
Differential Effects of Halofuginone Enantiomers on Muscle Fibrosis and Histopathology in Duchenne Muscular Dystrophy

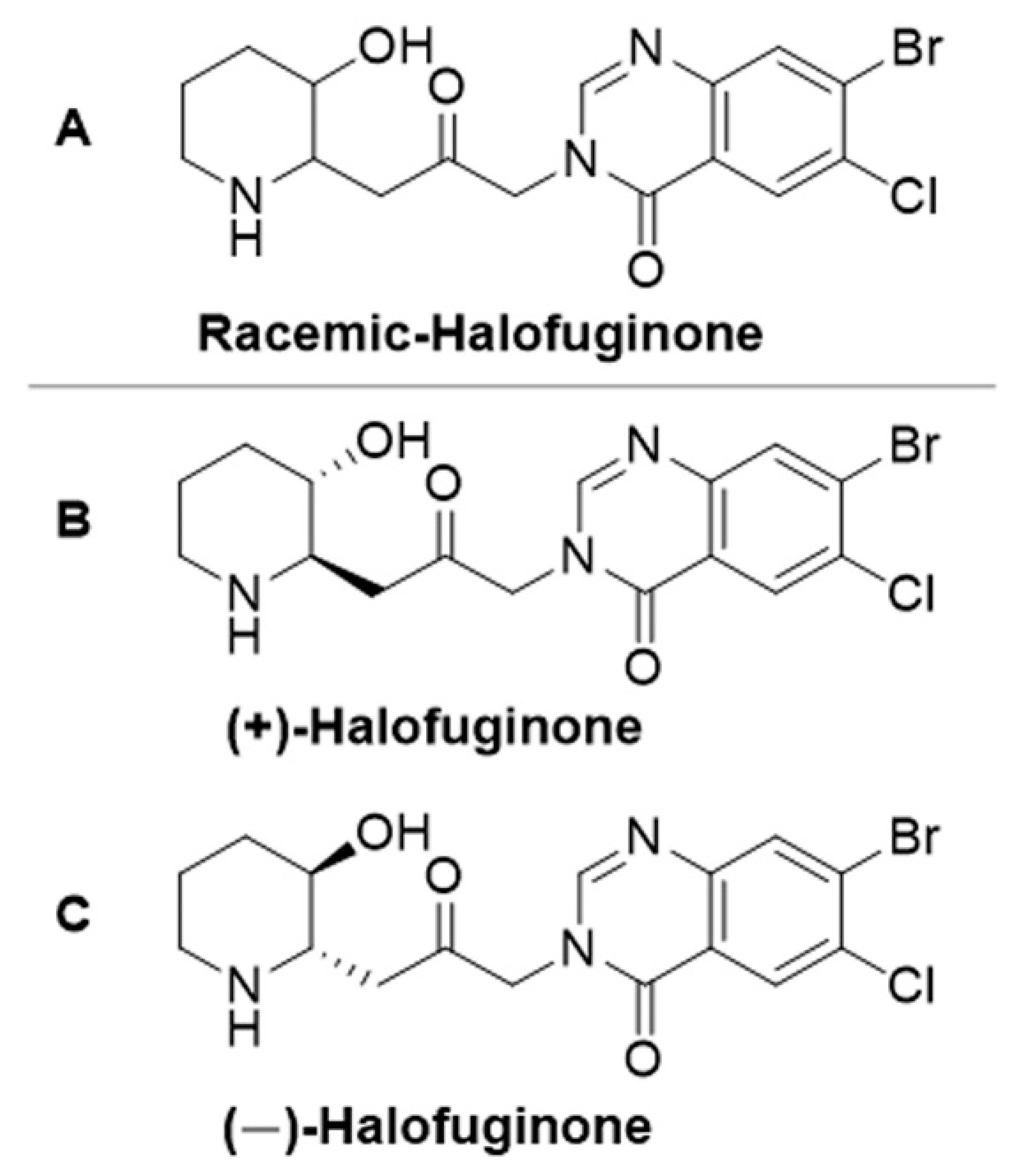{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
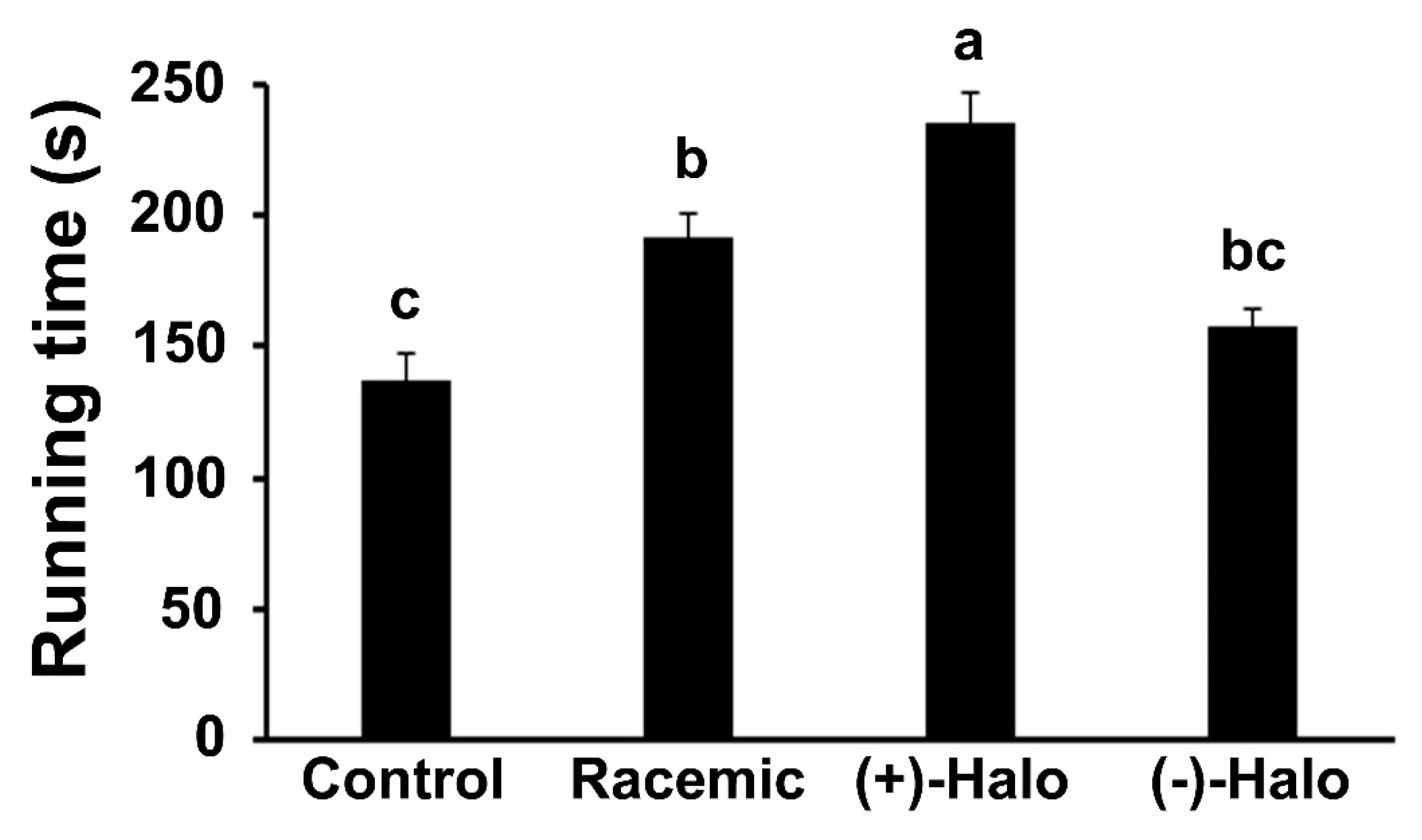
2.1. Motor Coordination
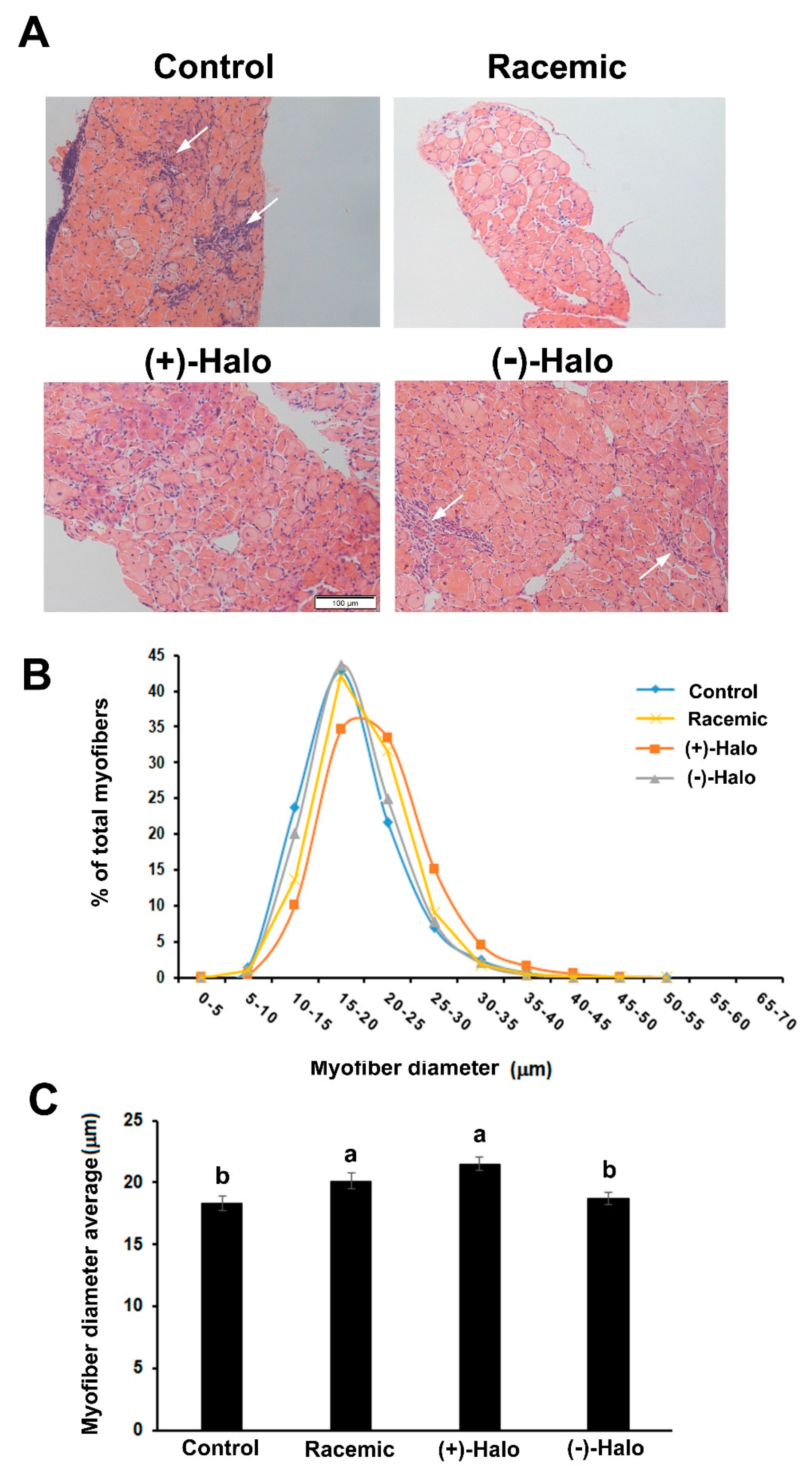
2.2. Sirius Red Staining
2.3. Morphometric Analyses
2.4. Cytotoxicity Assay in C2 Myogenic Cells
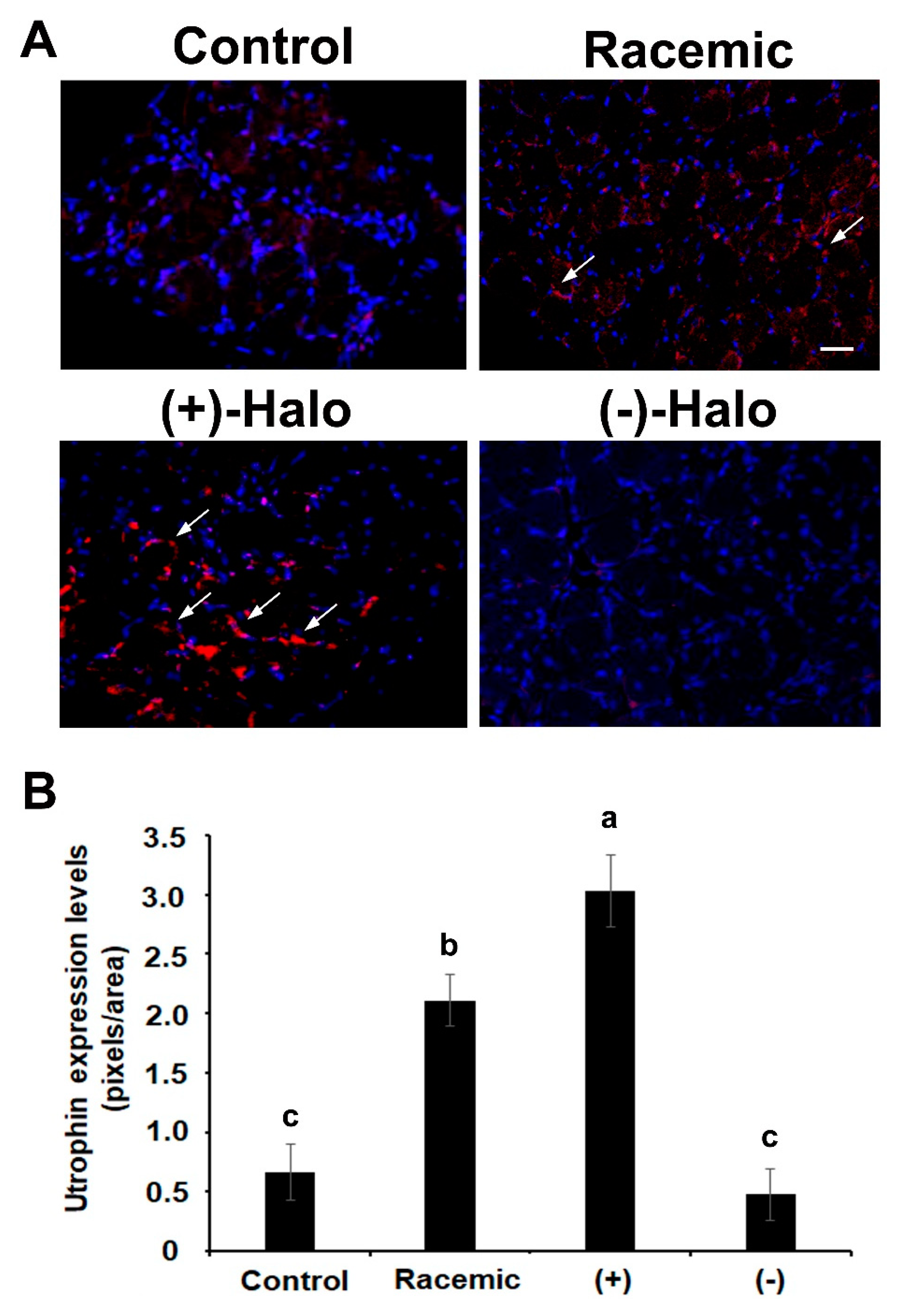
2.5. Utrophin Expression
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animals and Experimental Design
4.3. Motor Coordination Assay
4.4. Histology and Fibrosis Evaluation
4.5. Immunohistochemistry
4.6. Myofiber Diameter Analysis
4.7. Toxicity Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BW | Body weight |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DMD | Duchenne muscular dystrophy |
| H&E | Hematoxylin and eosin |
| ip | Intraperitoneally |
| LDH | Lactate dehydrogenase |
References
- Hantai, D.; Labat-Robert, J.; Grimaud, J.A.; Fardeau, M. Fibronectin, laminin, type I, III and IV collagens in Duchenne’s muscular dystrophy, congenital muscular dystrophies and congenital myopathies: An immunocytochemical study. Connect. Tissue Res. 1985, 13, 273–281. [Google Scholar] [CrossRef]
- Spector, I.; Zilberstein, Y.; Lavy, A.; Genin, O.; Barzilai-Tutsch, H.; Bodanovsky, A.; Halevy, O.; Pines, M. The involvement of Collagen triple helix repeat containing 1 in muscular dystrophies. Am. J. Pathol. 2013, 182, 905–916. [Google Scholar] [CrossRef]
- Lessa, T.B.; de Abreu, D.K.; Bertassoli, B.M.; Ambrósio, C.E. Diaphragm: A vital respiratory muscle in mammals. Ann. Anat. 2016, 205, 122–127. [Google Scholar] [CrossRef]
