
Plasticity in Intrinsic Excitability of Hypothalamic Magnocellular Neurosecretory Neurons in Late-Pregnant and Lactating Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
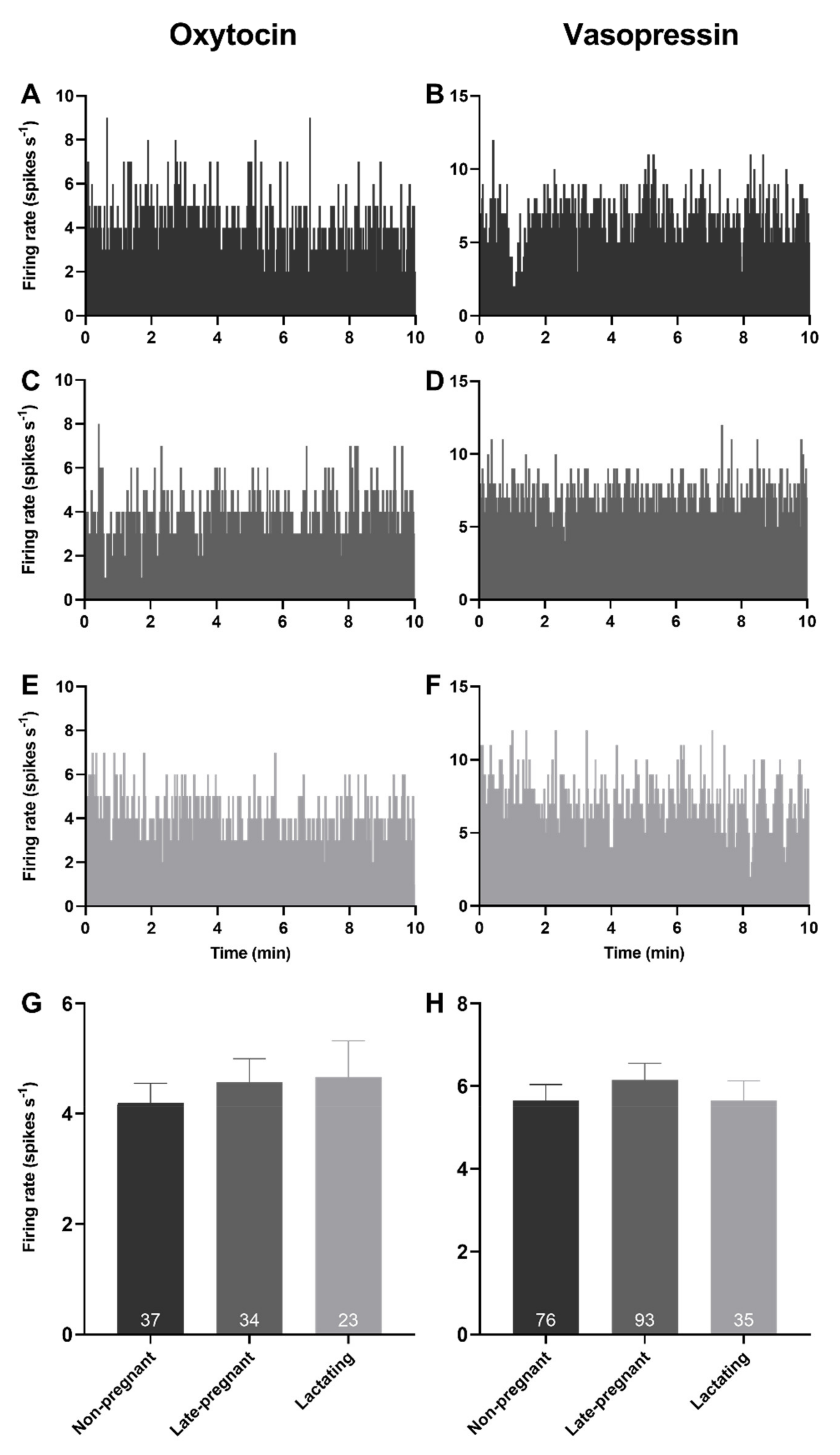
2.1. Basal Firing Rate of Oxytocin and Vasopressin Neurons in Non-Pregnant, Late-Pregnant, and Lactating Rats
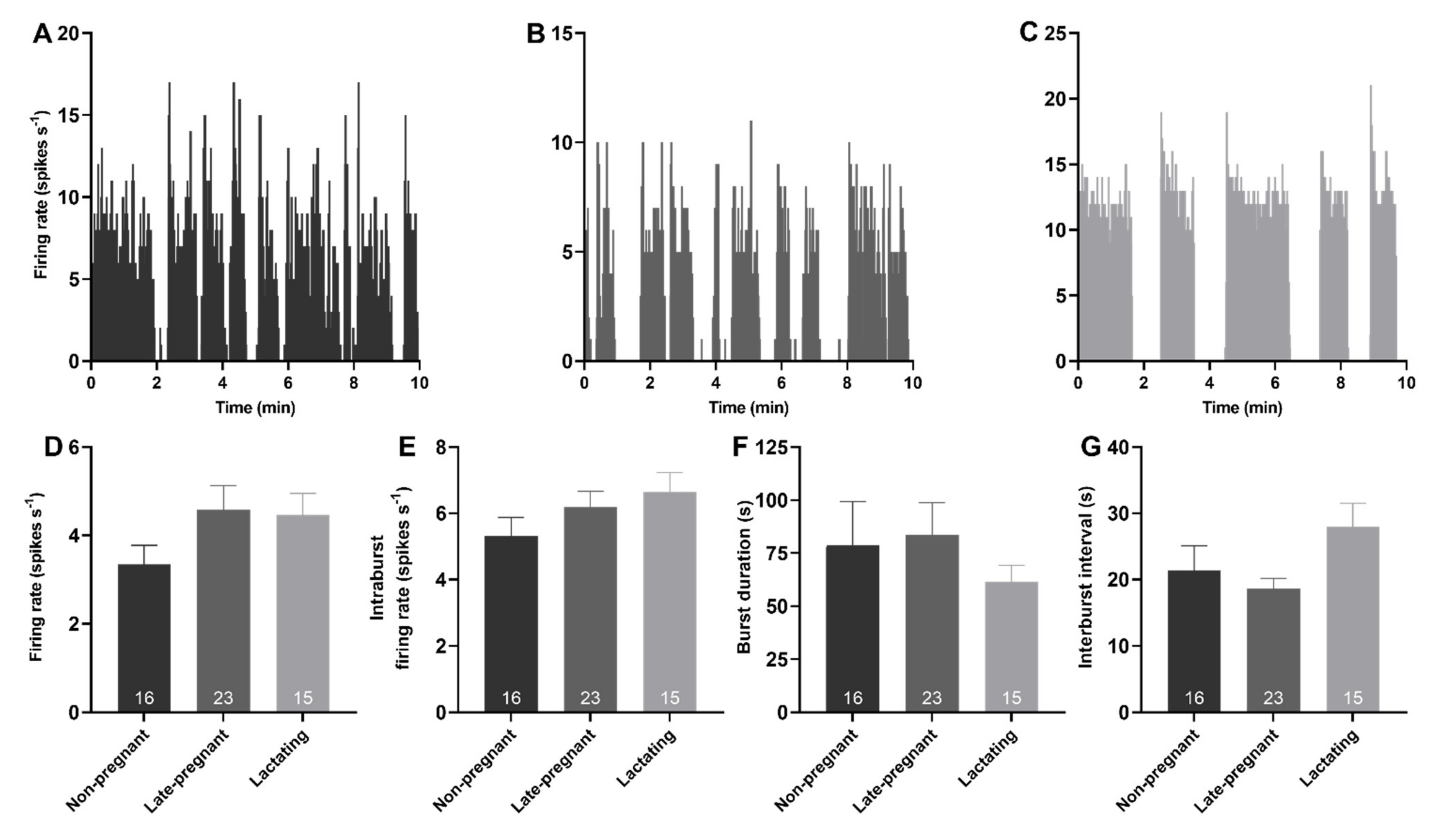
2.2. Phasic Activity of Vasopressin Neurons in Non-Pregnant, Late-Pregnant, and Lactating Rats
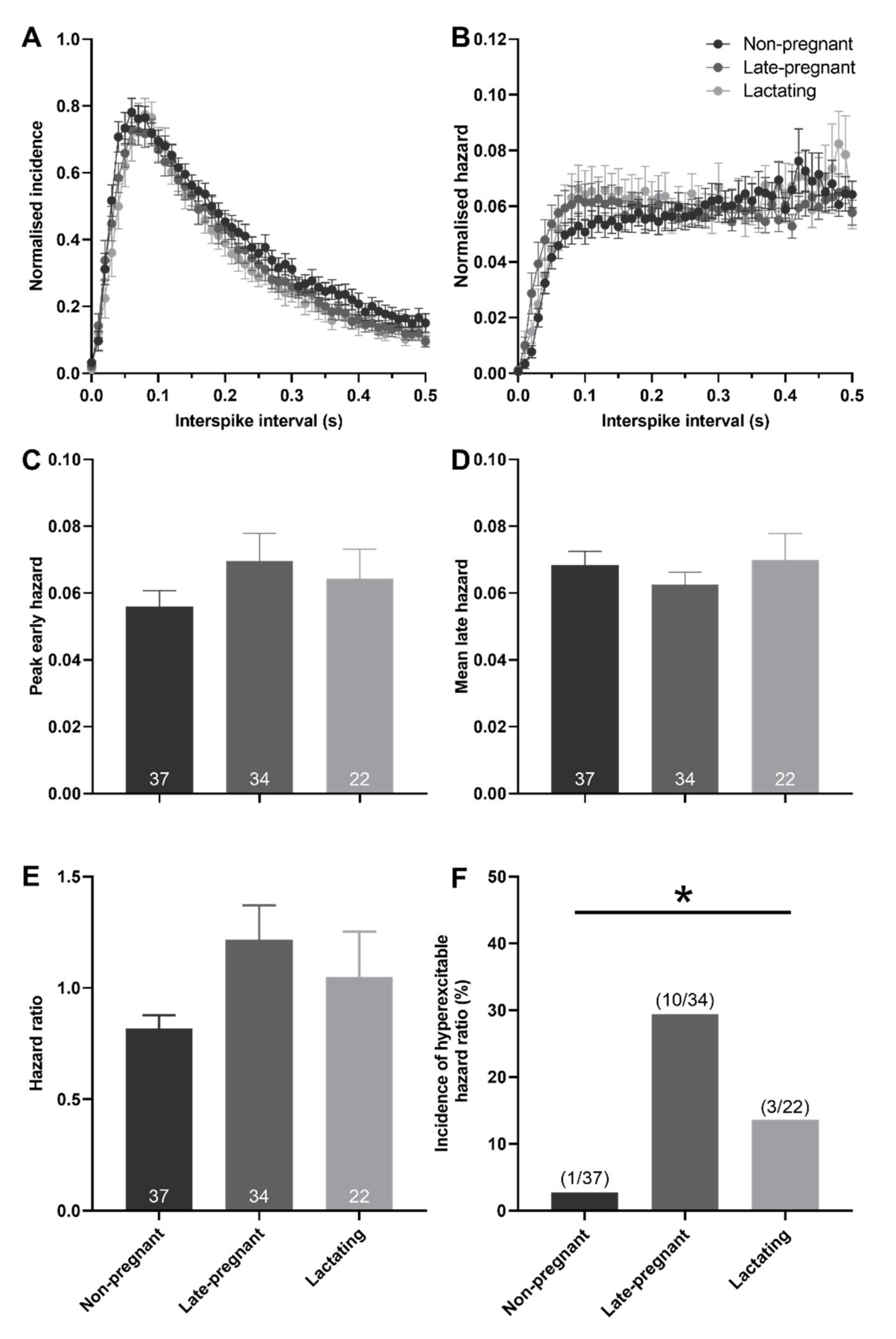
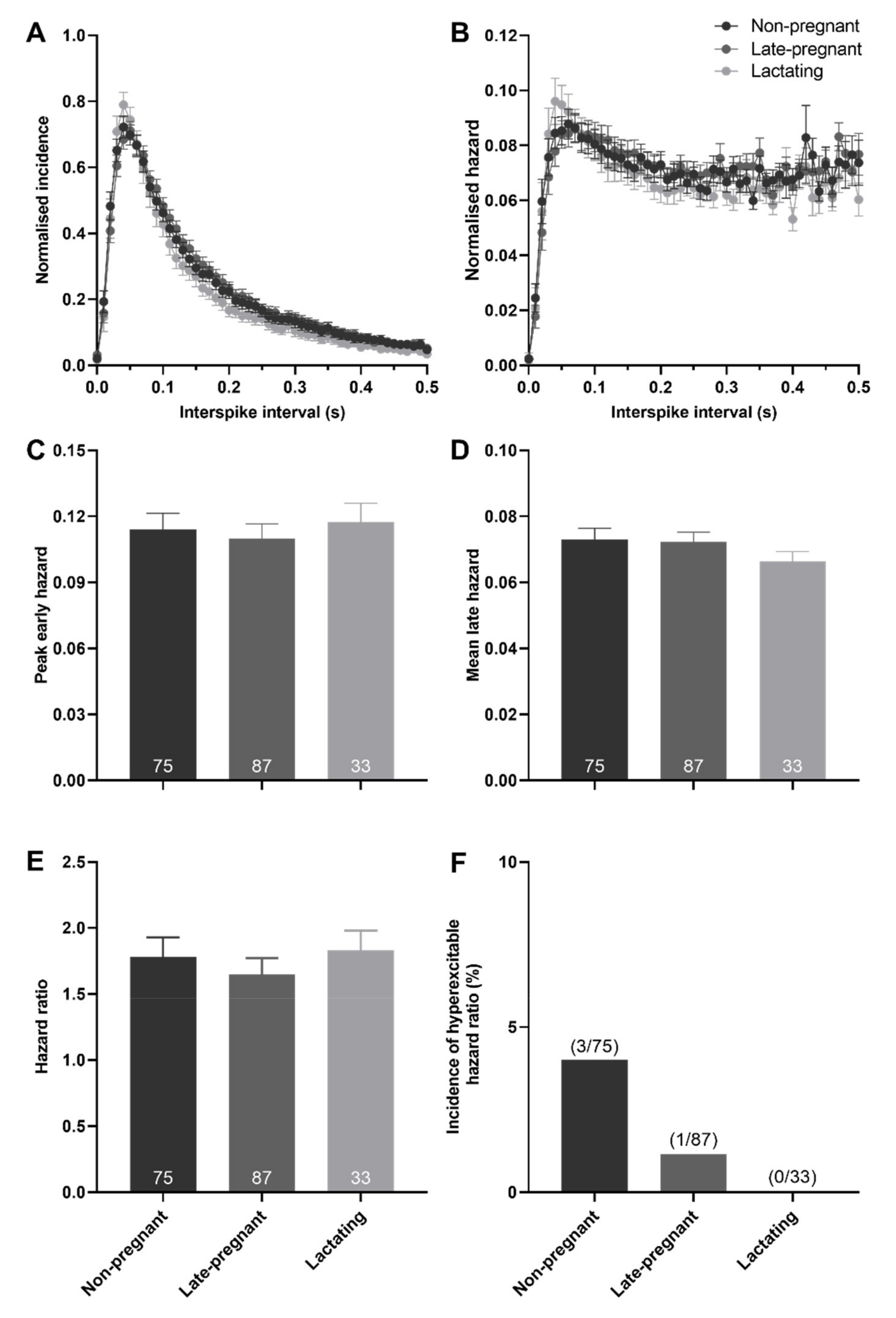
2.3. Post-Spike Excitability in Oxytocin and Vasopressin Neurons in Non-Pregnant, Late-Pregnant, and Lactating Rats
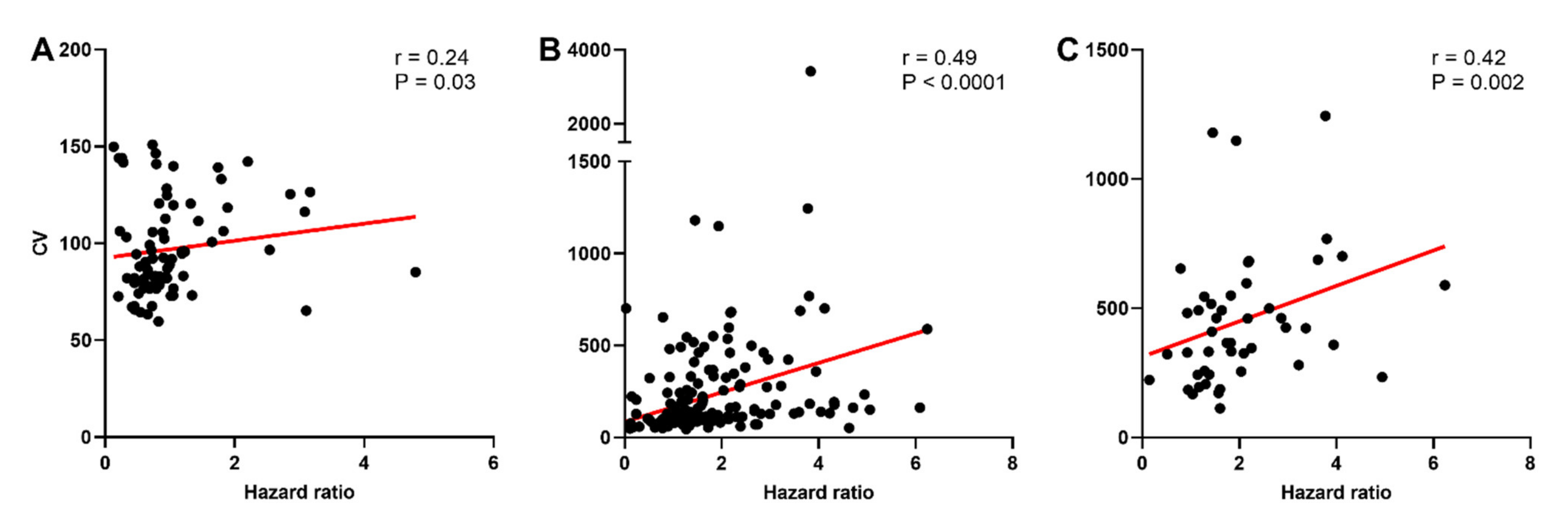
2.4. Variability of Spike Firing in Oxytocin and Vasopressin Neurons in Non-Pregnant, Late-Pregnant, and Lactating Rats
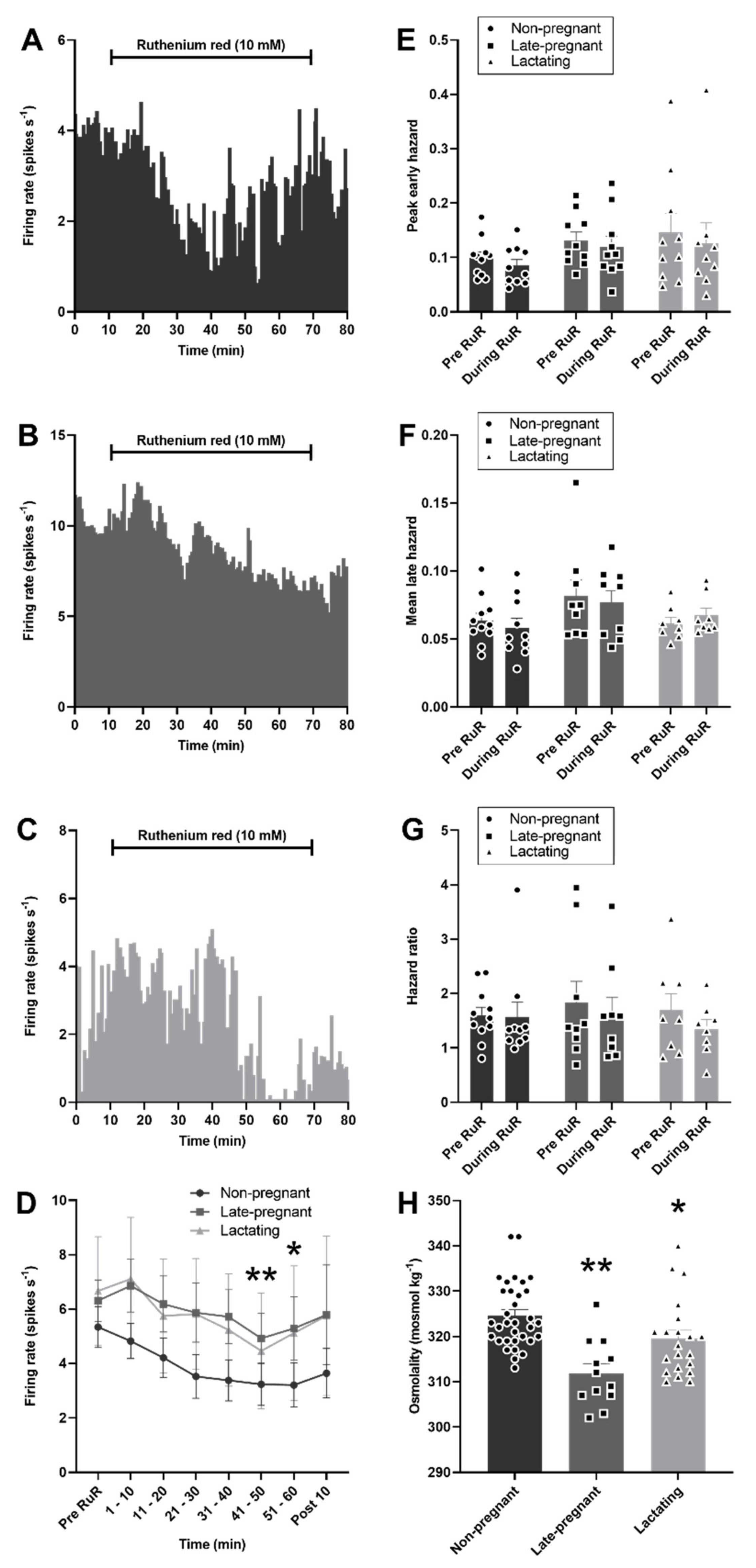
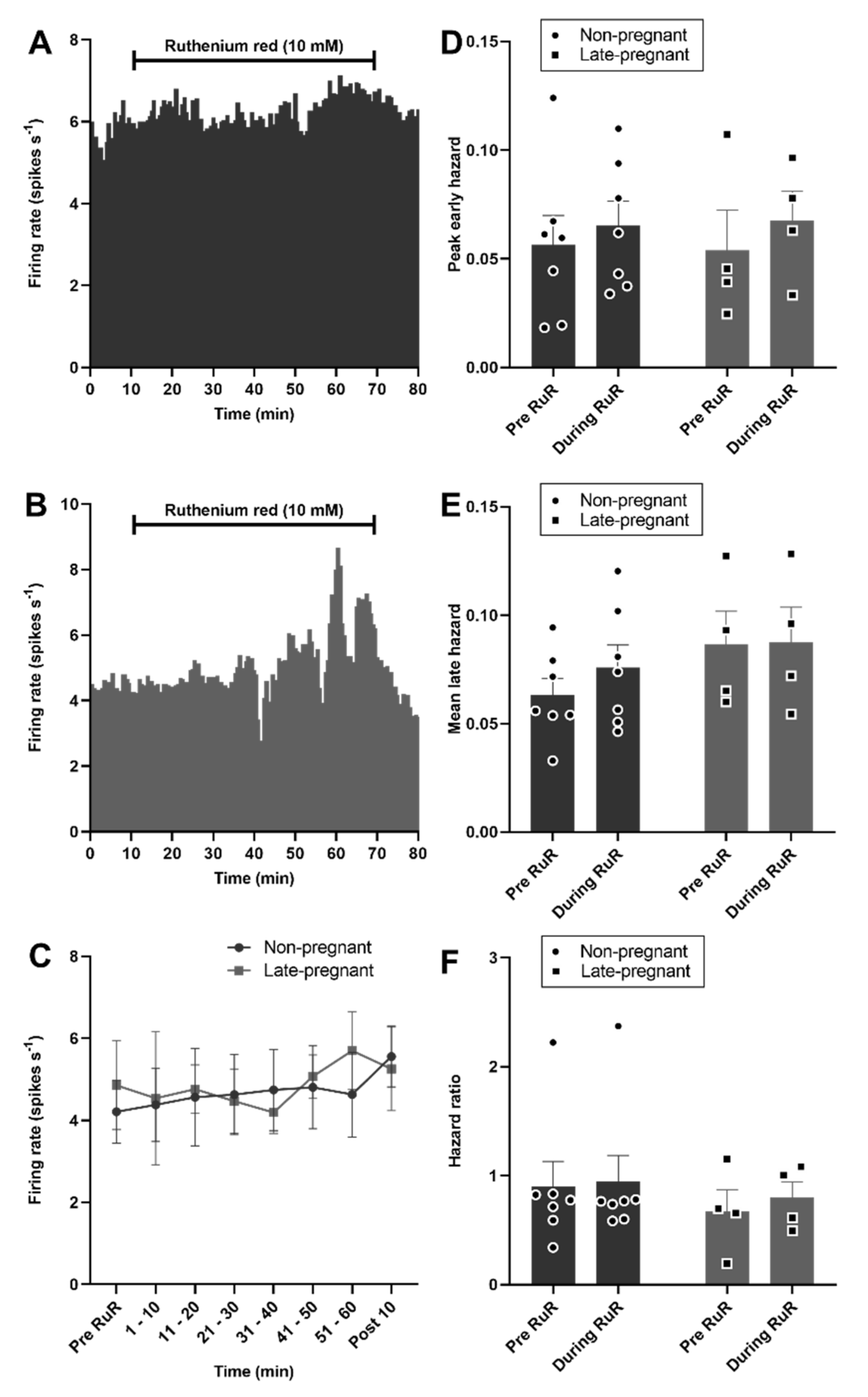
2.5. Ruthenium Red Effects on Oxytocin and Vasopressin Neuron Activity in Non-Pregnant, Late-Pregnant, and Lactating Rats
2.6. ΔN-TRPV1 mRNA and Vasopressin mRNA Expression in the Supraoptic Nucleus of Non-Pregnant and Late-Pregnant Rats
3. Discussion
3.1. Increased Post-Spike Hyperexcitability in Oxytocin Neurons in Late Pregnancy and Lactation
3.2. Increased Excitability in Vasopressin Neurons in Late Pregnancy and Lactation
4. Methods
4.1. Animals
4.2. In Vivo Electrophysiology
4.3. Hazard Function
4.4. Post-Spike Hyperexcitability
4.5. Coefficient of Variation
4.6. RNAscope
4.7. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, C.H. Magnocellular neurons and posterior pituitary function. Compr. Physiol. 2016, 6, 1701–1741. [Google Scholar]
- Brunton, P.J.; Arunachalam, S.; Russel, J.A. Control of neurohypophysial hormone secretion, blood osmolality and volume in pregnancy. J. Physiol. Pharmacol. 2008, 59 (Suppl. 8), 27–45. [Google Scholar]
- Verbalis, J.G. Disorders of body water homeostasis. Best Pract. Res. Clin. Endocrinol. Metab. 2003, 17, 471–503. [Google Scholar] [CrossRef]
