
Seleno-Functionalization of Quercetin Improves the Non-Covalent Inhibition of Mpro and Its Antiviral Activity in Cells against SARS-CoV-2

 , , ,
, , ,  , , , ,
, , , ,  ,
, 
 , , , and
, , , and  add
Show full author list
add
Show full author list
Abstract
:1. Introduction
2. Results and Discussion
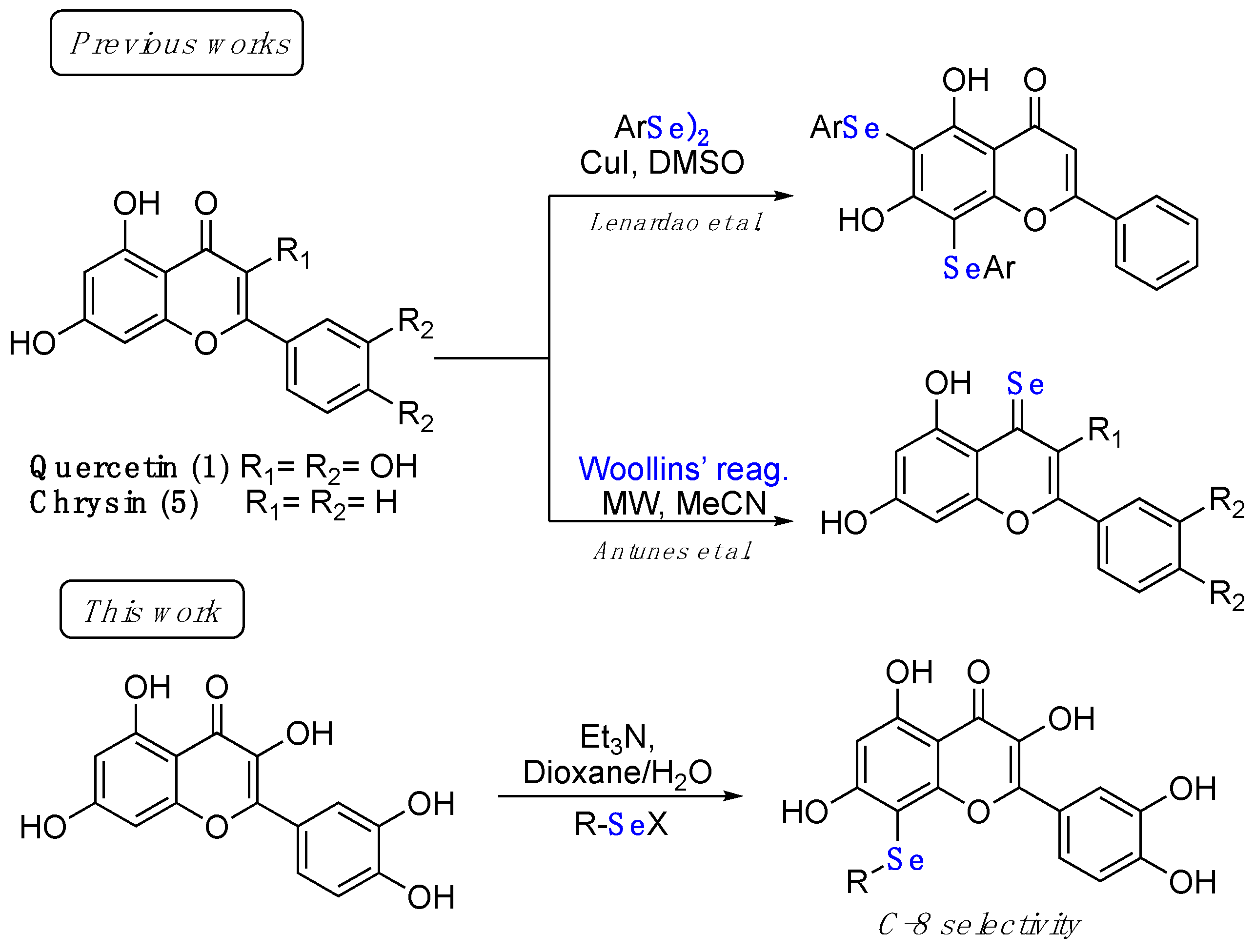
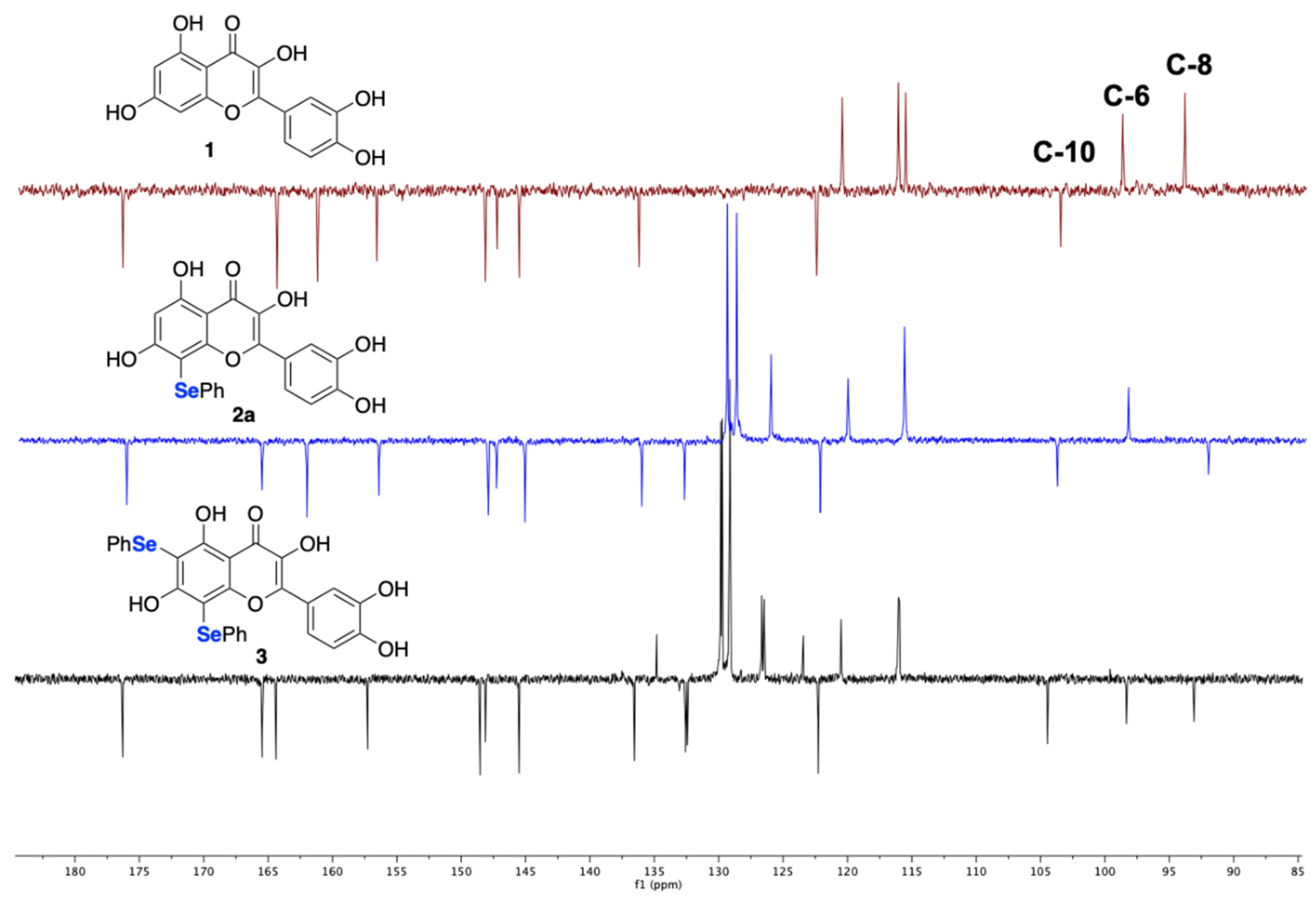
2.1. Synthesis
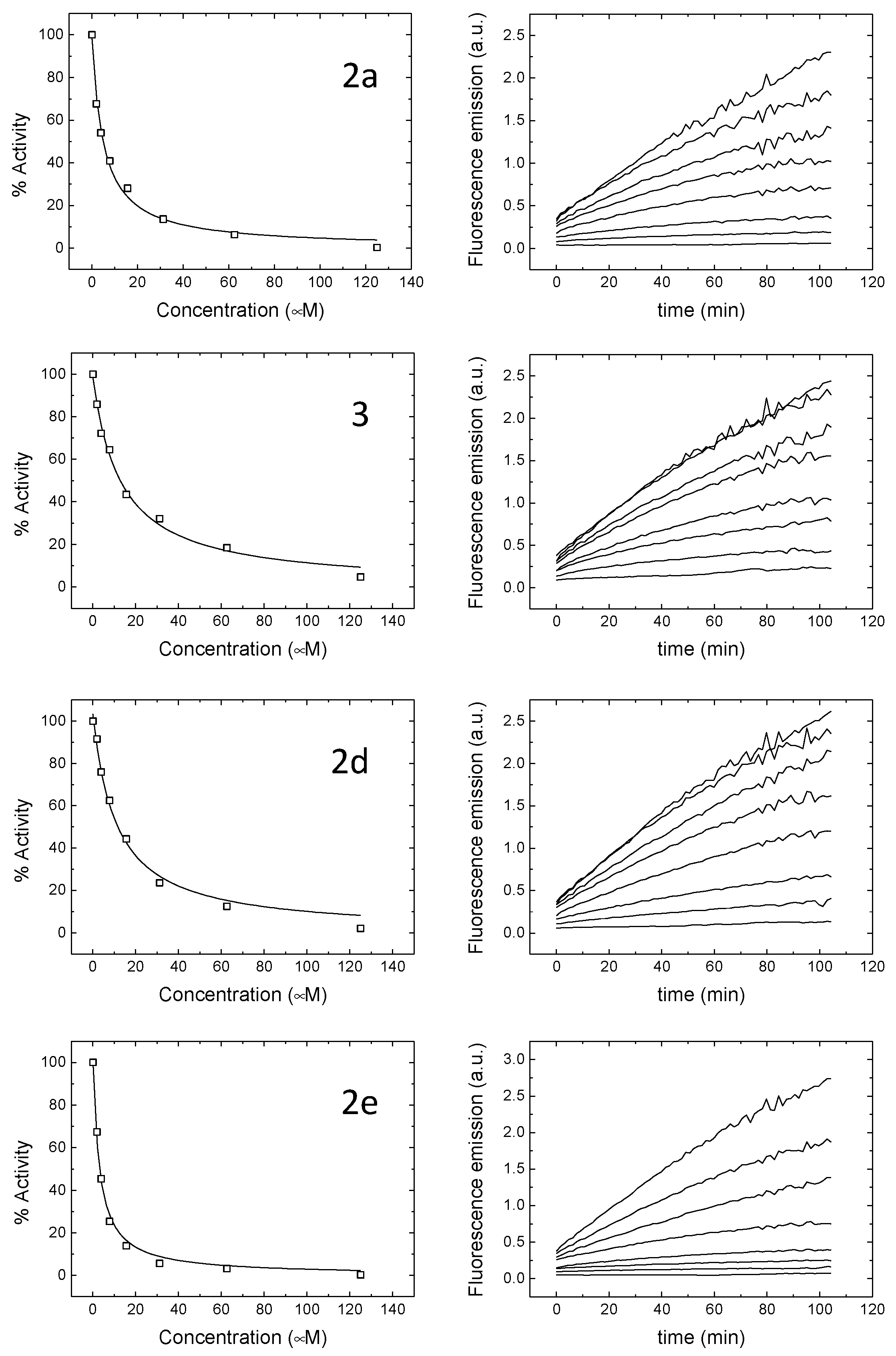
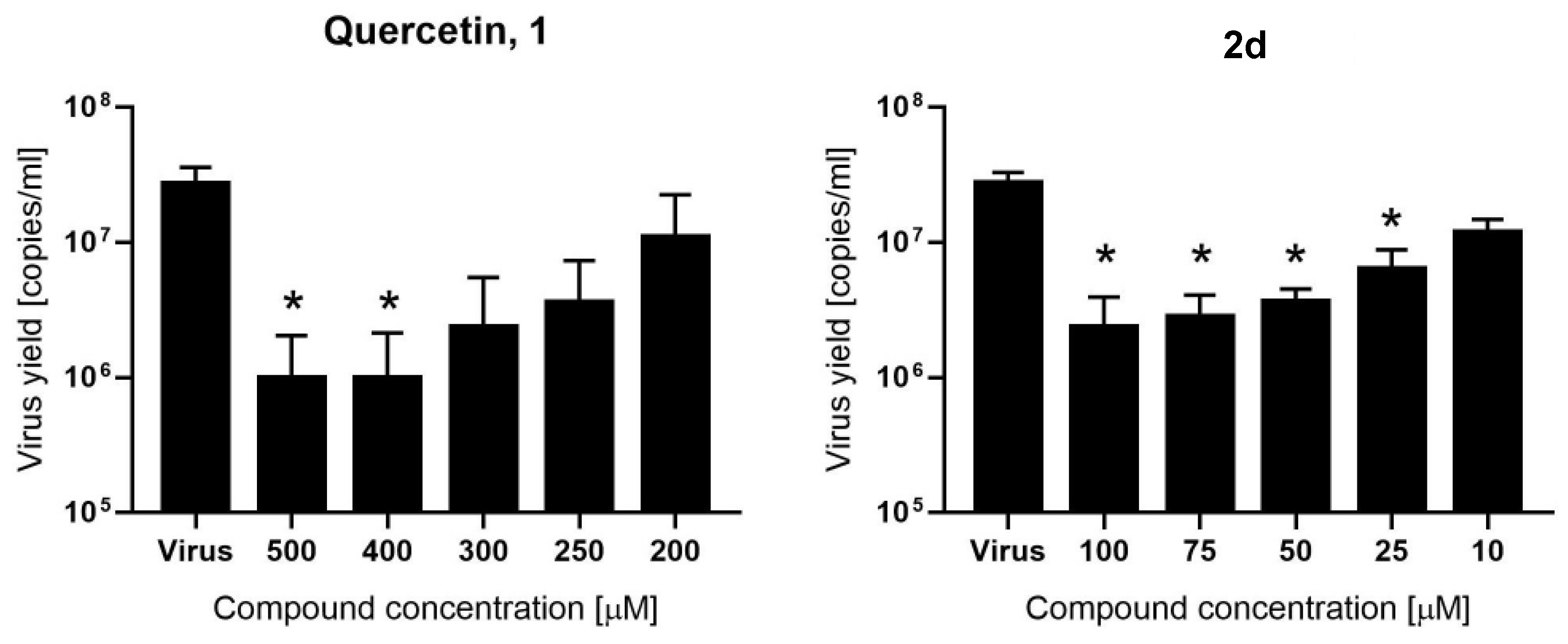
2.2. Evaluation of Mpro Inhibition and Antiviral Assay
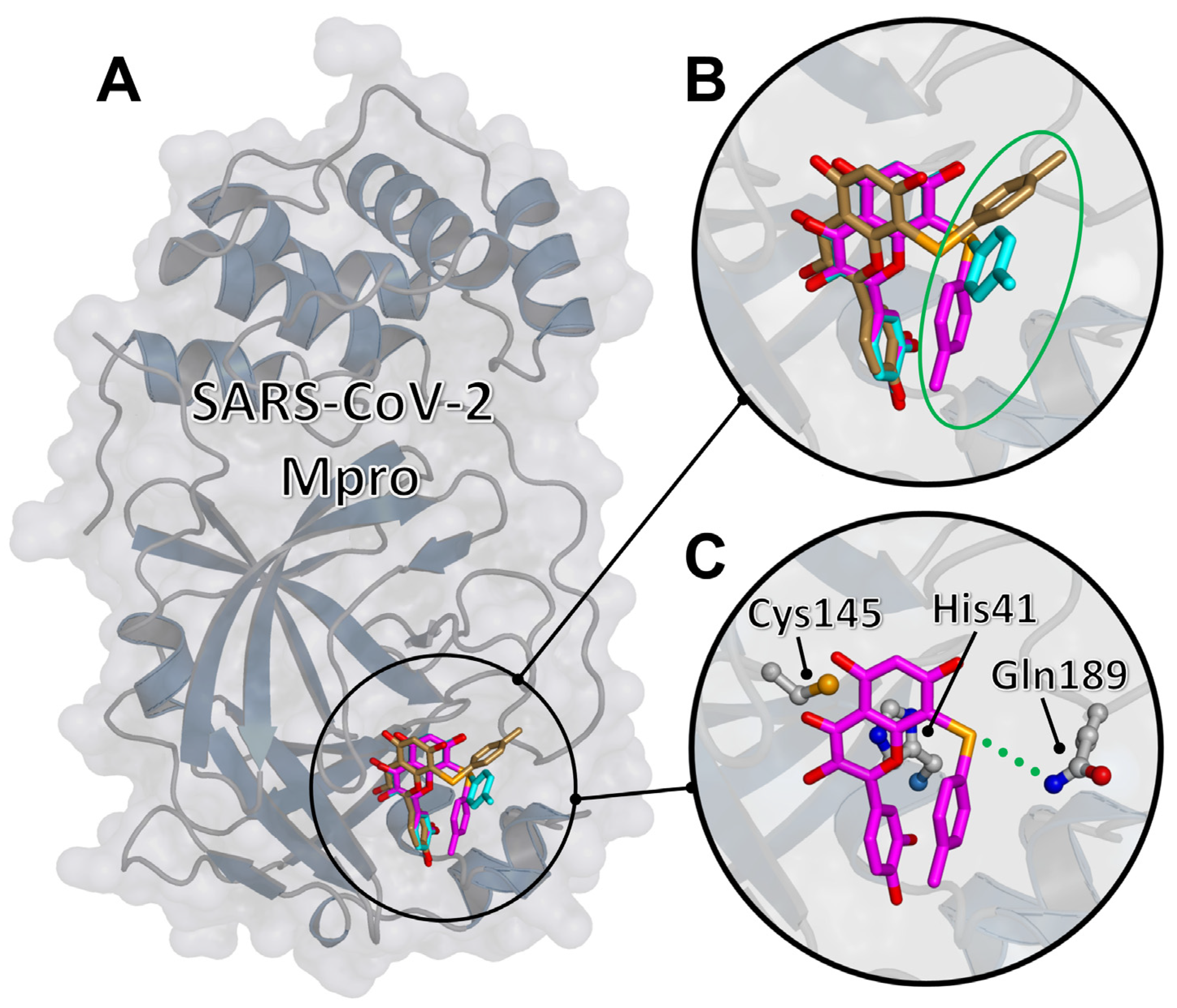
2.3. Molecular Docking of the Representative Compound 2d to SARS-CoV-2 Mpro
3. Materials and Methods
3.1. Synthesis: General Remarks
3.1.1. General Procedure for the In Situ Formation of RSeCl (5b–f)
3.1.2. General Procedure for the Seleno-Functionalization of 1
3.1.3. Physical Data
3.2. SARS-CoV-2 Mpro Expression and Purification
3.3. SARS-CoV-2 Mpro Proteolytic Activity Assay
3.4. SARS-CoV-2 Mpro Inhibition Assay
3.5. Molecular Simulations
3.6. Cells and Viruses
3.7. Evaluation of Viral Infection
3.8. Isolation of Nucleic Acids, Reverse Transcription, and Quantitative PCR
3.9. Cell Viability Assay
3.10. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Hirano, R.; Sasamoto, W.; Matsumoto, A.; Itakura, H.; Igarashi, O.; Kondo, K. Antioxidant Ability of Various Flavonoids against DPPH Radicals and LDL Oxidation. J. Nutr. Sci. Vitaminol. 