- Pines, M.; Spector, I. Halofuginone–the multifaceted molecule. Molecules 2015, 20, 573–594. [Google Scholar] [CrossRef] [PubMed]
- Pearce, M.; Blake, D.J.; Tinsley, J.M.; Byth, B.C.; Campbell, L.; Monaco, A.P.; Davies, K.E. The utrophin and dystrophin genes share similarities in genomic structure. Hum. Mol. Genet. 1993, 2, 1765–1772. [Google Scholar] [CrossRef] [PubMed]
- Grady, R.M.; Teng, H.; Nichol, M.C.; Cunningham, J.C.; Wilkinson, R.S.; Sanes, J.R. Skeletal and cardiac myopathies in mice lacking utrophin and dystrophin: A model for Duchenne muscular dystrophy. Cell 1997, 90, 729–738. [Google Scholar] [CrossRef] [Green Version]
- Moorwood, C.; Khurana, T.S. Duchenne muscular dystrophy drug discovery–the application of utrophin promoter activation screening. Exp. Opin. Drug Discov. 2013, 8, 569–581. [Google Scholar] [CrossRef]
- Levi, O.; Genin, O.; Angelini, C.; Fanin, M.; Halevy, O.; Pines, M. Inhibition of muscle fibrosis results in increases in both utrophin levels and the number of revertant myofibers in Duchenne muscular dystrophy. Oncotarget 2015, 6, 23249–23260. [Google Scholar] [CrossRef] [PubMed]
- Wellner, G.; Mordecai, S.; Evans, P.; Genin, O.; Pines, M.; Halevy, O. Hydroxy group requirement for halofuginone-dependent inhibition of muscle fibrosis and improvement of histopathology in the mdx mouse model for Duchenne muscular dystrophy. Histol. Histopathol. J. 2019, 34, 791–801. [Google Scholar]
- Bylo, M.; Farewell, R.; Coppenrath, V.A.; Yogaratnam, D. A Review of deflazacort for patients with Duchenne muscular dystrophy. Ann. Pharmacother. 2020, 54, 788–794. [Google Scholar] [CrossRef]