- Davison, J.M.; Shiells, E.A.; Philips, P.R.; Lindheimer, M.D. Serial evaluation of vasopressin release and thirst in human pregnancy. Role of human chorionic gonadotrophin in the osmoregulatory changes of gestation. J. Clin. Investig. 1988, 81, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Durr, J.A.; Stamoutsos, B.; Lindheimer, M.D. Osmoregulation during pregnancy in the rat. Evidence for resetting of the threshold for vasopressin secretion during gestation. J. Clin. Investig. 1981, 68, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Forsling, M.L.; Montgomery, H.; Halpin, D.; Windle, R.J.; Treacher, D.F. Daily patterns of secretion of neurohypophysial hormones in man: Effect of age. Exp. Physiol. 1998, 83, 409–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summerlee, A.J. Extracellular recordings from oxytocin neurones during the expulsive phase of birth in unanaesthetized rats. J. Physiol. 1981, 321, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Summerlee, A.J.; Lincoln, D.W. Electrophysiological recordings from oxytocinergic neurones during suckling in the unanaesthetized lactating rat. J. Endocrinol. 1981, 90, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Wakerley, J.B.; Lincoln, D.W. The milk-ejection reflex of the rat: A 20- to 40-fold acceleration in the firing of paraventricular neurones during oxytocin release. J. Endocrinol. 1973, 57, 477–493. [Google Scholar] [CrossRef]
- Voisin, D.L.; Bourque, C.W. Integration of sodium and osmosensory signals in vasopressin neurons. Trends Neurosci. 2002, 25, 199–205. [Google Scholar] [CrossRef]
- Bourque, C.W. Central mechanisms of osmosensation and systemic osmoregulation. Nat. Rev. Neurosci. 2008, 9, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Naeini, R.S.; Witty, M.F.; Seguela, P.; Bourque, C.W. An N-terminal variant of Trpv1 channel is required for osmosensory transduction. Nat. Neurosci. 2006, 9, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Oliet, S.H.; Bourque, C.W. Mechanosensitive channels transduce osmosensitivity in supraoptic neurons. Nature 1993, 364, 341–343. [Google Scholar] [CrossRef]
