
A Role for Xanthurenic Acid in the Control of Brain Dopaminergic Activity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
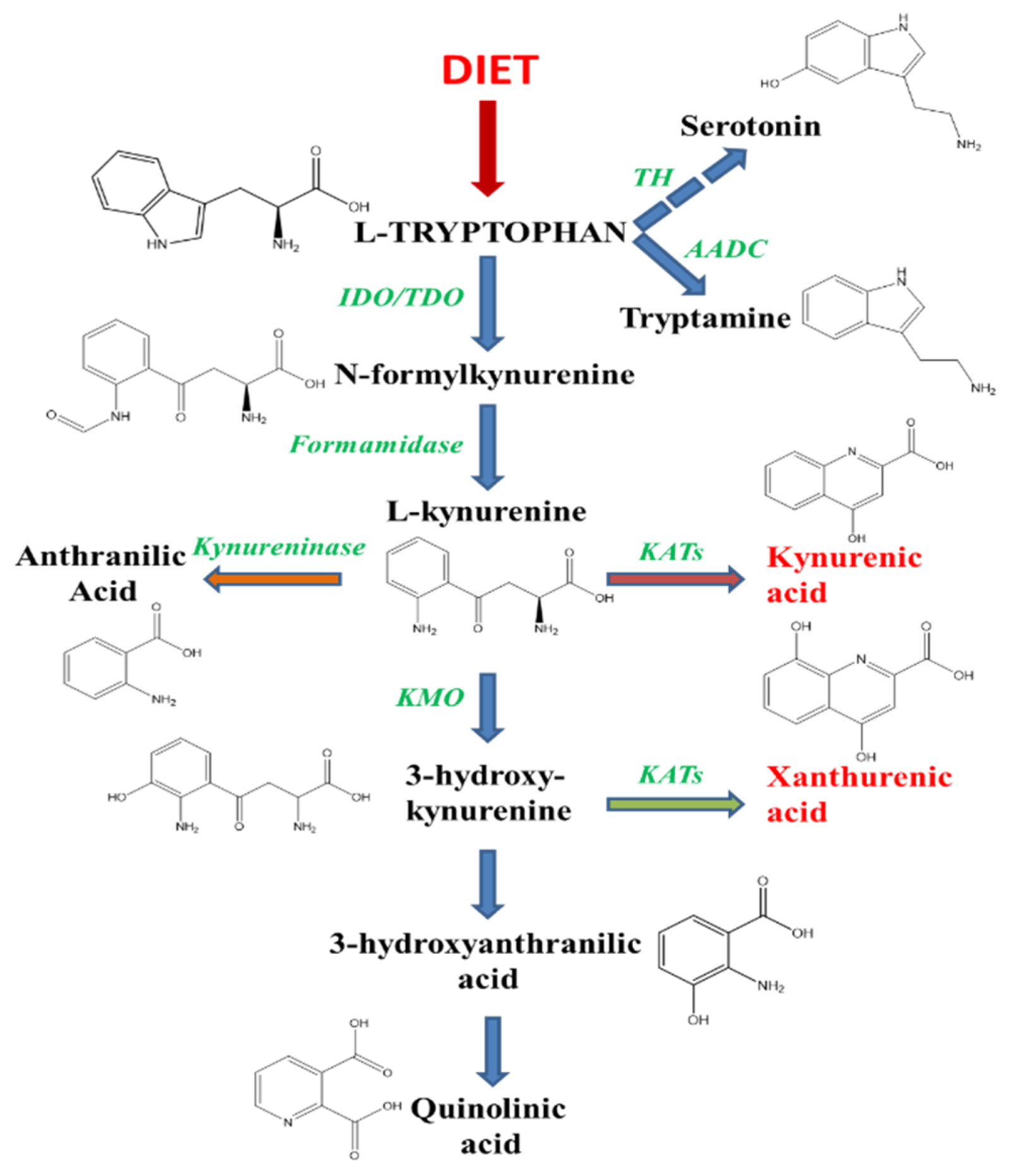
:1. Introduction
2. Results
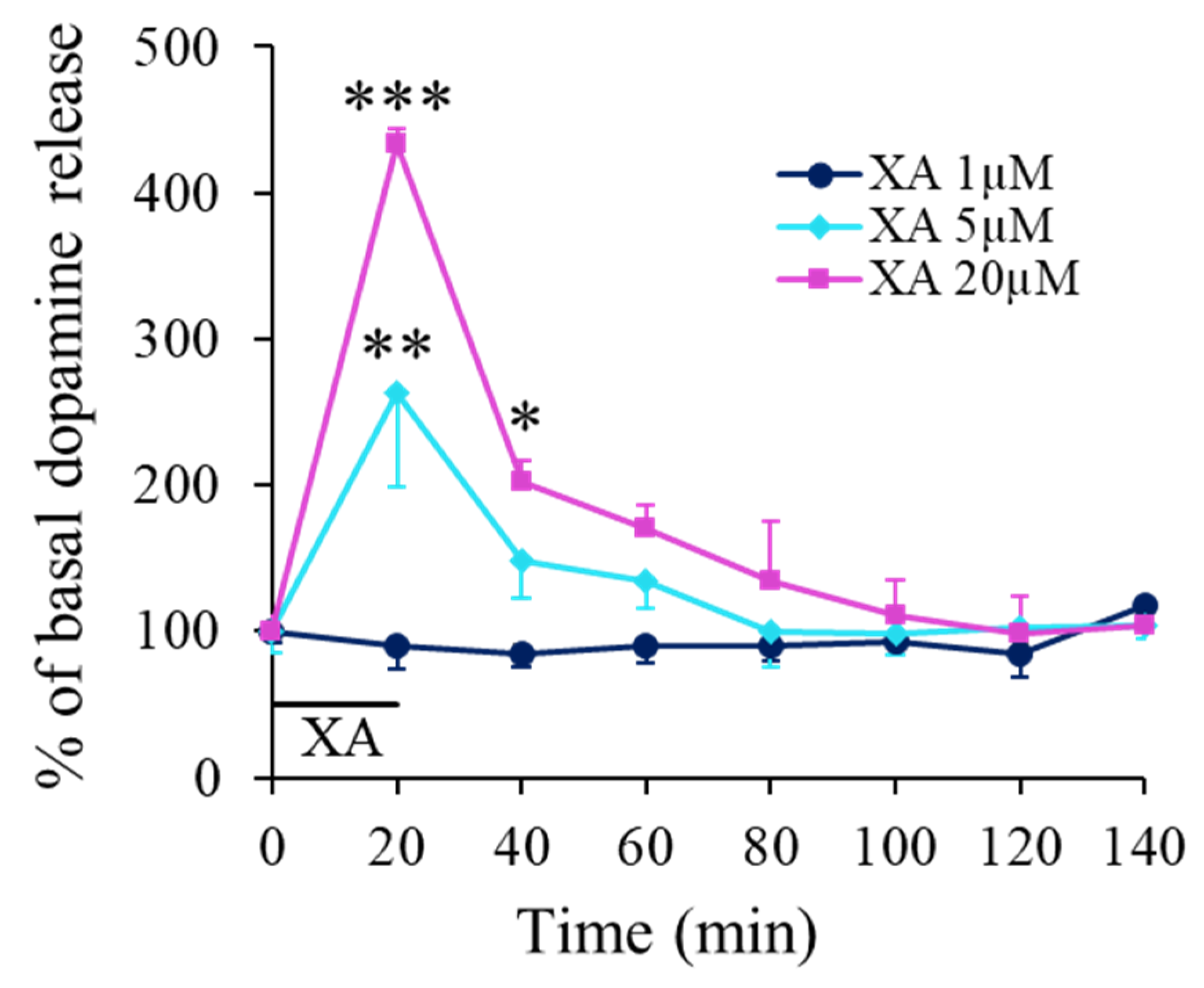
2.1. XA Retro-Dialysis Induces an Increase in Dopamine Extracellular Concentration in the Rat Prefrontal Cortex
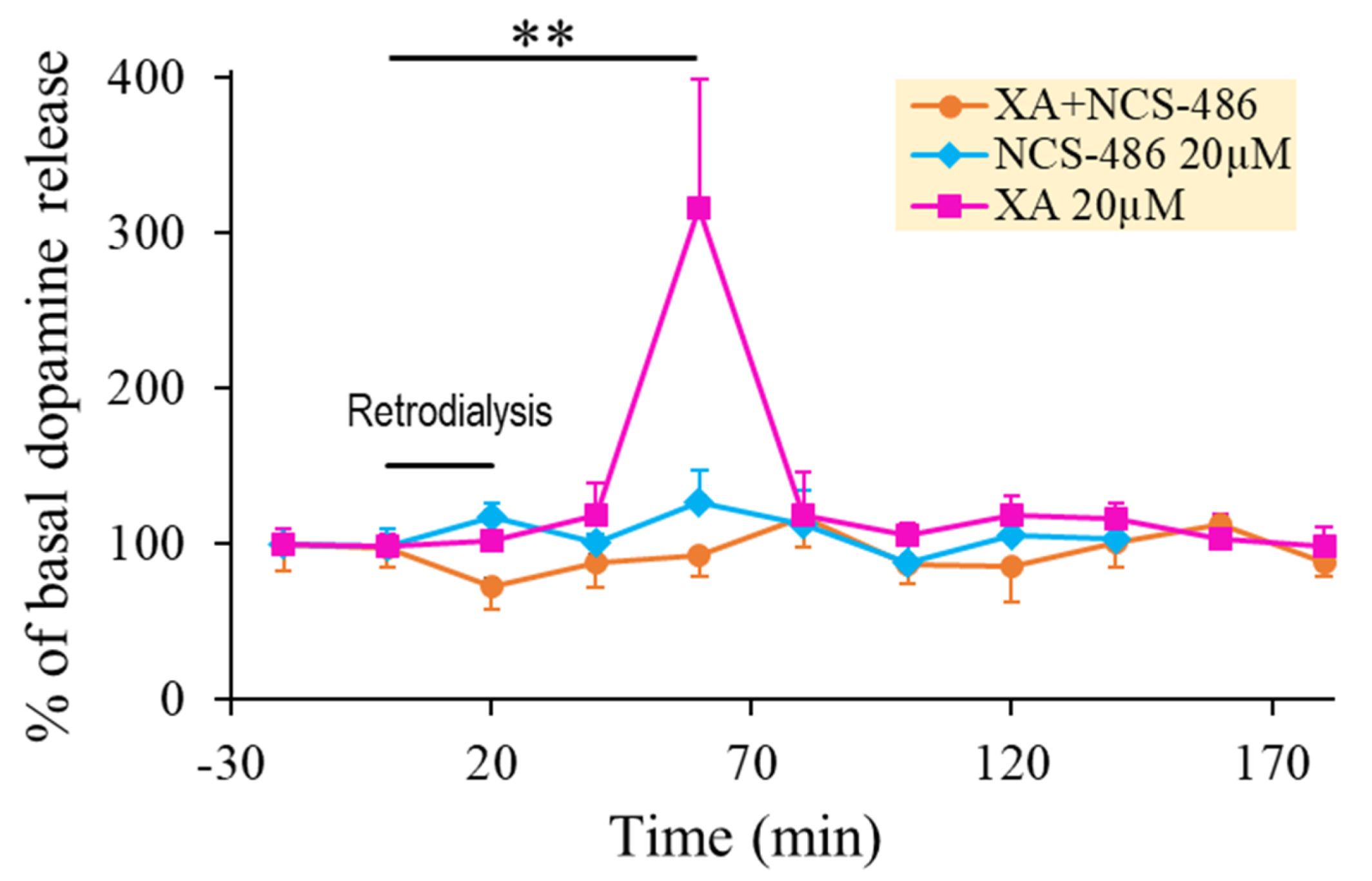
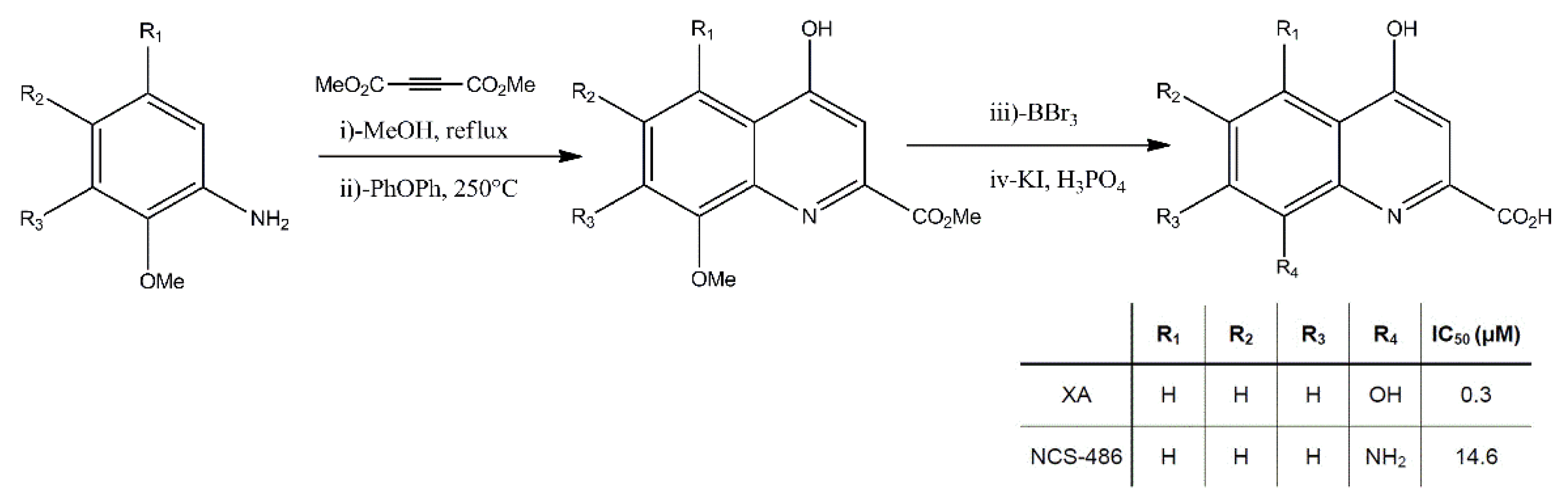
2.2. The Effect of XA Local Application on Dopamine Release in the Prefrontal Cortex Was Blocked by the Antagonist NCS-486
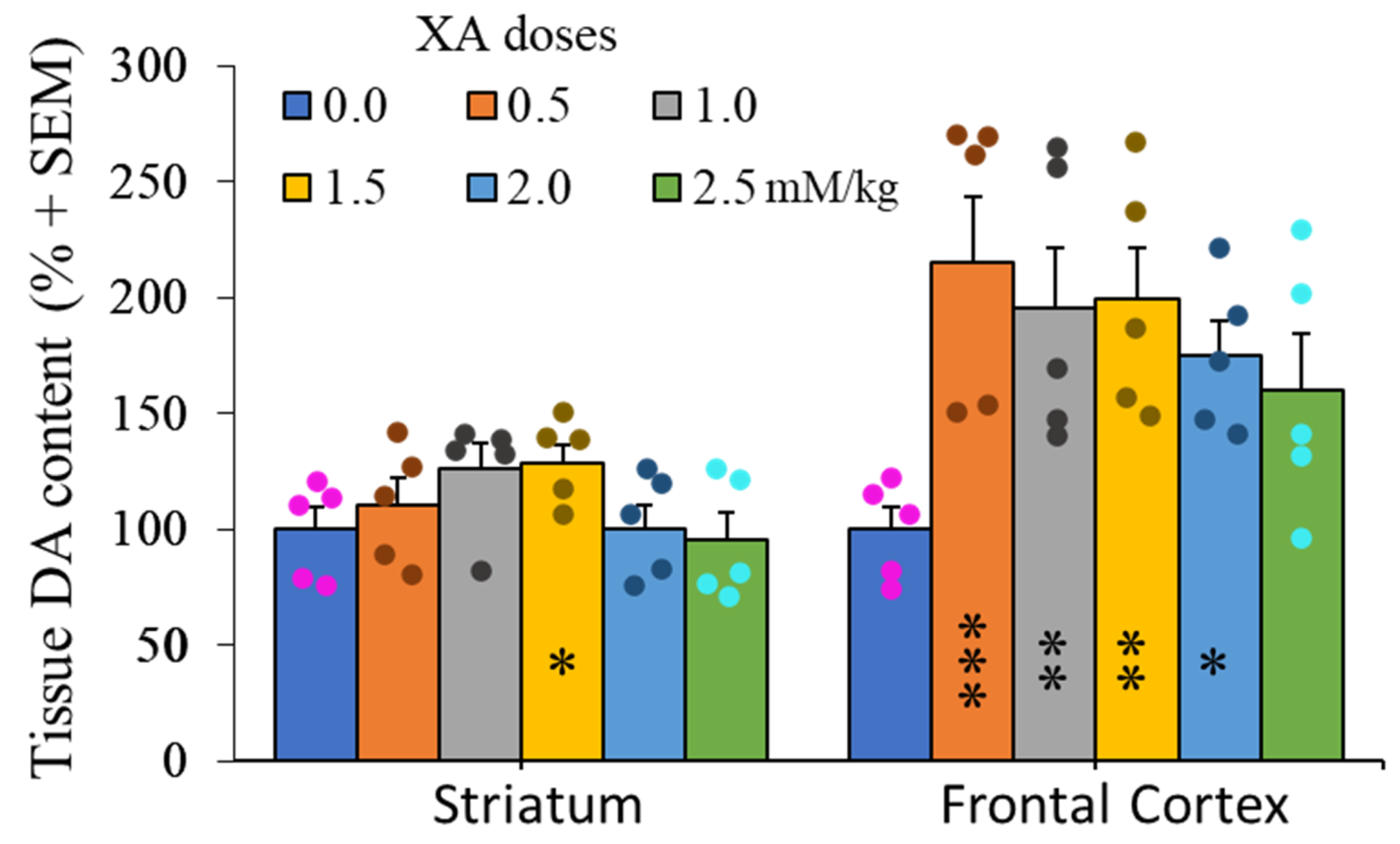
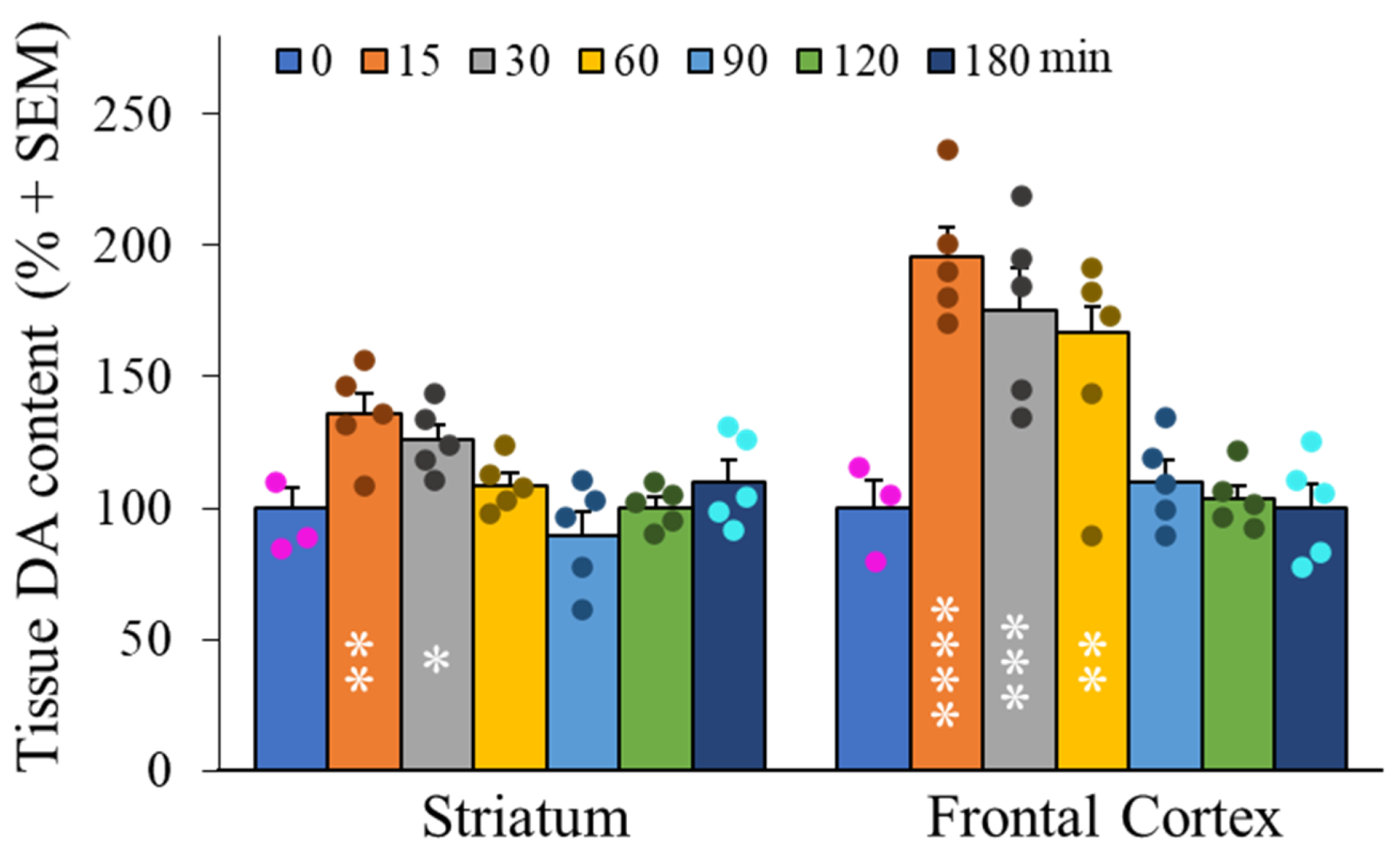
2.3. Doses-Effect of XA Administration on Frontal Cortex and Striatum Dopamine Contents
2.4. Kinetics of Dopamine Variation in the Frontal Cortex and Striatum after Peripheral Administration of XA
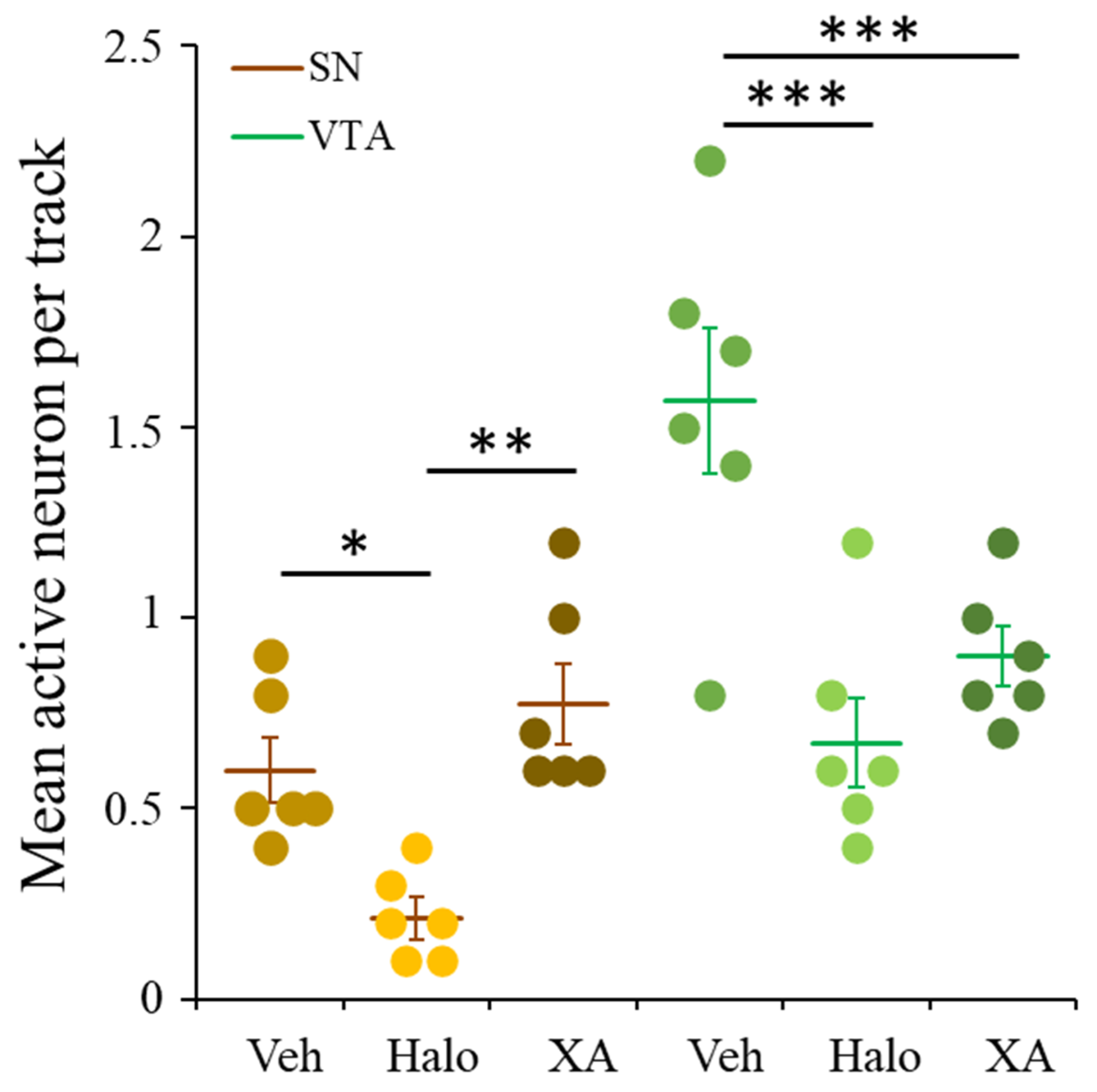
2.5. XA Repeated Treatment Reduced Spontaneous Firing in VTA but Not in SN
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Materials
4.3. Surgical Procedures
4.4. Microdialysis Procedures
4.5. Dopamine Analysis
4.6. In Vitro Recovery Experiments
4.7. Histology
4.8. Electrophysiological Recording
4.8.1. Animal Treatments
4.8.2. Surgical, Stereotaxic, and Recording Procedures
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Li, Y.; Hu, N.; Yang, D.; Oxenkrug, G.; Yang, Q. Regulating the balance between the kynurenine and serotonin pathways of tryptophan metabolism. FEBS J. 2017, 284, 948–966. [Google Scholar] [CrossRef] [Green Version]
