
Mitochondrial Function Are Disturbed in the Presence of the Anticancer Drug, 3-Bromopyruvate

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
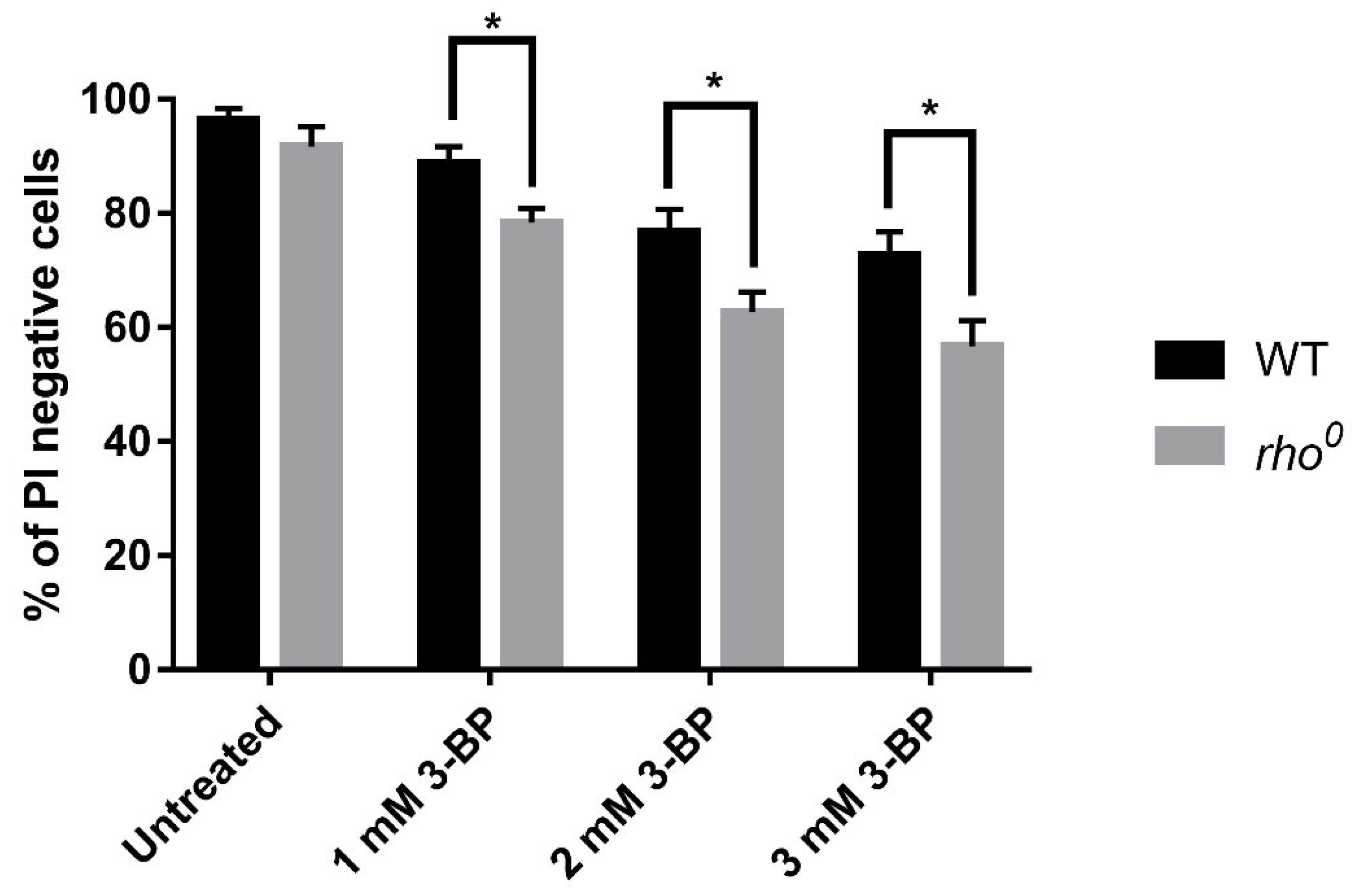
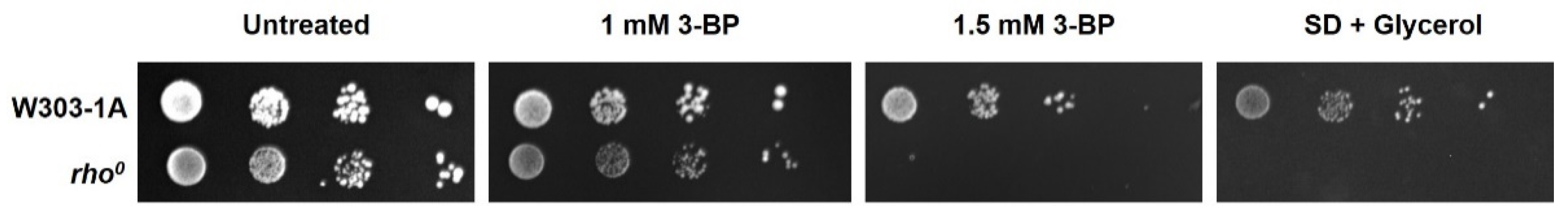
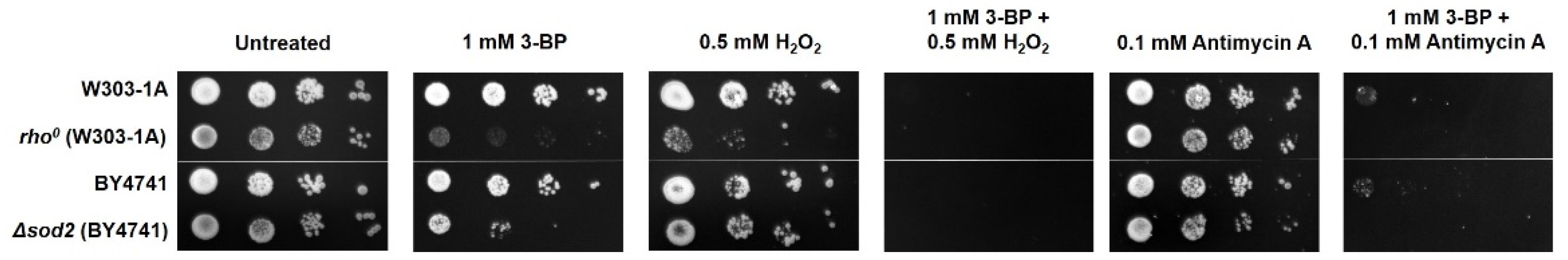
2.1. The Role of Functional Yeast Mitochondria in Response to 3-BP—Viability and Sensitivity of W303-1A and rho0 Strains to 3-BP
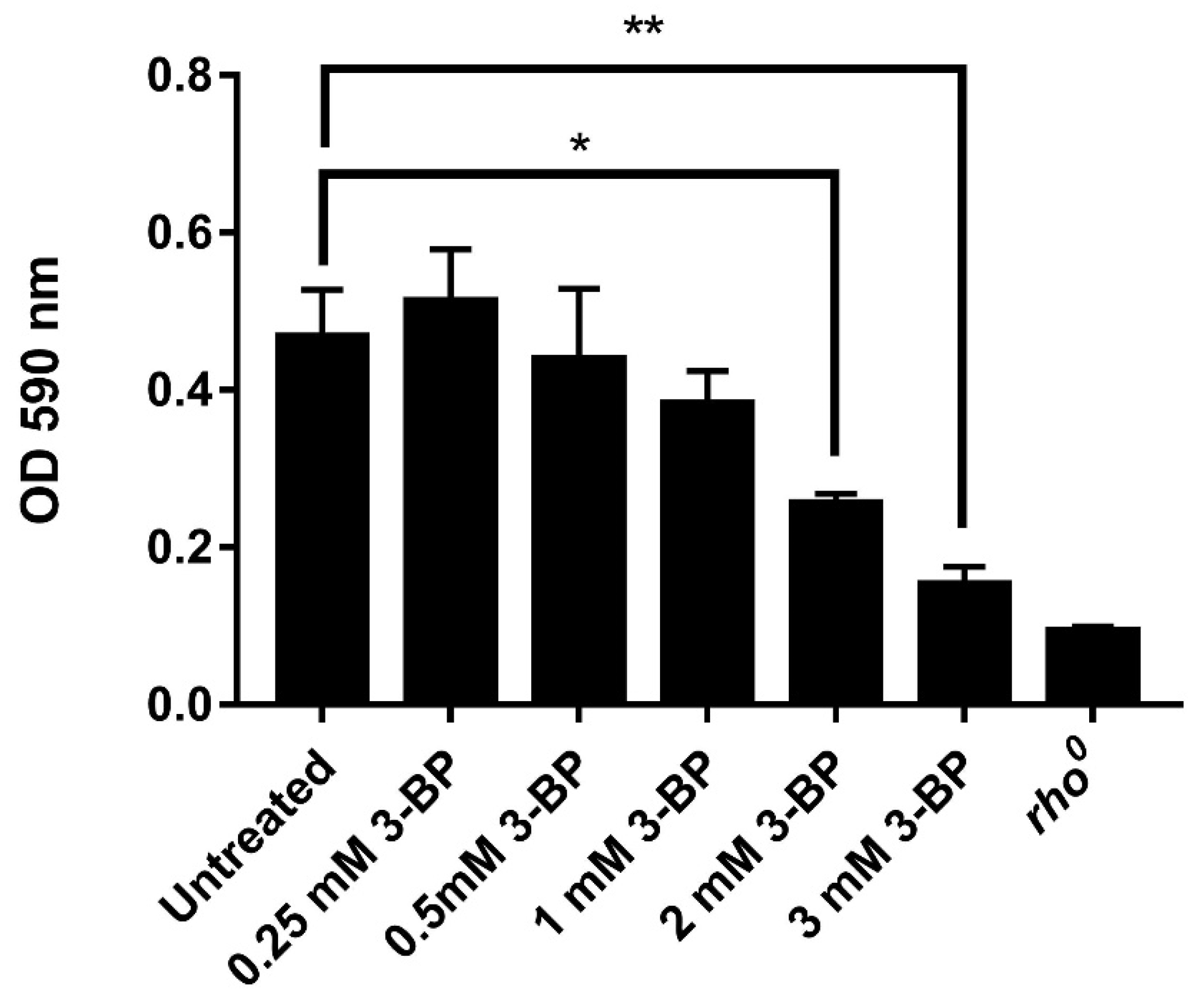
2.2. 3-BP Disturbs Mitochondrial Function and Reduces Metabolic Activity in Yeast
S. cerevisiae Cells
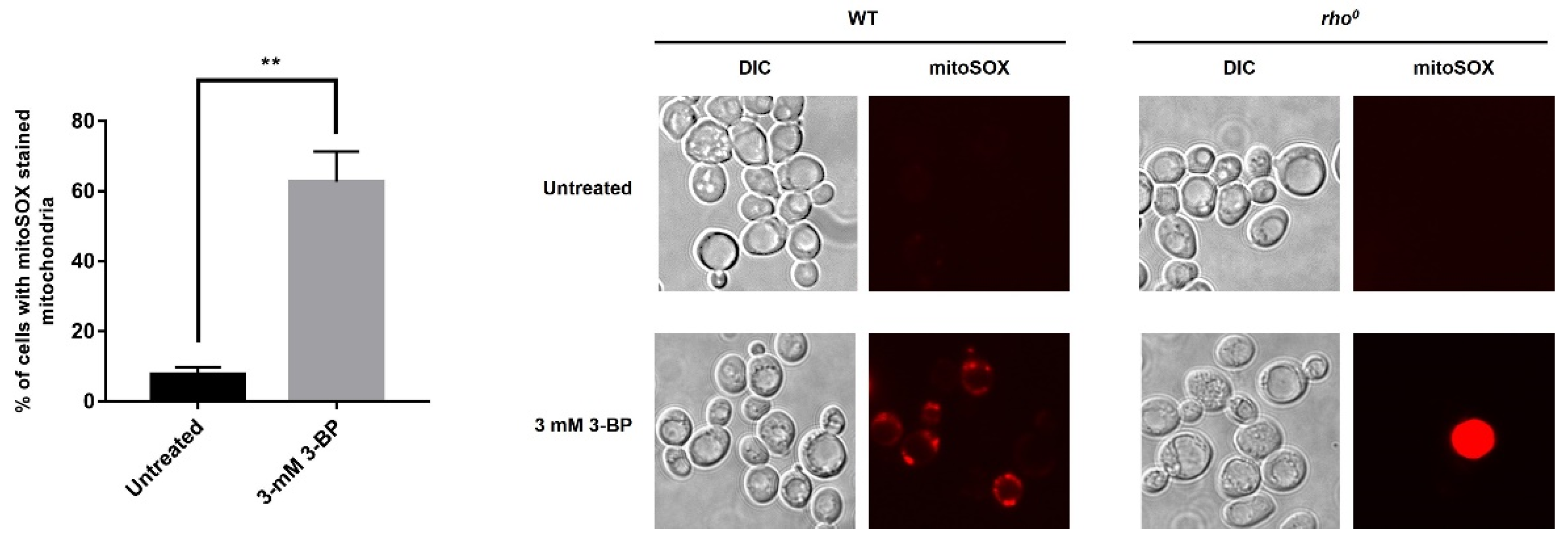
2.3. Exposure to 3-BP Leads to Generation of Superoxide Anion in Mitochondria
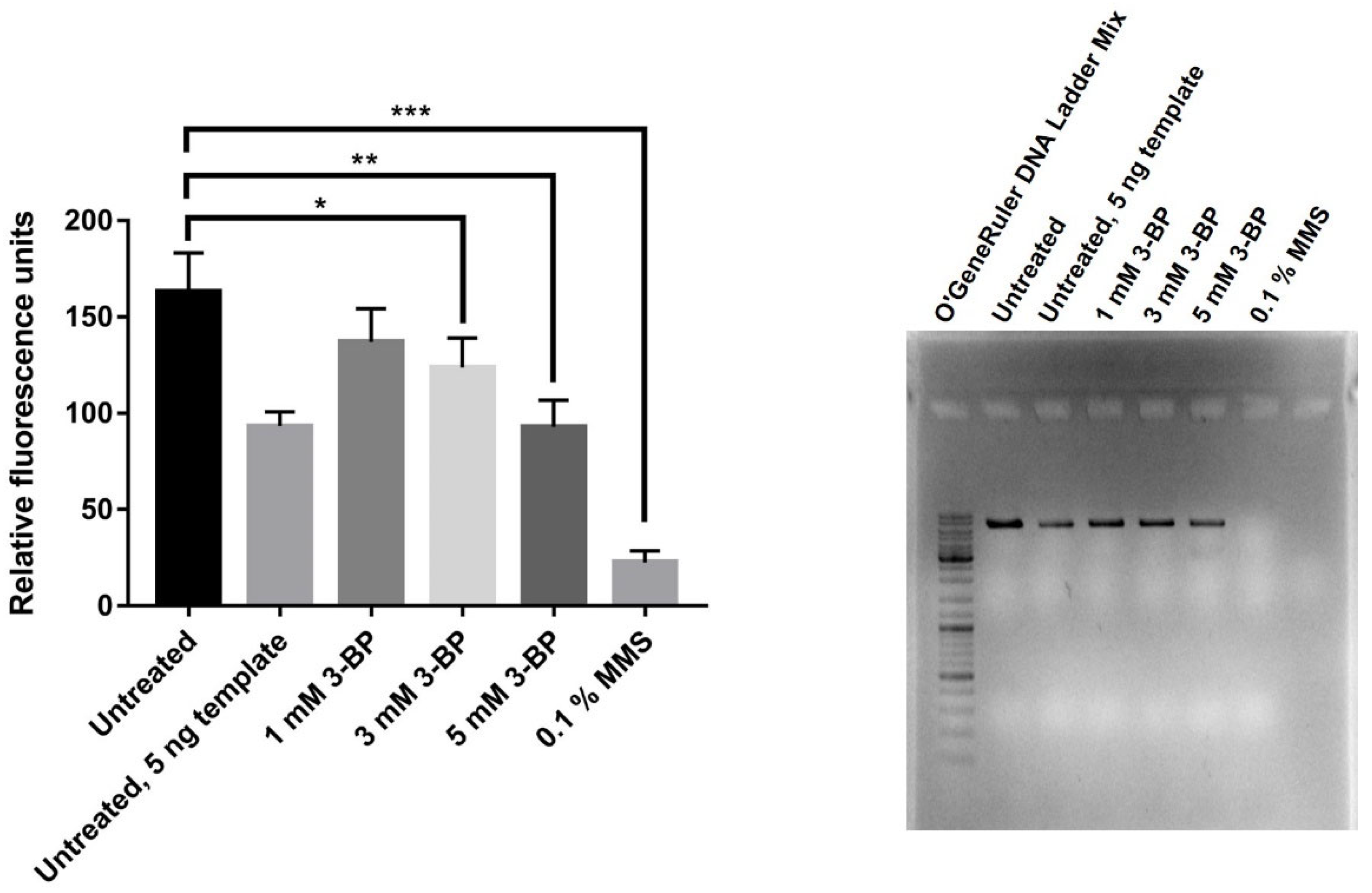
2.4. 3-BP Induces Lesion Formation in mtDNA in Yeast
2.5. Compounds Inhibiting Electron Transport Chain (ETC) and Inducing Oxidative Stress Work Synergistically with 3-BP
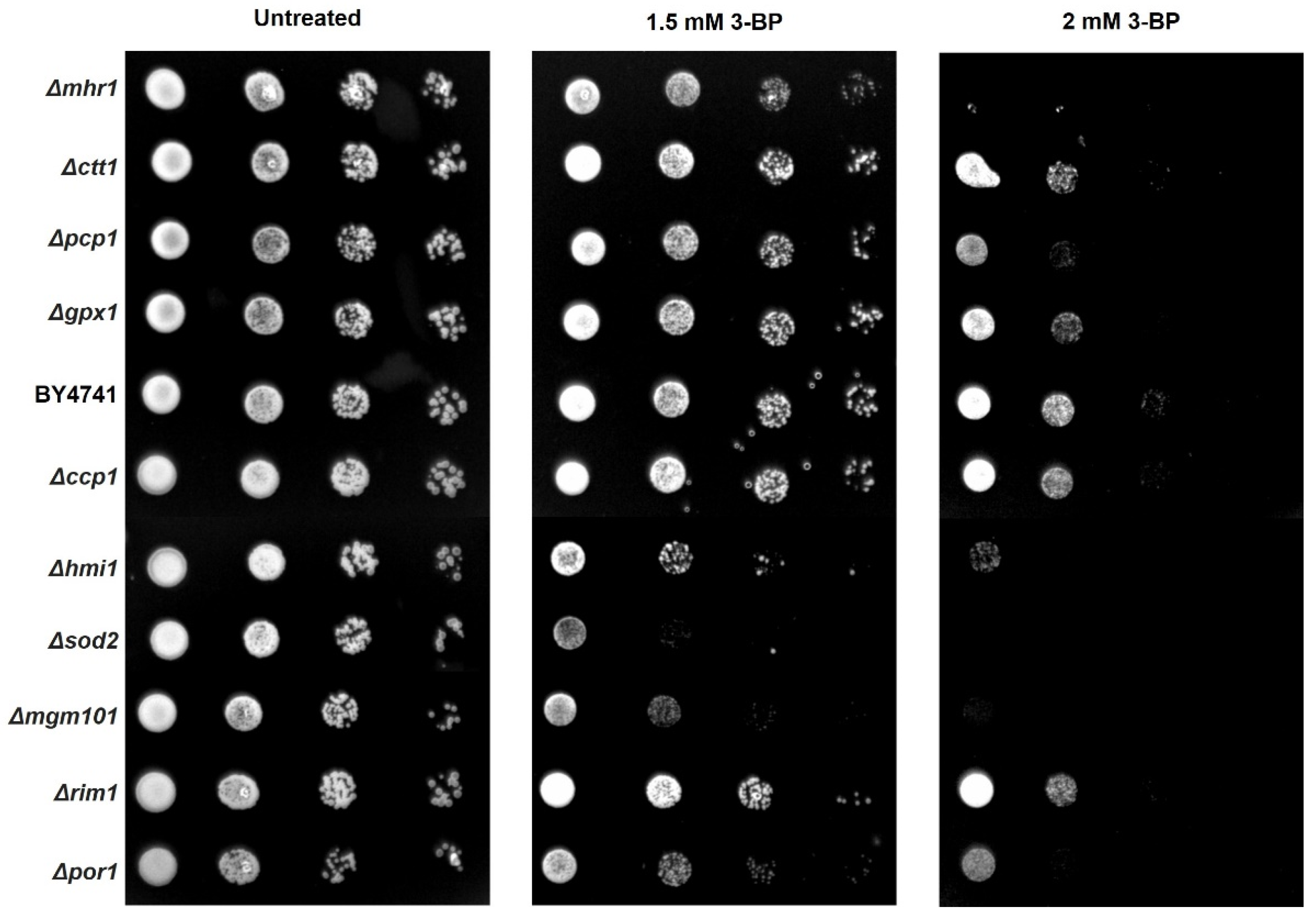
2.6. Role of mtDNA Maintenance, mtDNA Repair Proteins and Antioxidant Enzymes in Tolerance to 3-BP
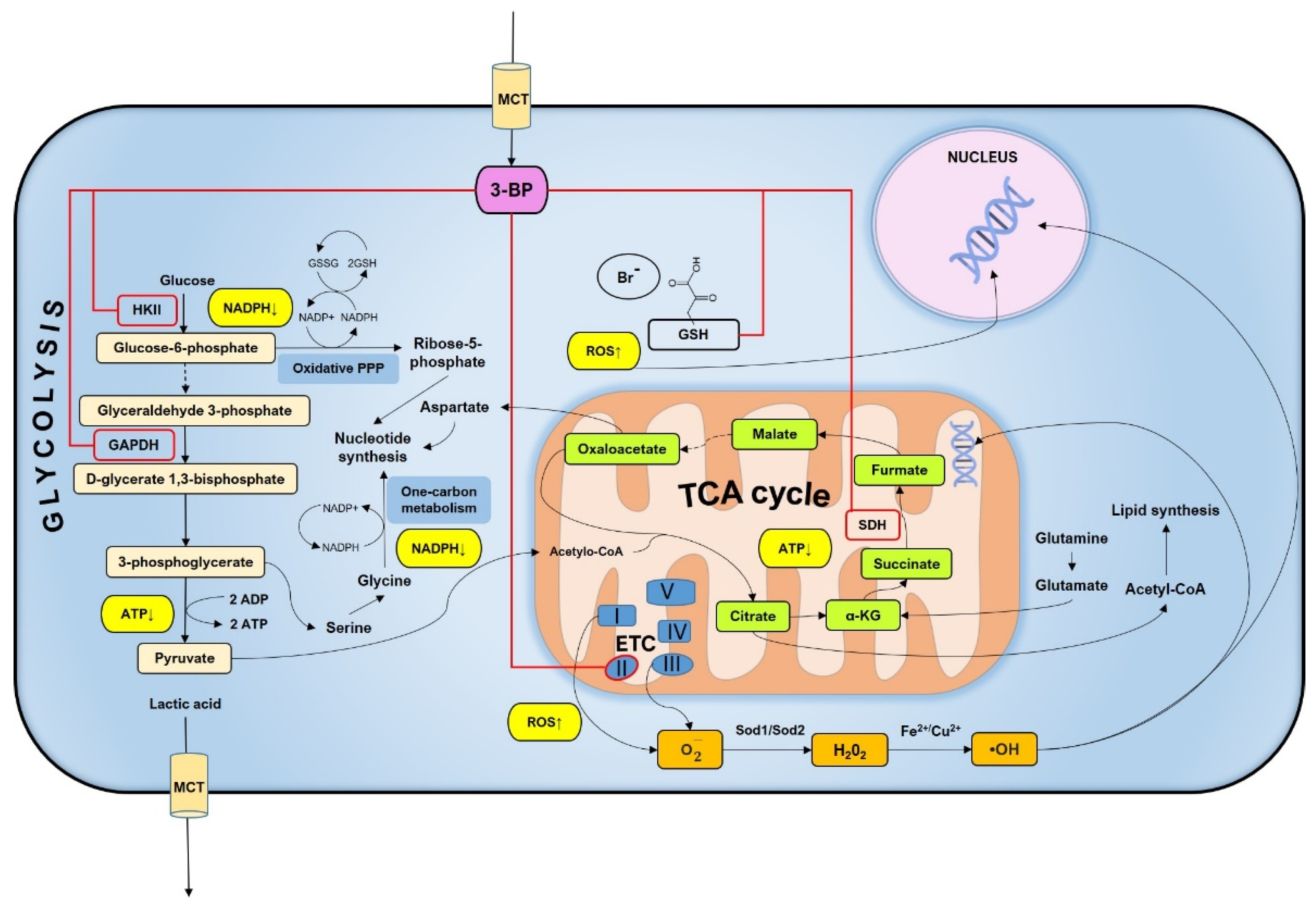
3. Discussion
4. Materials and Methods
4.1. The Yeast Strains, Growth Conditions and Cell Treatment
4.2. Cell Viability Assay—Propidium Iodide (PI) Staining
4.3. Yeast Sensitivity Test (Spot Test)
4.4. Mitochondrial ROS Measurements
4.5. Quantitative Polymerase Chain Reaction
- 1 µL of 10 ng/µL template DNA
- 12.5 µL of LongAmp® Taq 2X Master Mix (New England Biolabs, Ipswich, MA, USA)
- 10 µL of Nuclease-free water
- 1 µL of 10 µM Sense primer (Genomed, Warszawa, Poland), 5′-GTG CGT ATA TTT CGT TGA TGC GT-3′
- 1 µL of 10µM Antisense primer (Genomed, Warszawa, Poland), 5′-GTC ACC ACC TCC TGC TAC TTC AA-3′
4.6. MTT Assay
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Queirós, O.; Preto, A.; Pacheco, A.; Pinheiro, C.; Azevedo-Silva, J.; Moreira, R.; Pedro, M.; Ko, Y.H.; Pedersen, P.L.; Baltazar, F.; et al. Butyrate activates the monocarboxylate transporter MCT4 expression in breast cancer cells and enhances the antitumor activity of 3-bromopyruvate. J. Bioenergy Biomembr. 