- Zhang, Z.; Kindrat, A.N.; Sharif-Naeini, R.; Bourque, C.W. Actin filaments mediate mechanical gating during osmosensory transduction in rat supraoptic nucleus neurons. J. Neurosci. 2007, 27, 4008–4013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greffrath, W.; Magerl, W.; Disque-Kaiser, U.; Martin, E.; Reuss, S.; Boehmer, G. Contribution of Ca2+-activated K+ channels to hyperpolarizing after-potentials and discharge pattern in rat supraoptic neurones. J. Neuroendocrinol. 2004, 16, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Greffrath, W.; Martin, E.; Reuss, S.; Boehmer, G. Components of after-hyperpolarization in magnocellular neurones of the rat supraoptic nucleus in vitro. J. Physiol. 1998, 513 Pt 2, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, K.; Bourque, C.W. Activity dependence and functional role of the apamin-sensitive K+ current in rat supraoptic neurones in vitro. J. Physiol. 1996, 494 Pt 2, 389–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teruyama, R.; Armstrong, W.E. Calcium-dependent fast depolarizing afterpotentials in vasopressin neurons in the rat supraoptic nucleus. J. Neurophysiol. 2007, 98, 2612–2621. [Google Scholar] [CrossRef] [Green Version]
- Cobbett, P.; Smithson, K.G.; Hatton, G.I. Immunoreactivity to vasopressin—But not oxytocin-associated neurophysin antiserum in phasic neurons of rat hypothalamic paraventricular nucleus. Brain Res. 1986, 362, 7–16. [Google Scholar] [CrossRef]
- Armstrong, W.E.; Smith, B.N.; Tian, M. Electrophysiological characteristics of immunochemically identified rat oxytocin and vasopressin neurones in vitro. J. Physiol. 1994, 475, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Augustine, R.A.; Ladyman, S.R.; Bouwer, G.T.; Alyousif, Y.; Sapsford, T.J.; Scott, V.; Kokay, I.C.; Grattan, D.R.; Brown, C.H. Prolactin regulation of oxytocin neurone activity in pregnancy and lactation. J. Physiol. 2017, 595, 3591–3605. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, L.E.; Leng, G.; Brown, C.H. Effects of neurotensin on the organisation of activity in supraoptic nucleus cells in virgin and lactating rats. J. Neuroendocrinol. 