- Vamos, E.; Pardutz, A.; Klivenyi, P.; Toldi, J.; Vecsei, L. The role of kynurenines in disorders of the central nervous system: Possibilities for neuroprotection. J. Neurol. Sci. 2009, 283, 21–27. [Google Scholar] [CrossRef]
- Schwarcz, R. Kynurenines and Glutamate: Multiple Links and Therapeutic Implications. Adv. Pharmacol. 2016, 76, 13–37. [Google Scholar]
- Stone, T.W.; Forrest, C.M.; Darlington, L.G. Kynurenine pathway inhibition as a therapeutic strategy for neuroprotection. FEBS J. 2012, 279, 1386–1397. [Google Scholar] [CrossRef] [PubMed]
- Sathyasaikumar, K.V.; Tararina, M.; Wu, H.Q.; Neale, S.A.; Weisz, F.; Salt, T.E.; Schwarcz, R. Xanthurenic Acid Formation from 3-Hydroxykynurenine in the Mammalian Brain: Neurochemical Characterization and Physiological Effects. Neuroscience 2017, 367, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarcz, R.; Stone, T.W. The kynurenine pathway and the brain: Challenges, controversies and promises. Neuropharmacology 2017, 112 Pt B, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Fujigaki, H.; Yamamoto, Y.; Saito, K. L-Tryptophan-kynurenine pathway enzymes are therapeutic target for neuropsychiatric diseases: Focus on cell type differences. Neuropharmacology 2017, 112 Pt B, 264–274. [Google Scholar] [CrossRef]
- Phillips, R.S.; Iradukunda, E.C.; Hughes, T.; Bowen, J.P. Modulation of Enzyme Activity in the Kynurenine Pathway by Kynurenine Monooxygenase Inhibition. Front. Mol. Biosci. 