- Matthews, E.; Brassington, R.; Kuntzer, T.; Jichi, F.; Manzur, A.Y. Corticosteroids for the treatment of Duchenne muscular dystrophy. Cochrane Database Syst. Rev. 2016, 5, CD003725. [Google Scholar] [CrossRef] [Green Version]
- Mendell, J.R.; Sahenk, Z.; Lehman, K.; Nease, C.; Lowes, L.P.; Miller, N.F.; Iammarino, M.A.; Alfano, L.N.; Nicholl, A.; Al-Zaidy, S.; et al. Assessment of systemic delivery of rAAVrh74.MHCK7.micro-dystrophin in children with Duchenne muscular dystrophy: A nonrandomized controlled trial. JAMA Neurol. 2020, 77, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, O.; Yokota, T. Restoring protein expression in neuromuscular conditions: A review assessing the current state of exon skipping/inclusion and gene therapies for Duchenne muscular dystrophy and spinal muscular atrophy. BioDrugs 2021, 7. [Google Scholar] [CrossRef]
- Morine, K.J.; Bish, L.T.; Pendrak, K.; Sleeper, M.M.; Barton, E.R.; Sweeney, H.L. Systemic myostatin inhibition via liver-targeted gene transfer in normal and dystrophic mice. PLoS ONE 2010, 5, e9176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crone, M.; Mah, J.K. Current and emerging therapies for Duchenne muscular dystrophy. Curr. Treat. Options Neurol. 2018, 20, 31. [Google Scholar] [CrossRef] [PubMed]
- Koepfli, J.B.; Mead, J.F.; Brockman, J.A., Jr. An alkaloid with high antimalarial activity from Dichroa febrifuga. J. Am. Chem. Soc. 1947, 69, 1837–1839. [Google Scholar] [CrossRef]