2004, 16, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.H.; Scott, V.; Ludwig, M.; Leng, G.; Bourque, C.W. Somatodendritic dynorphin release: Orchestrating activity patterns of vasopressin neurons. Biochem. Soc. Trans. 2007, 35 Pt 5, 1236–1242. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.H.; Stern, J.E.; Jackson, K.L.; Bull, P.M.; Leng, G.; Russell, J.A. Morphine withdrawal increases intrinsic excitability of oxytocin neurons in morphine-dependent rats. Eur. J. Neurosci. 2005, 21, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Augustine, R.A.; Seymour, A.J.; Campbell, R.E.; Grattan, D.R.; Brown, C.H. Integrative neuro-humoral regulation of oxytocin neuron activity in pregnancy and lactation. J. Neuroendocrinol. 2018, 30, e12569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, J.E.; Armstrong, W.E. Changes in the electrical properties of supraoptic nucleus oxytocin and vasopressin neurons during lactation. J. Neurosci. 1996, 16, 4861–4871. [Google Scholar] [CrossRef] [Green Version]
- Teruyama, R.; Armstrong, W.E. Changes in the active membrane properties of rat supraoptic neurones during pregnancy and lactation. J. Neuroendocrinol. 2002, 14, 933–944. [Google Scholar] [CrossRef]
- Brown, C.H.; Bourque, C.W. Mechanisms of rhythmogenesis: Insights from hypothalamic vasopressin neurons. Trends Neurosci. 2006, 29, 108–115. [Google Scholar] [CrossRef]
- Sabatier, N.; Brown, C.H.; Ludwig, M.; Leng, G. Phasic spike patterning in rat supraoptic neurones in vivo and in vitro. J. Physiol. 2004, 558, 161–180. [Google Scholar] [CrossRef]
- Royo, J.M.; Brown, C.H.; Leng, G.; MacGregor, D.J. Oxytocin neurones: Intrinsic mechanisms governing the regularity of spiking activity. J. Neuroendocrinol. 2016, 28. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.; Fontanaud, P.; Moos, F.C. The variability of basal action potential firing is positively correlated with bursting in hypothalamic oxytocin neurones. J. Neuroendocrinol. 2000, 12, 506–520. [Google Scholar] [CrossRef]