2019, 6, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhard, J.F., Jr. Pharmacological manipulation of brain kynurenine metabolism. Ann. N. Y. Acad. Sci. 2004, 1035, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Amaral, M.; Levy, C.; Heyes, D.J.; Lafite, P.; Outeiro, T.F.; Giorgini, F.; Leys, D.; Scrutton, N.S. Structural basis of kynurenine 3-monooxygenase inhibition. Nature 2013, 496, 382–385. [Google Scholar] [CrossRef] [PubMed]
- Nematollahi, A.; Sun, G.; Jayawickrama, G.S.; Church, W.B. Kynurenine Aminotransferase Isozyme Inhibitors: A Review. Int. J. Mol. Sci. 2016, 17, 946. [Google Scholar] [CrossRef]
- Rossi, F.; Miggiano, R.; Ferraris, D.M.; Rizzi, M. The Synthesis of Kynurenic Acid in Mammals: An Updated Kynurenine Aminotransferase Structural KATalogue. Front. Mol. Biosci. 2019, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Jayawickrama, G.S.; Sadig, R.R.; Sun, G.; Nematollahi, A.; Nadvi, N.A.; Hanrahan, J.R.; Gorrell, M.D.; Church, W.B. Kynurenine Aminotransferases and the Prospects of Inhibitors for the Treatment of Schizophrenia. Curr. Med. Chem. 2015, 22, 2902–2918. [Google Scholar] [CrossRef]
- Bohar, Z.; Toldi, J.; Fulop, F.; Vecsei, L. Changing the face of kynurenines and neurotoxicity: Therapeutic considerations. Int. J. Mol. Sci. 2015, 16, 9772–9793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tufvesson-Alm, M.; Schwieler, L.; Schwarcz, R.; Goiny, M.; Erhardt, S.; Engberg, G. Importance of kynurenine 3-monooxygenase for spontaneous firing and pharmacological responses of midbrain dopamine neurons: Relevance for schizophrenia. Neuropharmacology 2018, 138, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Schwieler, L.; Engberg, G.; Erhardt, S. Clozapine modulates midbrain dopamine neuron firing via interaction with the NMDA receptor complex. Synapse 2004, 52, 114–122. [Google Scholar] [CrossRef]