2012, 44, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Birsoy, K.; Wang, T.; Possemato, R.; Yilmaz, O.H.; Koch, C.E.; Chen, W.W.; Hutchins, A.W.; Gultekin, Y.; Peterson, T.R.; Carette, J.E.; et al. MCT1 mediated transport of a toxic molecule is an effective strategy for targeting glycolytic tumors. Nat. Genet. 2013, 45, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Lis, P.; Zarzycki, M.; Ko, Y.H.; Casal, M.; Pedersen, P.L.; Goffeau, A.; Ułaszewski, S. Transport and cytotoxicity of the anticancer drug 3-bromopyruvate in the yeast Saccharomyces cerevisiae. J. Bioenerg. Biomembr. 2012, 44, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, P.L.; Mathupala, S.; Rempel, A.; Geschwind, J.F.; Ko, Y.H. Mitochondrial bound type II hexokinase: A key player in the growth and survival of many cancers and an ideal prospect for therapeutic intervention. Biochim. Biophys. Acta. 2002, 1555, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Mathupala, S.P.; Ko, Y.H.; Pedersen, P.L. Hexokinase II: Cancer’s double-edged sword acting as both facilitator and gatekeeper of malignancy when bound to mitochondria. Oncogene 2006, 25, 4777–4786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathupala, S.P.; Ko, Y.H.; Pedersen, P.L. Hexokinase-2 bound to mitochondria: Cancer’s stygian link to the “Warburg effect” and a pivotal target for effective therapy. Semin. Cancer Biol. 2009, 19, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, Y.H.; Smith, B.L.; Wang, Y.; Pomper, M.G.; Rini, D.A.; Torbenson, M.S.; Hullihen, J.; Pedersen, P.L. Advanced cancers: Eradication in all cases using 3-bromopyruvate therapy to deplete ATP. Biochem. Biophys. Res. Commun. 2004, 324, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy-Kanniappan, S.; Kunjithapatham, R.; Geschwind, J.F. Anticancer efficacy of the metabolic blocker 3-bromopyruvate: Specific molecular targeting. Anticancer Res. 2013, 33, 13–20. [Google Scholar] [PubMed]
- Macchioni, L.; Davidescu, M.; Roberti, R.; Corazzi, L. The energy blockers 3-bromopyruvate and lonidamine: Effects on bioenergetics of brain mitochondria. J. Bioenerg Biomembr. 2014, 46, 389–394. [Google Scholar] [CrossRef] [PubMed]
