
Preconceptional Immunization Can Modulate Offspring Intrathymic IL-17-Producing γδT Cells with Epigenetic Implications Mediated by microRNAs


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
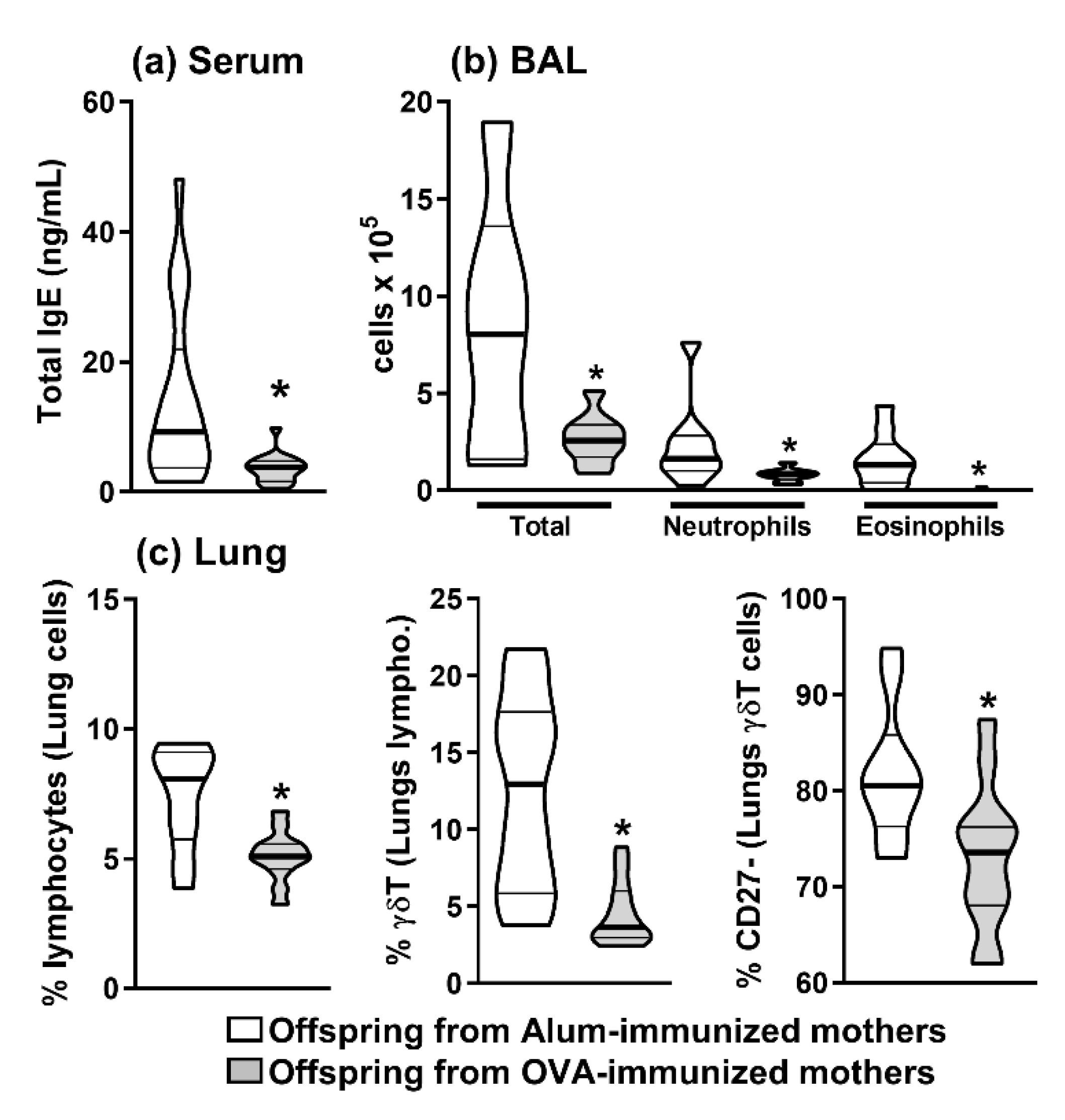
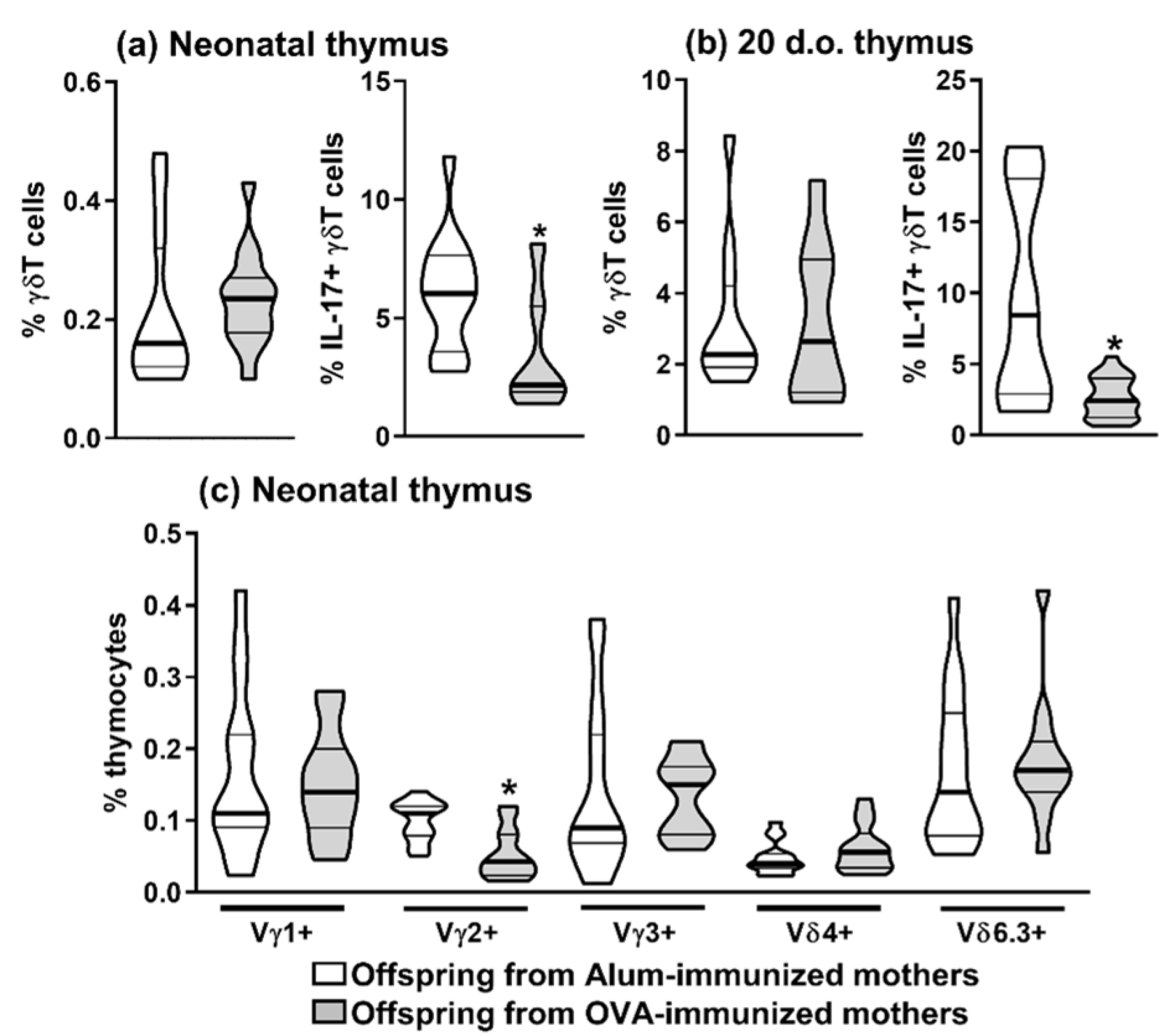
2.1. Maternal Immunization Could Inhibit Offspring Allergy and Reduce the Frequency of Offspring Thymic IL-17-Producing γδT Cells
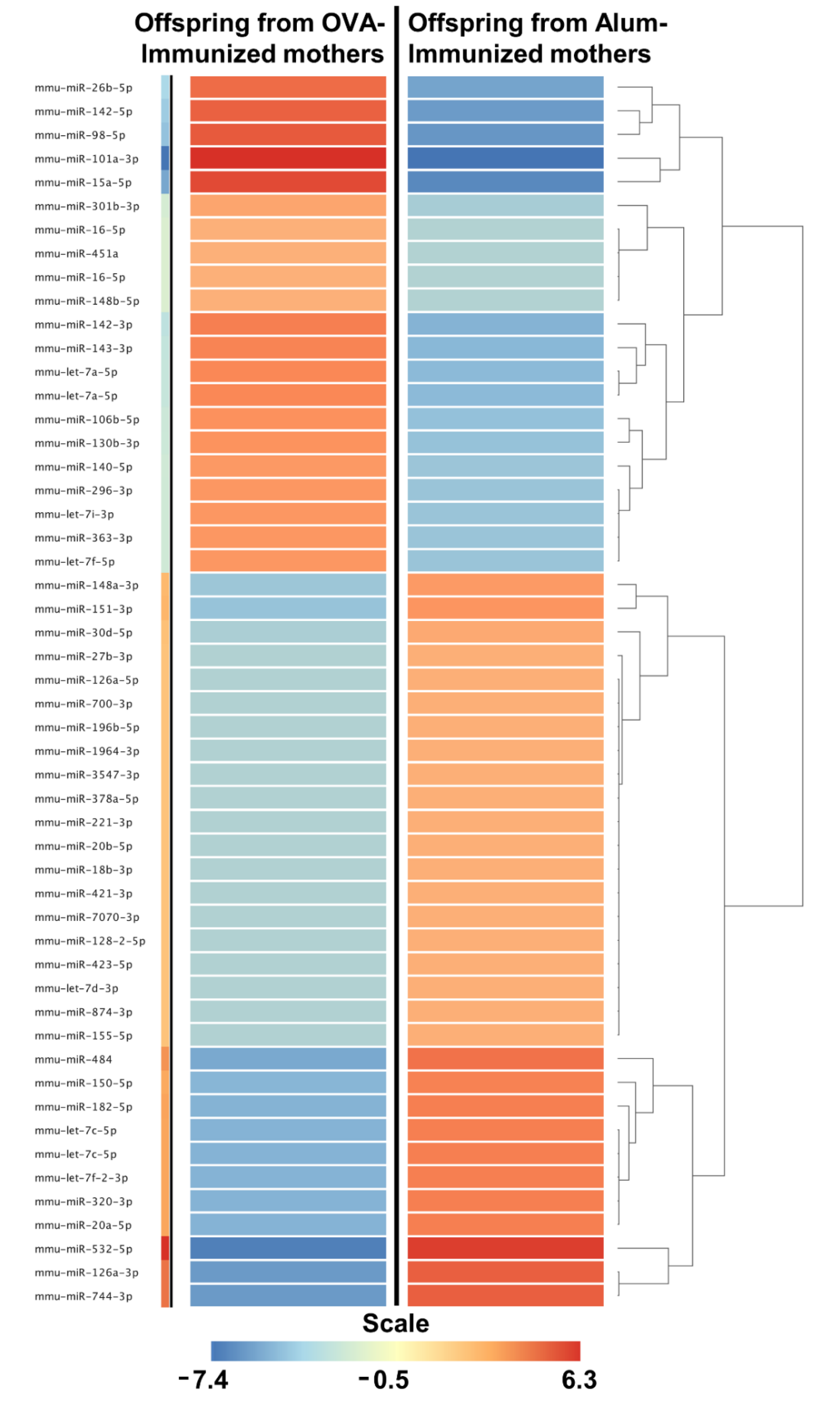
2.2. Maternal Immunization Could Induce the Expression of IL-17 Inhibition-Related miRNAs on Offspring Thymus
3. Discussion
4. Materials and Methods
4.1. Immunization
4.2. Flow Cytometry
4.3. sRNA Sequencing
4.3.1. RNA Extraction
4.3.2. sRNA Construction and Sequencing
4.3.3. sRNA Data Analysis and Interpretation
4.3.4. Target Genes and Gene Ontology (GO) Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fusaro, A.E.; de Brito, C.A.; Taniguchi, E.F.; Muniz, B.P.; Victor, J.R.; Orii, N.M.; da Silva Duarte, A.J.; Sato, M.N. Balance between early life tolerance and sensitization in allergy: Dependence on the timing and intensity of prenatal and postnatal allergen exposure of the mother. Immunology 2009, 128, e541–e550. [Google Scholar] [CrossRef]
- Matson, A.P.; Zhu, L.; Lingenheld, E.G.; Schramm, C.M.; Clark, R.B.; Selander, D.M.; Thrall, R.S.; Breen, E.; Puddington, L. Maternal transmission of resistance to development of allergic airway disease. J. Immunol. 2007, 179, 1282–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhasselt, V.; Milcent, V.; Cazareth, J.; Kanda, A.; Fleury, S.; Dombrowicz, D.; Glaichenhaus, N.; Julia, V. Breast milk-mediated transfer of an antigen induces tolerance and protection from allergic asthma. Nat. Med. 2008, 14, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Ellertsen, L.K.; Nygaard, U.C.; Melkild, I.; Løvik, M. Maternal allergen immunisation to prevent sensitisation in offspring: Th2-polarising adjuvants are more efficient than a Th1-polarising adjuvant in mice. BMC Immunol. 2010, 11, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Victor, J.R.; Muniz, B.P.; Fusaro, A.E.; de Brito, C.A.; Taniguchi, E.F.; Duarte, A.J.S.; Sato, M.N. Maternal immunization with ovalbumin prevents neonatal allergy development and up-regulates inhibitory receptor Fc gamma RIIB expression on B cells. BMC Immunol. 2010, 11. [Google Scholar] [CrossRef] [Green Version]