- Erhardt, S.; Schwieler, L.; Nilsson, L.; Linderholm, K.; Engberg, G. The kynurenic acid hypothesis of schizophrenia. Physiol. Behav. 2007, 92, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Wonodi, I.; Schwarcz, R. Cortical kynurenine pathway metabolism: A novel target for cognitive enhancement in Schizophrenia. Schizophr Bull. 2010, 36, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Olsson, S.K.; Andersson, A.S.; Linderholm, K.R.; Holtze, M.; Nilsson-Todd, L.K.; Schwieler, L.; Olsson, E.; Larsson, K.; Engberg, G.; Erhardt, S. Elevated levels of kynurenic acid change the dopaminergic response to amphetamine: Implications for schizophrenia. Int. J. Neuropsychopharmacol. 2009, 12, 501–512. [Google Scholar] [CrossRef] [Green Version]
- McCutcheon, R.A.; Abi-Dargham, A.; Howes, O.D. Schizophrenia, Dopamine and the Striatum: From Biology to Symptoms. Trends Neurosci. 2019, 42, 205–220. [Google Scholar] [CrossRef] [Green Version]
- Rassoulpour, A.; Wu, H.Q.; Ferre, S.; Schwarcz, R. Nanomolar concentrations of kynurenic acid reduce extracellular dopamine levels in the striatum. J. Neurochem. 2005, 93, 762–765. [Google Scholar] [CrossRef]
- Wu, H.Q.; Rassoulpour, A.; Schwarcz, R. Kynurenic acid leads, dopamine follows: A new case of volume transmission in the brain? J. Neural Transm. (Vienna) 2007, 114, 33–41. [Google Scholar] [CrossRef]
- Roussel, G.; Bessede, A.; Klein, C.; Maitre, M.; Mensah-Nyagan, A.G. Xanthurenic acid is localized in neurons in the central nervous system. Neuroscience 2016, 329, 226–238. [Google Scholar] [CrossRef]