- El Sayed, S.M.; El-Magd, R.A.; Shishido, Y.; Chung, S.P.; Sakai, T.; Watanabe, H.; Kagami, S.; Fukui, K. D-amino acid oxidase gene therapy sensitizes glioma cells to the antiglycolytic effect of 3-bromopyruvate. Cancer Gene Ther. 2012, 19, 1. [Google Scholar] [CrossRef] [Green Version]
- Ihrlund, L.S.; Hernlund, E.; Khan, O.; Shoshan, M.C. 3-Bromopyruvate as inhibitor of tumour cell energy metabolism and chemopotentiator of platinum drugs. Mol. Oncol. 2008, 2, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Ahn, K.J.; Kim, J.A.; Kim, H.M.; Lee, J.D.; Lee, J.M.; Kim, S.J.; Park, J.H. Role of reactive oxygen species-mediated mitochondrial dysregulation in 3-bromopyruvate induced cell death in hepatoma cells. J. Bioenerg. Biomembr. 2008, 40, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, Z.; Zou, X.; Lan, Y.; Sun, X.; Wang, X.; Zhao, S.; Jiand, C.; Liu, H. Mechanisms underlying 3-bromopyruvate-induced cell death in colon cancer. J. Bioenerg. Biomembr. 2015, 47, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Sadowska-Bartosz, I.; Bartosz, G. Effect of 3-bromopyruvic acid on human erythrocyte antioxidant defense system. Cell Biol. Int. 2013, 37, 1285–1290. [Google Scholar] [CrossRef] [PubMed]
- Sadowska-Bartosz, I.; Szewczyk, R.; Jaremko, L.; Jaremko, M.; Bartosz, G. Anticancer agent 3-bromopyruvic acid forms a conjugate with glutathione. Pharmacol. Rep. 2016, 68, 502–505. [Google Scholar] [CrossRef]
- Niedźwiecka, K.; Dyląg, M.; Augustyniak, D.; Majkowska-Skrobek, G.; Cal-Bąkowska, M.; Ko, Y.H.; Ułaszewski, S. Glutathione may have implications in the design of 3-bromopyruvate treatment protocols for both fungal and algal infections as well as multiple myeloma. Oncotarget 2016, 7, 65614–65626. [Google Scholar] [CrossRef] [Green Version]
- El Sayed, S.M.; Baghdadi, H.; Zolaly, M.; Almaramhy, H.H.; Ayat, M.; Donki, J.G. The promising anticancer drug 3-bromopyruvate is metabolized through glutathione conjugation which affects chemoresistance and clinical practice: An evidence-based view. Med. Hypotheses 2017, 100, 67–77. [Google Scholar] [CrossRef]