- De Lima Lira, A.A.; de Oliveira, M.G.; de Oliveira, L.M.; da Silva Duarte, A.J.; Sato, M.N.; Victor, J.R. Maternal immunization with ovalbumin or Dermatophagoides pteronyssinus has opposing effects on Fc gamma RIIb expression on offspring B cells. Allergy Asthma Clin. Immunol. 2014, 10. [Google Scholar] [CrossRef] [Green Version]
- Fusaro, A.; Maciel, M.; Victor, J.; Oliveira, C.; Duarte, A.; Sato, M. Influence of maternal murine immunization with Dermatophagoides pteronyssinus extract on the type I hypersensitivity response in offspring. Int. Arch. Allergy Immunol. 2002, 127, 208–216. [Google Scholar] [CrossRef]
- Uthoff, H.; Spenner, A.; Reckelkamm, W.; Ahrens, B.; Wölk, G.; Hackler, R.; Hardung, F.; Schaefer, J.; Scheffold, A.; Renz, H.; et al. Critical role of preconceptional immunization for protective and nonpathological specific immunity in murine neonates. J. Immunol. 2003, 171, 3485–3492. [Google Scholar] [CrossRef] [Green Version]
- Victor, J.; Fusaro, A.; Duarte, A.; Sato, M. Preconception maternal immunization to dust mite inhibits the type I hypersensitivity response of offspring. J. Allergy Clin. Immunol. 2003, 111, 269–277. [Google Scholar] [CrossRef] [PubMed]
- de-Oliveira, M.G.; de LimaSgnotto, A.A.; da RessureiçãoInoue, F.; SabôDuarte, A.H.; da SilvaVictor, A.J.; Russo, J. Preconception immunization can modulate intracellular Th2 cytokine profile in offspring: In vivo influence of interleukin 10 and B/T cell collaboration. Cent. Eur. J. Immunol. 2018, 43, 10. [Google Scholar] [CrossRef]
- Fusaro, A.E.; Brito, C.A.; Victor, J.R.; Rigato, P.O.; Goldoni, A.L.; Duarte, A.J.S.; Sato, M.N. Maternal-fetal interaction: Preconception immunization in mice prevents neonatal sensitization induced by allergen exposure during pregnancy and breastfeeding. Immunology 2007, 122, 107–115. [Google Scholar] [CrossRef]