2001, 47, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Yeh, S.-L.; Wang, W.-Y.; Huang, C.-H.; Hu, M.-L. Pro-oxidative effect of β-carotene and the interaction with flavonoids on UVA-induced DNA strand breaks in mouse fibroblast C3H10T1/2 cells. J. Nutr. Biochem. 2005, 16, 729–735. [Google Scholar] [CrossRef]
- Pietta, P.-G. Flavonoids as Antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3289–3303. [Google Scholar] [CrossRef]
- Lenardão, E.J.; Santi, C.; Sancineto, L. New Frontiers in Organoselenium Compounds; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-92404-5. [Google Scholar]
- Nagata, H.; Takekoshi, S.; Takagi, T.; Honma, T.; Watanabe, K. Antioxidative action of flavonoids, quercetin and catechin, mediated by the activation of glutathione peroxidase. Tokai J. Exp. Clin. Med. 1999, 24, 1–11. [Google Scholar]
- Andersen, K.G.; Rambaut, A.; Lipkin, W.I.; Holmes, E.C.; Garry, R.F. The proximal origin of SARS-CoV-2. Nat. Med. 2020, 26, 450–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of Mpro from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Pacuła, A.J.; Mangiavacchi, F.; Sancineto, L.; Lenardão, E.; Ścianowski, J.; Santi, C. An Update on “Selenium Containing Compounds from Poison to Drug Candidates: A Review on the GPx-like Activity”. Curr. Chem. Biol. 2016, 9, 97–112. [Google Scholar] [CrossRef]