- Hirschhaeuser, F.; Sattler, U.G.; Mueller-Klieser, W. Lactate: A metabolic key player in cancer. Cancer Res. 2011, 71, 6921–6925. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, P.L. 3-Bromopyruvate (3BP) a fast acting, promising, powerful, specific, and effective “small molecule” anti-cancer agent taken from labside to bedside: Introduction to a special issue. J. Bioenerg. Biomembr. 2012, 44, 1–6. [Google Scholar] [CrossRef]
- Moore, J.D.; Yazgan, O.; Ataian, Y.; Krebs, J.E. Diverse roles for histone H2A modifications in DNA damage response pathways in yeast. Genetics 2007, 176, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Majkowska-Skrobek, G.; Augustyniak, D.; Lis, P.; Bartkowiak, A.; Gonchar, M.; Ko, Y.H.; Pedersen, P.L.; Goffeau, A.; Ułaszewski, S. Killing multiple myeloma cells with the small molecule 3-bromopyruvate: Implications for therapy. Anticancer Drugs 2014, 25, 673–682. [Google Scholar] [CrossRef]
- Lanz, M.C.; Dibitetto, D.; Smolka, M.B. DNA damage kinase signaling: Checkpoint and repair at 30 years. Embo J. 2019, 38, e101801. [Google Scholar] [CrossRef]
- Xue, C.; Wang, W.; Crickard, J.B.; Moevus, C.J.; Kwon, Y.; Sung, P.; Greene, E.C. Regulatory control of Sgs1 and Dna2 during eukaryotic DNA end resection. Proc. Natl. Acad. Sci. USA 2019, 116, 6091–6100. [Google Scholar] [CrossRef] [Green Version]
- Cal, M.; Matyjaszczyk, I.; Litwin, I.; Augustyniak, D.; Ogórek, R.; Ko, Y.; Ułaszewski, S. The anticancer drug 3-bromopyruvate induces DNA damage potentially through reactive oxygen species in yeast and in human cancer cells. Cells 2020, 9, 1161. [Google Scholar] [CrossRef]
- Birsoy, K.; Wang, T.; Chen, W.; Freinkman, E.; Abu-Remaileh, M.; Sabatini, D.M. An essential role of the mitochondrial electron transport chain in cell proliferation is to enable aspartate synthesis. Cell 2015, 162, 540–551. [Google Scholar] [CrossRef] [Green Version]
- Van der Bliek, A.M.; Sedensky, M.M.; Morgan, P.G. Cell biology of the mitochondrion. Genetics 2017, 207, 843–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malina, C.; Larsson, C.; Nielsen, J. Yeast mitochondria: An overview of mitochondrial biology and the potential of mitochondrial systems biology. FEMS Yeast Res. 2018, 18, 5. [Google Scholar] [CrossRef]