- Garcia de Oliveira, M.; da Ressureição Sgnotto, F.; Rodrigues de Sousa, T.; Fagundes, B.O.; Duarte, A.J.D.S.; Victor, J.R. Preconceptional immunization with an allergen inhibits offspring thymic Th17 cells maturation without influence on Th1 and Th2 cells. Eur. Cytokine Netw. 2020. [Google Scholar] [CrossRef]
- Ness-Schwickerath, K.J.; Morita, C.T. Regulation and function of IL-17A- and IL-22-producing γδ T cells. Cell Mol. Life Sci. 2011, 68, 2371–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Victor, J.R.; Lezmi, G.; Leite-de-Moraes, M. New Insights into Asthma Inflammation: Focus on iNKT, MAIT, and γδT Cells. Clin. Rev. Allergy Immunol. 2020. [Google Scholar] [CrossRef]
- de Sousa, T.R.; Victor, J.R. Natural Self-Ligand Gamma Delta T Cell Receptors (γδTCRs) Insight: The Potential of Induced IgG. Vaccines 2020, 8, 436. [Google Scholar] [CrossRef]
- Belkadi, A.; Dietrich, C.; Machavoine, F.; Victor, J.R.; Leite-de-Moraes, M. γδ T cells amplify Blomia tropicalis-induced allergic airway disease. Allergy 2019, 74, 395–398. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, M.G.; de Lima Lira, A.A.; da Ressureição Sgnotto, F.; Inoue, A.H.S.; Santos, L.S.; Nakamatsu, B.Y.; Duarte, A.J.D.S.; Leite-de-Moraes, M.; Victor, J.R. Maternal IgG impairs the maturation of offspring intrathymic IL-17-producing γδT cells: Implications for murine and human allergies. Clin. Exp. Allergy 2019. [Google Scholar] [CrossRef]
- Victor, J.R. Allergen-specific IgG as a mediator of allergy inhibition: Lessons from mother to child. Hum. Vaccines Immunother. 2017, 13, 507–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Victor, J.R. Do different IgG repertoires play a role in B- and T-cell functional modulation during ontogeny? The “hooks without bait” theory. Immunol. Cell Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Victor, J.R. Influence of maternal immunization with allergens on the thymic maturation of lymphocytes with regulatory potential in children: A broad field for further exploration. J. Immunol. Res. 2014, 2014, 780386. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.H.S.; Lira, A.A.L.; de-Oliveira, M.G.; de Sousa, T.R.; Sgnotto, F.D.R.; Duarte, A.J.D.S.; Victor, J.R. The Potential of IgG to Induce Murine and Human Thymic Maturation of IL-10+ B Cells (B10) Revealed in a Pilot Study. Cells 2020, 9, 2239. [Google Scholar] [CrossRef]
- Santos, L.S.; Sgnotto, F.D.R.; Sousa, T.R.; Orfali, R.L.; Aoki, V.; Duarte, A.J.D.S.; Victor, J.R. IgG from atopic dermatitis patients induces non-atopic infant thymic invariant natural killer T (iNKT) cells to produce IL-4, IL-17, and IL-10. Int. J. Derm. 2019. [Google Scholar] [CrossRef]
- Da Ressureição Sgnotto, F.; Souza Santos, L.; Rodrigues de Sousa, T.; Feitosa de Lima, J.; Mara da Silva Oliveira, L.; Saeed Sanabani, S.; José da Silva Duarte, A.; Russo Victor, J. IgG From HIV-1-Exposed Seronegative and HIV-1-Infected Subjects Differently Modulates IFN-γ Production by Thymic T and B Cells. J. Acquir. Immune Defic. Syndr. 2019, 82, e56–e60. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.S.; Sgnotto, F.D.R.; Inoue, A.H.S.; Padreca, A.F.; Menghini, R.P.; Duarte, A.J.D.S.; Victor, J.R. IgG from Non-atopic Individuals Induces In Vitro IFN-γ and IL-10 Production by Human Intra-thymic γδT Cells: A Comparison with Atopic IgG and IVIg. Arch. Immunol. Ther. Exp. 2019. [Google Scholar] [CrossRef] [PubMed]
- Sgnotto, F.D.R.; de Oliveira, M.G.; Lira, A.A.L.; Inoue, A.H.S.; Titz, T.O.; Orfali, R.L.; Bento-de-Souza, L.; Sato, M.N.; Aoki, V.; Duarte, A.J.S.; et al. IgG from atopic dermatitis patients induces IL-17 and IL-10 production in infant intrathymic TCD4 and TCD8 cells. Int. J. Derm. 2018, 57, 434–440. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, M.G.; Lira, A.A.L.; Sgnotto, F.D.R.; Inoue, A.H.S.; Beltrame, G.R.; da Silva, D.; Menghini, R.P.; Duarte, A.J.D.S.; Victor, J.R. Maternal immunization downregulates offspring TCD4 regulatory cells (Tregs) thymic maturation without implications for allergy inhibition. Scand. J. Immunol. 