- McLaughlin, N.P.; Evans, P.; Pines, M. The chemistry and biology of febrifugine and halofuginone. Bioorg. Med. Chem. 2014, 22, 1993–2004. [Google Scholar] [CrossRef]
- Pines, M. Targeting TGFβ signaling to inhibit fibroblasts activation as a therapy for fibrosis and cancer. Expert Opin. Drug Discov. 2008, 3, 11–20. [Google Scholar] [CrossRef]
- Roffe, S.; Hagai, Y.; Pines, M.; Halevy, O. Halofuginone inhibits Smad3 phosphorylation via the PI3K/Akt and MAPK/ERK pathways in muscle cells: Effect on myotube fusion. Exp. Cell Res. 2010, 316, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Pines, M.; Nagler, A. Halofuginone—A novel anti-fibrotic therapy. Drug Future 1996, 21, 569–599. [Google Scholar]
- Pines, M.; Halevy, O. Halofuginone and muscular dystrophy. Histol. Histopathol. 2011, 26, 135–146. [Google Scholar]
- Turgeman, T.; Hagai, Y.; Huebner, K.; Anderson, J.; Genin, O.; Nagler, A.; Halevy, O.; Pines, M. Prevention of muscle fibrosis and improvement in muscle performance in the mdx mouse by halofuginone. Neuromuscul. Disord. 2008, 18, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Nevo, Y.; Halevy, O.; Genin, O.; Turgeman, T.; Harel, M.; Biton, E.; Rief, S.; Pines, M. Inhibition of fibrosis in laminin-α2-deficient congenital muscular dystrophy mice: Effect of halofuginone. Muscle Nerve 2010, 42, 218–229. [Google Scholar] [CrossRef]
- Halevy, O.; Genin, O.; Barzilai-Tutsch, H.; Pima, Y.; Levi, O.; Moshe, I.; Pines, M. Inhibition of muscle fibrosis and improvement of muscle histopathology in dysferlin knock-out mice treated with halofuginone. Histol. Histopathol. 2013, 28, 211–226. [Google Scholar] [PubMed]