- Pan, Y. Mitochondria, reactive oxygen species, and chronological aging: A message from yeast. Exp. Gerontol. 2011, 46, 847–852. [Google Scholar] [CrossRef]
- Dell’Antone, P. Targets of 3-bromopyruvate, a new, energy depleting, anticancer agent. Med. Chem. 2009, 5, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Jardim-Messeder, D.; Moreira-Pacheco, F. 3-Bromopyruvic acid inhibits tricarboxylic acid cycle and glutaminolysis in HepG2 cells. Anticancer Res. 2016, 36, 2233–2241. [Google Scholar] [PubMed]
- Lasserre, J.-P.; Dautant, A.; Aiyar, R.S.; Kucharczyk, R.; Glatigny, A.; Tribouillard–Tanvier, D.; Rytka, J.; Blondel, M.; Skoczen, N.; Reynier, P.; et al. Yeast as a system for modeling mitochondrial disease mechanisms and discovering therapies. Dis. Model. Mech. 2015, 8, 509–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larosa, V.; Remacle, C. Insights into the respiratory chain and oxidative stress. Biosci. Rep. 2018, 38, BSR20171492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luk, E.; Yang, M.; Jensen, L.T.; Bourbonnais, Y.; Culotta, V.C. Manganese activation of superoxide dismutase 2 in the mitochondria of Saccharomyces cerevisiae. J. Biol. Chem. 2005, 280, 22715–22720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Formenko, D.E.; Koc, A.; Agisheva, N.; Jacobsen, M.; Kaya, A.; Malinouski, M.; Rutherford, J.C.; Siu, K.-L.; Jin, D.-Y.; Winge, D.R.; et al. Thiol peroxidases mediate specific genome-wide regulation of gene expression in response to hydrogen peroxide. Proc. Natl. Acad. Sci. USA 2011, 108, 2729–2734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwolek-Mirek, M.; Zadrag-Tecza, R. Comparison of methods used for assessing the viability and vitality of yeast cells. FEMS Yeast Res. 2014, 1068–1079. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.V.; Tan, A.S. Characterization of the cellular reduction of 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT): Subcellular localization, substrate dependence, and involvement of mitochondrial electron transport in MTT reduction. Arch. Biochem. Biophys. 1993, 303, 474–482. [Google Scholar] [CrossRef]