- Parnham, M.J.; Sies, H. The early research and development of ebselen. Biochem. Pharmacol. 2013, 86, 1248–1253. [Google Scholar] [CrossRef] [PubMed]
- Abian, O.; Ortega-Alarcon, D.; Jimenez-Alesanco, A.; Ceballos-Laita, L.; Vega, S.; Reyburn, H.T.; Rizzuti, B.; Velazquez-Campoy, A. Structural stability of SARS-CoV-2 3CLpro and identification of quercetin as an inhibitor by experimental screening. Int. J. Biol. Macromol. 2020, 164, 1693–1703. [Google Scholar] [CrossRef]
- Rizzuti, B.; Grande, F.; Conforti, F.; Jimenez-Alesanco, A.; Ceballos-Laita, L.; Ortega-Alarcon, D.; Vega, S.; Reyburn, H.T.; Abian, O.; Velazquez-Campoy, A. Rutin Is a Low Micromolar Inhibitor of SARS-CoV-2 Main Protease 3CLpro: Implications for Drug Design of Quercetin Analogs. Biomedicines 2021, 9, 375. [Google Scholar] [CrossRef]
- Sancineto, L.; Mariotti, A.; Bagnoli, L.; Marini, F.; Desantis, J.; Iraci, N.; Santi, C.; Pannecouque, C.; Tabarrini, O. Design and Synthesis of DiselenoBisBenzamides (DISeBAs) as Nucleocapsid Protein 7 (NCp7) Inhibitors with anti-HIV Activity. J. Med. Chem. 2015, 58, 9601–9614. [Google Scholar] [CrossRef]
- Sancineto, L.; Iraci, N.; Tabarrini, O.; Santi, C. NCp7: Targeting a multitasking protein for next-generation anti-HIV drug development part 1: Covalent inhibitors. Drug Discov. Today 2018, 23, 260–271. [Google Scholar] [CrossRef]