- Balog, E.; Jenei, G.; Gellert, L.; Ono, E.; Vecsei, L.; Toldi, J.; Kis, Z. Species-specific neuronal localization of kynurenine aminotransferase-2 in the mouse cerebellum. Neurochem. Int. 2021, 142, 104920. [Google Scholar] [CrossRef]
- Gobaille, S.; Kemmel, V.; Brumaru, D.; Dugave, C.; Aunis, D.; Maitre, M. Xanthurenic acid distribution, transport, accumulation and release in the rat brain. J. Neurochem. 2008, 105, 982–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taleb, O.; Maammar, M.; Brumaru, D.; Bourguignon, J.J.; Schmitt, M.; Klein, C.; Kemmel, V.; Maitre, M.; Mensah-Nyagan, A.G. Xanthurenic acid binds to neuronal G-protein-coupled receptors that secondarily activate cationic channels in the cell line NCB-20. PLoS ONE 2012, 7, e48553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neale, S.A.; Copeland, C.S.; Uebele, V.N.; Thomson, F.J.; Salt, T.E. Modulation of hippocampal synaptic transmission by the kynurenine pathway member xanthurenic acid and other VGLUT inhibitors. Neuropsychopharmacology 2013, 38, 1060–1067. [Google Scholar] [CrossRef] [PubMed]
- Copeland, C.S.; Neale, S.A.; Salt, T.E. Actions of Xanthurenic acid, a putative endogenous Group II metabotropic glutamate receptor agonist, on sensory transmission in the thalamus. Neuropharmacology 2013, 66, 133–142. [Google Scholar] [CrossRef]
- Fazio, F.; Lionetto, L.; Curto, M.; Iacovelli, L.; Cavallari, M.; Zappulla, C.; Ulivieri, M.; Napoletano, F.; Capi, M.; Corigliano, V.; et al. Xanthurenic Acid Activates mGlu2/3 Metabotropic Glutamate Receptors and is a Potential Trait Marker for Schizophrenia. Sci. Rep. 2015, 5, 17799. [Google Scholar] [CrossRef] [Green Version]
