in pancreatic tissues of streptozotocin-induced diabetic rats. Endocrine 2013, 44, 359–368. [Google Scholar] [CrossRef]
- Bashir, N.; Manoharan, V.; Miltonprabu, S. Grape seed proanthocyanidins protects against cadmium induced oxidative pancreatitis in rats by attenuating oxidative stress, inflammation and apoptosis via Nrf-2/HO-1 signaling. J. Nutr. Biochem. 2016, 32, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Rajappa, R.; Sireesh, D.; Salai, M.B.; Ramkumar, K.M.; Sarvajayakesavulu, S.; Madhunapantula, S.V. Treatment With Naringenin Elevates the Activity of Transcription Factor Nrf2 to Protect Pancreatic β-Cells From Streptozotocin-Induced Diabetes In Vitro and In Vivo. Front. Pharmacol. 2019, 9, 1562. [Google Scholar] [CrossRef] [Green Version]
- Goto, T.; Horita, M.; Nagai, H.; Nagatomo, A.; Nishida, N.; Matsuura, Y.; Nagaoka, S. Tiliroside, a glycosidic flavonoid, inhibits carbohydrate digestion and glucose absorption in the gastrointestinal tract. Mol. Nutr. Food Res. 2011, 56, 435–445. [Google Scholar] [CrossRef]
- Ojelabi, O.A.; Lloyd, K.P.; De Zutter, J.K.; Carruthers, A. Red wine and green tea flavonoids are cis-allosteric activators and competitive inhibitors of glucose transporter 1 (GLUT1)-mediated sugar uptake. J. Biol. Chem. 2018, 293, 19823–19834. [Google Scholar] [CrossRef] [Green Version]
- Zygmunt, K.; Faubert, B.; MacNeil, J.; Tsiani, E. Naringenin, a citrus flavonoid, increases muscle cell glucose uptake via AMPK. Biochem. Biophys. Res. Commun. 2010, 398, 178–183. [Google Scholar] [CrossRef]
- Kim, M.S.; Hur, H.J.; Kwon, D.Y.; Hwang, J.-T. Tangeretin stimulates glucose uptake via regulation of AMPK signaling pathways in C2C12 myotubes and improves glucose tolerance in high-fat diet-induced obese mice. Mol. Cell. Endocrinol. 2012, 358, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Go, G.-W.; Imm, J.-Y. Promotion of Glucose Uptake in C2C12 Myotubes by Cereal Flavone Tricin and Its Underlying Molecular Mechanism. J. Agric. Food Chem. 2017, 65, 3819–3826. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Yamashita, Y.; Nakamura, A.; Croft, K.; Ashida, H. Quercetin and its metabolite isorhamnetin promote glucose uptake through different signalling pathways in myotubes. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Prasath, G.S.; Pillai, S.I.; Subramanian, S.P. Fisetin improves glucose homeostasis through the inhibition of gluconeogenic enzymes in hepatic tissues of streptozotocin induced diabetic rats. Eur. J. Pharmacol. 2014, 740, 248–254. [Google Scholar] [CrossRef]
- Prince, P.S.M.; Kamalakkannan, N. Rutin improves glucose homeostasis in streptozotocin diabetic tissues by altering glycolytic and gluconeogenic enzymes. J. Biochem. Mol. Toxicol. 2006, 20, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Alkhalidy, H.; Moore, W.; Wang, A.; Luo, J.; McMillan, R.P.; Wang, Y.; Zhen, W.; Hulver, M.W.; Liu, D. Kaempferol ameliorates hyperglycemia through suppressing hepatic gluconeogenesis and enhancing hepatic insulin sensitivity in diet-induced obese mice. J. Nutr. Biochem. 2018, 58, 90–101. [Google Scholar] [CrossRef]