- Moos, F.; Fontanaud, P.; Mekaouche, M.; Brown, D. Oxytocin neurones are recruited into co-ordinated fluctuations of firing before bursting in the rat. Neuroscience 2004, 125, 391. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.; Moos, F. Onset of bursting in oxytocin cells in suckled rats. J. Physiol. 1997, 503 Pt 3, 625–634. [Google Scholar] [CrossRef]
- Davison, J.M.; Vallotton, M.B.; Lindheimer, M.D. Plasma osmolality and urinary concentration and dilution during and after pregnancy: Evidence that lateral recumbency inhibits maximal urinary concentrating ability. Br. J. Obstet. Gynaecol. 1981, 88, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Weitzman, R.E.; Leake, R.D.; Rubin, R.T.; Fisher, D.A. The effect of nursing on neurohypophyseal hormone and prolactin secretion in human subjects. J. Clin. Endocrinol Metab. 1980, 51, 836–839. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Koizumi, N.; Hirose, H.; Hokao, R.; Takemura, N.; Motoyoshi, S. Changes in plasma arginine vasopressin concentration during lactation in rats. Comp. Med. 2000, 50, 277–280. [Google Scholar] [PubMed]
- Koehler, E.M.; McLemore, G.L.; Tang, W.; Summy-Long, J.Y. Osmoregulation of the magnocellular system during pregnancy and lactation. Am. J. Physiol. 1993, 264 Pt 2, R555–R560. [Google Scholar] [CrossRef]
- Tkachenko, O.; Shchekochikhin, D.; Schrier, R.W. Hormones and hemodynamics in pregnancy. Int. J. Endocrinol. Metab. 2014, 12, e14098. [Google Scholar] [CrossRef] [Green Version]
- Barron, W.M.; Schreiber, J.; Lindheimer, M.D. Effect of ovarian sex steroids on osmoregulation and vasopressin secretion in the rat. Am. J. Physiol. 1986, 250 Pt 1, E352–E361. [Google Scholar] [CrossRef]
- Barron, W.M.; Lindheimer, M.D. Osmoregulation in pseudopregnant and prolactin-treated rats: Comparison with normal gestation. Am. J. Physiol. 1988, 254 Pt 2, R478–R484. [Google Scholar] [CrossRef]
- Zhao, S.; Malmgren, C.H.; Shanks, R.D.; Sherwood, O.D. Monoclonal antibodies specific for rat relaxin. VIII. Passive immunization with monoclonal antibodies throughout the second half of pregnancy reduces water consumption in rats. Endocrinology 1995, 136, 1892–1897. [Google Scholar] [CrossRef]