2018, e12721. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, M.G.; Oliveira, L.M.; Lira, A.A.L.; Sgnotto, F.D.R.; Duarte, A.J.D.S.; Sato, M.N.; Victor, J.R. Preconception allergen sensitization can induce B10 cells in offspring: A potential main role for maternal IgG. Allergy Asthma Clin. Immunol. 2017, 13, 22. [Google Scholar] [CrossRef] [Green Version]
- Sgnotto, F.D.R.; Oliveira, M.G.; Lira, A.A.L.; Bento-de-Souza, L.; Duarte, A.J.D.S.; Victor, J.R. Low doses of IgG from atopic individuals can modulate in vitro IFN-γ production by human intra-thymic TCD4 and TCD8 cells: An IVIg comparative approach. Hum. Vaccin. Immunother. 2017, 13, 1563–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribot, J.C.; deBarros, A.; Pang, D.J.; Neves, J.F.; Peperzak, V.; Roberts, S.J.; Girardi, M.; Borst, J.; Hayday, A.C.; Pennington, D.J.; et al. CD27 is a thymic determinant of the balance between interferon-gamma- and interleukin 17-producing gammadelta T cell subsets. Nat. Immunol. 2009, 10, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Ribot, J.C.; Silva-Santos, B. Differentiation and activation of γδ T Lymphocytes: Focus on CD27 and CD28 costimulatory receptors. Adv. Exp. Med. Biol. 2013, 785, 95–105. [Google Scholar] [CrossRef]
- Cahill, R.N.; Kimpton, W.G.; Washington, E.A.; Walker, I.D. Origin and development of the gamma delta T-cell system in sheep: A critical role for the thymus in the generation of TcR diversity and tissue tropism. Semin. Immunol. 1996, 8, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Carding, S.R.; Allan, W.; Kyes, S.; Hayday, A.; Bottomly, K.; Doherty, P.C. Late dominance of the inflammatory process in murine influenza by gamma/delta + T cells. J. Exp. Med. 1990, 172, 1225–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carding, S.R.; Egan, P.J. Gammadelta T cells: Functional plasticity and heterogeneity. Nat. Rev. Immunol. 2002, 2, 336–345. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, R.L.; Born, W.K. gammadelta T cell subsets: A link between TCR and function? Semin. Immunol. 2010, 22, 193–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonneville, M.; O’Brien, R.L.; Born, W.K. Gammadelta T cell effector functions: A blend of innate programming and acquired plasticity. Nat. Rev. Immunol. 2010, 10, 467–478. [Google Scholar] [CrossRef]