- Sundaram, R.; Nandhakumar, E.; Banu, H.H. Hesperidin, a citrus flavonoid ameliorates hyperglycemia by regulating key enzymes of carbohydrate metabolism in streptozotocin-induced diabetic rats. Toxicol. Mech. Methods 2019, 29, 644–653. [Google Scholar] [CrossRef]
- Takikawa, M.; Kurimoto, Y.; Tsuda, T. Curcumin stimulates glucagon-like peptide-1 secretion in GLUTag cells via Ca2+/calmodulin-dependent kinase II activation. Biochem. Biophys. Res. Commun. 2013, 435, 165–170. [Google Scholar] [CrossRef]
- Kato, M.; Tani, T.; Terahara, N.; Tsuda, T. The Anthocyanin Delphinidin 3-Rutinoside Stimulates Glucagon-Like Peptide-1 Secretion in Murine GLUTag Cell Line via the Ca2+/Calmodulin-Dependent Kinase II Pathway. PLoS ONE 2015, 10, e0126157. [Google Scholar] [CrossRef]
- Kim, K.-S.; Yang, H.J.; Lee, I.-S.; Kim, K.-H.; Park, J.; Jeong, H.-S.; Kim, Y.; Ahn, K.S.; Na, Y.-C.; Jang, H.-J. The aglycone of ginsenoside Rg3 enables glucagon-like peptide-1 secretion in enteroendocrine cells and alleviates hyperglycemia in type 2 diabetic mice. Sci. Rep. 2015, 5, 18325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, A.; AlKhalidy, H.; Luo, J.; Moomaw, E.; Neilson, A.P.; Liu, D. Flavone Hispidulin Stimulates Glucagon-Like Peptide-1 Secretion and Ameliorates Hyperglycemia in Streptozotocin-Induced Diabetic Mice. Mol. Nutr. Food Res. 2020, 64, e1900978. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, S.-T.; Yin, Y.-C.; Xing, S.; Li, W.-N.; Fu, X.-Q. Hypoglycemic effect and mechanism of isoquercitrin as an inhibitor of dipeptidyl peptidase-4 in type 2 diabetic mice. RSC Adv. 2018, 8, 14967–14974. [Google Scholar] [CrossRef] [Green Version]
- Song, W.-Y.; Aihara, Y.; Hashimoto, T.; Kanazawa, K.; Mizuno, M. (−)-Epigallocatechin-3-gallate induces secretion of anorexigenic gut hormones. J. Clin. Biochem. Nutr. 2015, 57, 164–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalivarathan, J.; Kalaivanan, K.; Chandrasekaran, S.P.; Nanda, D.; Ramachandran, V.; Venkatraman, A.C. Apigenin modulates hippocampal CREB-BDNF signaling in high fat, high fructose diet-fed rats. J. Funct. Foods 2020, 68, 103898. [Google Scholar] [CrossRef]
- Kato, M.; Nishikawa, S.; Ikehata, A.; Dochi, K.; Tani, T.; Takahashi, T.; Imaizumi, A.; Tsuda, T. Curcumin improves glucose tolerance via stimulation of glucagon-like peptide-1 secretion. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Rehman, K.; Ali, M.B.; Akash, M.S.H. Genistein enhances the secretion of glucagon-like peptide-1 (GLP-1) via downregulation of inflammatory responses. Biomed. Pharmacother. 2019, 112, 108670. [Google Scholar] [CrossRef]
- Kwon, E.-Y.; Choi, M.-S. Luteolin Targets the Toll-Like Receptor Signaling Pathway in Prevention of Hepatic and Adipocyte Fibrosis and Insulin Resistance in Diet-Induced Obese Mice. Nutrients 2018, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalitha, N.; Sadashivaiah, B.; Ramaprasad, T.R.; Singh, S.A. Anti-hyperglycemic activity of myricetin, through inhibition of DPP-4 and enhanced GLP-1 levels, is attenuated by co-ingestion with lectin-rich protein. PLoS ONE 2020, 15, e0231543. [Google Scholar] [CrossRef] [Green Version]