- Zhao, L.; Roche, P.J.; Gunnersen, J.M.; Hammond, V.E.; Tregear, G.W.; Wintour, E.M.; Beck, F. Mice without a functional relaxin gene are unable to deliver milk to their pups. Endocrinology 1999, 140, 445–453. [Google Scholar] [CrossRef]
- Sunn, N.; Egli, M.; Burazin, T.C.; Burns, P.; Colvill, L.; Davern, P.; Denton, D.A.; Oldfield, B.J.; Weisinger, R.S.; Rauch, M.; et al. Circulating relaxin acts on subfornical organ neurons to stimulate water drinking in the rat. Proc. Natl. Acad. Sci. USA 2002, 99, 1701–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Way, S.A.; Leng, G. Relaxin increases the firing rate of supraoptic neurones and increases oxytocin secretion in the rat. J. Endocrinol. 1992, 132, 149. [Google Scholar] [CrossRef]
- Chakfe, Y.; Bourque, C.W. Excitatory peptides and osmotic pressure modulate mechanosensitive cation channels in concert. Nat. Neurosci. 2000, 3, 572–579. [Google Scholar] [CrossRef]
- Prager-Khoutorsky, M.; Khoutorsky, A.; Bourque, C.W. Unique interweaved microtubule scaffold mediates osmosensory transduction via physical interaction with TRPV1. Neuron 2014, 83, 866–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leng, G.; Brown, C.H.; Bull, P.M.; Brown, D.; Scullion, S.; Currie, J.; Blackburn-Munro, R.E.; Feng, J.; Onaka, T.; Verbalis, J.G.; et al. Responses of magnocellular neurons to osmotic stimulation involves coactivation of excitatory and inhibitory input: An experimental and theoretical analysis. J. Neurosci. 2001, 21, 6967–6977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, H.; Vriens, J.; Prenen, J.; Droogmans, G.; Voets, T.; Nilius, B. Anandamide and arachidonic acid use epoxyeicosatrienoic acids to activate TRPV4 channels. Nature 2003, 424, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.R.; MacLennan, D.H. Identification of calmodulin-, Ca2+-, and ruthenium red-binding domains in the Ca2+ release channel (ryanodine receptor) of rabbit skeletal muscle sarcoplasmic reticulum. J. Biol. Chem. 1994, 269, 22698–22704. [Google Scholar] [CrossRef]