- Curto, M.; Lionetto, L.; Fazio, F.; Corigliano, V.; Comparelli, A.; Ferracuti, S.; Simmaco, M.; Nicoletti, F.; Baldessarini, R.J. Serum xanthurenic acid levels: Reduced in subjects at ultra high risk for psychosis. Schizophr. Res. 2019, 208, 465–466. [Google Scholar] [CrossRef] [PubMed]
- Kanchanatawan, B.; Thika, S.; Sirivichayakul, S.; Carvalho, A.F.; Geffard, M.; Maes, M. In Schizophrenia, Depression, Anxiety, and Physiosomatic Symptoms Are Strongly Related to Psychotic Symptoms and Excitation, Impairments in Episodic Memory, and Increased Production of Neurotoxic Tryptophan Catabolites: A Multivariate and Machine Learning Study. Neurotox. Res. 2018, 33, 641–655. [Google Scholar] [PubMed]
- Jiang, H.; Leger, J.M.; Huc, I. Aromatic delta-peptides. J. Am. Chem. Soc. 2003, 125, 3448–3449. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C.R.; Emson, P.C. AChE-stained horizontal sections of the rat brain in stereotaxic coordinates. J. Neurosci. Methods. 1980, 3, 129–149. [Google Scholar] [CrossRef]
- Bunney, B.S.; Aghajanian, G.K.; Roth, R.H. Comparison of effects of L-dopa, amphetamine and apomorphine on firing rate of rat dopaminergic neurones. Nat. New Biol. 1973, 245, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Kegel, M.E.; Bhat, M.; Skogh, E.; Samuelsson, M.; Lundberg, K.; Dahl, M.L.; Sellgren, C.; Schwieler, L.; Engberg, G.; Schuppe-Koistinen, I.; et al. Imbalanced kynurenine pathway in schizophrenia. Int. J. Tryptophan