- Jastrzebska, I.; Mellea, S.; Salerno, V.; Grzes, P.A.; Siergiejczyk, L.; Niemirowicz-Laskowska, K.; Bucki, R.; Monti, B.; Santi, C. PhSeZnCl in the Synthesis of Steroidal β-Hydroxy-Phenylselenides Having Antibacterial Activity. Int. J. Mol. Sci. 2019, 20, 2121. [Google Scholar] [CrossRef] [Green Version]
- Sancineto, L.; Piccioni, M.; De Marco, S.; Pagiotti, R.; Nascimento, V.; Braga, A.L.; Santi, C.; Pietrella, D. Diphenyl diselenide derivatives inhibit microbial biofilm formation involved in wound infection. BMC Microbiol. 2016, 16, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, S.F.; Padilha, N.B.; Thurow, S.; Roehrs, J.A.; Savegnago, L.; de Souza, M.N.; Fronza, M.G.; Collares, T.; Buss, J.; Seixas, F.K.; et al. Ultrasound-promoted copper-catalyzed synthesis of bis-arylselanyl chrysin derivatives with boosted antioxidant and anticancer activities. Ultrason. Sonochem. 2017, 39, 827–836. [Google Scholar] [CrossRef]
- Martins, I.L.; Charneira, C.; Gandin, V.; Ferreira Da Silva, J.L.; Justino, G.C.; Telo, J.P.; Vieira, A.J.S.C.; Marzano, C.; Antunes, A.M.M. Selenium-containing chrysin and quercetin derivatives: Attractive scaffolds for cancer therapy. J. Med. Chem. 2015, 58, 4250–4265. [Google Scholar] [CrossRef]
- Akolli, A.; Moretti, S.; Mangiavacchi, F.; Monti, B.; Santi, C. Mild and Green Protocol for Selective Deuteration of Quercetin-3-ORutinoside (Rutin) Under Aqueous Basic Conditions. Curr. Green Chem. 2019, 6, 147–151. [Google Scholar] [CrossRef]