- Tang, Y.; Zucker, R.S. Mitochondrial involvement in post-tetanic potentiation of synaptic transmission. Neuron 1997, 18, 483–491. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Hatton, G.I. Ca2+ release from internal stores: Role in generating depolarizing after-potentials in rat supraoptic neurones. J. Physiol. 1997, 498 Pt 2, 339–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, M.; Leng, G. Autoinhibition of supraoptic nucleus vasopressin neurons in vivo: A combined retrodialysis/electrophysiological study in rats. Eur. J. Neurosci. 1997, 9, 2532–2540. [Google Scholar] [CrossRef]
- Lindheimer, M.D.; Barron, W.M.; Davison, J.M. Osmoregulation of thirst and vasopressin release in pregnancy. Am. J. Physiol. 1989, 257 Pt 2, F159–F169. [Google Scholar]
- Nedungadi, T.P.; Carreno, F.R.; Walch, J.D.; Bathina, C.S.; Cunningham, J.T. Region-specific changes in transient receptor potential vanilloid channel expression in the vasopressin magnocellular system in hepatic cirrhosis-induced hyponatraemia. J. Neuroendocrinol. 2012, 24, 642–652. [Google Scholar] [CrossRef]
- Rutter, A.R.; Ma, Q.P.; Leveridge, M.; Bonnert, T.P. Heteromerization and colocalization of TrpV1 and TrpV2 in mammalian cell lines and rat dorsal root ganglia. Neuroreport 2005, 16, 1735–1739. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Yang, F.; Takanishi, C.L.; Zheng, J. Thermosensitive TRPV channel subunits coassemble into heteromeric channels with intermediate conductance and gating properties. J. Gen. Physiol. 2007, 129, 191–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.H.; Han, S.Y.; Moaddab, M.; Scott, V.; Schwenke, D.O. Peptidergic control of oxytocin and vasopressin neurons and its role in reproductive and hypertension-associated plasticity. In Neurophysiology of Neuroendocrine Neuron; Armstrong, W.E., Tasker, J.G., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2014; pp. 65–86. [Google Scholar]