- Casanova-Martí, À.; Serrano, J.; Blay, M.T.; Terra, X.; Ardévol, A.; Pinent, M. Acute selective bioactivity of grape seed proanthocyanidins on enteroendocrine secretions in the gastrointestinal tract. Food Nutr. Res. 2017, 61, 1321347. [Google Scholar] [CrossRef] [Green Version]
- González-Abuín, N.; Martínez-Micaelo, N.; Blay, M.; Ardévol, A.; Pinent, M. Grape-Seed Procyanidins Prevent the Cafeteria-Diet-Induced Decrease of Glucagon-Like Peptide-1 Production. J. Agric. Food Chem. 2014, 62, 1066–1072. [Google Scholar] [CrossRef]
- Dao, T.M.; Waget, A.; Klopp, P.; Serino, M.; Vachoux, C.; Pechere, L.; Drucker, D.J.; Champion, S.; Barthélemy, S.; Barra, Y.; et al. Resveratrol Increases Glucose Induced GLP-1 Secretion in Mice: A Mechanism which Contributes to the Glycemic Control. PLoS ONE 2011, 6, e20700. [Google Scholar] [CrossRef]
- Lee, J.E.; Mitchell, A.E. Pharmacokinetics of Quercetin Absorption from Apples and Onions in Healthy Humans. J. Agric. Food Chem. 2012, 60, 3874–3881. [Google Scholar] [CrossRef]
- Trakooncharoenvit, A.; Tanaka, S.; Mizuta, E.; Hira, T.; Hara, H. Water-soluble dietary fibers enhance bioavailability of quercetin and a fiber derived from soybean is most effective after long-term feeding in rats. Eur. J. Nutr. 2020, 59, 1389–1398. [Google Scholar] [CrossRef]
- Trakooncharoenvit, A.; Hara, H.; Hira, T. Combination of α-Glycosyl-Isoquercitrin and Soybean Fiber Promotes Quercetin Bioavailability and Glucagon-like Peptide-1 Secretion and Improves Glucose Homeostasis in Rats Fed a High-Fat High-Sucrose Diet. J. Agric. Food Chem. 2021, 69, 5907–5916. [Google Scholar] [CrossRef]


{kind=link}
| Peptide Sequence | Models | Concentration Used (mM) | Source | Reference |
|---|---|---|---|---|
| TKAVEH | STC-1 | 0.1 | bovine hemoglobin | Caron, J. Food Res Int 2016 [72] |
| ANVST | STC-1 | 1 | bovine hemoglobin | Caron, J. Food Res Int 2016 [72] |
| KAAVT | STC-1 | 1 | bovine hemoglobin | Caron, J. Food Res Int 2016 [72] |
| YGAE | STC-1 | 1 | bovine hemoglobin | Caron, J. Food Res Int 2016 [72] |
| GPVRGPFPIIV | GLUTag | 5 | β-casein | Komatsu, Y. Food Chem 2019 [50] |
| PFL | STC-1 | 2 | egg white protein | Santos-Hernández, M. Food Chem 2020 [60] |
| RVASMASEKM | STC-1 | 2 | egg white protein | Santos-Hernández, M. Food Chem 2020 [60] |
| LKPT | STC-1 | 1 | tilapia byproduct | Theysgeur, S. Molecules 2020 [58] |
| GGGG | NCl-H716 | 20 | synthetic | Le Nevé, B. Regul Pept 2011 [59] |
| AAAA | NCl-H716 | 10 | synthetic | Le Nevé, B. Regul Pept 2011 [59] |
| GWGG | NCl-H716 | 10 | synthetic | Le Nevé, B. Regul Pept 2011 [59] |
| Glycylsarcosine | Murine colon primary culture | 10 | synthetic | Diakogiannaki, E. Diabetologia 2013 [33] |
| LGG | Murine colon primary culture | 10 | synthetic | Diakogiannaki, E. Diabetologia 2013 [33] |
| GF | Murine colon primary culture | 10 | synthetic | Diakogiannaki, E. Diabetologia 2013 [33] |
| Compound | Models | Treatment | Effect | Reference |
|---|---|---|---|---|