- Sancineto, L.; Palomba, M.; Bagnoli, L.; Marini, F.; Santi, C. Advances in Electrophilic Organochalcogen Reagents. Curr. Org. Chem. 2015, 20, 122–135. [Google Scholar] [CrossRef]
- Wawer, I.; Zielinska, A. 13C-CP-MAS-NMR studies of flavonoids. I. Solid-state conformation of quercetin, quercetin 5′-sulphonic acid and some simple polyphenols. Solid State Nucl. Magn. Reson. 1997. [Google Scholar] [CrossRef]
- Krasowska, D.; Begini, F.; Santi, C.; Mangiavacchi, F.; Drabowicz, J.; Sancineto, L. Ultrasound-assisted synthesis of alkali metals diselenides (M2Se2) and their application for the gram-scale preparation of 2,2’-diselenobis(benzoic acid). Arkivoc 2019, 2019, 24–37. [Google Scholar] [CrossRef]
- Sharpless, K.B.; Young, M.W. Olefin synthesis. Rate enhancement of the elimination of alkyl aryl selenoxides by electron-withdrawing substituents. J. Org. Chem. 1975, 40, 947–949. [Google Scholar] [CrossRef]
- Klayman, D.L.; Griffin, T.S. Reaction of selenium with sodium borohydride in protic solvents. A Facile Method for the introduction of selenium into organic molecules. J. Am. Chem. Soc. 1973, 95, 197–199. [Google Scholar] [CrossRef]
- Fonseca, S.F.; Lima, D.B.; Alves, D.; Jacob, R.G.; Perin, G.; Lenardão, E.J.; Savegnago, L. Synthesis, characterization and antioxidant activity of organoselenium and organotellurium compound derivatives of chrysin. New J. Chem. 2015, 39, 3043–3050. [Google Scholar] [CrossRef]