- Barzilai-Tutsch, H.; Genin, O.; Pines, P.; Halevy, O. Early pathological signs in young dysf-/-mice are improved by halofuginone. Neuromuscul. Disord. 2020, 30, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Pines, M.; Levi, O.; Genin, O.; Lavy, A.; Angelini, C.; Allamand, V.; Halevy, O. Elevated expression of moesin in muscular dystrophies. Am. J. Pathol. 2017, 187, 654–664. [Google Scholar] [CrossRef] [Green Version]
- Sundrud, M.S.; Koralov, S.B.; Feuerer, M.; Calado, D.P.; Kozhaya, A.E.; Rhule-Smith, A.; Lefebvre, R.E.; Unutmaz, D.; Mazitschek, R.; Waldner, H.; et al. Halofuginone inhibits TH17 cell differentiation by activating the amino acid starvation response. Science 2009, 324, 1334–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, T.L.; Zocco, D.; Sundrud, M.S.; Hendrick, M.; Edenius, M.; Yum, J.; Kim, Y.J.; Lee, H.K.; Cortese, J.F.; Wirth, D.F.; et al. Halofuginone and other febrifugine derivatives inhibit prolyl-tRNA synthase. Nat. Chem. Biol. 2012, 12, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Sun, L.; Yang, X.-L.; Schimmel, P. ATP-directed capture of bioactive herbal-based medicine on human tRNA synthetase. Nature 2013, 494, 121–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidan, R.K.; Smullen, S.; Evans, P. Asymmetric synthesis of (+)- and (–)-deoxyfebrifugine and deoxyhalofuginone. Tetrahedron Lett. 2015, 56, 6433–6435. [Google Scholar] [CrossRef]
- Barzilai-Tutsch, H.; Dewulf, M.; Lamaze, C.; Butler Browne, G.; Pines, M.; Halevy, O. A promotive effect for halofuginone on membrane repair and synaptotagmin-7 levels in muscle cells of dysferlin-null mice. Hum. Mol. Gent. 2018, 16, 2817–2829. [Google Scholar] [CrossRef] [PubMed]