- Yazgan, O.; Krebs, J.E. Mitochondrial and nuclear genomic integrity after oxidative damage in Saccharomyces cerevisiae. Front. Biosci. 2012, 17, 1079–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, X.; Xue, D.; Li, N.; Clark-Walker, G.D. A functional core of the mitochondrial genome maintenance protein Mgm101p in Saccharomyces cerevisiae determined with a temperature-conditional allele. FEMS Yeast Res. 2007, 7, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Ling, F.; Bradshaw, E.; Yoshida, M. Prevention of mitochondrial genomic instability in yeast by the mitochondrial recombinase Mhr1. Sci. Rep. 2019, 9, 5433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramanagoudr-Bhojappa, R.; Blair, L.P.; Tackett, A.J.; Raney, K.D. Physical and functional interaction between yeast Pif1 helicase and Rim1 single-stranded DNA binding protein. Nucleic Acids Res. 2013, 41, 1029–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, Y.H.; Verhoeven, H.A.; Lee, M.J.; Corbin, D.J.; Vogl, T.J.; Pedersen, P.L. A translational study “case report” on the small molecule “energy blocker” 3-bromopyruvate (3BP) as a potent anticancer agent: From bench side to bedside. J. Bioenerg. Biomembr. 2012, 44, 163–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, Y.H.; Niedźwiecka, K.; Casal, M.; Pedersen, P.L.; Ułaszewski, S. 3-bromopyruvate (3BP) as a potent anticancer therapy in honor and memory of the late Professor Andre Goffeau. Yeast 2018, 36, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Annesley, S.J.; Fisher, P.R. Mitochondria in health and disease. Cells 2019, 8, 680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, C.A.; Soares, E.V. Mitochondria are the main source and one of the targets of Pb(lead)-induced oxidative stress in the yeast Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2014, 98, 5153–5160. [Google Scholar] [CrossRef] [Green Version]
- Vujcic, M.; Shroff, M.; Singh, K.K. Genetic determinants of mitochondrial response to arsenic in yeast Saccharomyces cerevisiae. Cancer Res. 2007, 67, 9740–9749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, E.; Ros, J.; Belli, G.; Cabiscol, E. Redox control and oxidative stress in yeast cells. Biochim. Biophys. Acta 2008, 1780, 1217–1235. [Google Scholar] [CrossRef]