- Narayan, K.; Sylvia, K.E.; Malhotra, N.; Yin, C.C.; Martens, G.; Vallerskog, T.; Kornfeld, H.; Xiong, N.; Cohen, N.R.; Brenner, M.B.; et al. Intrathymic programming of effector fates in three molecularly distinct γδ T cell subtypes. Nat. Immunol. 2012, 13, 511–518. [Google Scholar] [CrossRef]
- Roark, C.L.; French, J.D.; Taylor, M.A.; Bendele, A.M.; Born, W.K.; O’Brien, R.L. Exacerbation of collagen-induced arthritis by oligoclonal, IL-17-producing gamma delta T cells. J. Immunol. 2007, 179, 5576–5583. [Google Scholar] [CrossRef] [Green Version]
- Gerber, D.J.; Azuara, V.; Levraud, J.P.; Huang, S.Y.; Lembezat, M.P.; Pereira, P. IL-4-producing gamma delta T cells that express a very restricted TCR repertoire are preferentially localized in liver and spleen. J. Immunol. 1999, 163, 3076–3082. [Google Scholar]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Malone, C.D.; Hannon, G.J. Small RNAs as guardians of the genome. Cell 2009, 136, 656–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, J.D.; Nistala, K.; Petermann, F.; Saran, N.; Chennupati, V.; Schmitz, S.; Korn, T.; Wedderburn, L.R.; Forster, R.; Krueger, A.; et al. Expression of miRNAs miR-133b and miR-206 in the Il17a/f locus is co-regulated with IL-17 production in alphabeta and gammadelta T cells. PLoS ONE 2011, 6, e20171. [Google Scholar] [CrossRef] [PubMed]
- Schmolka, N.; Papotto, P.H.; Romero, P.V.; Amado, T.; Enguita, F.J.; Amorim, A.; Rodrigues, A.F.; Gordon, K.E.; Coroadinha, A.S.; Boldin, M.; et al. MicroRNA-146a controls functional plasticity in gammadelta T cells by targeting NOD1. Sci. Immunol. 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lira, A.A.L.; de-Oliveira, M.G.; Inoue, A.H.S.; Beltrame, G.R.; Duarte, A.J.D.S.; Victor, J.R. Preconceptional allergen immunization can induce offspring IL-17 secreting B cells (B17): Do they share similarities with regulatory B10 cells? Allergol. Immunopathol. 2018, 46, 454–459. [Google Scholar] [CrossRef]