- Peregrine, A.S. Chemotherapy and delivery systems: Haemoparasites. Vet. Parasitol. 1994, 54, 223–248. [Google Scholar] [CrossRef]
- Van de Casteele, M.; Roskams, T.; Van der Elst, I.; Van Pelt, J.F.; Fevery, J.; Nevens, F. Halofuginone can worsen liver fibrosis in bile duct obstructed rats. Liver Int. 2004, 24, 502–509. [Google Scholar] [CrossRef] [PubMed]
- de Jonge, M.J.; Dumez, H.; Verweij, J.; Yarkoni, S.; Snyder, D.; Lacombe, D.; Marréaud, S.; Yamaguchi, T.; Punt, C.J.; van Oosterom, A. Phase I and pharmacokinetic study of halofuginone, an oral quinazolinone derivative in patients with advanced solid tumours. Eur. J. Cancer 2006, 42, 1768–1774. [Google Scholar] [CrossRef] [PubMed]
- Holland, R.E. Some infectious causes of diarrhea in young farm animals. Clin. Microbiol. Rev. 1990, 3, 345–375. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.R.; Heckeroth, A.R.; Najdrowski, M.; Daugschies, A.; Schollmeyer, D.; Miculka, C. (2R,3S)-(+)- and (2S,3R)-(-)-Halofuginone lactate: Synthesis, absolute configuration, and activity against Cryptosporidium parvum. Bioorganic Med. Chem. Lett. 2007, 17, 4140–4143. [Google Scholar] [CrossRef] [PubMed]
- Smullen, S.; Evans, P. An asymmetric synthesis of febrifugine, halofuginone and their hemiketal isomers. Tetrahedron 2017, 73, 5493–5499. [Google Scholar] [CrossRef]
- Pines, M.; Snyder, D.; Yarkoni, S.; Nagler, A. Halofuginone to treat fibrosis in chronic graft versus host disease and scleroderma. Biol. Blood Marrow Transplant. 2003, 9, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Yaffe, D.; Saxel, O. Serial passaging and differentiation of myogenic cells isolated from dystrophic mouse muscle. Nature 1977, 270, 725–727. [Google Scholar] [CrossRef]