- Horn, T.F.W.; Engelmann, M. In vivo microdialysis for nonapeptides in rat brain—A practical guide. Methods 2001, 23, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Scott, V.; Bishop, V.R.; Leng, G.; Brown, C.H. Dehydration-induced modulation of kappa-opioid inhibition of vasopressin neurone activity. J. Physiol. 2009, 587, 5679–5689. [Google Scholar] [CrossRef]
- Ladyman, S.R.; Augustine, R.A.; Scherf, E.; Phillipps, H.R.; Brown, C.H.; Grattan, D.R. Attenuated hypothalamic responses to alpha-melanocyte stimulating hormone during pregnancy in the rat. J. Physiol. 2016, 594, 1087–1101. [Google Scholar] [CrossRef] [Green Version]
- Scott, V.; Brown, C.H. Kisspeptin activation of supraoptic nucleus neurons in vivo. Endocrinology 2011, 152, 3862–3870. [Google Scholar] [CrossRef] [Green Version]
- Seymour, A.J.; Scott, V.; Augustine, R.A.; Bouwer, G.T.; Campbell, R.E.; Brown, C.H. Development of an excitatory kisspeptin projection to the oxytocin system in late pregnancy. J. Physiol. 2017, 595, 825–838. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perkinson, M.R.; Augustine, R.A.; Bouwer, G.T.; Brown, E.F.; Cheong, I.; Seymour, A.J.; Fronius, M.; Brown, C.H. Plasticity in Intrinsic Excitability of Hypothalamic Magnocellular Neurosecretory Neurons in Late-Pregnant and Lactating Rats. Int. J. Mol. Sci. 2021, 22, 7140. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137140
Perkinson MR, Augustine RA, Bouwer GT, Brown EF, Cheong I, Seymour AJ, Fronius M, Brown CH. Plasticity in Intrinsic Excitability of Hypothalamic Magnocellular Neurosecretory Neurons in Late-Pregnant and Lactating Rats. International Journal of Molecular Sciences. 2021; 22(13):7140. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137140
Chicago/Turabian StylePerkinson, Michael R., Rachael A. Augustine, Gregory T. Bouwer, Emily F. Brown, Isaiah Cheong, Alexander J. Seymour, Martin Fronius, and Colin H. Brown. 2021. "Plasticity in Intrinsic Excitability of Hypothalamic Magnocellular Neurosecretory Neurons in Late-Pregnant and Lactating Rats" International Journal of Molecular Sciences 22, no. 13: 7140. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137140