- Ba, L.A.; Döring, M.; Jamier, V.; Jacob, C. Tellurium: An element with great biological potency and potential. Org. Biomol. Chem. 2010, 8, 4203. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Mishra, K.K.; Singh, S.K.; Ghosh, P.; Ghosh, D.; Das, A. The nature of selenium hydrogen bonding: Gas phase spectroscopy and quantum chemistry calculations. Phys. Chem. Chem. Phys. 2017, 19, 24179–24187. [Google Scholar] [CrossRef]
- Kuo, C.-J.; Chi, Y.-H.; Hsu, J.T.-A.; Liang, P.-H. Characterization of SARS main protease and inhibitor assay using a fluorogenic substrate. Biochem. Biophys. Res. Commun. 2004, 318, 862–867. [Google Scholar] [CrossRef]
- Paketurytė, V.; Petrauskas, V.; Zubrienė, A.; Abian, O.; Bastos, M.; Chen, W.-Y.; Moreno, M.J.; Krainer, G.; Linkuvienė, V.; Sedivy, A.; et al. Uncertainty in protein–ligand binding constants: Asymmetric confidence intervals versus standard errors. Eur. Biophys. J. 2021. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009. [Google Scholar] [CrossRef] [Green Version]
- Rappe, A.K.; Casewit, C.J.; Colwell, K.S.; Goddard, W.A.; Skiff, W.M. UFF, a full periodic table force field for molecular mechanics and molecular dynamics simulations. J. Am. Chem. Soc. 1992, 114, 10024–10035. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyykkö, P.; Atsumi, M. Molecular Single-Bond Covalent Radii for Elements 1-118. Chem. A Eur. J. 2009, 15, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Santofimia-Castaño, P.; Rizzuti, B.; Pey, Á.L.; Soubeyran, P.; Vidal, M.; Urrutia, R.; Iovanna, J.L.; Neira, J.L. Intrinsically