- Bodanovsky, A.; Guttman, N.; Barzilai-Tutcsh, H.; Levy, O.; Pines, M.; Halevy, O. Halofuginone improves cell survival in muscular dystrophies. Biochem. Biophys. Acta Mol. Cell Res. 2014, 1843, 1339–1347. [Google Scholar] [CrossRef] [Green Version]
- Barzilai-Tutsch, H.; Bodanovsky, A.; Maimon, H.; Pines, M.; Halevy, O. Halofuginone promotes satellite cell activation and survival in muscular dystrophies. Biochem. Biophys. Acta. Mol. Basis Dis. 2016, 1862, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rybalka, E.; Timpani, C.A.; Cooke, M.B.; Williams, A.D.; Hayes, A. Defects in mitochondrial ATP synthesis in dystrophin-deficient mdx skeletal muscles may be caused by complex I insufficiency. PLoS ONE 2014, 9, e115763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubinin, M.V.; Talanov, E.Y.; Tenkov, K.S.; Starinets, V.S.; Mikheeva, I.B.; Sharapov, M.G.; Belosludtsev, K.N. Duchenne muscular dystrophy is associated with the inhibition of calcium uniport in mitochondria and an increased sensitivity of the organelles to the calcium-induced permeability transition. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165674. [Google Scholar] [CrossRef]
- Hughes, M.C.; Ramos, S.V.; Turnbull, P.C.; Rebalka, I.A.; Cao, A.; Monaco, C.M.F.; Varah, N.E.; Edgett, B.A.; Huber, J.S.; Tadi, P.; et al. Early myopathy in Duchenne muscular dystrophy is associated with elevated mitochondrial H2O2 emission during impaired oxidative phosphorylation. J. Cachexia Sarcopenia Muscle 2019, 10, 