- Rasmussen, A.K.; Chatterjee, A.; Rasmussen, L.J.; Singh, K.K. Mitochondria-mediated nuclear mutator phenotype in Saccharomyces cerevisiae. Nucleic Acids Res. 2003, 31, 3909–3917. [Google Scholar] [CrossRef] [PubMed]
- Caldecott, K.W. DNA single-strand break repair. Exp. Cell Res. 2014, 329, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Pâques, F.; Haber, J.E. Multiple pathways of recombination induced by double-strand breaks in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 1999, 63, 349–404. [Google Scholar] [CrossRef] [Green Version]
- Thomas, D.; Scot, A.D.; Barbey, R.; Padula, M.; Boiteux, S. Inactivation of OGG1 increases the incidence of GC TA transversions in Saccharomyces cerevisiae: Evidence for endogenous oxidative damage to DNA in eukaryotic cells. Mol. Gen. Genet. 1997, 254, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Grant, C.M.; MacIver, F.H.; Dawes, I.W. Mitochondrial function is required for resistance to oxidative stress in the yeast Saccharomyces cerevisiae. FEBS Lett. 1997, 410, 219–222. [Google Scholar] [CrossRef]
- Kaniak-Golik, A.; Skoneczna, A. Mitochondria–nucleus network for genome stability. Free Radic Biol Med. 2015, 82, 73–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rendekova, J.; Ward, T.A.; Simonicova, L.; Thomas, P.H.; Nosek, J.; Tomaska, L.; McHugh, P.J.; Chovanec, M. Mgm101: A double-duty Rad52-like protein. Cell Cycle 2016, 23, 3169–3176. [Google Scholar] [CrossRef] [PubMed]
- Herlan, M.; Vogel, F.; Bornhovd, C.; Neupert, W.; Reichert, A.S. Processing of Mgm1 by the rhomboid-type protease Pcp1 is required for maintenance of mitochondrial morphology and of mitochondrial DNA. J. Biol. Chem. 2003, 278, 27781–27788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedman, T.; Kuusk, S.; Kivi, S.; Sedman, J. A DNA helicase required for maintenance of the functional mitochondrial genome in Saccharomyces cerevisiae. Mol. Cell Biol. 2000, 20, 1816–1824. [Google Scholar] [CrossRef] [Green Version]