- Xiong, N.; Kang, C.; Raulet, D.H. Positive selection of dendritic epidermal gammadelta T cell precursors in the fetal thymus determines expression of skin-homing receptors. Immunity 2004, 21, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Cobb, B.S.; Hertweck, A.; Smith, J.; O’Connor, E.; Graf, D.; Cook, T.; Smale, S.T.; Sakaguchi, S.; Livesey, F.J.; Fisher, A.G.; et al. A role for Dicer in immune regulation. J. Exp. Med. 2006, 203, 2519–2527. [Google Scholar] [CrossRef]
- Mildner, A.; Chapnik, E.; Varol, D.; Aychek, T.; Lampl, N.; Rivkin, N.; Bringmann, A.; Paul, F.; Boura-Halfon, S.; Hayoun, Y.S.; et al. MicroRNA-142 controls thymocyte proliferation. Eur. J. Immunol. 2017, 47, 1142–1152. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Liu, J.; Liu, X.; Huang, E.; Yang, J.; Qian, J.; Zhang, D.; Liu, R.; Chu, Y. MicroRNA 15a/16-1 suppresses aryl hydrocarbon receptor-dependent interleukin-22 secretion in CD4. Hepatology 2018, 67, 1027–1040. [Google Scholar] [CrossRef] [Green Version]
- Zou, Q.; Yang, M.; Yu, M.; Liu, C. Influences of Regulation of miR-126 on Inflammation, Th17/Treg Subpopulation Differentiation, and Lymphocyte Apoptosis through Caspase Signaling Pathway in Sepsis. Inflammation 2020, 43, 2287–2300. [Google Scholar] [CrossRef]
- Chen, X.; Yu, X.; Li, X.; Li, L.; Li, F.; Guo, T.; Guan, C.; Miao, L.; Cao, G. MiR-126 targets IL-17A to enhance proliferation and inhibit apoptosis in high-glucose-induced human retinal endothelial cells. Biochem. Cell Biol. 2020, 98, 277–283. [Google Scholar] [CrossRef]
- Dinesh, P.; Kalaiselvan, S.; Sujitha, S.; Rasool, M. MicroRNA-532-3p Regulates Pro-Inflammatory Human THP-1 Macrophages by Targeting ASK1/p38 MAPK Pathway. Inflammation 2021, 44, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Valadão de Souza, D.R.; Pessôa, R.; Nascimento, A.; Nukui, Y.; Pereira, J.; Casseb, J.; Penalva de Oliveira, A.C.; da Silva Duarte, A.J.; Clissa, P.B.; Sanabani, S.S. Small RNA profiles of HTLV-1 asymptomatic carriers with monoclonal and polyclonal rearrangement of the T-cell antigen receptor γ-chain using massively parallel sequencing: A pilot study. Oncol. Lett. 2020, 20, 2311–2321. [Google Scholar] [CrossRef]