643–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moulin, M.; Ferreiro, A. Muscle redox disturbances and oxidative stress as pathomechanisms and therapeutic targets in early-onset myopathies. Semin. Cell Dev. Biol. 2017, 64, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Terrill, J.R.; Radley-Crabb, H.G.; Iwasaki, T.; Lemckert, F.A.; Arthur, P.G.; Grounds, M.D. Oxidative stress and pathology in muscular dystrophies: Focus on protein thiol oxidation and dysferlinopathies. FEBS J. 2013, 280, 4149–4164. [Google Scholar] [CrossRef]
- Dubowitz, V. Muscle Biopsy: A Practical Approach; Balliere-Tindal: Philadelphia, PA, USA, 1985. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mordechay, S.; Smullen, S.; Evans, P.; Genin, O.; Pines, M.; Halevy, O. Differential Effects of Halofuginone Enantiomers on Muscle Fibrosis and Histopathology in Duchenne Muscular Dystrophy. Int. J. Mol. Sci. 2021, 22, 7063. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137063
Mordechay S, Smullen S, Evans P, Genin O, Pines M, Halevy O. Differential Effects of Halofuginone Enantiomers on Muscle Fibrosis and Histopathology in Duchenne Muscular Dystrophy. International Journal of Molecular Sciences. 2021; 22(13):7063. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137063
Chicago/Turabian StyleMordechay, Sharon, Shaun Smullen, Paul Evans, Olga Genin, Mark Pines, and Orna Halevy. 2021. "Differential Effects of Halofuginone Enantiomers on Muscle Fibrosis and Histopathology in Duchenne Muscular Dystrophy" International Journal of Molecular Sciences 22, no. 13: 7063. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137063