disordered chromatin protein NUPR1 binds to the C-terminal region of Polycomb RING1B. Proc. Natl. Acad. Sci. USA 2017, 114, E6332–E6341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | ||||||
| Entry | Electrophile (E) | Base (B) | 1:E:B | Time (h) | T (°C) | Yield % (2a:3) |
| 1 | PhSeCl | Tris | 1:2:5 | 72 | rt | 18 (18:traces) |
| 2 | PhSeOTf a | Tris | 1:2:5 | 24 | rt | 11 (8:3) |
| 3 | NPSeP | Tris | 1:2:5 | 4 | rt | 47 (22:25) |
| 4 | PhSeCl | Tris | 1:2:5 | 4 | 80 | 11 (11:trace) |
| 5 | NPSeP | Tris | 1:2:5 | 4 | 80 | 35 (10:25) |
| 6 b | PhSeCl | Tris | 1:2:5 | 2 | rt | 33 (33:trace) |
| 7 b | NPSeP | Tris | 1:2:5 | 1 | rt | 81 (30:51) |
| 8 b | PhSeCl | Tris | 1:2:10 | 2 | rt | 22 (22:trace) |
| 9 b | PhSeCl | Et3N | 1:2:5 | 2 | rt | 66 (55:11) |
| 10 b | PhSeCl | Arginine c | 1:2:5 | 2 | rt | 44 (44:trace) |
| 11 | PhSeCl | NaOH | 1:2:2 | 6 | rt | 9 (9:0) |
 | ||
|---|---|---|
| Entry | Product | Yield (%) |
| 1 |  | 58 |
| 2 |  | 48 |
| 3 |  | 46 |
| 4 |  | 32 |
| 5 |  | 21 |
| Ki (μM) | CI95,Ki | IC50 | CI95,IC50 | |
|---|---|---|---|---|
| Quercetin 1 | 7.4 | [5.8, 9.5] | 21 | [17, 28] |
| 2a | 1.8 | [1.4, 2.2] | 5.1 | [4.1, 6.3] |
| 2b | ND | ND | ||
| 2c | 8.6 | [6.9, 11] | 24 | [20, 31] |
| 2d | 3.8 | [3.0, 5.0] | 11 | [8.5, 14.2] |
| 2e | 1.1 | [0.85, 1,3] | 3.0 | [2.4, 3.9] |
| 3 | 4.6 | [3.7, 5.8] | 13 | [11, 16] |