Res. 2014, 7, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Erhardt, S.; Pocivavsek, A.; Repici, M.; Liu, X.C.; Imbeault, S.; Maddison, D.C.; Thomas, M.A.R.; Smalley, J.L.; Larsson, M.K.; Muchowski, P.J.; et al. Adaptive and Behavioral Changes in Kynurenine 3-Monooxygenase Knockout Mice: Relevance to Psychotic Disorders. Biol. Psychiatry 2017, 82, 756–765. [Google Scholar] [CrossRef] [Green Version]
- Kubicova, L.; Chobot, V. Potential of kynurenine metabolites in drug development against neurodegenerative diseases. Neural Regen. Res. 2021, 16, 308–309. [Google Scholar]
- Fazio, F.; Lionetto, L.; Curto, M.; Iacovelli, L.; Copeland, C.S.; Neale, S.A.; Bruno, V.; Battaglia, G.; Salt, T.E.; Nicoletti, F. Cinnabarinic acid and xanthurenic acid: Two kynurenine metabolites that interact with metabotropic glutamate receptors. Neuropharmacology 2017, 112 Pt B, 365–372. [Google Scholar] [CrossRef]
- Neale, S.A.; Copeland, C.S.; Salt, T.E. Effect of VGLUT inhibitors on glutamatergic synaptic transmission in the rodent hippocampus and prefrontal cortex. Neurochem. Int. 2014, 73, 159–165. [Google Scholar] [CrossRef]
- Gimenez-Gomez, P.; Perez-Hernandez, M.; Gutierrez-Lopez, M.D.; Vidal, R.; Abuin-Martinez, C.; O’Shea, E.; Colado, M.I. Increasing kynurenine brain levels reduces ethanol consumption in mice by inhibiting dopamine release in nucleus accumbens. Neuropharmacology 2018, 135, 581–591. [Google Scholar] [CrossRef]