| Apigenin | High fat-high fructose diet-fed rats | 1.5 mg/kg BW, intraperitoneal, 30 days | Plasma GLP-1↑ | Kalivarathan, J. J Func Foods 2020 [115] |
| Curcumin | GLUTag cells | 10–50 µM, 2 h | GLP-1 secretion↑ | Takikawa, M. Biochem Biophys Res Commun 2013 [109] |
| Curcumin | Rats | 1.5 mg/kg BW, oral | Plasma GLP-1↑ Glucose tolerance↑ | Kato, M. Mol Nutr Food Res 2017 [116] |
| Delphinidin 3-rutinoside | GLUTag Cells | 10–100 µM, 2 h | GLP-1 secretion↑ | Kato, M. PLoS One 2015 [110] |
| Epigallocatechin-3-gallate | Caco-2 cells Murine ileal tissue | 300 µM, 2 h 1 mM, 45 min | GLP-1 secretion↑ GLP-1 secretion↑ | Song, WY. J Clin Biochem Nutr 2015 [114] |
| Genistein (combined with metformin) | Alloxan-induced diabetic rats | 20 mg/kg BW/day, intraperitoneal, 30 days | Serum and intestinal GLP-1↑ Glucose tolerance↑ | Rehman, K. Biomed Pharmacother 2019 [117] |
| Ginsenoside metabolite, Rg3 | NCI-H716 cells db/db mice | 1–25 µM, 1 h 0.5 mg/kg BW, oral | GLP-1 secretion↑ Plasma GLP-1↑ Glucose tolerance↑ | Kim, KS. Sci Rep. 2015 [111] |
| Hispidulin | GLUTag cells Mouse ileum crypts STZ-treated mice | 1–50 µM, 1 h 1–50 µM, 1 h 20 mg/kg BW/day, oral, 6 weeks | GLP-1 secretion↑ GLP-1 secretion↑ Plasma GLP-1↑ Glucose tolerance↑ | Wang, Y. Mol Nutr Food Res 2020 [112] |
| Isoquercitrin | NCI-H716 cells HFD-fed, STZ-treated mice | 10–100 µM, 90 min 20–80 mg/kgBW/day, oral, 8 weeks | GLP-1 secretion↑ Plasma GLP-1↑ Plasma DPP-4↓ Plasma Glucose↓ | Zang, L. RSC Adv 2018 [113] |
| Luteolin | HFD-fed mice | Supplemented in diet, 0.005%, 16 weeks | Plasma GLP-1↑ Glucose tolerance↑ | Kwon, EY. Nutrients 2018 [118] |
| Myricetin | HFD-fed, STZ-treated rats | 20 mg/kg BW/day, oral, 4 weeks | Plasma GLP-1↑ Plasma and tissue DPP-4↓ | Lalitha, N. PLoS One 2020 [119] |
| Proanthocyanidin Gallic acid | Rat ileal segment | 0.2 mg/mL, 1 h 31 µg/mL, 1 h | GLP-1 secretion↑ GLP-1 secretion↑ | Casanova-Martí, À. Food Nutr Res 2017 [120] |
| Procyanidin | Cafeteria diet-fed rats | 25 mg/kg BW/day, oral, 12 weeks | Intestinal GLP-1↑ | González-Abuín, N. J Agric Food Chem 2014 [121] |
| Resveratrol | HFD-fed mice | Supplemented in diet, 60 mg/kg BW/day, 5 weeks | Plasma GLP-1↑ Glucose tolerance↑ | Dao, TM. PLoS One 2011 [122] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hira, T.; Trakooncharoenvit, A.; Taguchi, H.; Hara, H. Improvement of Glucose Tolerance by Food Factors Having Glucagon-Like Peptide-1 Releasing Activity. Int. J. Mol. Sci. 2021, 22, 6623. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126623
Hira T, Trakooncharoenvit A, Taguchi H, Hara H. Improvement of Glucose Tolerance by Food Factors Having Glucagon-Like Peptide-1 Releasing Activity. International Journal of Molecular Sciences. 2021; 22(12):6623. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126623
Chicago/Turabian StyleHira, Tohru, Aphichat Trakooncharoenvit, Hayate Taguchi, and Hiroshi Hara. 2021. "Improvement of Glucose Tolerance by Food Factors Having Glucagon-Like Peptide-1 Releasing Activity" International Journal of Molecular Sciences 22, no. 12: 6623. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126623