- Nascimento, A.; Valadão de Souza, D.R.; Pessôa, R.; Pietrobon, A.J.; Nukui, Y.; Pereira, J.; Casseb, J.; Penalva de Oliveira, A.C.; Loureiro, P.; da Silva Duarte, A.J.; et al. Global expression of noncoding RNome reveals dysregulation of small RNAs in patients with HTLV-1-associated adult T-cell leukemia: A pilot study. Infect. Agent Cancer 2021, 16, 4. [Google Scholar] [CrossRef]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. miRWalk: An online resource for prediction of microRNA binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de-Sousa, T.R.; Pessôa, R.; Nascimento, A.; Fagundes, B.O.; Sgnotto, F.d.R.; Duarte, A.J.d.S.; Sanabani, S.S.; Victor, J.R. Preconceptional Immunization Can Modulate Offspring Intrathymic IL-17-Producing γδT Cells with Epigenetic Implications Mediated by microRNAs. Int. J. Mol. Sci. 2021, 22, 6633. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126633
de-Sousa TR, Pessôa R, Nascimento A, Fagundes BO, Sgnotto FdR, Duarte AJdS, Sanabani SS, Victor JR. Preconceptional Immunization Can Modulate Offspring Intrathymic IL-17-Producing γδT Cells with Epigenetic Implications Mediated by microRNAs. International Journal of Molecular Sciences. 2021; 22(12):6633. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126633
Chicago/Turabian Stylede-Sousa, Thamires Rodrigues, Rodrigo Pessôa, Andrezza Nascimento, Beatriz Oliveira Fagundes, Fábio da Ressureição Sgnotto, Alberto José da Silva Duarte, Sabri Saeed Sanabani, and Jefferson Russo Victor. 2021. "Preconceptional Immunization Can Modulate Offspring Intrathymic IL-17-Producing γδT Cells with Epigenetic Implications Mediated by microRNAs" International Journal of Molecular Sciences 22, no. 12: 6633. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126633