- Badawy, A.A. Kynurenine Pathway of Tryptophan Metabolism: Regulatory and Functional Aspects. Int. J. Tryptophan Res. 2017, 10, 1178646917691938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heredi, J.; Berko, A.M.; Jankovics, F.; Iwamori, T.; Iwamori, N.; Ono, E.; Horvath, S.; Kis, Z.; Toldi, J.; Vecsei, L.; et al. Astrocytic and neuronal localization of kynurenine aminotransferase-2 in the adult mouse brain. Brain Struct. Funct. 2017, 222, 1663–1672. [Google Scholar] [CrossRef] [Green Version]
- Chuhma, N.; Mingote, S.; Kalmbach, A.; Yetnikoff, L.; Rayport, S. Heterogeneity in Dopamine Neuron Synaptic Actions Across the Striatum and Its Relevance for Schizophrenia. Biol. Psychiatry 2017, 81, 43–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gainetdinov, R.R.; Hoener, M.C.; Berry, M.D. Trace Amines and Their Receptors. Pharmacol. Rev. 2018, 70, 549–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taleb, O.; Maammar, M.; Klein, C.; Maitre, M.; Mensah-Nyagan, A.G. A Role for Xanthurenic Acid in the Control of Brain Dopaminergic Activity. Int. J. Mol. Sci. 2021, 22, 6974. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136974
Taleb O, Maammar M, Klein C, Maitre M, Mensah-Nyagan AG. A Role for Xanthurenic Acid in the Control of Brain Dopaminergic Activity. International Journal of Molecular Sciences. 2021; 22(13):6974. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136974
Chicago/Turabian StyleTaleb, Omar, Mohammed Maammar, Christian Klein, Michel Maitre, and Ayikoe Guy Mensah-Nyagan. 2021. "A Role for Xanthurenic Acid in the Control of Brain Dopaminergic Activity" International Journal of Molecular Sciences 22, no. 13: 6974. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22136974