| 6 | ND | ND | ||
| 7a | ND | ND | ||
| 8 | 1.1 | [0.80, 1.6] | 3.3 | [2.3, 4.8] |
| 9 | 0.77 | [0.57, 1.0] | 2.2 | [1.7, 3.2] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mangiavacchi, F.; Botwina, P.; Menichetti, E.; Bagnoli, L.; Rosati, O.; Marini, F.; Fonseca, S.F.; Abenante, L.; Alves, D.; Dabrowska, A.; et al. Seleno-Functionalization of Quercetin Improves the Non-Covalent Inhibition of Mpro and Its Antiviral Activity in Cells against SARS-CoV-2. Int. J. Mol. Sci. 2021, 22, 7048. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137048
Mangiavacchi F, Botwina P, Menichetti E, Bagnoli L, Rosati O, Marini F, Fonseca SF, Abenante L, Alves D, Dabrowska A, et al. Seleno-Functionalization of Quercetin Improves the Non-Covalent Inhibition of Mpro and Its Antiviral Activity in Cells against SARS-CoV-2. International Journal of Molecular Sciences. 2021; 22(13):7048. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137048
Chicago/Turabian StyleMangiavacchi, Francesca, Pawel Botwina, Elena Menichetti, Luana Bagnoli, Ornelio Rosati, Francesca Marini, Sérgio F. Fonseca, Laura Abenante, Diego Alves, Agnieszka Dabrowska, and et al. 2021. "Seleno-Functionalization of Quercetin Improves the Non-Covalent Inhibition of Mpro and Its Antiviral Activity in Cells against SARS-CoV-2" International Journal of Molecular Sciences 22, no. 13: 7048. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137048