- Van Meerloo, J.; Kaspers, G.J.; Cloos, J. Cell sensitivity assays: The MTT assay. Methods Mol. Biol. 2011, 731, 237–245. [Google Scholar] [PubMed]
- Xu, R.H.; Pelicano, H.; Zhou, Y.; Carew, J.S.; Feng, L.; Bhalla, K.N.; Keating, M.J.; Huang, P. Inhibition of glycolysis in cancer cells: A novel strategy to overcome drug resistance associated with mitochondrial respiratory defect and hypoxia. Cancer Res. 2005, 65, 613–621. [Google Scholar] [PubMed]
- Santos, J.H.; Mandavilli, B.S.; Van Houten, B. Measuring oxidative mtDNA damage and repair using quantitative PCR. Methods Mol. Biol. 2002, 197, 159–176. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Genotype | Source |
|---|---|---|
| W303-1A | MATa ade2-1 can1-100 ura3-1 his3-11,15 leu2-3, 112trp1-1 RAD5 | R. Rothstein |
| rho0 | W303-1A, rho0 | This work |
| BY4741 | MAT a, Δhis3; Δleu2; Δmet15; Δura3 | Euroscarf |
| Δmhr1 | BY4741, Δmhr1 | Euroscarf |
| Δctt1 | BY4741, Δctt1 | Euroscarf |
| Δpcp1 | BY4741, Δpcp1 | Euroscarf |
| Δgpx1 | BY4741, Δgpx1 | Euroscarf |
| Δccp1 | BY4741, Δccp1 | Euroscarf |
| Δhmi1 | BY4741, Δhmi1 | Euroscarf |
| Δsod2 | BY4741, Δsod2 | Euroscarf |
| Δmgm101 | BY4741, Δmgm101 | Euroscarf |
| Δpor1 | BY4741, Δpor1 | Euroscarf |
| Δrim1 | BY4741, Δrim1 | Euroscarf |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cal, M.; Matyjaszczyk, I.; Filik, K.; Ogórek, R.; Ko, Y.; Ułaszewski, S. Mitochondrial Function Are Disturbed in the Presence of the Anticancer Drug, 3-Bromopyruvate. Int. J. Mol. Sci. 2021, 22, 6640. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126640
Cal M, Matyjaszczyk I, Filik K, Ogórek R, Ko Y, Ułaszewski S. Mitochondrial Function Are Disturbed in the Presence of the Anticancer Drug, 3-Bromopyruvate. International Journal of Molecular Sciences. 2021; 22(12):6640. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126640
Chicago/Turabian StyleCal, Magdalena, Irwin Matyjaszczyk, Karolina Filik, Rafał Ogórek, Young Ko, and Stanisław Ułaszewski. 2021. "Mitochondrial Function Are Disturbed in the Presence of the Anticancer Drug, 3-Bromopyruvate" International Journal of Molecular Sciences 22, no. 12: 6640. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126640